Introduction
Pancreatic ductal adenocarcinoma (PDAC) is one of
the most lethal and malignant tumors of the digestive system. The
rate of successful PDAC resection is less than 20% and most
patients present with disease recurrence. The 5-year survival rate
of PDAC is only ~5% despite therapeutic advances in diagnosis and
treatment (1). PDAC is the fourth
leading cause of cancer-related deaths in the US with an estimate
of 37,390 deaths closely following the number of 43,920 diagnoses
in 2012 (2) and an estimate of
38,460 deaths closely following the number of 45,220 diagnoses in
2013 (3). Moreover, PDAC has been
projected to advance as one of the leading 3 cancer killers in 2030
(4). PDAC is characterized by
infiltration of polymorphonuclear neutrophils (PMNs) into the
desmoplastic stroma in most cancer samples. PMN infiltration may
create a pro-inflammatory microenvironment affecting tumor
progression by diverse mechanisms (5,6).
PMNs consist of various enzymes, including human
neutrophil elastase (HNE) (7,8). HNE
is a neutral protease of the serine protease superfamily and is
chiefly stored in neutrophilic granulocytes. In addition, various
cancer cells contain HNE (9,10).
Normally, HNE does not injure cells, but HNE may have an important
regulatory role in local inflammatory responses and dysregulation
causing HNE to be accumulated and produce chronic inflammatory
disease (11). Inflammation
contributes to numerous cancers such as lung, bladder,
gastrointestinal tract, skin and vulva cancer (12). Inflammation may promote cancer
growth, tissue invasion and metastasis, characterized by reactive
oxygen species and cell cycling induction. Cancer arising from
inflammation may be associated with a worse prognosis (13). α1-antitrypsin, an endogenous
inhibitor of HNE, is implicated in α1-antitrypsin-HNE imbalances
and this is thought to create a favorable tissue environment for
carcinogens and tumor progression, promoting the growth, survival,
metastasis of tumor cells (11,14).
Inhibition of HNE by its specific inhibitor was found to
significantly suppress growth, motility and chemotaxis of the
pancreatic carcinoma cell line Capan-1 (15).
Caffeic acid phenethyl ester (CAPE) is an active
component of the honey bee resin, propolis, and is reported to have
anti-inflammatory, neuroprotective, hepatoprotective,
cardioprotective, immunomodulatory and anticancer effects (16,17).
The tumor-specific cytotoxicity of CAPE has been reported as CAPE
can inhibit tumor cell growth, infiltration and metastasis in
vivo and in vitro, including central nervous system,
gastrointestinal system and breast cancers as well as leukemias
(18,19). CAPE modulates the inflammatory
response by reducing c-jun-N-terminal kinase and nuclear factor
(NK)-κB activation and decreasing cyclooxygenase (COX)-2 expression
(16). However, how this occurs is
unclear. Thus, we investigated the contribution of inflammation to
pancreatic cancer progression and the novel effects of CAPE on HNE,
as well as the interaction between CAPE and HNE.
Materials and methods
Cell culture and treatment
The PDAC PANC-1 cell line was used for in
vitro experiments. Cells were maintained in Dulbeccos modified
essential medium (DMEM) plus 10% fetal bovine serum (FBS) (both
from Gibco, Grand Island, NY, USA), 100 U/ml penicillin and 100
mg/ml streptomycin in a humidified incubator with 5% CO2
in air at 37̊C. Cells were treated with different concentrations of
HNE (Merck Group, Darmstadt, Germany) and/or CAPE (R&D
Systems/Tocris, Minneapolis, MN, USA) for 24 or 48 h, and then
evaluated.
Cell viability assay
Cytotoxicity was measured using the CellTiter
96® AQueous One Solution Cell Proliferation
Assay kit (Promega, Madison, WI, USA). Briefly, PANC-1 cells
growing in log-phase were trypsinized and seeded at 5,000
cells/well into 96-well plates, and allowed to adhere for 24 h.
Media were replaced by fresh medium or medium with HNE and/or CAPE,
and incubated for 48 h. One-fifth of the volume of CellTiter
96® AQueous One solution was added to each
well and incubated for an additional 3 h. The resulting color was
assayed at 490 nm using a microplate reader (Bio-Rad Laboratories,
Inc., Hercules, CA, USA). Blank control wells were used for zeroing
the absorbance.
Wound-healing assay of PANC-1 cell
migration
PANC-1 cells stably transfected with GFP were seeded
into 6-well plates. Confluent monolayers were scratched using a
sterile 1,000-µl pipette tip to create a uniform cell-free zone in
each well. Suspended cells were removed by washing with normal
growth medium. Wounded monolayers were then incubated in the
presence or absence of 10 nM HNE and decreasing concentrations of
CAPE (27 µM with 1.5-fold dilutions to 8 µM). A scratch wound was
captured after 24 or 48 h using an Olympus microscope in 3 fields
of view at a magnification of ×40. The recovered area was measured
using PhotoShop software.
Elastase assay
HNE activity was measured with an enzyme assay in
96-multiwell plates. Briefly, 50 µl substrate solution (1.4 mM
MeO-Suc-Ala-Ala-Pro-Val-pNA in Tris-HCl buffer, pH 7.5) was mixed
with 140 µl test solution (test substances solubilized in Tris-HCl
buffer, pH 7.5) and vortexed. After the addition of 10 µl HNE
solution, the samples were incubated for 1 h at 37̊C. The reaction
was stopped by the addition of 200 µl soybean trypsin inhibitor
solution (2 mg/ml Tris-HCl buffer, pH 7.5) and placed in an ice
bath. After vortexing, the absorbance was read at 405 nm.
