Introduction
Chondrosarcoma, as the second most frequently
occurring primary malignant bone tumor, is characterized by the
secretion of a cartilage-like extracellular matrix. This tumor
usually appears on the pelvis, scapula, or ribs, and presents
diverse clinicopathological behaviors based on the degree of
differentiation. Because of the abundant cartilage-like matrix,
chondrosarcoma exhibits resistance to radiotherapy and
chemotherapy, and surgical resection remains the most effective
approach in chondrosarcoma treatment (1). Thus, to improve the prognosis of
chondrosarcoma, it is of great value and urgency to clarify the
potential pathogenesis mechanism and to develop novel
therapies.
Primary cilia, as microtubule-based organelles, are
found in most eukaryotic cell membranes, and function as ‘antennae’
to detect multifarious stimulation in the extracellular
environment, such as mechanical stress and biochemical signaling
changes (2,3). They have been proved as a vital part
of the control center to regulate intracellular signaling
transduction such as Wnt, Hedgehog, platelet-derived growth factor,
and mammalian target or rapamycin pathway (4). Recently, many studies demonstrated a
potential relationship between primary cilia disassembly and
tumorigenesis. Cilia deficiency has shown a close link with breast
cancer, gastrointestinal tumor, basal cell carcinoma, renal
carcinoma, and rhabdomyosarcoma (5,6).
Besides, de Andrea et al revealed deficient ciliogenesis in
the early period of chondrosarcomagenesis (7). However, the exact mechanism leading to
deciliation in chondrosarcomagenesis and effects of cilia
restoration in chondrosarcoma development remain poorly
understood.
Primary cilia assembly is generally initiated at the
G1/G0 phase of the cell cycle, and the centriole transforms into a
basal body and ciliary axoneme (8,9).
Ciliogenesis and elongation processes require the coordination of
microtubule assembly and protein modification (10,11).
HDAC6, as a special member of the HDAC family, unlike others
interacting with histones, mainly deacetylates microtubule, heat
shock protein 90, cortactin, and redox-related regulatory proteins
(12). Recent studies demonstrated
that overexpression of HDAC6 also correlated with tumorigenesis;
for example, HDAC6 can promote breast cancer cell metastasis by
upregulating cell mobility (13).
Besides, inhibition of HDAC6 in multiple medulloblastoma and
cholangiocarcinoma contribute to induction of apoptosis (14,15).
HDAC6 can serve as a biomarker to evaluate tumor prognosis. In this
study, Tubastatin A, a selective HDAC6 inhibitor, was used to treat
tumor cells. Tubastatin A was reported substantially more selective
(over 1000-fold selectivity) than other isozymes except HDAC8
(57-fold selectivity) (16).
This study found that abnormal expression of HDAC6
existed in human chondrosarcoma tissues, and targeting inhibition
of HDAC6 could significantly suppress chondrosarcoma cell
proliferation and invasion. The potential mechanism may affect
ciliogenesis via Aurora A-HDAC6 cascade. This study aimed to
illustrate a feasible therapeutic method for chondrosarcoma by
regulating primary cilia assembly through inhibiting HDAC6
activation.
Materials and methods
This study was approved by the ethics committee of
the Tongji Hospital, Tongji Medical College, Huazhong University of
Science and Technology, Wuhan, China.
Cells and reagents
Human chondrosarcoma cell line SW1353 and
osteosarcoma cell line MG63 were purchased from the Type Culture
Collection of the Chinese Academy of Sciences (Shanghai, China).
Cells were cultured in Dulbecco's modified Eagle's medium/nutrient
mixture F-12 containing 10% fetal bovine serum and 100 U/ml
penicillin-streptomycin antibiotic solution. HDAC6 inhibitor
Tubastatin A, and Aurora A kinase activator anacardic acid and
inhibitor Danuscrtib (PHA-739358) were all purchased from
Selleckchem (Houston, TX, USA).
Cell proliferation test
Cell Counting Kit-8 (CCK-8, Boster, Wuhan, China)
was used to evaluate cell proliferation capacity, according to
manufacturer's instructions: 2000 cells per well were plated in
96-well plates. The cells were then cultured in a 100 µl culture
medium with different concentrations of Tubastatin A for 12–48 h.
Then, 10 µl CCK-8 was added and the optical density value
(absorbance at 450 nm wavelength) was measured using an enzyme
microplate reader.
Transwell invasion assay
The Transwell assay was used to assess the invasion
capacity of chondrosarcoma cells. The upper filter membrane was
coated with 25 mg Matrigel (BD Biosciences, San Jose, CA, USA) at
37°C for 30 min. The lower chambers were filled with a culture
medium containing 10% fetal bovine serum. SW1353 cells were first
transferred to 100 nM siRNA or negative control siRNA for 48 h and
then starved in a serum-free medium for 4 h. After digestion,
1×105 cells were transferred onto the upper membrane and
stimulated with 10 µM Tubastatin A. After 24 h, the Matrigel was
gently wiped from the upper surface, and the cells were stained
with crystal violet (Boster). Cells in 10 random visual fields were
counted under a microscope (Olympus, Tokyo, Japan).
Immunohistochemical studies
Human chondrosarcoma samples were fixed in 4%
paraformaldehyde, then embedded in paraffin and sectioned for
immunohistochemical assays. The experimental processes were
conducted using standard techniques based on previous studies
(17). Primary antibodies HDAC6
(1:100 dilution; Beyotime, Shanghai, China) and IFT88 (1:100
dilution; Abgent, San Diego, CA, USA) were stained, and Ki67 was
stained as a marker for tumor proliferation (1:200 dilution; Cell
Signaling Technology, Danvers, MA, USA). These sections were
observed under a microscopic magnification of ×200.
Immunofluorescence assay
Chondrosarcoma SW1353 and osteosarcoma MG63 cells of
the proper density were inoculated on cover glasses and stimulated
with different reagents. On the basis of immunofluorescence
standard processes, primary cilia were confirmed using primary
anti-acetylated α-tubulin antibody (1:300 dilution; Abcam,
Cambridge, UK) and double stained with cilia IFT88 (1:100 dilution;
Abgent). CY3-conjugated goat anti-mouse and FITC-conjugated goat
anti-rabbit immunoglobulin G (IgG) were used as secondary antibody
at a dilution of 1:100, and nuclear staining with 1 µg/µl
4,6-diamidino-2-phenylindole (DAPI). Images were visualized and
recorded with a fluorescent microscope.
