Introduction
Osteosarcoma (OS) is the most common primary bone
malignancy in children, adolescents and young adults with a high
tendency of local invasion and distant metastases (1,2). The
introduction of chemotherapy has led to a dramatic improvement in
prognosis for patients with localized osteosarcoma; long-term
survival rates of less than 20% improved to 65–70% after the advent
of multi-agent chemotherapy regimens in the 1980s (3). However, in the last three decades, the
efficacy of treatment has remained unchanged (4). Multiple factors are involved in the
complex process of tumorigenesis (5), thus, uncovering the molecular
mechanisms of OS may help to identify effective therapies for OS
treatment.
Membrane transporters (MTs) are proteins that
regulate the transport of endogenous molecules and xenobiotics
across the cell membrane (6).
Recently, MTs have attracted significant attention for their key
roles in tumor growth, survival relapse and drug resistance
(7,8). MTs may be grouped into various
classes, with the most important classes being ion channels
(9), ABC transporters (10), water channels (11), pumps (such as sodium potassium
pumps) (12) and the solute
carriers (SLCs), which, by far, is the largest group of
transporters (13).
SLC3A2 (solute carrier family 3 member 2), also
known as CD98 heavy chain, is a type of SLC. Recent studies have
reported that SLC3A2 is overexpressed and correlated with tumor
progression in numerous human cancers, including head and neck
squamous cell carcinoma (14),
biliary tract cancer (15),
non-small cell lung cancer (16),
and triple-negative breast cancer (17). However, the role of SLC3A2 in OS
progression is still unknown.
In this study, we report for the first time that
SLC3A2 is upregulated in human OS. Additionally, we found that
knockdown of SLC3A2 resulted in growth inhibition in OS cell lines
and led to PI3K/Akt signaling pathway dysregulation, which
indicates that SLC3A2 may play a crucial role in the growth of OS
by influencing the PI3K/Akt signaling pathway.
Materials and methods
Cell lines and cell culture
The human osteosarcoma cell lines MNNG/HOS, MG63 and
U2OS were used in this study. All cell lines were incubated at 37°C
in a humidified atmosphere containing 5% CO2. MNNG/HOS
and MG63 cells were cultured in Dulbecco's modified Eagle's medium
(DMEM) containing 10% fetal bovine serum (FBS South America origin;
Bio-West, Logan, UT, USA), 100 U/ml penicillin (Sigma-Aldrich, St.
Louis, MO, USA) and 100 mg/ml streptomycin (Sigma-Aldrich), and
U2OS cells were cultured in RPMI-1640 medium containing 10% FBS,
100 U/ml penicillin and 100 mg/ml streptomycin.
Human osteosarcoma samples
In total, 50 human osteosarcoma samples and the
adjacent non-cancerous tissues were obtained from patients who
underwent treatment at the Department of Orthopedics, Shanghai Jiao
Tong University Affiliated Sixth People's Hospital (Shanghai,
China) between 2013 and 2014. To avoid deviation from the
heterogeneity of the tumor body, we obtained tumor tissue samples
from several sections of the tumor. More than 80% of cancerous
section were obtained. The samples were frozen and stored in liquid
nitrogen after resection. All the procedures were approved by the
Ethics Committee of the Shanghai Jiao Tong University Affiliated
Sixth People's Hospital and under consensus agreement.
RNA isolation and qRT-PCR assays
The total RNA of cells and human tissue samples was
extracted using TRIzol reagent (Invitrogen, Carlsbad, CA, USA) and
quantified by Nanodrop 2000 (Thermo Fisher Scientific, Waltham, MA,
USA). The first-stand cDNA was synthesized using the PrimeScript RT
reagent kit (Takara, Shiga, Japan) and was used as a template for
the PCR detection. RT-PCR was performed with SYBR Green Premix Ex
Taq (Takara) according to the manufacturer's protocol. All
reactions were performed three times in a 10-µl reaction volume.
The expression level of genes was measured using the comparative Ct
method. The information of the primer sequences used was described
as follows: SLC3A2 forward, ACCCCTGTTTTCAGCTACGG and reverse, GGTC
TTCACTCTGGCCCTTC; β-actin forward, TTGTTACA GGAAGTCCCTTGCC and
reverse, ATGCTATCACCTCC CCTGTGTG.
Cell transfection
The siRNA targeting SLC3A2 as well as a negative
control were synthesized by RiboBio Co., Ltd. (Guangzhou, China).
The siRNA sequences targeting SLC3A2 were: forward primer,
5′-AGAUGAAGAUAGUCAAGAA-3′, and reverse primer,
5′-UUCUUGACUAUCUUCAUCU-3′. RNAi-Max (Invitrogen) was used in the
transfection. All the steps were applied in accordance with the
manual.
Cell proliferation and cell cycle
assays
For cell proliferation assay, 3000 cells transfected
with targeted siRNA or si-NC were seeded in each well of a 96-well
microplate 2 days after the transfection. To measure the cell
viability, we added 10 µl Cell Counting Kit-8 (CCK-8, Dojindo
Molecular Technologies, Inc., Kumamoto, Japan) and 100 µl DMEM (for
MNNG/HOS) or RPMI-1640 (for U2OS) to each well and incubated for 2
h. The absorbance at a wavelength of 450 nm was measured by a
microplate reader (Model 680, Bio-Rad Laboratories, Hercules, CA,
USA). For cell cycle assay, cells were fixed in 70% ethanol for 24
h at −20°C two days after the transfection. Then, staining solution
containing 50 µg/ml RNase A (BD LSRII, San Jose, CA, USA) and 50
µg/ml propidium iodide (PI) (Biolegend, San Diego, CA, USA) was
used to treat the cells. All the experiments were performed three
times.
