Introduction
Chemotherapeutic agents that target microtubules are
mainly divided into two categories based on their functions. One
agent, Taxol (paclitaxel), is a microtubule stabilizer which leads
to obstacles in chromosome separation and the cell cycle process by
promoting microtubule polymerization. Taxol is widely prescribed
for the treatment of solid tumors including advanced ovarian and
breast cancer, non-small cell lung cancer (NSCLC) and Kaposi's
sarcoma (1,2). The other is vinblastine (VBL), a
microtubule destabilizer vinca alkaloid, which causes the
dysfunction of cell proliferation by predominantly binding to α/β
tubulin in order to disassemble microtubules. VBL is commonly
applied to treat malignant lymphoma, osteosarcoma, breast, ovarian,
gastric and lung cancer, Hodgkin's disease and choriocarcinoma
(3,4). Thereby, both Taxol and VBL interact
with tubulin while in opposite patterns.
Op18/stathmin is a small molecular biomarker and is
highly expressed in many carcinomas. Op18/stathmin integrates and
relays intracellular and extracellular stimuli to regulate the
equilibrium of microtubule dynamics through phosphorylated
inactivation and dephosphorylated activation at 4 serine sites
including Ser16, Ser25, Ser38 and Ser63, which is vital to the
maintenance of malignant phenotypes in tumors (5–7). Our
previous study confirmed that high expression of Op18/stathmin was
significantly correlated with Taxol resistance in 5 randomly
selected human epithelial-derived carcinoma cell lines, including
nasopharyngeal carcinoma CNE1, gastric MGC, breast cancer MCF-7,
and hepatocellular carcinoma Hep3B-2 cells, as well as in NSCLC
NCI-H1299 cells (8).
The present study aimed to compare the curative
effects of VBL and Taxol in non-small cell lung cancer cells and to
explore the correlation between drug sensitivity and Op18/stathmin
signaling.
Materials and methods
Cell culture
NSCLC NCI-H1299 and A549 cells were cultured in
RPMI-1640 medium supplemented with 10% (v/v) fetal bovine serum
(FBS) and 1% penicillin/streptomycin and maintained at 37̊C in 5%
CO2. Both cell lines were presented as a gift from Dr
Yongguang Tao from the Cancer Research Institute of Central South
University. NCI-H1299 was confirmed to be a Taxol-resistant cell
line with high expression of Op18/stathmin in our previous study
(8).
Antibodies and chemical reagents
Primary antibodies included rabbit polyclonal
anti-stathmin (Calbiochem, La Jolla, CA, USA), rabbit monoclonal
anti-stathmin (phospho-Ser16, -Ser25, -Ser38 and -Ser63) (Abcam,
Cambridge, MA, USA), mouse monoclonal anti-caspase 8, rabbit
monoclonal anti-caspase 9, rabbit monoclonal anti-PP2A C subunit
(Cell Signaling Technology, Danvers, MA, USA), rabbit
anti-phosphoserine (Invitrogen, Carlsbad, CA, USA), part antibodies
including rabbit polyclonal anti-Bcl-2, rabbit polyclonal
anti-NF-κB, mouse monoclonal anti β-actin, mouse monoclonal
anti-caspase 3, mouse monoclonal anti-interleukin-10 (IL-10), which
were purchased from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA,
USA).
Secondary antibodies included rabbit anti-mouse
HRP-conjugated IgG and goat anti-rabbit HRP-conjugated IgG (Santa
Cruz Biotechnology, Inc.).
Taxol (sc-201439) and VBL (sc-491749) were also
obtained from Santa Cruz Biotechnology, Inc., and were dissolved in
dimethyl sulfoxide (DMSO) at appropriate concentrations, and were
stored at −20°C.
Assessments of cell apoptosis
Upon reaching 80% confluency, the cells were treated
with Taxol or VBL (100 nM) for 24 h, and the solvent DMSO was used
as control. Then, the treated cells were harvested and stained with
500 µl specific binding buffer containing 5 µl Annexin V-FITC and 5
µl propidium iodide for 10 min at room temperature in the dark.
Analysis of cell apoptosis was performed by flow cytometry at the
The Second Xiangya Hospital Affiliated with Central South
University.
MTT assays
Cells (5,000/well) in a logarithmic growth phase
were planted into 96-well plates and treated with Taxol or VBL (100
nM). Next, 10 µl of 5 mg/ml 3-(4,5-dimethylthiazol-yl)-2,5-diphenyl
tetrazolium bromide (MTT) was added to each well at time points of
0, 24, 48 and 72 h, and the cells were subsequently incubated at
37̊C for 4 h. After the culture supernatants were removed, 100 µl
DMSO was added for 10 min, and the absorbance was measured at a
490-nm wavelength. Six parallel wells were performed.
Cellular proliferative viability was calculated
using the following formulation: Relative proliferative percentages
(%) = (ODtreatment/ODcontrol) × 100%. The
percentage of the DMSO control was designed as 100% at all time
points.
Colony formation assays
Cells (2,000/well) were cultured in media with 10 nM
Taxol or VBL in 6-well plates for 1–2 weeks. The culture was
suspended when obvious colonies were observed by the naked eye.
Cells were fixed with methanol, and then stained with 0.1% crystal
violet for 30 min. Colonies with >50 cells/colony were counted
using an inverted microscope. The experiment was carried out in
triplicate independently. The relative colony formation efficiency
(%) = (colony number/2,000) × 100%.
Wound healing assays
Cells were grown in 6-well plates to a monolayer.
The cell monolayer was then wounded by creating a scratch using a
200-µl pipette tip. Old medium was replaced with fresh medium
containing 100 nM Taxol or VBL. The status of wound healing was
monitored at time points of 0, 12, 24 and 36 h, and images were
captured for assaying the capability of cell migration in a
2-dimensional plane.
Transwell assays
Capability of cell migration in a 3-dimensional
space was assessed by Transwell assays according to the protocol
described in a previous study (8).
Approximately 4×104 cells were seeded in 200 µl
RPMI-1640 medium with 0.2% FBS in the upper chambers of a
Transwell, while 800 µl media with 10% FBS was placed in the lower
chambers. Twenty-four hours later, the media were removed and the
cells were wiped off from the upper chambers using a cotton swab.
The remaining migrated cells on the back of the bottom membrane in
the upper chamber were fixed with 100% methanol and stained with
0.1% crystal violet. Finally, the Transwell plates were mounted and
observed under an inverted microscope, and the migrated cells were
counted and images were captured.
Western blot analysis
Protein extracts were collected according to our
previous studies (9,10). Briefly, cells were lysed with lysis
buffer (50 mM Tris-HCl pH 8.0, 1 mM EDTA, 2% SDS, 5 mM DTT, 10 mM
PMSF), and then denatured in boiling water for sonication. Protein
concentrations were determined using BCA protein assay reagent
(Pierce, Rockford, IL, USA).