Western blot analysis
According to manufacturer's instructions, total cell
proteins (20 µg/lane) were loaded and separated using sodium
dodecyl sulfate-polyacrylamide gels and transferred onto
polyvinylidene difluoride membranes. Next, they were incubated with
the following primary antibodies: acetylated α-tubulin (1:1000
dilution; Abcam), IFT88 (1:300 dilution; Abgent), Cyclin D1 (1:100
dilution; Boster), PCNA (1:1000 dilution; Cell Signaling
Technology), Aurora A (1:1000 dilution; Beyotime), phosphorylate
Aurora A (1:1000 dilution; Beyotime), and GAPDH (1:500 dilution;
Boster). Then they were incubated with secondary antibody
horseradish peroxidase-labeled goat anti-rabbit and goat anti-mouse
IgG (1:5000 dilution; Boster). All the protein bands were
visualized using an enhanced chemiluminescence system (Bio-Rad,
Philadelphia, PA, USA).
siRNA transfection
SW1353 chondrosarcoma cells and MG63 osteosarcoma
cells were transfected with 100 nM siRNA targeting HDAC6 or with a
scrambled sequence (negative control siRNA) according to the
manufacturer's instructions (siRNA were synthesized by the Ribobio
Co., Ltd., Guangzhou, China). The efficiency of the knockdown
specific gene was evaluated by western blot assays.
Xenograft experiment
SW1353 cells (1×107) were harvested and
suspended in 100 µl culture medium, and subcutaneously inoculated
in 4 weeks old female nude mice. Compound treatment was given after
tumor cell inoculation for 1 week. Twelve mice were randomly
divided into two groups and, respectively, treated with Tubastatin
A (40 mg/kg) or equivalent solvent (DMSO dissolved in saline). Ten
days later, tumors were removed and the weights and volumes were
measured.
Statistical analysis
Mean values and 95% confidence intervals were
calculated, and data are presented as mean ± standard deviation.
The Student's t-test or one-way analysis of variance was used to
analyze the differences. All statistical analyses were performed
using SPSS 20.0 software (IBM, Armonk, NY, USA) and a P-value
<0.05 was defined as statistically significant.
Results
Highly expressed HDAC6 exists in human
chondrosarcoma tissues
HDAC6 is a crucial enzyme in regulating protein
deacetylation and gene expression in epigenetic modification.
Intraflagellar transport (IFT) is a cilia-related motor-driven
protein mainly transporting signaling factors from the top of
axoneme to the basal body (18).
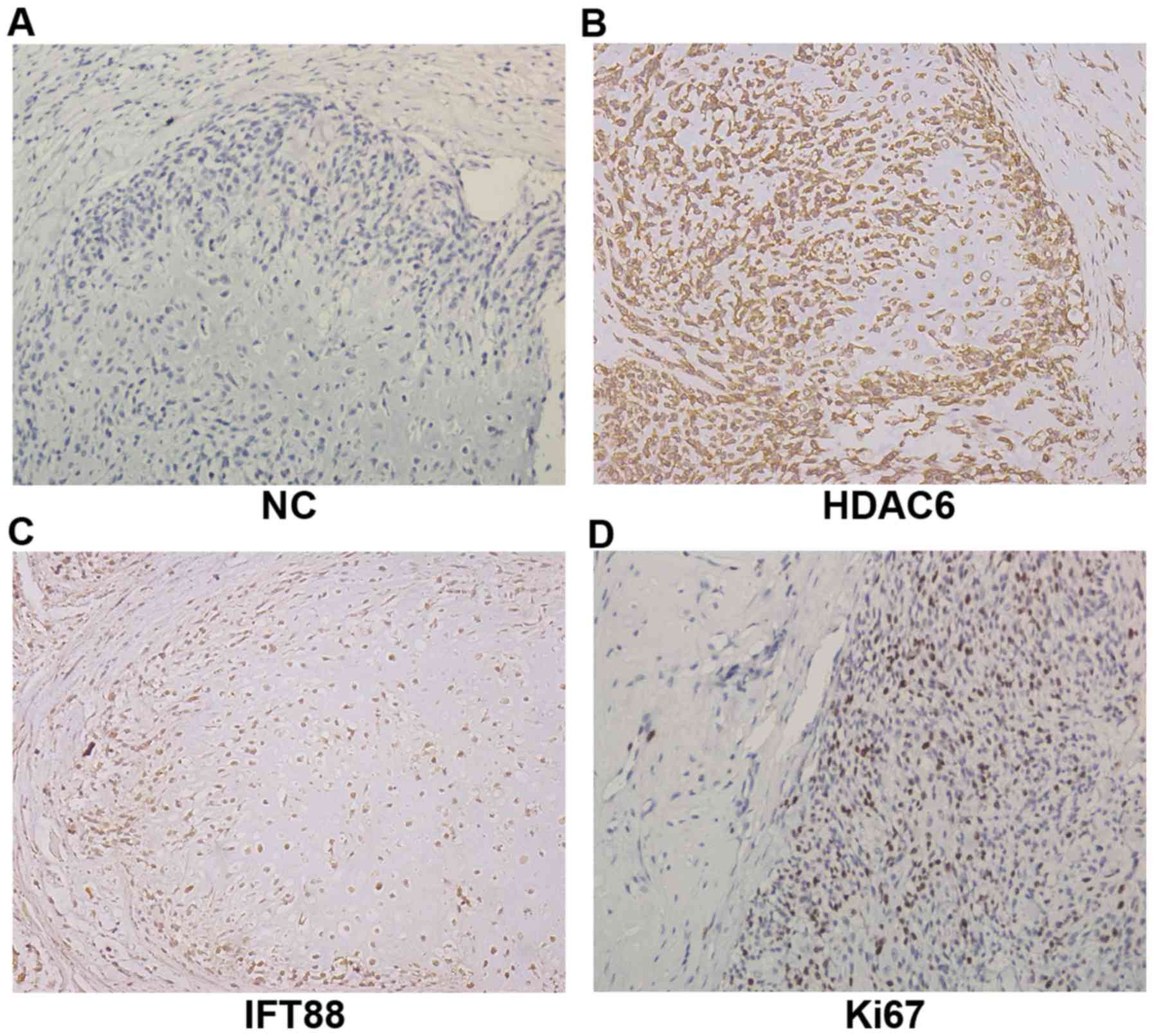
Immunohistochemical results are shown in Fig. 1. Compared with the negative control
(Fig. 1A), the obvious expression
of HDAC6 was found in chondrosarcoma tissues (Fig. 1B), with the relatively lower
expression of IFT88 (Fig. 1C). At
the same time, the definite proportion expression of Ki67 showed
the proliferation capacity of chondrosarcoma cells and reflected
the degree of malignancy (Fig. 1D).