Colony formation assay
OS cells transfected with targeted siRNA or si-NC
were collected 2 days after the transfection and were planted in
6-well plates at a density of 1000 cell/well. The cells were
cultured for 2 weeks at a constant temperature of 37°C in a
humidified atmosphere containing 5% CO2 to form
colonies. Colonies were fixed in 95% methanol and stained with 0.1%
crystal violet for 20 min, respectively. The colonies were then
counted and photographed. The assay was repeated three times.
Western blot analysis and PathScan
intracellular signaling array kit
A mixture of PhosSTOP (Roche, Basel, Switzerland),
T-PER protein extraction reagent (Thermo Fisher Scientific) and
Complete Mini (Roche) was used to extract the cell lysates. Equal
amounts of protein (20 µg) were loaded and separated using 8%
sodium dodecyl sulfate polyacrylamide gel electrophoresis
(SDS-PAGE) and then transferred to nitrocellulose membranes
(Millipore). After blocking in 5% milk with phosphate-buffered
saline for 1 h, the membranes were incubated with primary
antibodies listed below at 4°C overnight: SLC3A2 (Proteintech,
1:500), PI3-kinase (Phospho-Tyr607) (Signalway Antibody, 1:500),
Akt (Phospho-Ser473, 1:500) or β-actin (Sigma-Aldrich, 1:20000).
The secondary antibody was anti-rabbit IgG (Sigma-Aldrich, 1:5000).
Next, the detection of the blots was performed using SuperSignal
West Femto Maximum Sensitivity Substrate (Thermo Fisher
Scientific). To analyze the signaling pathways influenced by SLC3A2
expression, we used PathScan Intracellular Signaling Array kit
(Cell Signaling Technology). This system is a slide-based antibody
array, based upon the sandwich immunoassay principle and allowing
for the simultaneous detection of 18 important well-characterized
signaling molecules when phosphorylated or cleaved. The experiment
was performed according to the manufacturer's instructions, and the
fluorescent images of the slide were then captured with Image
Studio Software (Licensor Inc.). Images were analyzed with ImageJ
software. Normalization was performed by subtracting the intensity
of the negative control dot from each value.
Immunohistochemistry (IHC)
As previously described (18), immunohistochemistry was performed
with the following steps: After being warmed in a 60°C oven,
de-waxed in three changes of xylene, and passaged through graded
ethanol (100, 95, and 70%), formalin-fixed and paraffin-embedded
sections came to a final wash in distilled water. Hydrogen peroxide
(3%) was used to quench the endogenous peroxidase acivity and BSA
was used to block for 30 min. The sections were incubated with an