Protein extracts (50 µg) were separated by 10%
SDS-PAGE, electrotransferred to nitrocellulose membrane, blocked
with buffer containing 5% non-fat milk and incubated with primary
antibodies overnight at 4̊C and HRP-conjugated secondary antibodies
for 1 h at room temperature and developed with an enhanced
chemiluminescence detection kit (Thermo Fisher Scientific, Waltham,
MA, USA).
IP-western blot assays
Cells were lysed in immunoprecipitation (IP) lysis
buffer (50 mM Tris-HCl, 150 mM NaCl, 10% NP-40, 1 mM EDTA, 10%
glycerol, 10 mM NaF, 1 mM Na3VO4, 1 mM DTT, 1
mM PMSF and protease inhibitor cocktail tablet), and then the
supernatant was collected for detection of the total level of
phospho-Op18/stathmin.
Magnetic immunobeads (cat#: 10002D; Thermo Fisher
Scientific) were used to bind the anti-stathmin antibody and
pull-down Op18/stathmin for western blot analysis as previously
described (9).
Autocrine IL-10 detection
Equal number of cells were planted in 6-well plates
and cultured. The old medium was replaced with fresh medium without
phenol red indicator in the presence of Taxol or VBL (100 nM) when
cells reached 80% confluency. Twenty-four hours later, the media
were collected and centrifuged at 13,000 rpm for 30 min. Autocrine
IL-10 in the separated supernatant was detected by ELISA using the
human IL-10 ELISA kit (Cusabio, Wuhan, China). All experiments were
carried out in triplicate.
Construction of the RNA interference
plasmid targeting Op18/stathmin
Plasmids pGCsilencer-U6/Neo/GFP-RNAi (RNAi) with the
RNAi gene targeting the coding region of Op18/stathmin at
5′-AGAGAAACTGACCCACAAA-3′ (GenBank no. 53305, sequence 374-393) and
pGC-silencer-U6/Neo/GFP-non (Non) with an inserted non-coding
sequence and pGC-silencer-U6/Neo/GFP (Blank) without any fragments
inserted, were constructed at an early stage and were introduced
into cells by Lipofectamine 2000 (Invitrogen) as described in our
previous studies (8,10).
Statistical analysis
All statistical calculations were performed using
the statistical software program SPSS 17.0. Differences with
p-values <0.05 were considered statistically significant.
Results
NCI-H1299 cells are more sensitive to
VBL than to Taxol
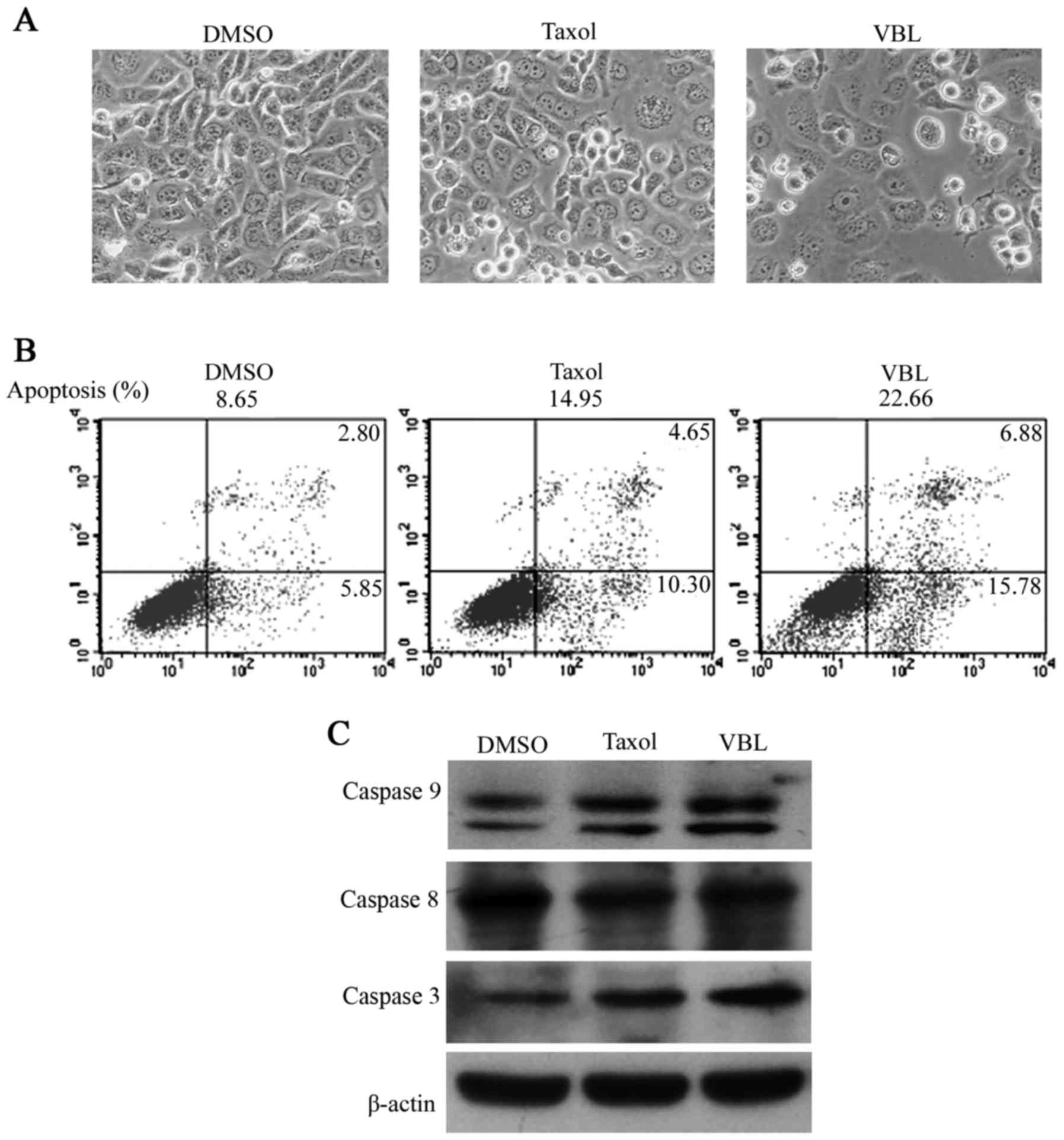
Morphological images showed that cells grew to
nearly complete confluency in the control DMSO, whereas cell
distribution became gradually dispersed with the appearance of a
few translucent floating cells in the Taxol treatment cell group. A
large number of suspended cells emerged and only a minority of
cells adhered to the wall at a low density in the VBL treatment
group (Fig. 1A).
Cell apoptotic ratios were 8.65, 14.95 and 22.66%
following treatment with the solvent (DMSO), Taxol and VBL,
respectively, in the NCI-H1299 cells as detected by FCM. The
apoptotic ratio was significantly higher for VBL treatment compared
with Taxol (Fig. 1B).