These results suggested that an abnormal activation of
deacetylation modification may exist in human chondrosarcoma, and
expression of HDAC6 may regulate tumor malignant biological
properties but show a negative correlation with cilium-specific
protein IFT88.
HDAC6 inhibition restrains
chondrosarcoma cell proliferation
To elucidate the effects of HDAC6 inhibition in
chondrosarcoma cell proliferation, HDAC6-specific inhibitor
Tubastatin A was used to stimulate chondrosarcoma cell line SW1353.
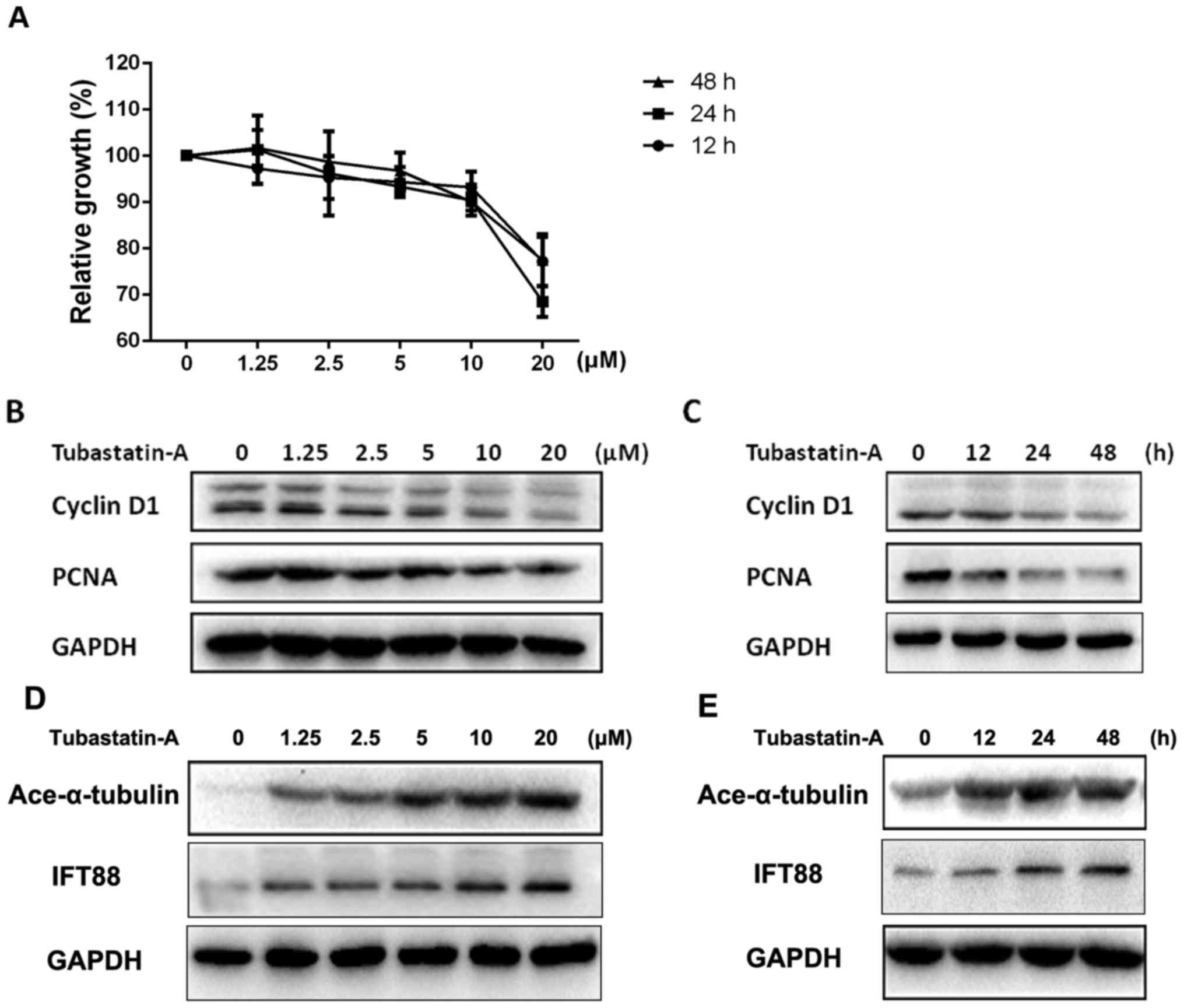
As shown in Fig. 2, CCK-8 and
western blot assays indicate that Tubastatin A could inhibit the
proliferation capacity of SW1353 cells in a concentration- and
time-dependent manner. The inhibitory effects increased gradually
with increasing concentrations of this drug. The tumor cell
proliferation speed slowed down with time (Fig. 2A-C). However, the cell proliferation
speed and related cyclin D1 and PCNA proteins tend to be stable
when stimulated after 24 h (Fig.
2C). Moreover, the expression of cilia-related protein
acetylated α-tubulin and IFT88 was increased as concentration
increased or time passed, and showed a negative tendency with the
proliferation capacity (Fig. 2D and
E). These results revealed that the inhibition of HDAC6
correlated with a decrease in chondrosarcoma cell proliferation but
could upregulate primary cilia-related protein expression.