antibody against SLC3A2 at a 1:50 dilution overnight in 4°C. The
slides were mounted for microscopic evaluation and photographed
after being counterstained with Gill's hematoxylin for 1 min and
dehydrated. The staining results for SLC3A2 protein were scored
according to an established 0–3 scale as listed below: 0, no
staining; 1, staining in <1% cells; 2, staining in 1–10% cells;
3, staining in >10% of cells. We considered the samples graded 0
and 1 as negative, and those graded as 2 and 3 as positive. In
total, 10 optical fields from three different sections were used
for each evaluation by two experienced colleagues,
respectively.
Statistical analysis
Data were analyzed with SPSS software (version 20.0)
(IBM Corp., Armonk, NY, USA). Student's t-test was used to compare
the differences between groups. Pearson's Chi-square or Continuity
Correction test was used to calculate the correlations between
parameters. All the data were graphically imaged by GraphPad Prism
5 software (Graphpad Software, Inc., La Jolla, CA, USA).
Results
Expression of SLC3A2 is upregulated in
OS clinical samples and cell lines
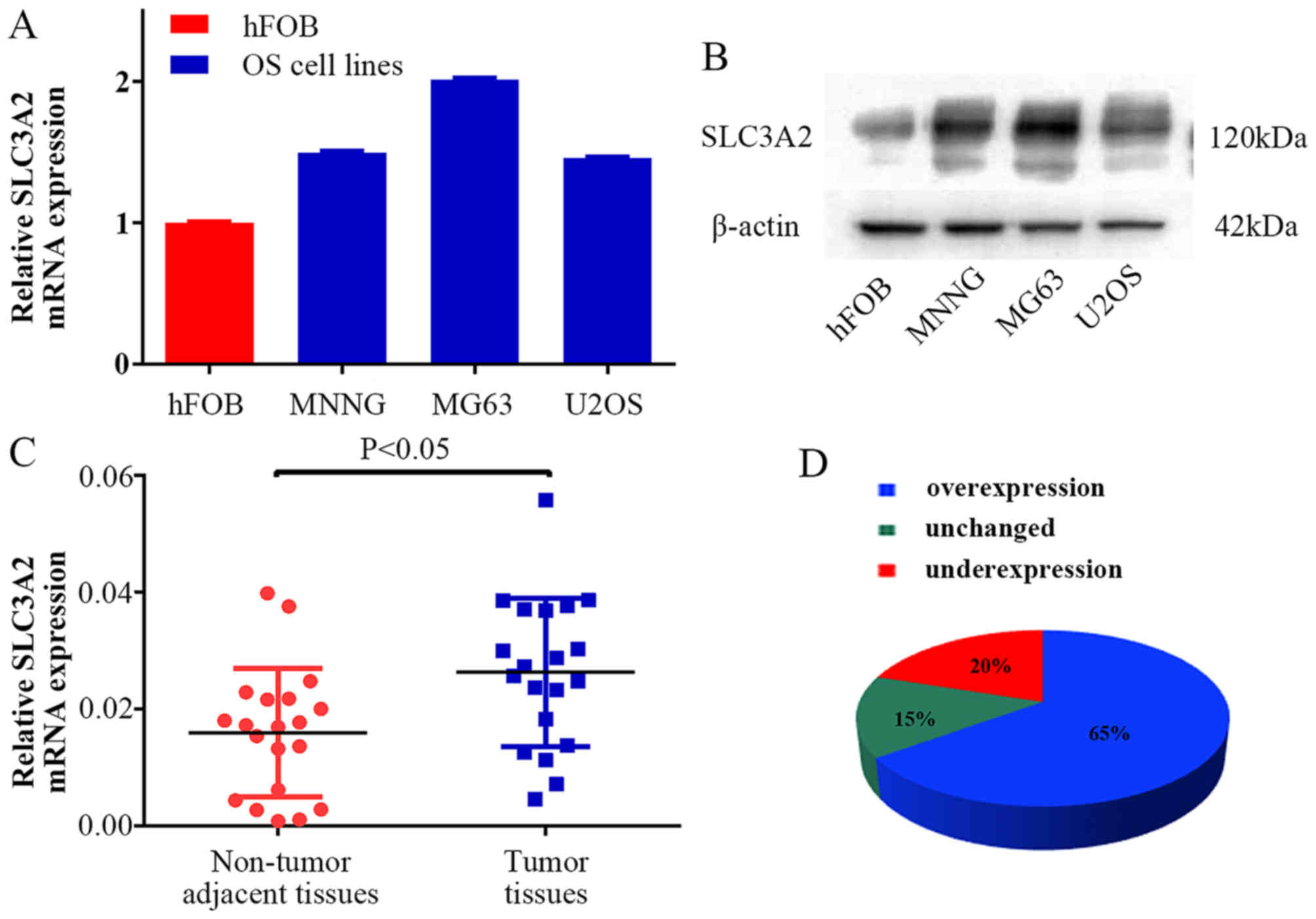
To clarify the expression level of SLC3A2 in OS, we
examined SLC3A2 expression at the mRNA and protein levels,
respectively. As a result, higher expression of SLC3A2 was observed
in the OS cell lines (MNNG/HOS, MG63, U2OS) compared with that
noted in the human osteoblast cell line (hFOB) (Fig. 1A and B). Moreover, the expression of
SLC3A2 in 20 pairs of human OS tissue samples and their
corresponding non-cancerous tissue controls were also analyzed
using quantitative RT-PCR (qRT-PCR). As shown in Fig. 1C and D, SLC3A2 expression was
significantly upregulated in tumor tissue samples compared to that
noted in the corresponding controls (65%, 13/20).
Knockdown of SLC3A2 leads to reduced
proliferation and cell cycle arrest in the G2/M phase
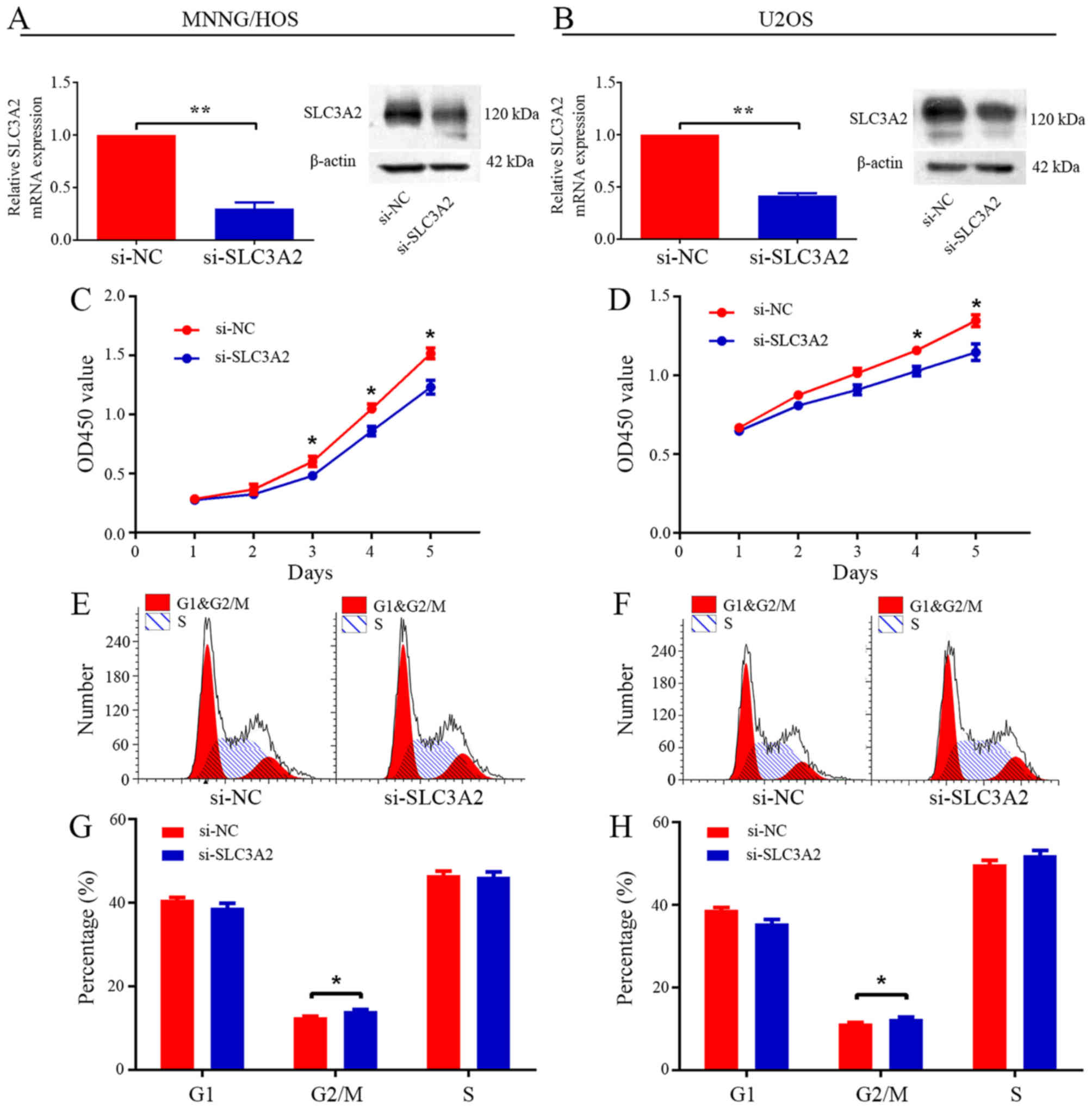
To ascertain the underlying role of SLC3A2 in
tumorigenesis, we first evaluated the effect of SLC3A2 on the
growth of OS cells in vitro. After knockdown of SLC3A2 by
specifically targeted siRNA (Fig. 2A
and B), the viability of the MNNG/HOS and U2OS cells was
clearly inhibited, as determined by CCK-8 assay (Fig. 2C and D). Cell cycle distribution was
analyzed by flow cytometry after transfection with si-NC or
si-SLC3A2. The results of the cell cycle analysis revealed that
knockdown of SLC3A2 expression resulted in G2/M phase arrest in
both the MNNG/HOS and U2OS cell lines (Fig. 2E-H). Thus, these results indicate
that SLC3A2 is essential for the proliferation of OS cells.