Western blot analysis showed that both Taxol and VBL
increased the expression levels of caspase 3 and 9, but to a
greater extent following VBL treatment, while no obvious changes in
caspase 8 were observed in the three treatment groups (Fig. 1C).
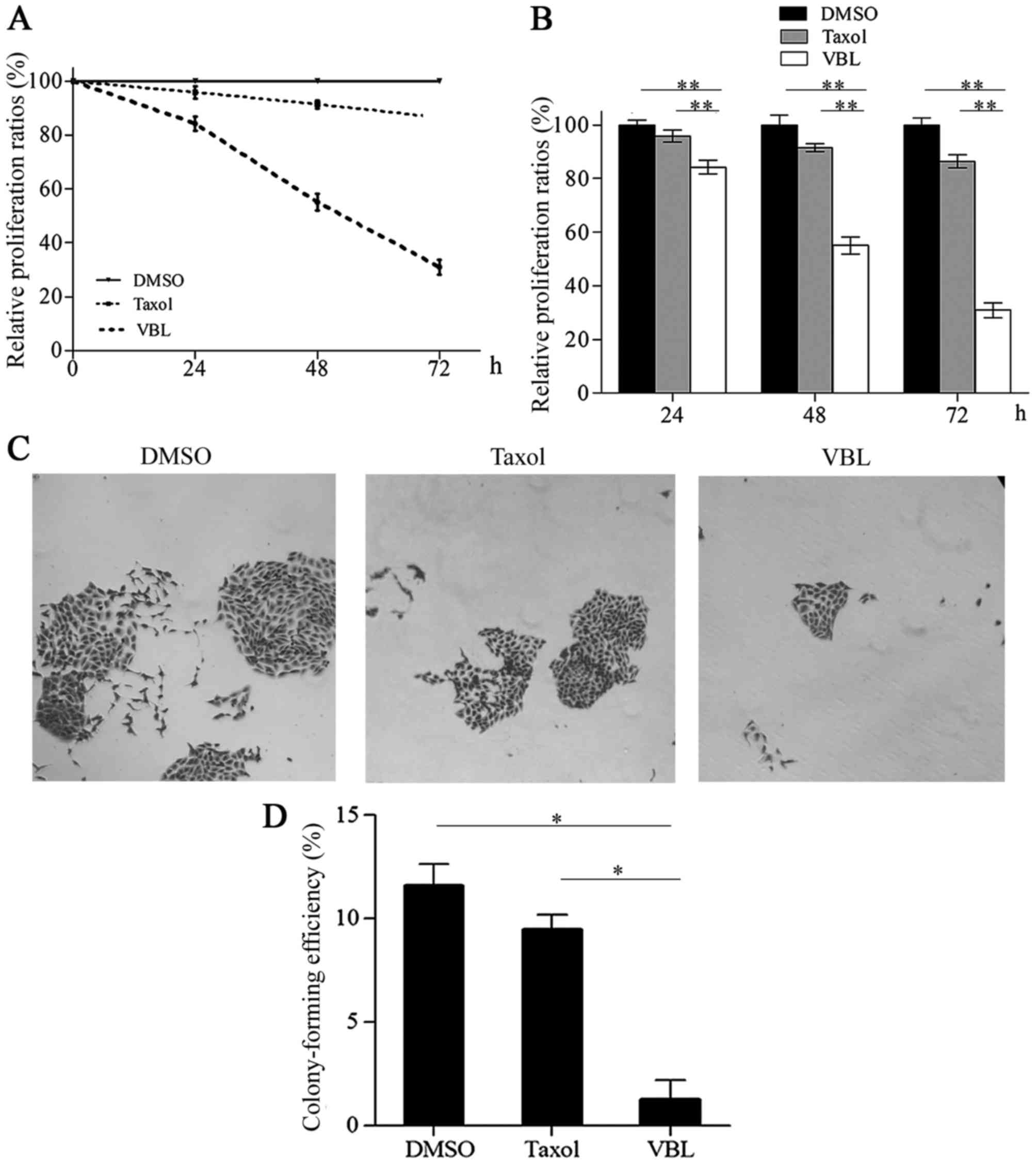
VBL significantly inhibits cell proliferation and
colony formation compared with Taxol. By contrast with DMSO, the
relative cell proliferation percentages were 84.24, 55.11 and
32.00%, respectively, for VBL treatment at time points of 24, 48
and 72 h, while these percentages were 95.86, 91.47 and 86.46%,
respectively, following Taxol treatment. The representative curve
steeply declined with increasing exposure time to VBL, whereas, it
declined slowly in the presence of Taxol (Fig. 2A). Statistical differences existed
in the proliferation percentages between VBL and that of the other
two groups for Taxol and DMSO at 24, 48 and 72 h (p<0.01)
(Fig. 2B).
Colony formation assays revealed that only a few
small colonies appeared in the VBL treatment group, while some
relative larger colonies emerged in the Taxol treatment group.
There were a large number of large colonies which generally merged
into an extensive one with unclear borders in the control DMSO
group (Fig. 2C). The mean colony
formation efficiency was 11.60, 9.47 and 1.07%, respectively, in
the DMSO, Taxol and VBL treatment group. VBL significantly reduced
the percentage of colony formation in comparison with the Taxol and
DMSO treatment groups (p<0.05) (Fig.
2D).