HDAC6 inhibition suppresses the
invasion capacity of chondrosarcoma cells
The invasion capacity reveals the malignant degree
of tumor cells and stands for aggressive destruction and
microenvironment adaptation. As chondrosarcoma often appears in
bone tissues, it results in osteolytic destruction and distant
metastasis (1). In this study,
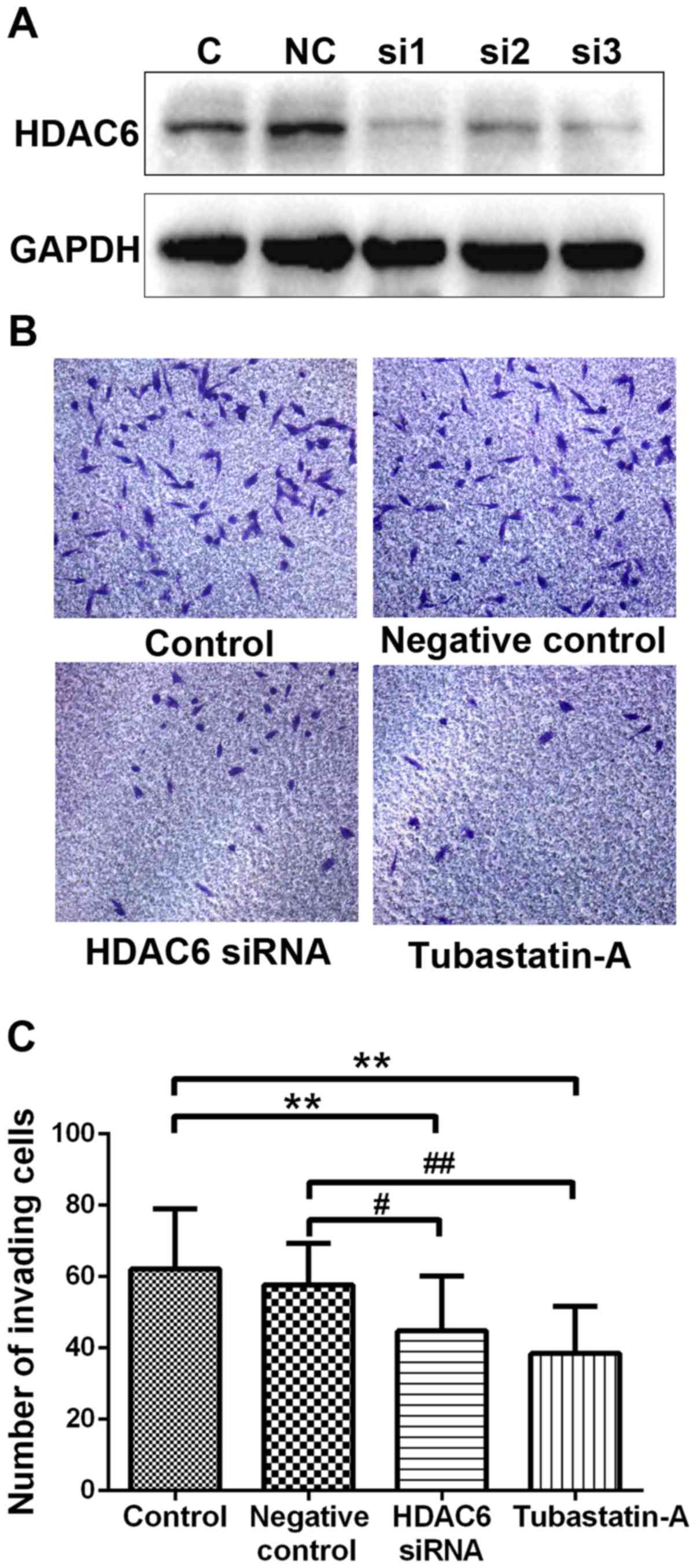
HDAC6 small interfering RNA (siRNA) and inhibitor were utilized to
assess the invasion capacity of chondrosarcoma cells. The results
showed that siRNA could effectively downregulate the endogenous
expression of HDAC6 (Fig. 3A).
Using 10 µM Tubastatin A or transferring HDAC6 siRNA could suppress
the invasion capacity of SW1353 cells compared with the control or
negative control groups (Fig. 3B).
The calculation of invading cells of different groups revealed that
the HDAC6 siRNA and Tubastatin A groups both had significant
differences with the control or negative control groups (Fig. 3C). This conclusion confirmed that
inhibiting HDAC6 associated with the invasion capacity of tumor
cells, and HDAC6 might be a vital regulator to induce osteolytic
destruction and metastasis.