Inhibition rates were calculated as a percentage of the controls
without inhibitors (20).
Sivelestat (Siv) sodium was added as a positive control.
Western blotting
Total protein was extracted using RIPA lysis
(Applygen Technologies, Inc., Beijing, China) buffer from PANC-1
cell lysates after co-incubation of 40 nM HNE and CAPE (18–60 µM)
for 48 h. After centrifuging and boiling at 100̊C for 5 min, equal
amounts of proteins were separated by 10% SDS-PAGE and transferred
to a polyvinylidene difluoride (PVDF) membrane (Millipore,
Billerica, MA, USA) for analysis of α1-antitrypsin (1:500; Santa
Cruz Biotechnology, Inc., Santa Cruz, CA, USA) and GAPDH (1:10,000;
Sigma-Aldrich, St. Louis, MO, USA). Slides were fixed and incubated
with the primary antibodies overnight at 4̊C with gentle agitation,
followed by secondary antibody reactions with DyLight 800-labeled
IgG [1:10,000; Cell Signaling Technology (CST) Danvers, MA, USA)].
Detection was evaluated using the Odyssey® SA Infrared
Imaging System.
RNA isolation and quantitative
real-time PCR
Total RNA was extracted using TRIzol reagent
(Invitrogen, Carlsbad, CA, USA) from PANC-1 cells at 80% confluence
after the co-incubation of 40 nM HNE and CAPE (18–60 µM) for 24 h.
Then, total RNA was retrotranscribed into first-strand cDNA using
the RevertAid First Strand cDNA Synthesis kit (Fermentas,
Burlington, ON, Canada). The primers used for quantitative
real-time PCR were as follows: α1-antitrypsin (sense primer,
5-CCCCACCCAGAGTTGCTC-3 and antisense primer,
5-GGTTAGGTGACAGCGGGTC-3); GAPDH (sense primer,
5-GGAGCGAGATCCCTCCAAAAT-3 and antisense primer,
5-GGCTGTTGTCATACTTCTCATGG-3). Quantitative real-time PCR was
performed using Mx3005P qPCR System (Agilent Technologies, Santa
Clara, CA, USA).
Molecular docking and dynamics
simulation
To assess the molecular interactions between CAPE
and HNE, a molecular docking study was carried out with CDOCKER
protocol from Discovery Studio™ 2.5 (DS; Accelrys Software Inc.,
San Diego, CA, USA). For ligand preparation, a 2-D structure of
CAPE was produced by KegDraw software. A force field was applied
and binding energy was minimized before the docking procedure. For
receptor preparation, the X-ray crystal structure of HNE was
obtained from the Protein Data Bank (PDB; http://www.rcsb.org/) with PDB ID: 1B0F. Then, a
ligand-based similarity search scheme was employed and the docking
protocol was performed as a default setting to avoid a potential
reduction in docking accuracy. Molecular dynamic simulations of HNE
and CAPE were performed using the standard dynamics cascade
protocol in DS. The initial structures of those two simulations
were the results of the ligand-receptor complex with the lowest
binding energy. The complex was solvated with water in a cubic box
with an explicit periodic boundary model to stimulate the
environment inside the cell. After creating harmonic restraint, a
standard dynamics cascade was performed including minimization,
heating, equilibration and production dynamics (21).
Surface plasmon resonance
analysis
Surface plasmon resonance (SPR) experiments of CAPE
binding to HNE were carried out on a BIAcore® 3000
(Biacore AB, Uppsala, Sweden) instrument using Sensor Chip CM5 (GE
Healthcare, Fairfield, CT, USA). Pipelines and the chip were
pretreated with phosphate-buffered saline (PBS)-ethyl pyruvate (EP)
running buffer (10 mM HEPES, 150 mM NaCl, 3 mM EDTA, 0.005%
surfactant P20 and deionized water, pH 7.4). For SPR analysis, ~500
resonance units of HNE were immobilized on the CM5 sensor chip
using 10 mM sodium acetate buffer with pH 5.5. Kinetics experiments
were carried out by injecting decreasing concentrations of CAPE
(120 µM with 2-fold dilutions to 7.5 µM) at a flow rate of 10
µl/min, and binding responses were recorded for 300 sec in
succession. Binding responses were recorded continuously with
response unit values. The association (Ka) and
dissociation (Kd) rate constants were elevated by BIA
evaluation software (Biacore AB) (21).
Statistical analysis
Significances of the differences between two groups
were evaluated by a Student's unpaired t-test and analysis of
variance (ANOVA) for comparisons among multiple groups using
GraphPad Prism version 5.01 (GraphPad Software, Inc., La Jolla, CA,
USA). All data are expressed as mean ± standard error of
measurement (SEM).