Silencing of SLC3A2 attenuates colony
formation capacity of OS cells
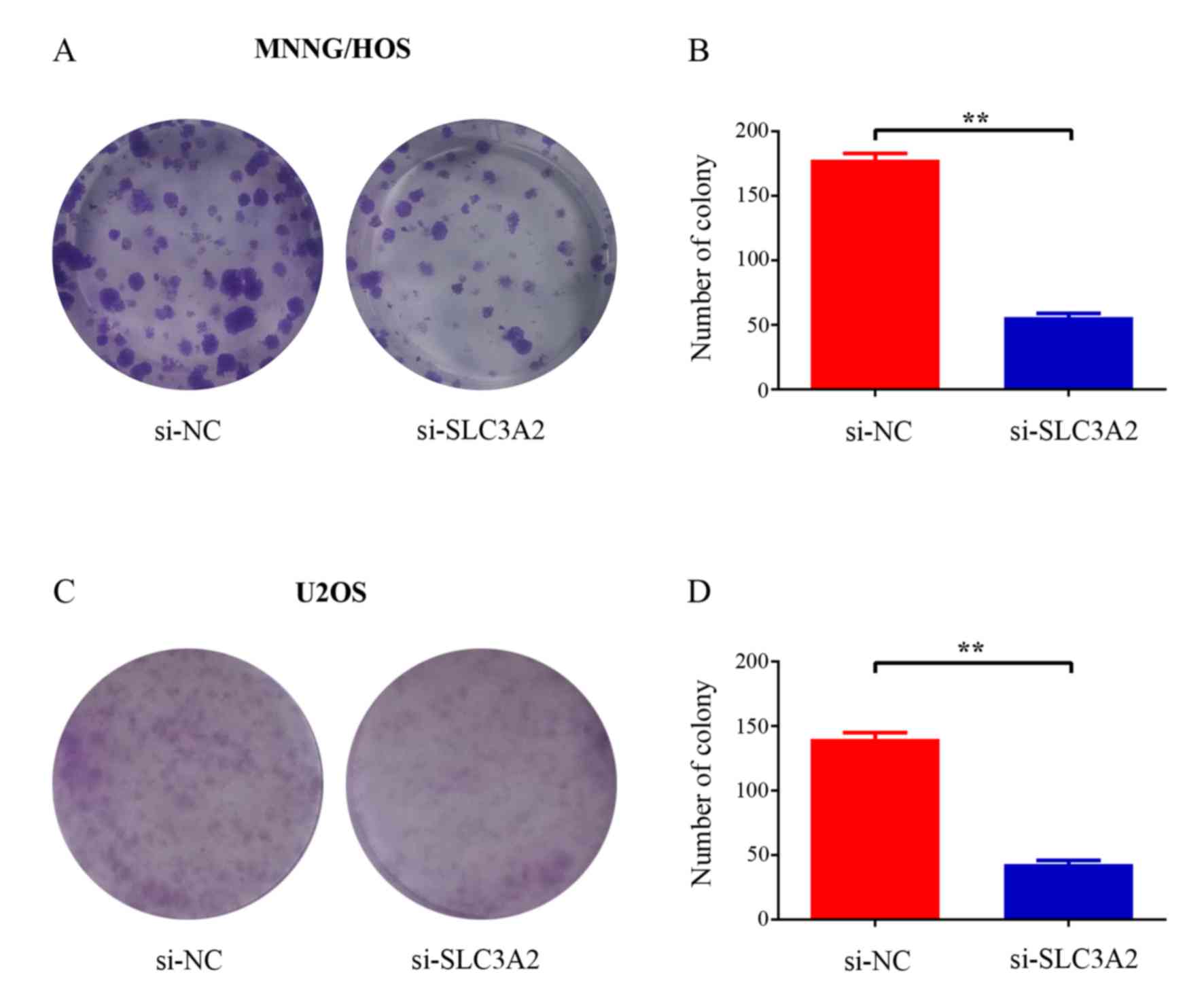
To further elucidate the potential role of SLC3A2 in
tumorigenesis, we determined whether the knockdown of SLC3A2
expression affects the colony formation ability of OS cells by
colony formation assay. As a result, both the size and the number
of colonies were markedly reduced in the SLC3A2-knockdown group
compared with these parameters in the control group (Fig. 3A and C). The number of colonies was
significantly reduced by 68.5 and 69.3% in the MNNG/HOS and U2OS
cells, respectively (Fig. 3B and
D). Taken together, these results demonstrated that SLC3A2 is
important for OS cell growth.