HDAC6 inhibition promotes ciliogenesis
by chondrosarcoma cells
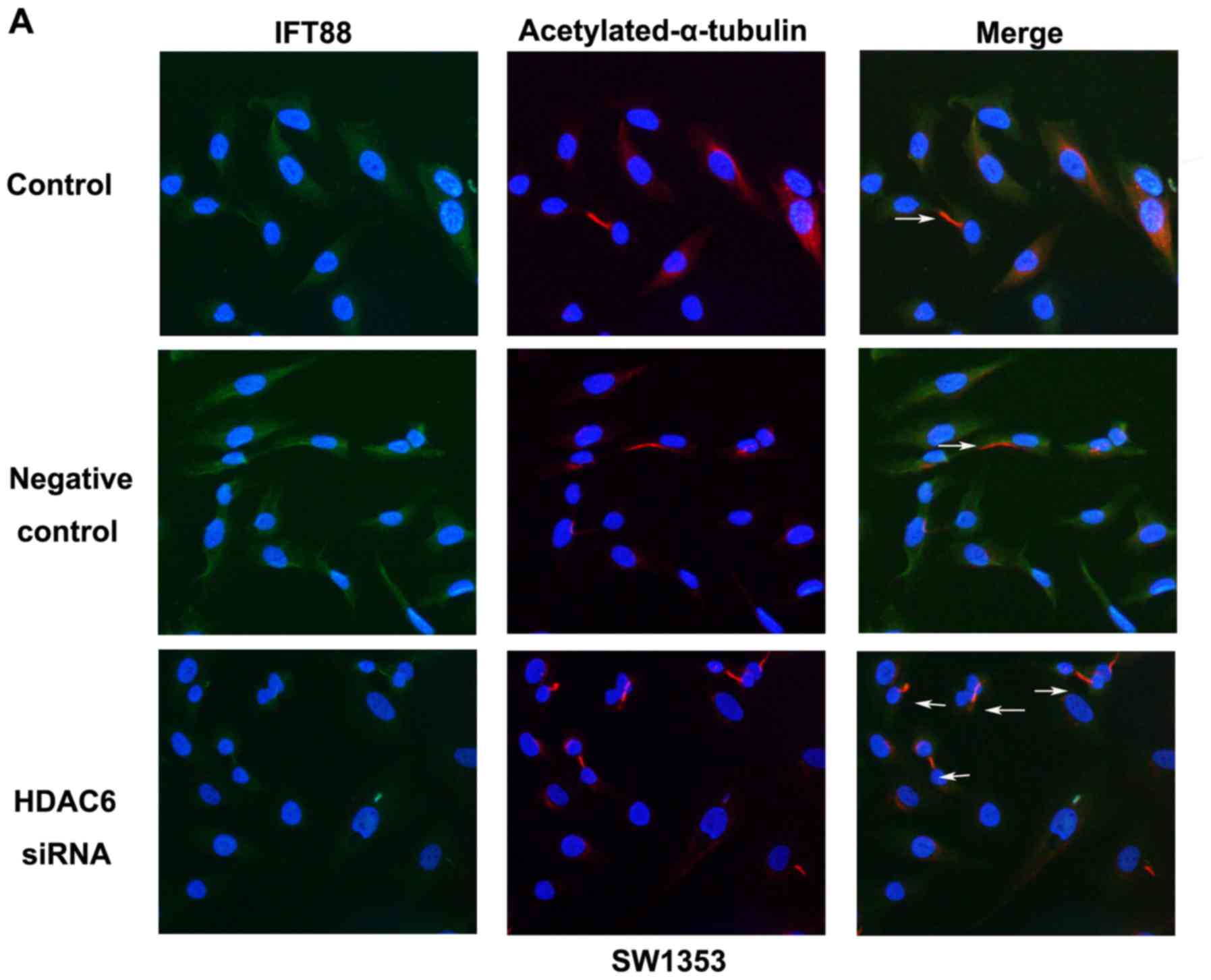
To investigate the function of endogenous HDAC6 in
regulating primary cilia assembly, the study used
immunofluorescence staining assays to detect cilia-related
structural protein acetylated α-tubulin and functional protein
IFT88 in SW1353 and calculated the proportion of ciliary cells. The
results showed that compared with the control or negative control
group, primary cilia could partly be induced by HDAC6 siRNA
(Fig. 4A). On amplifying part of an
image (Fig. 4A, HDAC6 siRNA), IFT88
was found to be widely spread in the cytoplasm but overexpressed
along axoneme acetylated α-tubulin, that is, primary cilia
(Fig. 4C). The frequency of ciliary
cells was 13.28±1.57% in the control group, 15.56±3.86% in the
negative control group, and 20.5±4.81% in the HDAC6 siRNA group
(Fig. 4D). These effects were also
observed in osteosarcoma cell line MG63 (Fig. 4B and E). The results revealed that
the chondrosarcoma cell line SW1353 showed a low level of primary
cilia, and knockdown of endogenous HDAC6 moderately increased cilia
restoration. The expression of IFT88 confirmed a tight correlation
with cilia assembly, and might be regulated by HDAC6.
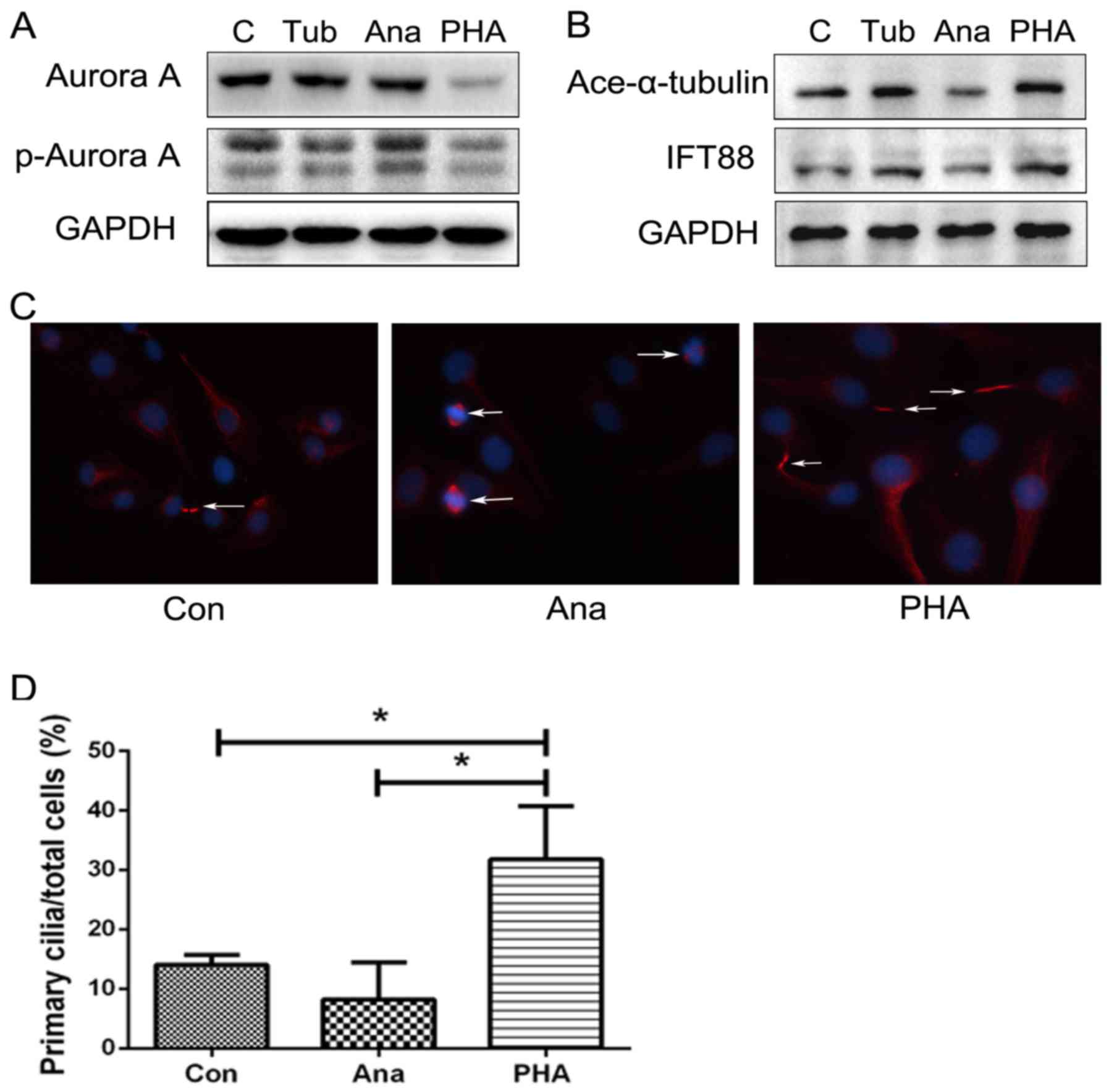
Aurora A-HDAC6 cascade regulates cilia
assembly in chondrosarcoma cells
Aurora A kinase is a centrosomal kinase that
regulates mitotic spindle organization through activation of cyclin
families and can effect HDAC6 activation through phosphorylation
modification (19). To clarify the
role of Aurora A-HDAC6 cascade in chondrosarcoma, HDAC6 inhibitor
Tubastatin A, and Aurora A activator anacardic acid and inhibitor
PHA-739358 were used to stimulate SW1353 cells, and cilia-related
proteins acetylated α-tubulin and IFT88 were evaluated. Results
revealed that the Aurora A activator upregulated phosphorylated
Aurora A, whereas with cilia-related proteins acetylated α-tubulin
and IFT88 decreased, and the inhibitor Danuscrtib showed a negative
tendency. However, HDAC6 inhibitor Tubastatin A had no obvious
effect on the upstream expression of Aurora A, but reduced
phosphorylated Aurora A to a certain extent (Fig. 5A and B), thus confirming that both
inhibitors could not only suppress Aurora A-HDAC6 cascade
activation but also effectively upregulated the primary
cilia-related protein modification of α-tubulin acetylation and
expression of IFT88. The Aurora A-HDAC6 cascade might play an
important role in regulating primary cilia restoration and
affecting cilia function. Immunofluorescence staining assays also
confirmed that Aurora A activation could be accompanied by lower