Results
CAPE inhibits tumor cell viability and
migration induced by HNE in PANC-1 cells
To measure the cell growth inhibition induced by
CAPE in pancreatic cancer cells with HNE in vitro, we
measured cell viability in PANC-1 cells using the CellTiter
96® AQueous One Solution cell proliferation
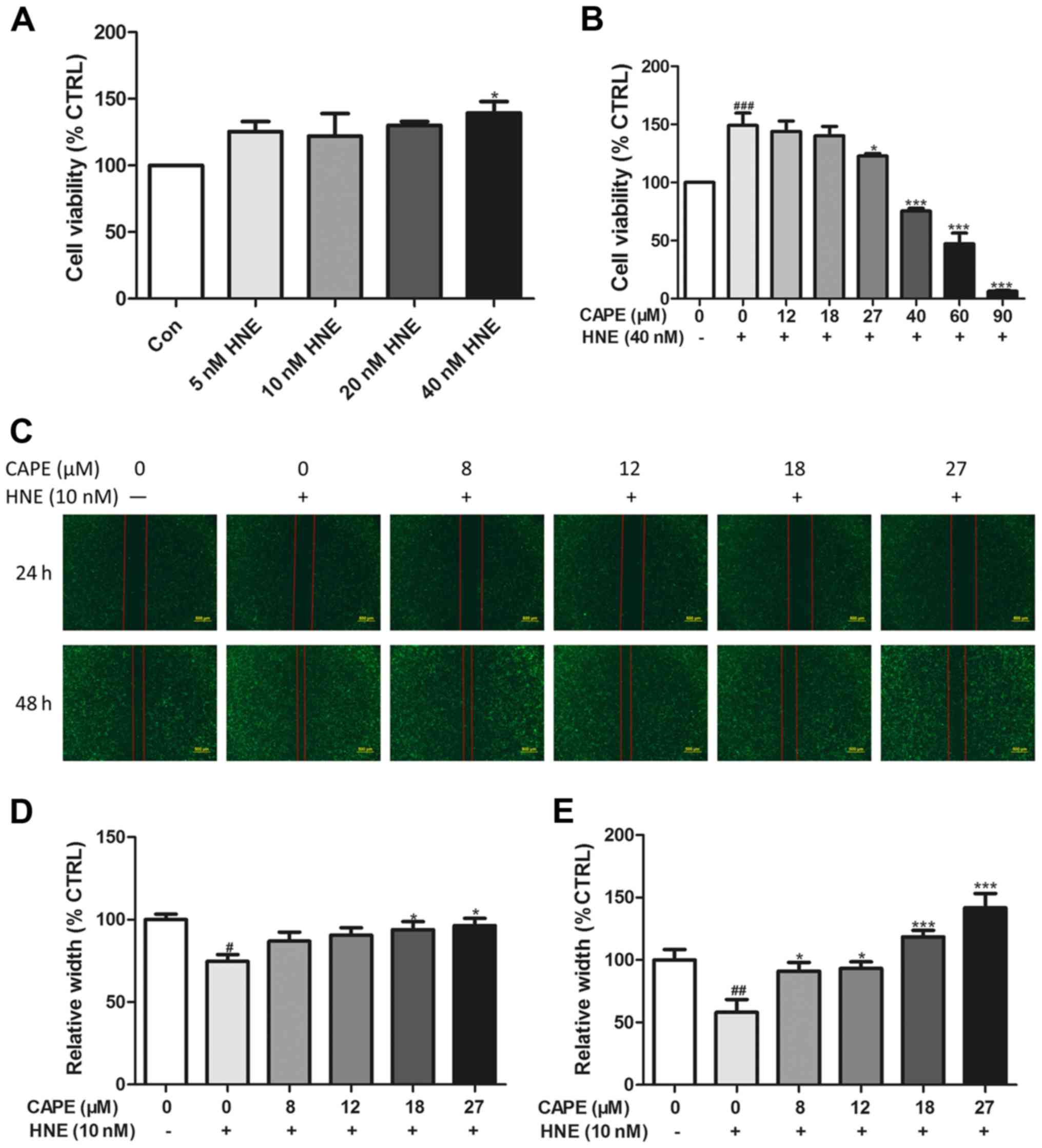
assay kit. As shown in Fig. 1A,
dose-response curves for the PANC-1 cells indicated that modest
concentrations of HNE (10–40 nM) enhanced tumor cell viability, and
that 40 nM HNE was the most efficacious, and thus this was used for
subsequent experiments. In the presence of 40 nM of HNE, cell
viability was significantly decreased with increasing CAPE doses
(27–90 µM), and lower concentrations had no appreciable effect
(Fig. 1B).
To determine whether CAPE inhibits the migration of
PANC-1 cells induced by HNE, migration capacity was assessed using
an in vitro wound-healing assay into 6-well plates. Fig. 1C-E shows that the migration of
PANC-1 cells incubated with 10 nM HNE was increased, whereas
incremental doses of CAPE (8–27 µM) inhibited the migration of
PANC-1 cells. From the cell viability and migration assays, HNE
induced cancer cell migration at lower doses, and increased cancer
cell viability at higher doses, data that agree with previous
research results (22). In
addition, our data showed that lower doses of CAPE inhibited
migration and higher doses of CAPE inhibited cell growth.