SLC3A2 regulates the PI3K/AKT
signaling pathway in OS cells
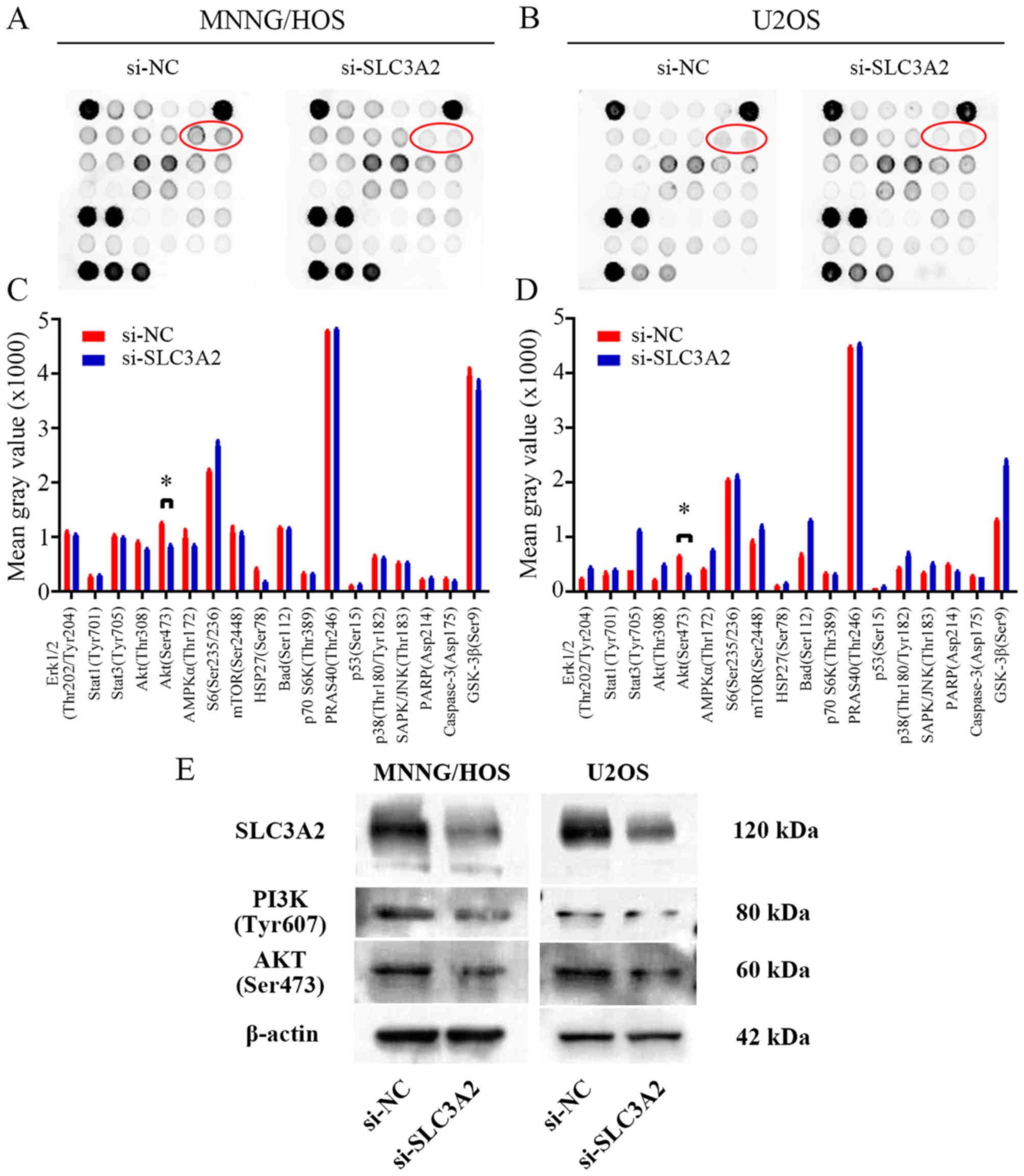
To further seek the potential molecular mechanism
through which knockdown of SLC3A2 expression suppresses OS cell
proliferation, we performed a signaling antibody array assay. As
shown in Fig. 4A-D, knockdown of
SLC3A2 expression led to reduced expression of p-AKT in the
MNNG/HOS and U2OS cell lines, suggesting that SLC3A2 may promote OS
cell proliferation through the PI3K/AKT signaling pathway. These
results were further supported by western blot analysis as shown in
Fig. 4E, where the expression of
PI3K and AKT were both downregulated by inhibition of SLC3A2.