expression of primary cilia, but showed a negative tendency with
Aurora A inhibitor treatment. Compared to DMSO control group
(14.06±1.62%), Aurora A activator could deacetylate axoneme
α-tubulin to induce cilia disassembly (8.22±7.27%) and lead cells
into a cell cycle, while low doses of Aurora A inhibitor could
promote primary cilia assembly (31.8±8.91%).
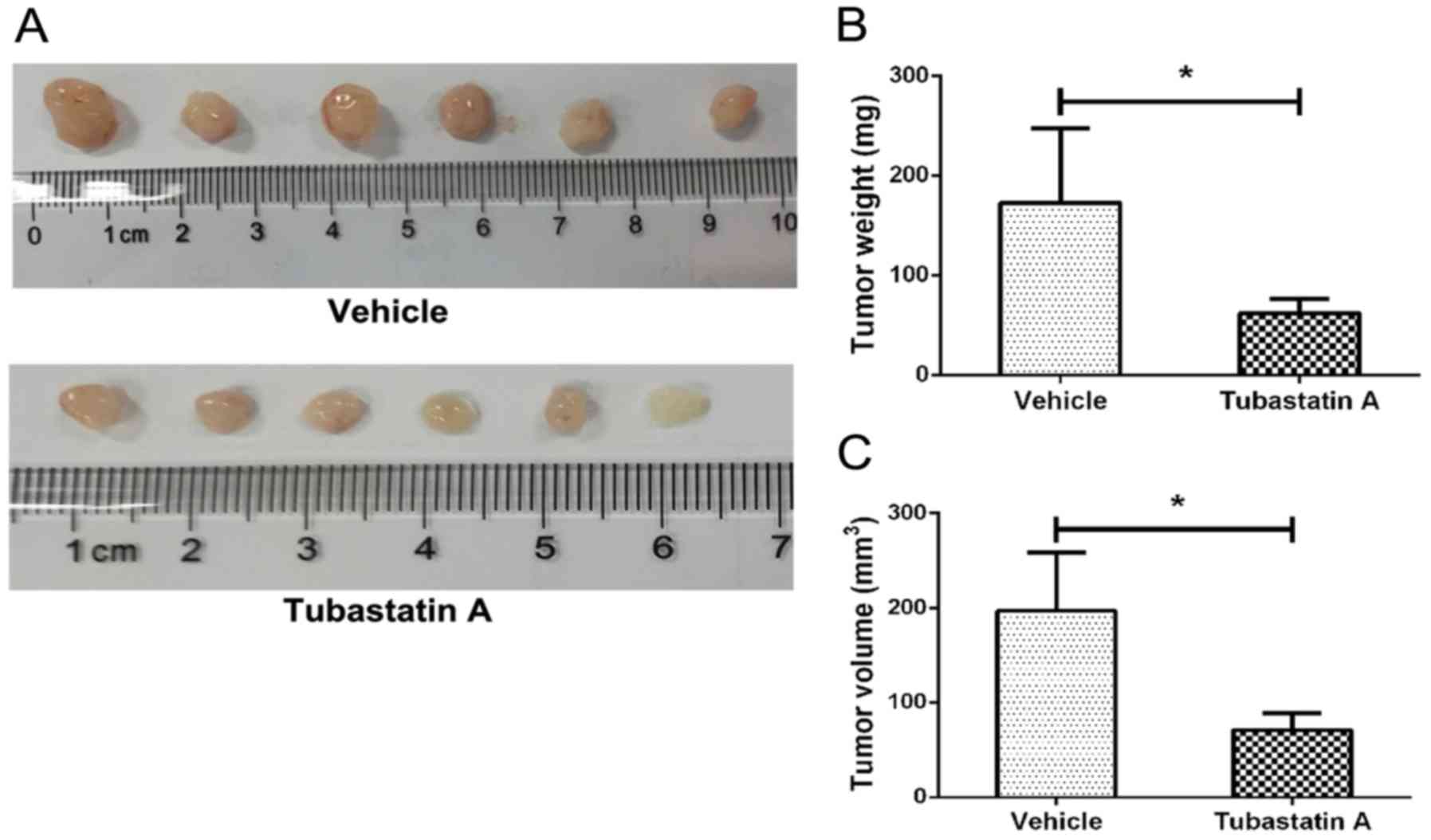
HDAC6 inhibitor suppresses
chondrosarcoma cell growth in vivo
To evaluate the potential clinical application of
HDAC6 inhibitor Tubastatin A, we injected chondrosarcoma SW1353
cells subcutaneously in athymic nude mice. After a 10-day treatment
with Tubastatin A or solvent, tumors were removed and the sizes of
tumors were compared. As the results showed (Fig. 6), compared with control group
(196.67±61.46 mm3, 172.500±62.167 mg), the average
volume of Tubastatin A treatment group was 70.83±17.94
mm3 and the average weight was 74.982±14.634 mg. There
was a significant difference between control group and Tubastatin A
treatment group revealing that HDAC6 inhibitor Tubastatin A could
inhibit chondrosarcoma cells growth in vivo.
Discussion
This study revealed the role of HDAC6 in
chondrosarcoma cell proliferation and invasion capacities, and
confirmed its function in regulating primary cilia restoration.
Targeting HDAC6 could achieve regulating ciliogenesis to improve
the prognosis of chondrosarcoma. Primary cilia, a cell superficial
multisensor, can detect mechanical and biochemical signaling in the
extracellular environment and is involved in regulating various
cellular processes (2,3,20).
Many studies demonstrated that primary cilia could affect
chondrocyte development. For example, primary cilia could have a
close link with Golgi apparatus and facilitate the secretion of
newly synthesized cartilage extracellular matrix directly to
maintain chondrocyte phenotype (21). Disruption of cilia-related IFT
proteins resulted in subsequent depletion of primary cilia and
epiphyseal plate dysplasia because of downregulating chondrocytes
proliferation capacity and accelerating hypertrophic
differentiation (22). The shape of
chondrocyte and columnar orientation were also disrupted by the
loss of primary cilia (22,23). Thus, evaluating the role of primary
cilia in cartilage development will help clarify the potential
mechanism of chondrosarcomagenesis.
Although the specific pathogenesis for
chondrosarcoma is still not clear, previous studies have found that
primary cilia deficiency existed in chondrosarcoma cells and the
suppression of endogenous expression of PTHrP could induce cilia
assembly, inhibiting chondrosarcoma cell proliferation and invasion
(24). Abnormal primary cilia