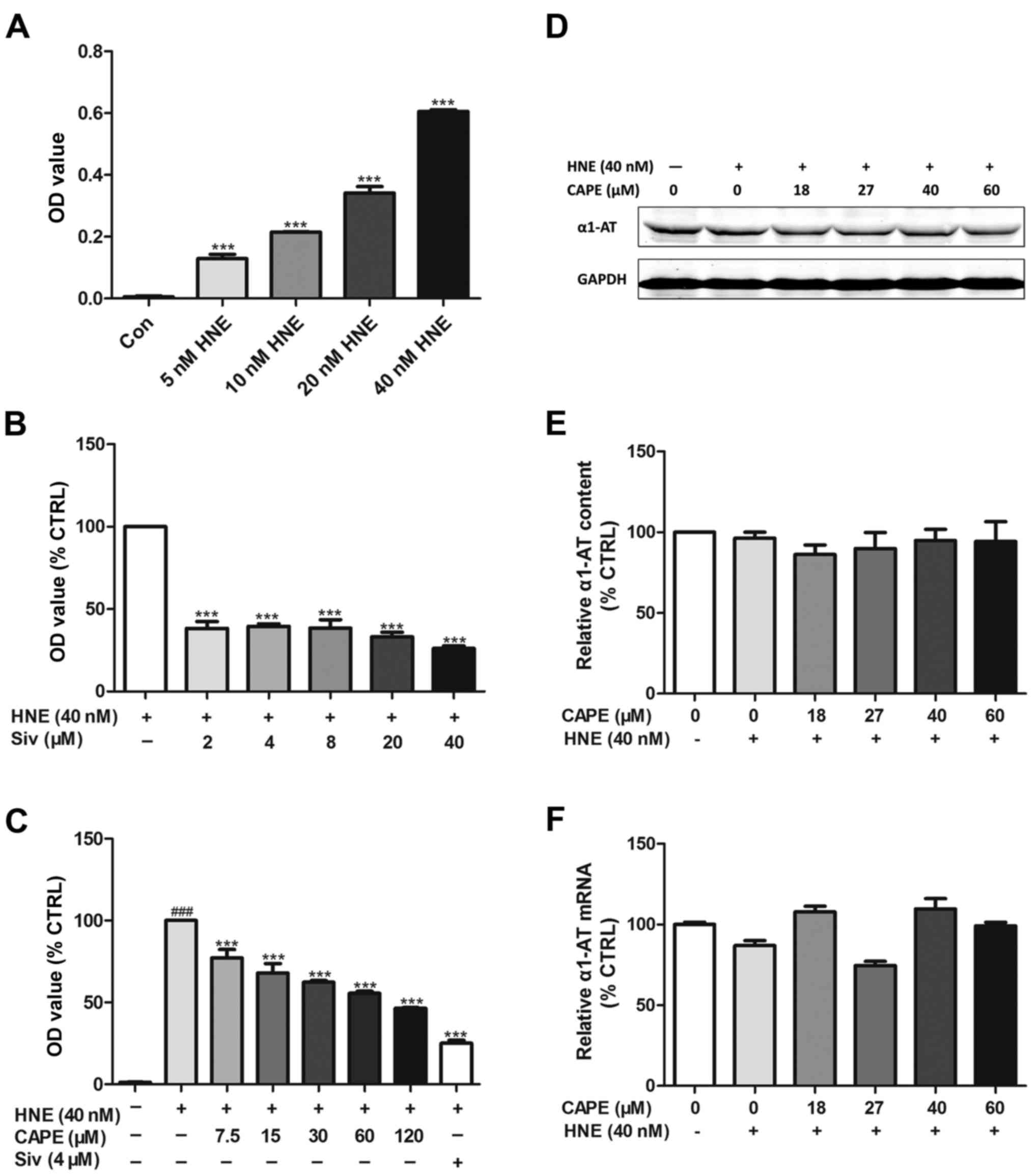
CAPE inhibits HNE activity (elastase
assay) in an α1-antitrypsin-independent manner
To study HNE inhibition using an elastase assay,
sivelestat sodium, a specific HNE inhibitor, was used as a positive
control (Fig. 2B). As shown in
Fig. 2A, HNE activity increased in
a concentration-dependent manner. In addition, Fig. 2C shows that HNE activity was also
significantly inhibited by CAPE in a concentration-dependent manner
(7.5–120 µM).
α1-antitrypsin is a serine protease inhibitor
(SERPIN) and the endogenous inhibitor of HNE. Several natural
products inhibit HNE through the regulation of α1-antitrypsin. To
investigate how HNE-induced growth and migration is inhibited by
CAPE, western blotting (Fig. 2D and
E) and quantitative real-time PCR (Fig. 2F) to determine whether
α1-antitrypsin was regulated by CAPE in PANC-1 cells. As shown in
Fig. 2D-F, with or without HNE,
CAPE (18–60 µM) did not modulate α1-antitrypsin expression at
either the transcriptional or translational level.