High expression of SLC3A2 correlates
with OS clinical stage and tumor size
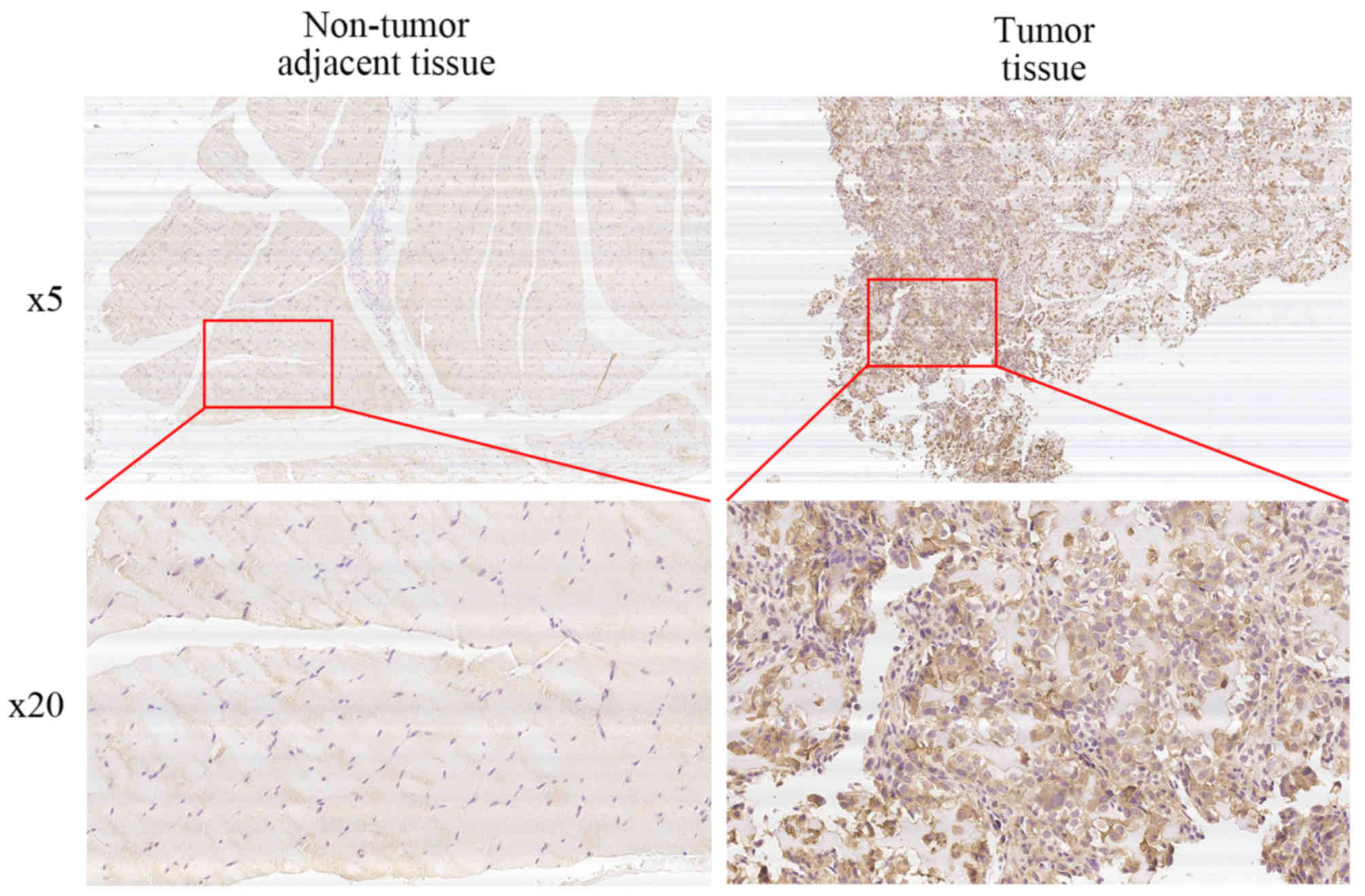
We performed immunohistochemical staining on 50
human osteosarcoma tissues and their corresponding non-cancerous
tissue controls. Representative examples of IHC for SLC3A2 in the
OS and non-cancerous tissues are shown in Fig. 5. The positive rate of IHC among OS
and non-cancerous tissues was 58% (29/50) and 14% (7/50),
respectively. In addition, 62% (31/50) of the matched cases showed
high SLC3A2 expression in the OS tissues compared to that observed
in the non-cancerous tissues. We then conducted stratified analyses
to assess SLC3A2 expression in the OS patients with specific
clinical characteristics as shown in Table I. While SLC3A2 expression was not
correlated with age, gender, anatomic location, degree of
malignancy or tumor necrosis rate, there were significant
correlations with clinical stage (P<0.05) and tumor size
(P<0.05) which achieved statistical significance. Taken
together, these results indicate that SLC3A2 is upregulated and
potentially plays a pivotal role in the growth and survival of
OS.
 | Table I.Correlation of the immunohistochemical
staining (IHC) for SLC3A2 and clinicopathologic parameters in 50
osteosarcoma patients. |
Table I.
Correlation of the immunohistochemical
staining (IHC) for SLC3A2 and clinicopathologic parameters in 50
osteosarcoma patients.
|
|
| IHC of SLC3A2 |
|
|
|
|
|---|
|
|
|
|
|
|
|
|
|---|
| Clinicopathologic
parameters | Number of cases | Positive | Negative | χ2 | P-value | Odds ratio | 95% CI |
|---|
| Age (years) |
|
|
|
|
|
|
|
|
<20 | 31 | 19 | 12 | 0.363 | 0.547 | – | – |
|
≥20 | 19 | 10 | 9 |
|
|
|
|
| Gender |
|
|
|
|
|
|
|
|
Male | 27 | 13 | 14 | 2.339 | 0.126 | – | – |
|
Female | 23 | 16 | 7 |
|
|
|
|
| Anatomic
location |
|
|
|
|
|
|
|
| Tibia/
Femur | 33 | 21 | 12 | 1.266 | 0.261 | – | – |
|
Elsewhere | 17 | 8 | 9 |
|
|
|
|
| Enneking stage |
|
|
|
|
|
|
|
|
I/II | 35 | 16 | 19 | 5.645 | 0.018a | 7.719 | 1.512–39.42 |
|
III | 15 | 13 | 2 |
|
|
|
|
| Tumor size
(cm3) |
|
|
|
|
|
|
|
|
<50 | 21 | 8 | 13 | 5.889 | 0.015a | 4.266 | 1.285–14.16 |
|
≥50 | 29 | 21 | 8 |
|
|
|
|
| Degree of
malignancy |
|
|
|
|
|
|
|
|
Low | 23 | 10 | 13 | 3.687 | 0.055 | – | – |
|
High | 27 | 19 | 8 |
|
|
|
|
| Tumor necrosis rate
(%) |
|
|
|
|
|
|
|
|
<90 | 27 | 17 | 10 | 0.593 | 0.441 | – | – |
|
≥90 | 23 | 12 | 11 |
|
|
|
|
Discussion
According to current knowledge, SLC proteins can
translocate diverse endogenous substrates, drugs and environmental
toxicants. As a result, they play crucial roles in the development
of tumor progression and therapy (19). Multiple SLC transporters have been
investigated in recent studies. A study by Babu et al
demonstrated that SLC6A14 is crucial for the maintenance of amino
acid nutrition and optimal mammalian target of rapamycin (mTOR)
signaling in ER+ breast cancer (20). Chen et al reported that
genomic polymorphisms of SLC29A3 are associated with overall
survival in advanced non-small cell lung cancer treated with
gemcitabine (21). Salaün et
al revealed that SLC20A1 has a direct biological role in the
downstream apoptotic signaling pathway induced by tumor necrosis
factor (TNF) (22).