assembly has been confirmed in human neoplastic chondrocytes such
as malignant chondrosarcoma and benign cartilage tumor enchondroma
(17).
HDAC6 is a special member of the HDAC family and
mainly deacetylates the α-tubulin structure (11). Microtubule, as a vital component of
the cytoskeleton, is indispensable to maintain cell morphology and
intercellular junction. In normal mitotic cell cycle, microtubule
plays a role in forming mitotic spindle, and its organizing center
is the centrosome. In the cell division process, microtubule could
depolymerize and remold periodically, and be accompanied by
reversible acetylation modification on Lys40 at N-terminal of
α-tubulin. This process could facilitate primary cilia and spindle
a periodic transformation during the cell cycle (8,9). Thus,
primary cilia assembly could be regulated by HDAC6-mediated
deacetylated modification of α-tubulin. Furthermore, cell division
cycle, morphology, and intercellular junction could also be
influenced by HDAC6, which is directly involved in regulating tumor
cell proliferation, invasion, and other malignant biological
behaviors (12).
Recently, many studies have confirmed that HDAC6
played an important role in tumorigenesis. For example, HDAC6 is
overexpressed in cholangiocarcinoma compared with normal
cholangiocytes, and inhibition of HDAC6 could reverse the malignant
phenotype of cholangiocarcinoma cells (14). Estrogen-mediated upregulation of
HDAC6 had a close relationship with breast cancer cell metastasis
capability and affected the prognosis of patients (13). The present research also revealed
that inhibiting HDAC6 could restore primary cilia assembly and
downregulate the malignant biological behavior of chondrosarcoma
cells such as proliferation and invasion capacities. Aurora A
kinase, as a centrosomal kinase, mainly regulates cell cycle
mitotic spindle organization. Its activation was confirmed to cause
phosphorylation and activation of HDAC6 (19,25–27).
This study revealed that regulating upstream Aurora A activation
led to a similar result in regulating cilia assembly, and HDAC6
inhibitor could affect Aurora A activation at some level. In the
future, we will clarify whether HDAC6 siRNA exerts an effect on
other HDAC family members, or whether there are other molecular
targets.
In conclusion, regulation of ciliogenesis through
targeting HDAC6 could be a promising therapeutic method for
chondrosarcoma treatment. Investigations targeting ciliogenesis
regulation may lead to a new direction in future tumorigenesis
research.
Acknowledgements
This study was supported by the National Natural
Science Foundation of China (grant nos. 81672652 and 81202121).