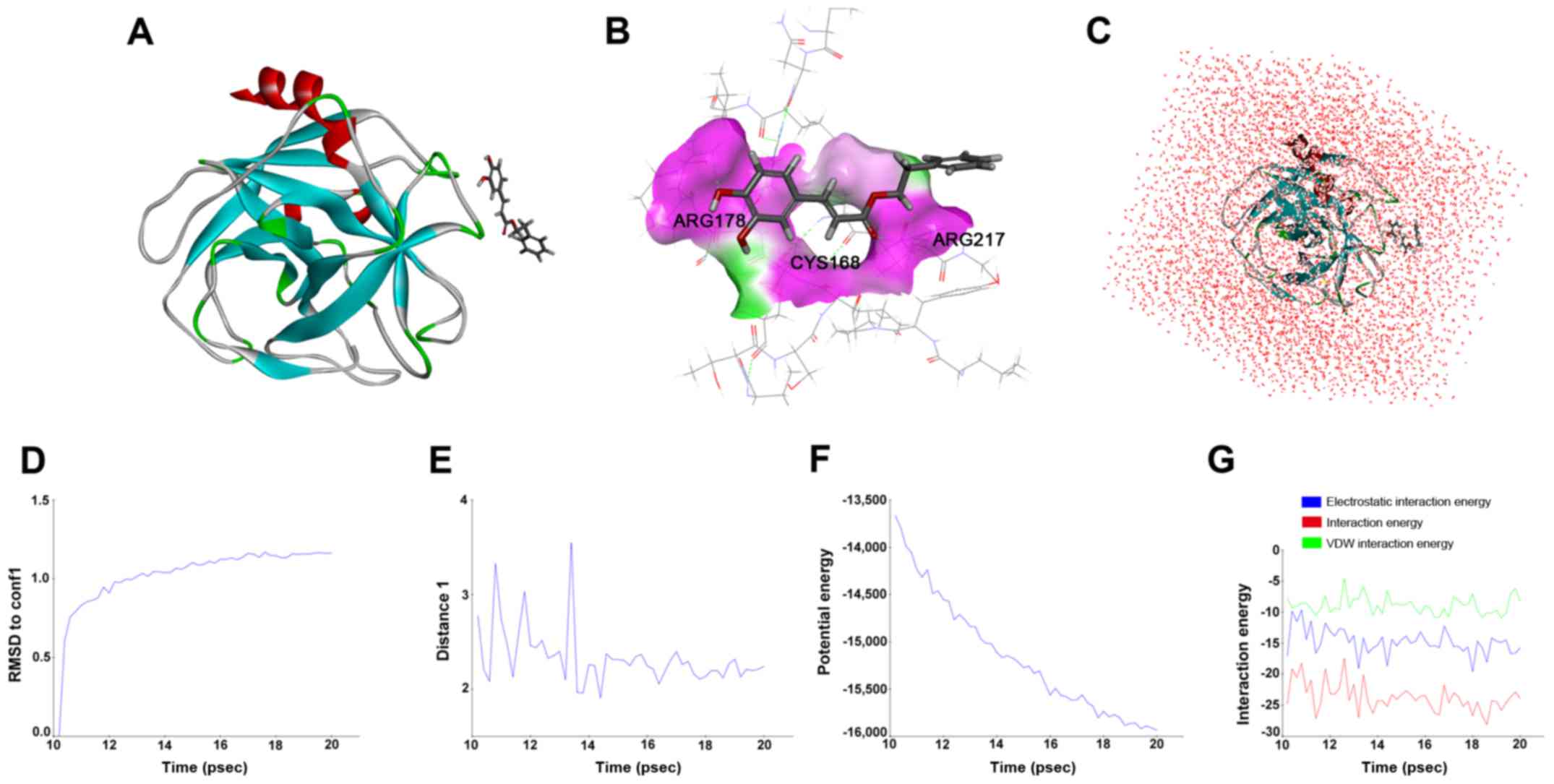
hNE is a direct protein-target for
CAPE in silico
We measured the specific binding of CAPE to HNE
using molecular dynamics. The in silico drug target docking
modeling analysis indicated that CAPE directly bound to the binding
pocket of HNE (Fig. 3A-C) with
25.6581 kcal/mol in its best binding confirmation. As shown in
Fig. 3B, ARG178, CYS168 and ARG217
play decisive roles in H-bond formation, which contributes to
stabilizing the complex of HNE and CAPE. RMSD reference of CAPE,
plotted in Fig. 3D, indicated that
the interaction of the receptor-ligand complex reached equilibrium
state after 14 psec. Similar results were obtained from interaction
analysis between the O of CAPE and HN in the amino residue of
ARG178 in HNE (Fig. 3E-G). Thus,
the residue of the catalytic site stabilizes the interaction
between CAPE and HNE.
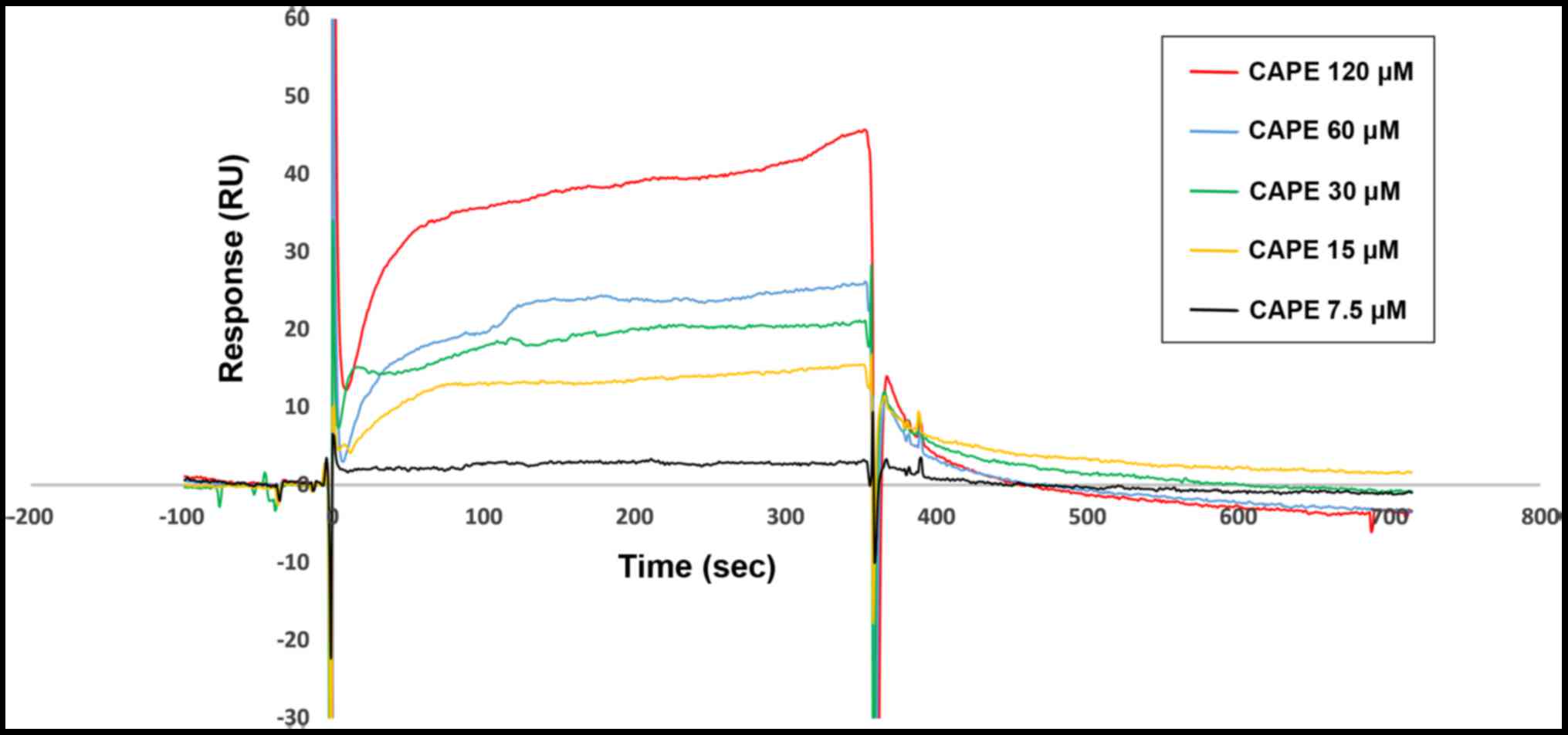
CAPE directly binds to HNE according
to SPR analysis
We determined the binding affinity of CAPE for HNE
based on SPR. Fig. 4 shows that the
response units were significantly increased with increasing CAPE
(7.5–120 µM), indicating that CAPE was able to directly bind to HNE
in a concentration-dependent manner. Ka and
Kd values for CAPE binding to immobilized HNE on the CM5
chip were 1.97×105 M−1 sec−1 and
2.35×10−2 sec−1, respectively. The
equilibrium dissociation constant (KD =
Kd/Ka) was 1.19×10−7 M. Thus, HNE
is a direct target of CAPE.
Discussion
We investigated the anti-migratory and anti-growth
effects of CAPE on HNE-induced PANC-1 cells. Using an elastase
assay, SPR and computer-aided drug design (CADD), we analyzed the
interaction between CAPE and HNE. Data showed that CAPE inhibited
the migration and viability of the HNE-induced PANC-1 cells. In
addition, CAPE solely and directly interacts with HNE.
CAPE, a polyphenolic natural product and active
component of propolis, has been reported to inhibit tumor
proliferation and metastasis (23),
as well as induce the apoptosis of various cancer cells in
vitro and in vivo, including central nervous system,
gastrointestinal system and breast cancer as well as leukemia
(18,24). CAPE has a range of molecular targets
and influences numerous biochemical and molecular processes and it
is documented to be safe for humans. Tumor cell lines are