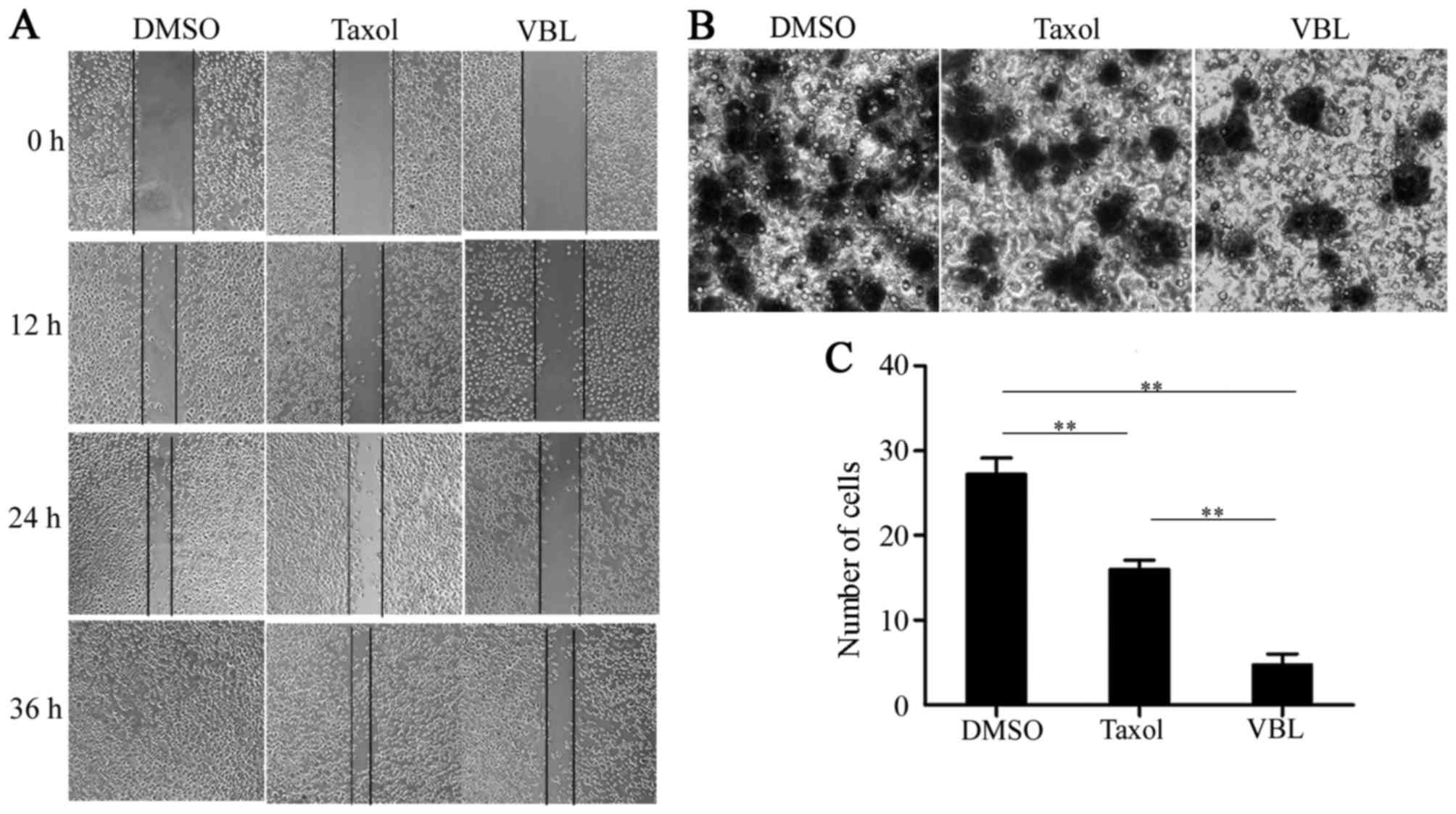
VBL notably suppresses cell migration
in contrast with Taxol
The scratch-wound trails obviously became narrow
with time in the three groups, and were completely recovered at
time point of 36 h in the DMSO treatment group, while the scratched
areas nearly healed for Taxol induction. However, a large area of
blank region still remained in the cells treated with VBL (Fig. 3A).
Transwell assay analysis revealed that the mean
number of invading cells was 27, 16 and 8, respectively, in the
DMSO, Taxol and VBL treatment group. The number of migrated cells
was greatly decreased following treatment with VBL when compared to
the Taxol treatment group, whereas Taxol also inhibited cell
migration in contrast with DMSO (Fig.
3B). Differences in the number of migrating cells were
significant between the VBL group and the other two treatment
groups; VBL was the most notable in inhibiting cell migration of a
3-dimensional space (p<0.01) (Fig.
3C).
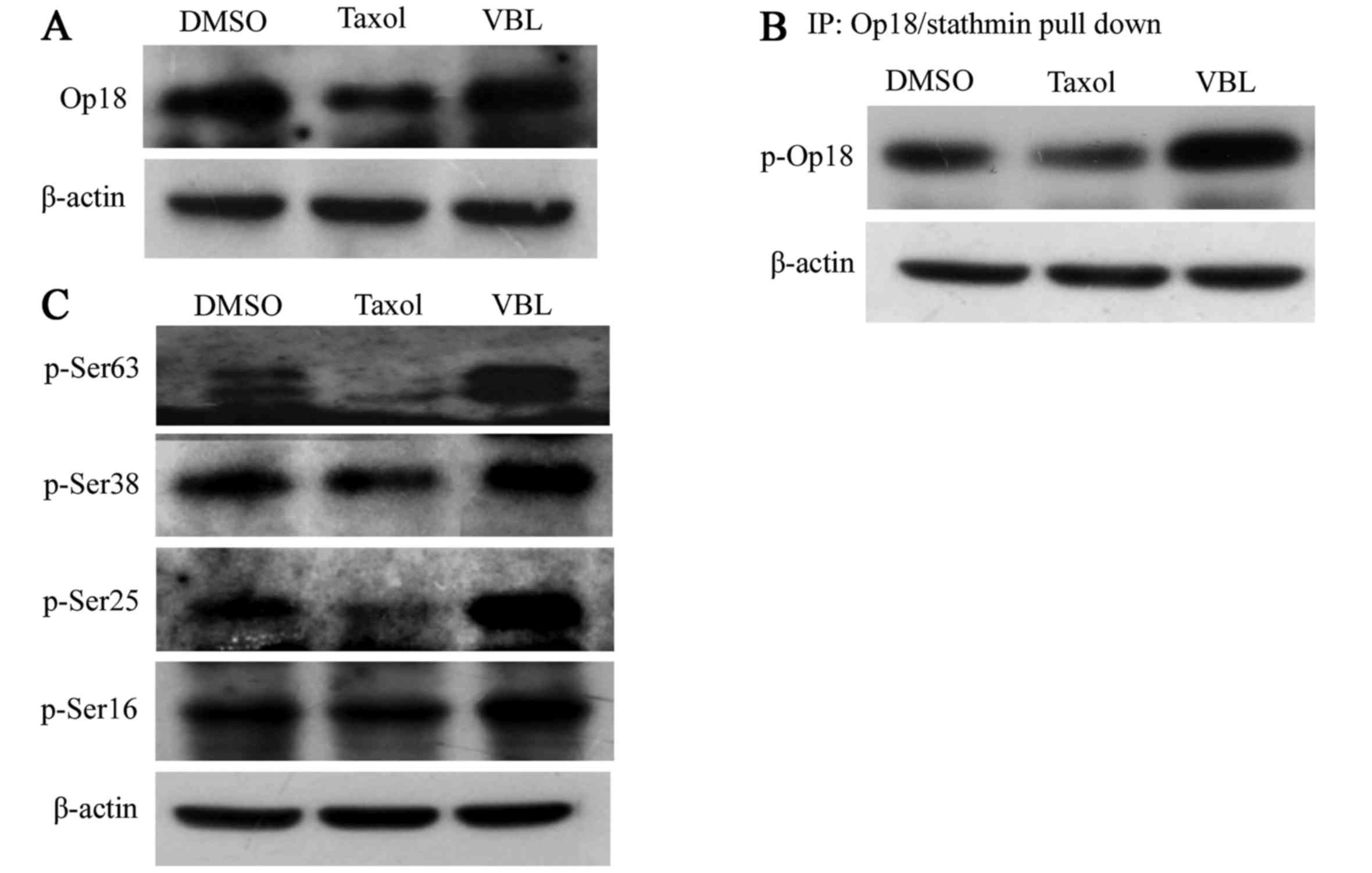
VBL promotes the phosphorylation of
Op18/stathmin, while Taxol decreases the expression and
phosphorylation of Op18/stathmine
Western blot analysis revealed that the expression
of Op18/stathmin was decreased by Taxol, but was not obviously
affected by VBL (Fig. 4A).