Glossary
Abbreviations
Abbreviations:
|
HDAC6
|
histone deacetylases 6
|
|
IFT88
|
intraflagellar transport protein
88
|
|
CCK-8
|
Cell Counting Kit-8
|
|
PCNA
|
proliferating cell nuclear antigen
|
References
|
1
|
Leddy LR and Holmes RE: Chondrosarcoma of
bone. Cancer Treat Res. 162:117–130. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Leucht P, Monica SD, Temiyasathit S,
Lenton K, Manu A, Longaker MT, Jacobs CR, Spilker RL, Guo H,
Brunski JB, et al: Primary cilia act as mechanosensors during bone
healing around an implant. Med Eng Phys. 35:392–402. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Muhammad H, Rais Y, Miosge N and Ornan EM:
The primary cilium as a dual sensor of mechanochemical signals in
chondrocytes. Cell Mol Life Sci. 69:2101–2107. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Zimmerman K and Yoder BK: SnapShot:
Sensing and signaling by cilia. Cell. 161:692–2.e1. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Seeger-Nukpezah T, Little JL, Serzhanova V
and Golemis EA: Cilia and cilia-associated proteins in cancer. Drug
Discov Today Dis Mech. 10:e135–e142. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hassounah NB, Bunch TA and McDermott KM:
Molecular pathways: The role of primary cilia in cancer progression
and therapeutics with a focus on Hedgehog signaling. Clin Cancer
Res. 18:2429–2435. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
de Andrea CE, Zhu JF, Jin H, Bovée JV and
Jones KB: Cell cycle deregulation and mosaic loss of Ext1 drive
peripheral chondrosarcomagenesis in the mouse and reveal an
intrinsic cilia deficiency. J Pathol. 236:210–218. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Izawa I, Goto H, Kasahara K and Inagaki M:
Current topics of functional links between primary cilia and cell
cycle. Cilia. 4:122015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ke YN and Yang WX: Primary cilium: An
elaborate structure that blocks cell division? Gene. 547:175–185.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yu F, Ran J and Zhou J: Ciliopathies: Does
HDAC6 represent a new therapeutic target? Trends Pharmacol Sci.
37:114–119. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ran J, Yang Y, Li D, Liu M and Zhou J:
Deacetylation of α-tubulin and cortactin is required for HDAC6 to
trigger ciliary disassembly. Sci Rep. 5:129172015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Valenzuela-Fernández A, Cabrero JR,
Serrador JM and Sánchez-Madrid F: HDAC6: A key regulator of
cytoskeleton, cell migration and cell-cell interactions. Trends
Cell Biol. 18:291–297. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhang Z, Yamashita H, Toyama T, Sugiura H,
Omoto Y, Ando Y, Mita K, Hamaguchi M, Hayashi S and Iwase H: HDAC6
expression is correlated with better survival in breast cancer.
Clin Cancer Res. 10:6962–6968. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Gradilone SA, Radtke BN, Bogert PS, Huang
BQ, Gajdos GB and LaRusso NF: HDAC6 inhibition restores ciliary
expression and decreases tumor growth. Cancer Res. 73:2259–2270.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Dhanyamraju PK, Holz PS, Finkernagel F,
Fendrich V and Lauth M: Histone deacetylase 6 represents a novel
drug target in the oncogenic Hedgehog signaling pathway. Mol Cancer
Ther. 14:727–739. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Butler KV, Kalin J, Brochier C, Vistoli G,
Langley B and Kozikowski AP: Rational design and simple chemistry
yield a superior, neuroprotective HDAC6 inhibitor, Tubastatin A. J
Am Chem Soc. 132:10842–10846. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ho L, Ali SA, Al-Jazrawe M, Kandel R,
Wunder JS and Alman BA: Primary cilia attenuate hedgehog signalling
in neoplastic chondrocytes. Oncogene. 32:5388–5396. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kawasaki M, Ezura Y, Hayata T, Notomi T,
Izu Y and Noda M: TGF-β suppresses Ift88 expression in chondrocytic
ATDC5 Cells. J Cell Physiol. 230:2788–2795. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Pugacheva EN, Jablonski SA, Hartman TR,
Henske EP and Golemis EA: HEF1-dependent Aurora A activation
induces disassembly of the primary cilium. Cell. 129:1351–1363.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Deren ME, Yang X, Guan Y and Chen Q:
Biological and chemical removal of primary cilia affects mechanical
activation of chondrogenesis markers in chondroprogenitors and
hypertrophic chondrocytes. Int J Mol Sci. 17:1882016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Poole CA, Jensen CG, Snyder JA, Gray CG,
Hermanutz VL and Wheatley DN: Confocal analysis of primary cilia
structure and colocalization with the Golgi apparatus in
chondrocytes and aortic smooth muscle cells. Cell Biol Int.
21:483–494. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Song B, Haycraft CJ, Seo HS, Yoder BK and
Serra R: Development of the post-natal growth plate requires
intraflagellar transport proteins. Dev Biol. 305:202–216. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yuan X and Yang S: Primary cilia and
intraflagellar transport proteins in bone and cartilage. J Dent
Res. 95:1341–1349. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Xiang W, Jiang T, Guo F, Xu T, Gong C,
Cheng P, Zhao L, Cheng W and Xu K: Evaluating the role of PTH in
promotion of chondrosarcoma cell proliferation and invasion by
inhibiting primary cilia expression. Int J Mol Sci. 15:19816–19831.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Mahankali M, Henkels KM, Speranza F and
Gomez-Cambronero J: A non-mitotic role for Aurora kinase A as a
direct activator of cell migration upon interaction with PLD, FAK
and Src. J Cell Sci. 128:516–526. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Al-Bataineh MM, Alzamora R, Ohmi K, Ho PY,
Marciszyn AL, Gong F, Li H, Hallows KR and Pastor-Soler NM: Aurora
kinase A activates the vacuolar H+-ATPase (V-ATPase) in kidney
carcinoma cells. Am J Physiol Renal Physiol. 310:F1216–F1228. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kozyreva VK, McLaughlin SL, Livengood RH,
Calkins RA, Kelley LC, Rajulapati A, Ice RJ, Smolkin MB, Weed SA
and Pugacheva EN: NEDD9 regulates actin dynamics through cortactin
deacetylation in an AURKA/HDAC6-dependent manner. Mol Cancer Res.
12:681–693. 2014. View Article : Google Scholar : PubMed/NCBI
|