significantly more sensitive to CAPE than non-cancerous cells
(19). An in vivo study
demonstrated that propolis has little acute oral toxicity
(LD50 2,000–7,300 g/kg for mice) (18). In addition to its antitumor effect,
CAPE also is an anti-inflammatory, antioxidant compound with
neuroprotective and immune modulatory functions (16).
Many molecules are involved in the antitumor
activity of CAPE, including NF-κB, JNK, caspase-3, activator
protein-1 (AP-1) and tumor necrosis factor (TNF). Cancer
progression is a complex process and its pathogenic mechanism
remains somewhat elusive, thus CAPE is considered to be a
multi-target natural compound. That CAPE may inhibit tumor
development via other mechanisms cannot be excluded. Thus, we
investigated the novel inflammation-related mechanisms of CAPE
involved in its antitumor activity in pancreatic cancer.
There is a growing awareness that inflammation is an
important risk factor for the development of cancer at many sites,
such as the lung, bladder, gastrointestinal tract, skin and vulva
(12). When Virchow observed the
presence of inflammatory cells within neoplastic tissue in the 19th
century, the link between inflammation and cancer was first
suggested, but the mechanism underlying this is only recently being
clarified (12,25). Inflammation plays a role in
promoting cancer growth, tissue invasion and metastasis, due to
reactive oxygen species and the induction of cell cycling. Cancer
arising in a background of inflammation may therefore be associated
with a worse prognosis (13).
PDAC is a lethal cancer with a poor prognosis. In
addition, a lack of early symptoms and distant metastatic spread by
the time of diagnosis and the intrinsic and acquired resistance to
conventional therapeutic modalities makes this a difficult cancer
to treat (26). Cellular mechanisms
contributing to pancreatic cancer development and progression are
undefined but inflammation is thought to be key particularly in the
context of repeated acute pancreatic injury. Acute pancreatitis
(AP) is a clinical syndrome which begins with acute injury to the
pancreas and injurious substances include alcohol, smoking, gall
stones, drugs and obesity. Recurrent acute pancreatic insult can
lead to chronic pancreatitis, which is a strong risk factor for
pancreatic cancer (27).