IP-western blotting showed that Taxol decreased the phosphorylation
of Op18/stathmin; however, VBL upregulated the total level of
phosphorylated Op18/stathmin (Fig.
4B).
VBL universally increased the level of
phosphorylated Op18/stathmin at all 4 serine sites including Ser63,
Ser38, Ser25 and Ser16, while Taxol mainly reduced phosphorylated
Op18/stathmin at Ser25 and Ser63 sites, particularly,
p-Ser63-Op18/stathmin was almost completely inhibited, yet Taxol
did not influence the phosphorylation of Op18/stathmin at Ser16 and
Ser38 (Fig. 4C).
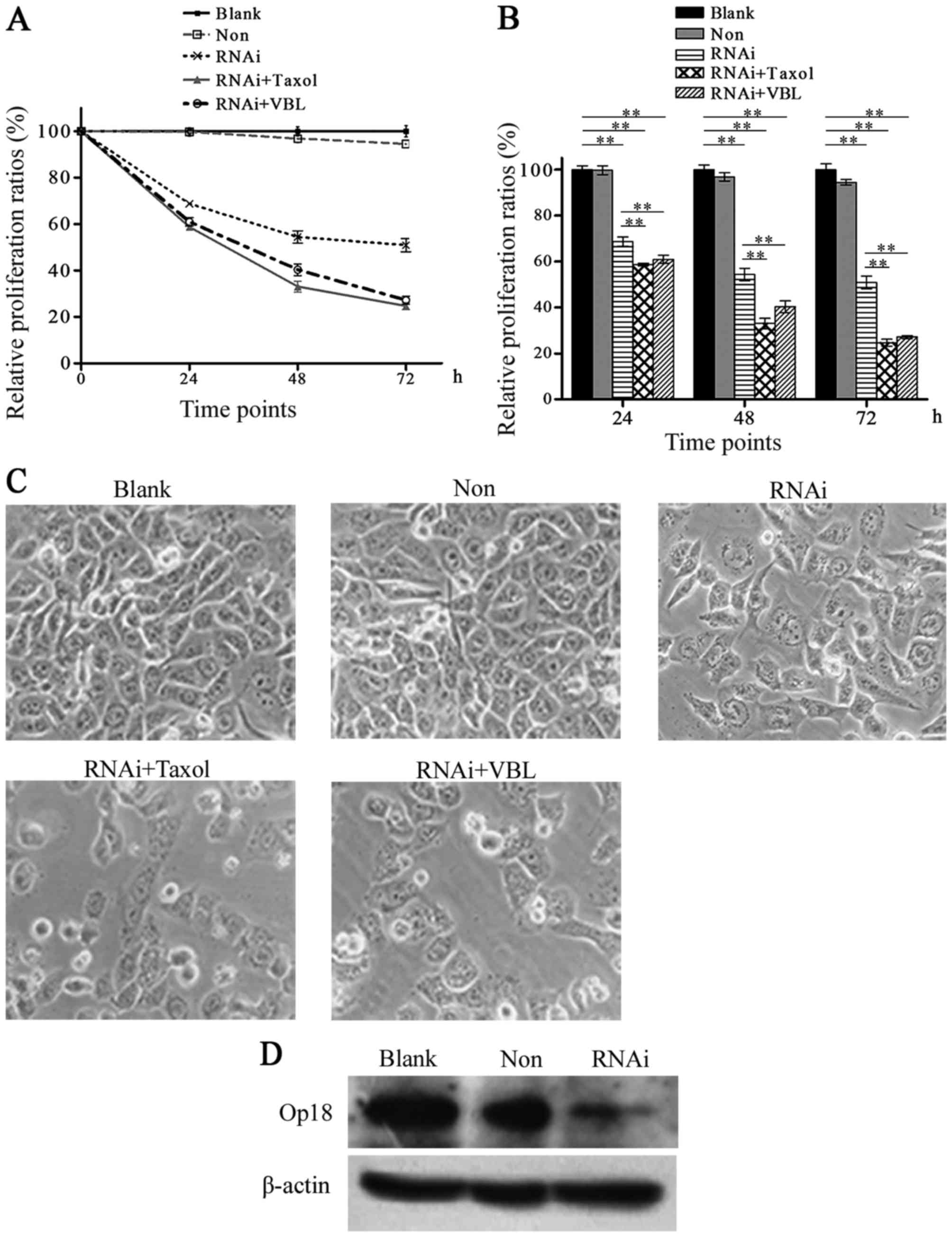
Op18/stathmin RNAi reduces the
difference in cell proliferation inhibition between VBL and
Taxol
MTT analysis demonstrated that the proliferation
ratios were 99.7, 96.79 and 94.48%, and 68.73, 54.38 and 50.88%, at
time points 24, 48 and 72 h in the non-silenced and the RNAi groups
respectively in comparison with the DMSO group. The representative
curve of the RNAi group declined rapidly, the curve of the
non-silenced group nearly overlapped the one of the Blank. After
co-treatment with Op18/stathmin RNAi and VBL or Taxol, the relative
proliferative ratios were 60.95, 40.32 and 27.16%, and 58.69, 33.20
and 24.66%, respectively, at the three time points, while the
corresponding curves were highly similar between VBL and Taxol,
which declined steeply compared with the one for single RNAi
(Fig. 5A). Histograms showed that
the difference was not significant between the Blank and the
non-silenced group, but was increased between the RNAi groups and
the blank control (p<0.01). The combination of RNAi and VBL or
Taxol significantly inhibited cell proliferation in contrast with
RNAi alone treatment (p<0.01), whereas Op18/stathmin RNAi
reduced the difference in cell proliferation inhibition between VBL
and Taxol at all time points (Fig.
5B).
Cell growth images show that cell proliferation was
obviously inhibited by RNAi, while the inhibition was increased
following co-treatment of RNAi and VBL or Taxol. Cells became much
sparser in the co-treated groups with RNAi than the RNAi alone
treatment group (Fig. 5C).
Western blot analysis showed that Op18/stathmin RNAi
effectively inhibited the expression of Op18/stathmin in the
NCI-H1299 cells (Fig. 5D).