In the present study, we demonstrated that SLC3A2,
one of the SLC transporters, was significantly upregulated in OS
clinical samples compared with that noted in non-cancerous tissues.
Moreover, we found that the expression of SLC3A2 was correlated
with OS Enneking stage and tumor size according to IHC staining. To
date, upregulation of SLC3A2 has been found in several different
tumor types. Garber et al examined the global gene
expression profiles of 67 human lung tumors and found that SLC3A2
was overexpressed in adenocarcinomas of the lung (23). Prager et al reported
increased SLC3A2 expression in different types of malignant renal
cell cancers (RCCs), lack of expression in benign renal oncocytoma
and the extent of expression were correlated directly with the
grade of malignancy (24). Yang
et al confirmed the upregulation of SLC3A2 in gastric cancer
cells by immunoblotting of a panel of 13 gastric cancer cell lines
and immunohistochemistry of tissue microarrays comprising 85
matched pairs of normal and tumor tissues (25). However, the association of SLC3A2
expression with OS has not yet been reported.
To investigate the function of SLC3A2 in OS cells,
we suppressed its expression using a targeted siRNA. Our
experimental data revealed that knockdown of SLC3A2 inhibited cell
proliferation through G2/M phase arrest, and impaired OS cell
colony formation capacity. Taken together, these results indicate
that SLC3A2 is essential for OS cell survival and acts as an
oncogene in OS. Consistently, SLC3A2 has been reported to enhance
proliferation in other types of cells. Wu et al recently
indicated that SLC3A2 was highly expressed and co-localized with
basigin on the human hepatocellular carcinoma (HCC) cell membrane,
and plays a critical role in promoting cell spreading and the
progression of hepatocellular carcinoma (26). SLC3A2 is also required for
proliferation of B and T lymphocytes (27,28),
vascular smooth muscle cells (29)
and intestinal epithelial cells (30).
It was previously reported that SLC3A2 is an
integrin-associated protein and contributes to integrin-dependent
cell spreading, cell migration, and protection from apoptosis
through activation of Akt and Rac GTPase, major contributors to
integrin-dependent signals involved in cell survival and cell
migration (31). Bulus et al
demonstrated that overexpression of SLC3A2 increased renal tubular
epithelial cell proliferation through Erk and p38 MAPK signaling
pathway (32). To further seek the
molecular mechanism by which SLC3A2 mediates OS cell proliferation,
we used a slide-based antibody array, which allows for the
simultaneous detection of 18 important and well-characterized
signaling molecules when phosphorylated or cleaved (33,34).
Reduced expression of Akt (Ser473) was found after knockdown of
SLC3A2 in both MNNG/HOS and U2OS cell lines. We then considered the
PI3K/Akt signaling pathway, which is crucial for cell growth and
survival (35). Western blotting
further affirmed our hypothesis that SLC3A2 influences OS
progression by regulating the PI3K/Akt signaling pathway.
In conclusion, we demonstrated in the present study
for the first time that SLC3A2 was upregulated in OS cell lines and
OS tissues, and that SLC3A2 expression was correlated with tumor
size and clinical stage of human OS. Knockdown of the expression of
SLC3A2 inhibited OS cell proliferation through the PI3K/Akt
signaling pathway. Importantly, these findings provide insight into
a novel therapeutic target for OS.
References
|
1
|
Moore DD and Luu HH: Osteosarcoma. Cancer
Treat Res. 162:65–92. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
He JP, Hao Y, Wang XL, Yang XJ, Shao JF,
Guo FJ and Feng JX: Review of the molecular pathogenesis of
osteosarcoma. Asian Pac J Cancer Prev. 15:5967–5976. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Isakoff MS, Bielack SS, Meltzer P and
Gorlick R: Osteosarcoma: Current treatment and a collaborative
pathway to success. J Clin Oncol. 33:3029–3035. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Allison DC, Carney SC, Ahlmann ER,
Hendifar A, Chawla S, Fedenko A, Angeles C and Menendez LR: A
meta-analysis of osteosarcoma outcomes in the modern medical era.
Sarcoma. 2012:7048722012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Baumhoer D: Molecular characterization of
osteosarcomas. Pathologe. 34:(Suppl 2). 260–263. 2013.(In German).
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ho RH and Kim RB: Transporters and drug
therapy: Implications for drug disposition and disease. Clin
Pharmacol Ther. 78:260–277. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Abdulhussein AA and Wallace HM: Polyamines
and membrane transporters. Amino Acids. 46:655–660. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zinzi L, Contino M, Cantore M, Capparelli
E, Leopoldo M and Colabufo NA: ABC transporters in CSC membranes as
a novel target for treating tumor relapse. Front Pharmacol.
5:1632014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yu FH and Catterall WA: The VGL-chanome: A
protein superfamily specialized for electrical signaling and ionic
homeostasis. Sci STKE. 2004:re152004.PubMed/NCBI
|
|
10
|
Montanari F and Ecker GF: Prediction of
drug-ABC-transporter interaction - Recent advances and future
challenges. Adv Drug Deliv Rev. 86:17–26. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wang F, Feng XC, Li YM, Yang H and Ma TH:
Aquaporins as potential drug targets. Acta Pharmacol Sin.
27:395–401. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Dunbar LA and Caplan MJ: Ion pumps in
polarized cells: Sorting and regulation of the Na+,
K+- and H+, K+-ATPases. J Biol
Chem. 276:29617–29620. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Fredriksson R, Nordström KJ, Stephansson
O, Hägglund MG and Schiöth HB: The solute carrier (SLC) complement
of the human genome: Phylogenetic classification reveals four major
families. FEBS Lett. 582:3811–3816. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Martens-de Kemp SR, Brink A, Stigter-van
Walsum M, Damen JM, Rustenburg F, Wu T, van Wieringen WN,
Schuurhuis GJ, Braakhuis BJ, Slijper M, et al: CD98 marks a
subpopulation of head and neck squamous cell carcinoma cells with
stem cell properties. Stem Cell Res (Amst). 10:477–488. 2013.