HNE degrades phagocytosed foreign organic molecules
within cells. Extracellular HNE can degrade various extracellular
proteins and induce production of inflammatory cytokines from
epithelial cells (28). Several
studies indicate that HNE concentration and activity are elevated
in AP, and are reliable markers of the severity of AP. Specific HNE
inhibitors, such as sivelestat, have potential for treating AP
(29). Sivelestat is the first drug
applied to systemic inflammatory response syndrome (SIRS) and
sivelestat may suppress breast and esophageal cancer and
cholangiocarcinoma and gastric carcinoma (30–33).
Previously, we reported that curcumin can inhibit
HNE with a similar CDOCKER interaction energy to that of CAPE.
However, the equilibrium dissociation constant (KD =
Kd/Ka) of CAPE binding to HNE
(1.19×10−7 M) was significantly lower than that of
curcumin (3.17×10−5 M) based on SPR. Thus, CAPE could
bind to HNE better than curcumin (21,22).
In addition, many other natural compounds have been screened as HNE
inhibitors, including flavonoids, caffeic acid derivatives,
phenolics, monoterpenes, sesquiterpenes, triterpenes and long-chain
fatty acids (11).
Traditional approaches to drug discovery rely on
screening of numerous compounds to identify a potential candidate,
which is an expensive and time consuming process. CADD approaches
have been widely employed to identify lead compounds and optimize
drug development against various targets (34). CADD uses rational drug design
methods, but biological studies are necessary to verify CADD data.
SPR is ideal for generating reliable data concerning interactions
between biomolecules and small ligands with real-time analysis and
detection of low-affinity target binders (35). CADD and SPR provide a fast,
inexpensive, and reliable method of drug discovery.
In conclusion, CAPE modulates tumor migration and
growth induced by HNE and CAPE blocks HNE activity directly
interacting with HNE. These data offer a new putative mechanism of
CAPE activity against tumor migration and growth and suggest that
CAPE may be a potential anticancer drug.
Acknowledgements
The present study was supported by the National
Natural Science Foundation of China (nos. 81473235, 81673453,
91129727, 81673486, 81270049 and 81373405), and the Research Fund
from the Ministry of Education of China (111 Projects no.
B07001).
References
|
1
|
Paulson AS, Cao Tran HS, Tempero MA and
Lowy AM: Therapeutic advances in pancreatic cancer.
Gastroenterology. 144:1316–1326. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel R, Naishadham D and Jemal A: Cancer
statistics, 2012. CA Cancer J Clin. 62:10–29. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Siegel R, Naishadham D and Jemal A: Cancer
statistics, 2013. CA Cancer J Clin. 63:11–30. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Rahib L, Smith BD, Aizenberg R, Rosenzweig
AB, Fleshman JM and Matrisian LM: Projecting cancer incidence and
deaths to 2030: The unexpected burden of thyroid, liver, and
pancreas cancers in the United States. Cancer Res. 74:2913–2921.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Reid MD, Basturk O, Thirabanjasak D,
Hruban RH, Klimstra DS, Bagci P, Altinel D and Adsay V:
Tumor-infiltrating neutrophils in pancreatic neoplasia. Mod Pathol.
24:1612–1619. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Gaida MM, Steffen TG, Günther F,
Tschaharganeh DF, Felix K, Bergmann F, Schirmacher P and Hänsch GM:
Polymorphonuclear neutrophils promote dyshesion of tumor cells and
elastase-mediated degradation of E-cadherin in pancreatic tumors.
Eur J Immunol. 42:3369–3380. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Sköld CM, Liu X, Umino T, Zhu Y, Ohkuni Y,
Romberger DJ, Spurzem JR, Heires AJ and Rennard SI: Human
neutrophil elastase augments fibroblast-mediated contraction of
released collagen gels. Am J Respir Crit Care Med. 159:1138–1146.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kouakou-Siransy G, Sahpaz S, Nguessan GI,
Datté JY, Brou JK, Gressier B and Bailleul F: Effects of
Alchornea cordifolia on elastase and superoxide anion
produced by human neutrophils. Pharm Biol. 48:128–133. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Sato T, Takahashi S, Mizumoto T, Harao M,
Akizuki M, Takasugi M, Fukutomi T and Yamashita J: Neutrophil
elastase and cancer. Surg Oncol. 15:217–222. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lucas SD, Costa E, Guedes RC and Moreira
R: Targeting COPD: Advances on low-molecular-weight inhibitors of
human neutrophil elastase. Med Res Rev. 33:(Suppl 1). E73–E101.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Siedle B, Hrenn A and Merfort I: Natural
compounds as inhibitors of human neutrophil elastase. Planta Med.
73:401–420. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
McKay CJ, Glen P and McMillan DC: Chronic
inflammation and pancreatic cancer. Best Pract Res Clin
Gastroenterol. 22:65–73. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Greer JB and Whitcomb DC: Inflammation and
pancreatic cancer: An evidence-based review. Curr Opin Pharmacol.
9:411–418. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Sun Z and Yang P: Role of imbalance
between neutrophil elastase and alpha 1-antitrypsin in cancer
development and progression. Lancet Oncol. 5:182–190. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kamohara H, Sakamoto K, Mita S, An XY and
Ogawa M: Neutrophil elastase inhibitor (ONO-5046.Na) suppresses the
proliferation, motility and chemotaxis of a pancreatic carcinoma
cell line, Capan-1. Res Commun Mol Pathol Pharmacol. 98:103–108.