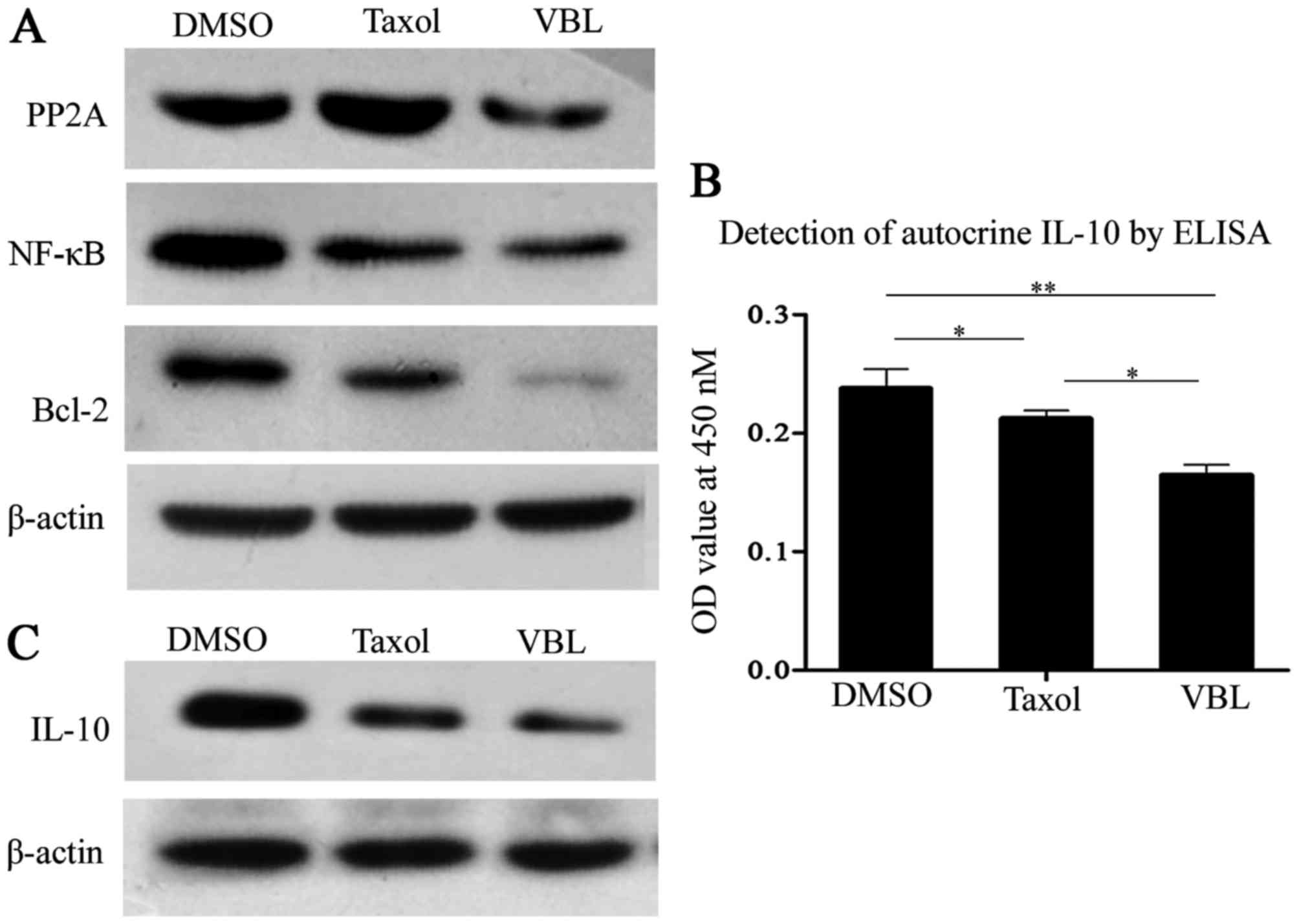
VBL markedly weakens the expression
levels of PP2A, Bcl-2, NF-κB and IL-10 and autocrine IL-10 in
comparison with Taxol
Western blot analysis showed that Taxol notably
enhanced the expression of protein phosphatase 2A (PP2A), but PP2A
was substantially decreased by VBL, while both Taxol and VBL
treatments decreased the expression of nuclear factor-κB (NF-κB),
B-cell lymphoma-2 (Bcl-2). However, the inhibitory effects were
more notable for VBL induction than Taxol (Fig. 6A).
ELISA demonstrated that autocrine IL-10 from cell
culture supernatants was decreased by Taxol and VBL, but VBL
induced more obvious inhibitory effects when compared with Taxol.
Histograms showed that the differences were significant between the
VBL and the other two treatment groups (Fig. 6B).
Western blot analysis showed that both Taxol and VBL
inhibited the expression of IL-10 in the NCI-H1299 cells, but VBL
was more effective than Taxol (Fig.
6C).
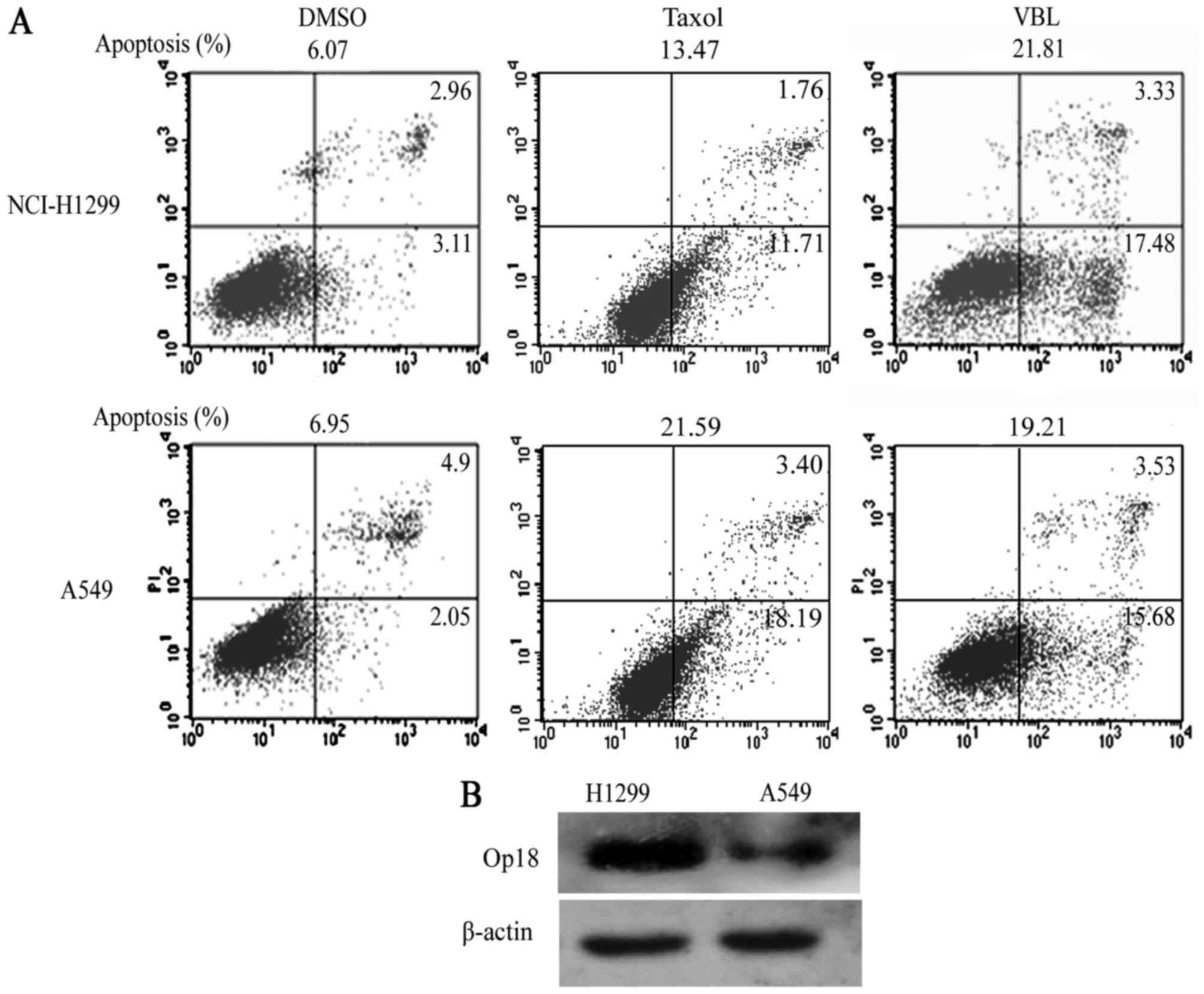
High expression of Op18/stathmin is
negatively correlated with the sensitivity of Taxol in the
different NSCLC cells, but only exerts a mild impact on VBL
FACS showed that cellular apoptotic percentages were
6.07, 13.47 and 21.81%, respectively, in the NCI-H1299 cells in the
DMSO, Taxol and VBL treatment groups, while they were 6.95, 21.59
and 19.21% respectively in A549 cells in the corresponding groups.