View Article : Google Scholar
|
|
15
|
Kaira K, Sunose Y, Oriuchi N, Kanai Y and
Takeyoshi I: CD98 is a promising prognostic biomarker in biliary
tract cancer. Hepatobiliary Pancreat Dis Int. 13:654–657. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Fei F, Li X, Xu L, Li D, Zhang Z, Guo X,
Yang H, Chen Z and Xing J: CD147-CD98hc complex contributes to poor
prognosis of non-small cell lung cancer patients through promoting
cell proliferation via the PI3K/Akt signaling pathway. Ann Surg
Oncol. 21:4359–4368. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Furuya M, Horiguchi J, Nakajima H, Kanai Y
and Oyama T: Correlation of L-type amino acid transporter 1 and
CD98 expression with triple negative breast cancer prognosis.
Cancer Sci. 103:382–389. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lin S, Guo Q, Wen J, Li C, Lin J, Cui X,
Sang N and Pan J: Survival analyses correlate stanniocalcin 2
overexpression to poor prognosis of nasopharyngeal carcinomas. J
Exp Clin Cancer Res. 33:262014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
He L, Vasiliou K and Nebert DW: Analysis
and update of the human solute carrier (SLC) gene superfamily. Hum
Genomics. 3:195–206. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Babu E, Bhutia YD, Ramachandran S,
Gnanaprakasam JP, Prasad PD, Thangaraju M and Ganapathy V: Deletion
of the amino acid transporter Slc6a14 suppresses tumour growth in
spontaneous mouse models of breast cancer. Biochem J. 469:17–23.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Chen X, Zhang L, Ren S, Li X, Zhou F, Li
W, Gao G, He Y and Zhou C: Genomic polymorphisms of SLC29A3
associated with overall survival in advanced non-small-cell lung
cancer treated with gemcitabine. Med Oncol. 31:8652014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Salaün C, Leroy C, Rousseau A, Boitez V,
Beck L and Friedlander G: Identification of a novel
transport-independent function of PiT1/SLC20A1 in the regulation of
TNF-induced apoptosis. J Biol Chem. 285:34408–34418. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Garber ME, Troyanskaya OG, Schluens K,
Petersen S, Thaesler Z, Pacyna-Gengelbach M, van de Rijn M, Rosen
GD, Perou CM, Whyte RI, et al: Diversity of gene expression in
adenocarcinoma of the lung. Proc Natl Acad Sci USA. 98:13784–13789.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Prager GW, Poettler M, Schmidinger M,
Mazal PR, Susani M, Zielinski CC and Haitel A: CD98hc (SLC3A2), a
novel marker in renal cell cancer. Eur J Clin Invest. 39:304–310.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yang Y, Toy W, Choong LY, Hou P, Ashktorab
H, Smoot DT, Yeoh KG and Lim YP: Discovery of SLC3A2 cell membrane
protein as a potential gastric cancer biomarker: Implications in
molecular imaging. J Proteome Res. 11:5736–5747. 2012.PubMed/NCBI
|
|
26
|
Wu B, Wang Y, Yang XM, Xu BQ, Feng F, Wang
B, Liang Q, Li Y, Zhou Y, Jiang JL, et al: Basigin-mediated
redistribution of CD98 promotes cell spreading and tumorigenicity
in hepatocellular carcinoma. J Exp Clin Cancer Res. 34:1102015.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Cantor J, Browne CD, Ruppert R, Féral CC,
Fässler R, Rickert RC and Ginsberg MH: CD98hc facilitates B cell
proliferation and adaptive humoral immunity. Nat Immunol.
10:412–419. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
28
|
Cantor J, Slepak M, Ege N, Chang JT and
Ginsberg MH: Loss of T cell CD98 H chain specifically ablates T
cell clonal expansion and protects from autoimmunity. J Immunol.
187:851–860. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Fogelstrand P, Féral CC, Zargham R and
Ginsberg MH: Dependence of proliferative vascular smooth muscle
cells on CD98hc (4F2hc, SLC3A2). J Exp Med. 206:2397–2406. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Nguyen HT, Dalmasso G, Torkvist L,
Halfvarson J, Yan Y, Laroui H, Shmerling D, Tallone T, D'Amato M,
Sitaraman SV, et al: CD98 expression modulates intestinal
homeostasis, inflammation, and colitis-associated cancer in mice. J
Clin Invest. 121:1733–1747. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
31
|
Feral CC, Nishiya N, Fenczik CA, Stuhlmann
H, Slepak M and Ginsberg MH: CD98hc (SLC3A2) mediates integrin
signaling. Proc Natl Acad Sci USA. 102:355–360. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Bulus N, Feral C, Pozzi A and Zent R: CD98
increases renal epithelial cell proliferation by activating MAPKs.
PLoS One. 7:e400262012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Montero JC, Esparís-Ogando A, Re-Louhau
MF, Seoane S, Abad M, Calero R, Ocaña A and Pandiella A: Active
kinase profiling, genetic and pharmacological data define mTOR as
an important common target in triple-negative breast cancer.
Oncogene. 33:148–156. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Yao W, Guan M, Jia J, Dai W, Lay YA,
Amugongo S, Liu R, Olivos D, Saunders M, Lam KS, et al: Reversing
bone loss by directing mesenchymal stem cells to bone. Stem Cells.
31:2003–2014. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Porta C, Paglino C and Mosca A: Targeting
PI3K/Akt/mTOR Signaling in Cancer. Front Oncol. 4:642014.
View Article : Google Scholar : PubMed/NCBI
|