1997.PubMed/NCBI
|
|
16
|
Tolba MF, Azab SS, Khalifa AE,
Abdel-Rahman SZ and Abdel-Naim AB: Caffeic acid phenethyl ester, a
promising component of propolis with a plethora of biological
activities: A review on its anti-inflammatory, neuroprotective,
hepatoprotective, and cardioprotective effects. IUBMB Life.
65:699–709. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chan GC, Cheung KW and Sze DMP: The
immunomodulatory and anticancer properties of propolis. Clin Rev
Allergy Immunol. 44:262–273. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Akyol S, Ozturk G, Ginis Z, Armutcu F,
Yigitoglu MR and Akyol O: In vivo and in vitro antineoplastic
actions of caffeic acid phenethyl ester (CAPE): Therapeutic
perspectives. Nutr Cancer. 65:515–526. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Watanabe MA, Amarante MK, Conti BJ and
Sforcin JM: Cytotoxic constituents of propolis inducing anticancer
effects: A review. J Pharm Pharmacol. 63:1378–1386. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Melzig MF, Pertz HH and Krenn L:
Anti-inflammatory and spasmolytic activity of extracts from
Droserae herba. Phytomedicine. 8:225–229. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Fan S, Xu Y and Li X, Tie L, Pan Y and Li
X: Opposite angiogenic outcome of curcumin against ischemia and
Lewis lung cancer models: In silico, in vitro and in vivo studies.
Biochim Biophys Acta. 1842:1742–1754. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Xu Y, Zhang J, Han J, Pan X, Cao Y, Guo H,
Pan Y, An Y and Li X: Curcumin inhibits tumor proliferation induced
by neutrophil elastase through the upregulation of α1-antitrypsin
in lung cancer. Mol Oncol. 6:405–417. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chen MJ, Shih SC, Wang HY, Lin CC, Liu CY,
Wang TE, Chu CH and Chen YJ: Caffeic Acid phenethyl ester inhibits
epithelial-mesenchymal transition of human pancreatic cancer cells.
Evid Based Complement Alternat Med. 2013:2709062013.PubMed/NCBI
|
|
24
|
Chen MJ, Chang WH, Lin CC, Liu CY, Wang
TE, Chu CH, Shih SC and Chen YJ: Caffeic acid phenethyl ester
induces apoptosis of human pancreatic cancer cells involving
caspase and mitochondrial dysfunction. Pancreatology. 8:566–576.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Hausmann S, Kong B, Michalski C, Erkan M
and Friess H: The role of inflammation in pancreatic cancer. Adv
Exp Med Biol. 816:129–151. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Momi N, Kaur S, Krishn SR and Batra SK:
Discovering the route from inflammation to pancreatic cancer.
Minerva Gastroenterol Dietol. 58:283–297. 2012.PubMed/NCBI
|
|
27
|
Kolodecik T, Shugrue C, Ashat M and
Thrower EC: Risk factors for pancreatic cancer: Underlying
mechanisms and potential targets. Front Physiol. 4:4152014.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Aikawa N and Kawasaki Y: Clinical utility
of the neutrophil elastase inhibitor sivelestat for the treatment
of acute respiratory distress syndrome. Ther Clin Risk Manag.
10:621–629. 2014.PubMed/NCBI
|
|
29
|
Novovic S, Andersen AM, Nord M, Astrand M,
Ottosson T, Jørgensen LN and Hansen MB: Activity of neutrophil
elastase reflects the progression of acute pancreatitis. Scand J
Clin Lab Invest. 73:485–493. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Nawa M, Osada S, Morimitsu K, Nonaka K,
Futamura M, Kawaguchi Y and Yoshida K: Growth effect of neutrophil
elastase on breast cancer: Favorable action of sivelestat and
application to anti-HER2 therapy. Anticancer Res. 32:13–19.
2012.PubMed/NCBI
|
|
31
|
Nishiyama J, Matsuda M, Ando S, Hirasawa
M, Suzuki T and Makuuchi H: The effects of the early administration
of sivelestat sodium, a selective neutrophil elastase inhibitor, on
the postoperative course after radical surgery for esophageal
cancer. Surg Today. 42:659–665. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Hanada N, Kusano S, Hori K and Momi H:
Good response in ARDS treated with sivelestat sodium hydrate during
chemotherapy for cholangiocarcinoma. Gan To Kagaku Ryoho.
34:1303–1306. 2007.(In Japanese). PubMed/NCBI
|
|
33
|
Wada Y, Yoshida K, Hihara J, Konishi K,
Tanabe K, Ukon K, Taomoto J, Suzuki T and Mizuiri H: Sivelestat, a
specific neutrophil elastase inhibitor, suppresses the growth of
gastric carcinoma cells by preventing the release of transforming
growth factor-alpha. Cancer Sci. 97:1037–1043. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Aparoy P, Reddy KK and Reddanna P:
Structure and ligand based drug design strategies in the
development of novel 5-LOX inhibitors. Curr Med Chem. 19:3763–3778.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Crauste C, Willand N, Villemagne B, Flipo
M, Willery E, Carette X, Dimala MM, Drucbert AS, Danze PM, Deprez
B, et al: Unconventional surface plasmon resonance signals reveal
quantitative inhibition of transcriptional repressor EthR by
synthetic ligands. Anal Biochem. 452:54–66. 2014. View Article : Google Scholar : PubMed/NCBI
|