NCI-H1299 cells presented notable resistance to Taxol when compared
with VBL, but the sensitivity of Taxol was similar to VBL in the
A549 cells (Fig. 7A).
Western blot analysis demonstrated that NCI-H1299
cells highly expressed Op18/stathmin compared with the A549 cells
(Fig. 7B).
Discussion
Op18/stathmin is thought to be closely associated
with cellular proliferation, differentiation and drug resistance of
tumors (11–14). Our previous studies demonstrated
that Op18/stathmin is regulated by multiple kinases including
cyclin dependent 2 (CDC2) and extracellular signal regulated kinase
(ERK), and that high expression of Op18/stathmin is positively
correlated with the resistance of Taxol in different
epithelial-derived carcinoma cells (8–10).
The present study demonstrated that NCI-H1299 cells
were more sensitive to VBL than to Taxol. VBL obviously promoted
cell apoptosis through initiating the activation of caspase 3 and
9, and inhibited cell proliferation, colony formation and cell
migration in contrast with Taxol. VBL mainly decreased the activity
of Op18/stathmin by triggering the phosphorylation of Op18/stathmin
at all 4 serine sites. On the contrary, Taxol decreased the
expression of Op18/stathmin and the phosphorylation at Ser25 and
Ser63. In short, VBL, completely different from Taxol, mediates the
Op18/stathmin signaling pathway by increasing the phosphorylation
of Op18/stathmin. Other studies also confirmed that the
phosphorylation of Op18/stathmin at 4 serine residues is involved
in cell apoptosis induced by exogenous tumor necrosis factor (TNF)
in mouse fibrosarcoma L929 cells, and wee-1 knockdown promoted the
sensitivity to VBL through augmenting phosphorylation of
Op18/stathmin at Ser25 and Ser63 in human breast carcinoma BT20ST1
cells. Blocking ERK by PD98059 enhanced the sensitivity to Taxol
through decreasing the expression of Op18/stathmin and the
phosphorylation in NCI-H1299 cells (9,15,16).
Previous studies demonstrated that silencing of
Op18/stathmin by RNAi promoted the sensitivity of breast cancer
BT549 cells to Taxol and VBL as well as the sensitivity of
nasopharyngeal carcinoma CNE1 cells to Taxol and high grade
differentiated CNE1 xenografted tumors in nude mice (16,17).
The present study found that Op18/stathmin RNAi reduced the
differences in cellular proliferation inhibition between VBL and
Taxol, which implies that Op18/stathmin signaling mediates the
sensitivities of both VBL and Taxol.
PP2A is an important serine/threonine phosphatase
acting to dephosphorylate Op18/stathmin (18). NF-κB is a transcription factor
involved in cell survival and proliferation, which interacts with
Op18/stathmin to promote tumor growth and predicts poor prognosis
of pancreatic cancer (19–21). IL-10 is a pleiotropic cytokine which
plays a vital role in tumor growth and drug resistance, of which
high expression is associated with poor prognosis in tumor patients
(22–24). Bcl-2 is a typical protein against
cellular apoptosis induced by chemotherapeutics (25,26).
The present study showed that the expression of PP2A was increased
by Taxol, but decreased by VBL, which coincided with the status of
phospho-Op18/stathmin for Taxol or VBL induction, while both Taxol
and VBL inhibited the expression levels of NF-κB, Bcl-2 and IL-10
as well as autocrine IL-10 in NCI-H1299 cells, but the inhibitory
effects were more obvious for VBL than for Taxol.
It has been reported that overexpression of
Op18/stathmin markedly decreases microtubule binding with Taxol and
increased binding with VBL, downregulated the sensitivity to Taxol,
but to VBL to a lesser extent in human breast cancer cells
(27,28). Others have found that high
expression of Op18/stathmin increased the sensitivity to vindesine
and vincristine (one of vinca alkaloids) in human lung carcinoma
cells, while crystallographic analysis showed that Op18/stathmin
increased VBL binding to tubulin. Conversely, VBL also increased
Op18/stathmin binding to tubulin in vitro (29–31).
The present study indicated that high expression of Op18/stathmin
was negatively associated with the sensitivity of Taxol in
different NSCLC cell lines, but exerted slight impact on VBL
cytotoxity, which is an attractive alternative for the treatment of
Taxol-resistant tumors with highly expressed Op18/stathmin.
Acknowledgements
The present study was funded by the National Natural
Science Foundation of China (grant no. 81272274), the Key Project
of Hunan Province Scientific Research of Colleges and Universities
(no. 12A018), and the Natural Science Foundation of Hunan Province
(no. 12JJ3104).
References
|
1
|
Hennessy BT, Coleman RL and Markman M:
Ovarian cancer. Lancet. 374:1371–1382. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Takashima S, Kiyoto S, Takahashi M, Hara
F, Aogi K, Ohsumi S, Mukai R and Fujita Y: Clinical experience with
nanoparticle albumin-bound paclitaxel, a novel taxane anticancer
agent, and management of adverse events in females with breast
cancer. Oncol Lett. 9:1822–1826. 2015.PubMed/NCBI
|
|
3
|
Coderch C, Morreale A and Gago F:
Tubulin-based structure- affinity relationships for antimitotic
Vinca alkaloids. Anticancer Agents Med Chem. 12:219–225. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Koontz MZ, Horning SJ, Balise R, Greenberg
PL, Rosenberg SA, Hoppe RT and Advani RH: Risk of therapy-related
secondary leukemia in Hodgkin lymphoma: The Stanford University
experience over three generations of clinical trials. J Clin Oncol.
31:592–598. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lin XC and Cao Y: Advances in the
regulation of the rignals on Op18/stathmin. Life Science Res.
11:195–999. 2007.
|
|
6
|
Lin XC, Liao Y, Yang J, Su L, Zou H, Zuo Y
and Zou L: Regulation of the drug resistance of carcinoma cells
mediated by Op18/stathmin. Chem Life. 33:265–268. 2013.
|
|
7
|
Nemunaitis J: Stathmin 1: A protein with
many tasks. New biomarker and potential target in cancer. Expert
Opin Ther Targets. 16:631–634. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lin X, Liao Y, Xie J, Liu S, Su L and Zou
H: Op18/stathmin is involved in the resistance of Taxol among
different epithelial carcinoma cell lines. Cancer Biother
Radiopharm. 29:376–386. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lin X, Liao Y, Chen X, Long D, Yu T and
Shen F: Regulation of oncoprotein 18/stathmin signaling by ERK
concerns the resistance to Taxol in non-small cell lung cancer
cells. Cancer Biother Radiopharm. 31:37–43. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lin X, Tang M, Tao Y, Li L, Liu S, Guo L,
Li Z, Ma X, Xu J and Cao Y: Epstein-Barr virus-encoded LMP1
triggers regulation of the ERK-mediated Op18/stathmin signaling
pathway in association with cell cycle. Cancer Sci. 103:993–999.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Gupta KK, Li C, Duan A, Alberico EO, Kim
OV, Alber MS and Goodson HV: Mechanism for the
catastrophe-promoting activity of the microtubule destabilizer
Op18/stathmin. Proc Natl Acad Sci USA. 110:20449–20454. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Hsu HP, Li CF, Lee SW, Wu WR, Chen TJ,
Chang KY, Liang SS, Tsai CJ and Shiue YL: Overexpression of
stathmin 1 confers an independent prognostic indicator in
nasopharyngeal carcinoma. Tumour Biol. 35:2619–2629. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Trovik J, Wik E, Stefansson IM,
Marcickiewicz J, Tingulstad S, Staff AC, Njolstad TS, Vandenput I,
Amant F, Akslen LA, et al: MoMaTec Study Group: Stathmin
overexpression identifies high-risk patients and lymph node
metastasis in endometrial cancer. Clin Cancer Res. 17:3368–3377.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Werner HM, Trovik J, Halle MK, Wik E,
Akslen LA, Birkeland E, Bredholt T, Tangen IL, Krakstad C and
Salvesen HB: Stathmin protein level, a potential predictive marker
for taxane treatment response in endometrial cancer. PLoS One.
9:e901412014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Vancompernolie K, Boonefaes T, Mann M,
Fiers W and Grooten J: Tumor necrosis factor-induced microtubule
stabilization mediated by hyperphosphorylated oncoprotein 18
promotes cell death. J Biochem. 275:3876–3882. 2000.
|
|
16
|
Alli E, Yang JM, Ford JM and Hait WN:
Reversal of stathmin- mediated resistance to paclitaxel and
vinblastine in human breast carcinoma cells. Mol Pharmacol.
71:1233–1240. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lin X, Yu T, Zhang LX, Chen SY, Chen X,
Liao Y, Dan L and Shen F: Silencing Op18/stathmin by RNA
interference promotes the sensitivity of nasopharyngeal carcinoma
cells to taxol and high grade differentiation of xenografted
tumours in nude mice. Basic Clin Pharmacol. 2016. View Article : Google Scholar
|
|
18
|
Tournebize R, Andersen SSL, Verde F, Dorée
M, Karsenti E and Hyman AA: Distinct roles of PP1 and PP2A-like
phosphatases in control of microtubule dynamics during mitosis.
EMBO J. 16:5537–5549. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hayden MS and Ghosh S: NF-κB, the first
quarter-century: Remarkable progress and outstanding questions.
Genes Dev. 26:203–234. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hoesel B and Schmid JA: The complexity of
NF-κB signaling in inflammation and cancer. Mol Cancer. 12:862013.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lu Y, Liu C, Cheng H, Xu Y, Jiang J, Xu J,
Long J, Liu L and Yu X: Stathmin, interacting with Nf-κB, promotes
tumor growth and predicts poor prognosis of pancreatic cancer. Curr
Mol Med. 14:328–339. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Itakura E, Huang RR, Wen DR, Paul E,
Wünsch PH and Cochran AJ: IL-10 expression by primary tumor cells
correlates with melanoma progression from radial to vertical growth
phase and development of metastatic competence. Mod Pathol.
24:801–809. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Mahipal A, Terai M, Berd D, Chervoneva I,
Patel K, Mastrangelo MJ and Sato T: Tumor-derived interleukin-10 as
a prognostic factor in stage III patients undergoing adjuvant
treatment with an autologous melanoma cell vaccine. Cancer Immunol
Immunother. 60:1039–1045. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Danoch H, Kalechman Y, Albeck M, Longo DL
and Sredni B: Sensitizing B- and T- cell lymphoma cells to
paclitaxel/abraxane-induced death by AS101 via inhibition of the
VLA-4-IL10-survivin axis. Mol Cancer Res. 13:411–422. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Moldoveanu T, Follis AV, Kriwacki RW and
Green DR: Many players in BCL-2 family affairs. Trends Biochem Sci.
39:101–111. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Delbridge ARD, Grabow S, Strasser A and
Vaux DL: Thirty years of BCL-2: Translating cell death discoveries
into novel cancer therapies. Nat Rev Cancer. 16:99–109. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Alli E, Bash-Babula J, Yang JM and Hait
WN: Effect of stathmin on the sensitivity to antimicrotubule drugs
in human breast cancer. Cancer Res. 62:6864–6869. 2002.PubMed/NCBI
|
|
28
|
Purani I, Tejada A and Mistry SJ: Effects
of combination of microtubule depolymerizing peptides and
vinblastine in breast cancer. Cancer Res. 73:(Suppl 8). S3275.
2013. View Article : Google Scholar
|
|
29
|
Nishio K, Nakamura T, Koh Y, Kanzawa F,
Tamura T and Saijo N: Oncoprotein 18 overexpression increases the
sensitivity to vindesine in the human lung carcinoma cells. Cancer.
91:1494–1499. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Devred F, Tsvetkov PO, Barbier P, Allegro
D, Horwitz SB, Makarov AA and Peyrot V: Stathmin/Op18 is a novel
mediator of vinblastine activity. FEBS Lett. 582:2484–2488. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Malesinski S, Tsvetkov PO, Kruczynski A,
Peyrot V and Devred F: Stathmin potentiates vinflunine and inhibits
Paclitaxel activity. PLoS One. 10:e01287042015. View Article : Google Scholar : PubMed/NCBI
|