Introduction
MicroRNAs (miRNAs) are a class of highly conserved
small non-coding single-stranded RNAs, and they are endogenously
expressed in many species. These miRNAs regulate gene expression by
targeting the 3′ untranslated regions (3′UTRs) of mRNAs (1–6).
Numerous studies have shown that miRNAs act as intrinsic regulators
of many cellular processes, including cell proliferation,
apoptosis, invasion and differentiation (7–12).
Aberrant expression of miRNAs has been found to be associated with
the development and progression of numerous types of cancers.
Existing research shows that aberrant expression of miRNAs has
prognostic significance for certain types of cancer, such as lung
and esophageal cancer, lymphocytic leukemia and neuroblastoma
(13–16).
Lung cancer is a major cause of cancer-related death
worldwide (17,18). Although chemotherapy and
molecular-targeted therapy have largely improved recently, the
survival rate of lung cancer patients remains poor. The
invasiveness and metastasis of non-small cell lung cancer (NSCLC)
cells are still critical challenges in clinical management.
Identifying novel and effective molecules that may suppress the
invasiveness and metastasis of lung cancer cells may facilitate the
development of anti-lung cancer strategies. Numerous studies have
shown that expression of many miRNAs is altered in lung
adenocarcinoma (19–21), indicating that deregulation of
miRNAs may play a role in the carcinogenesis of lung
adenocarcinoma. We previously conducted an miRNA chip-based
expression analysis of lung adenocarcinoma tissues and found that
miR-125a-5p expression in lung adenocarcinoma tissues was largely
lower than that in paired non-tumor tissues. Previous studies have
also shown that miR-125a suppresses tumor growth, invasion and
metastasis in cervical cancer, oral squamous cell and
hepatocellular carcinoma, and lung cancer (22–25).
Based on bioinformatic analyses, we hypothesized
that neural precursor cell expressed, developmentally downregulated
9 (NEDD9) may act as a target of miR-125a-5p. NEDD9 is a
non-catalytic scaffolding protein, which is a member of the
Crk-associated substrate (CAS) family (26).
The effects of miR-125a-5p on invasiveness and
metastasis of lung adenocarcinoma cells are unclear to date. In the
present study, we investigated miR-125a-5p expression levels in
tumor and normal tissues from 68 lung adenocarcinoma patients, and
observed its concomitant alteration in regards to lung
adenocarcinoma cell proliferation, apoptosis and metastasis.
Materials and methods
Patient sample collection
The present study was approved by the Ethics
Committee of Henan University and informed consent was obtained
from all patients. Sixty-eight lung samples (lung adenocarcinoma
and adjacent non-tumor lung tissues) were obtained from The First
Affiliated Hospital of Zhengzhou University (Zhengzhou, China), and
the Cancer Hospital of Henan Province (Zhengzhou, China) between
November 2013 and March 2015. All the samples were frozen in liquid
nitrogen immediately after resection until use. None of the
patients enrolled in the study had received chemotherapy or
radiotherapy. All tumors were histopathologically confirmed as lung
adenocarcinoma by two independent qualified clinical pathologists.
The clinical data of 68 cases of lung adenocarcinoma are shown in
Table I.
 | Table I.Relative expression of miR-125a and
NEDD9 in lung adenocarcinoma tissues and the correlation with
clinicopathological parameters of 68 cases with lung
adenocarcinoma. |
Table I.
Relative expression of miR-125a and
NEDD9 in lung adenocarcinoma tissues and the correlation with
clinicopathological parameters of 68 cases with lung
adenocarcinoma.
| Parameter | n | miR-125a | P-value | NEDD9 mRNA | P-value |
|---|
| Sex |
|
Male | 42 | 0.361±0.056 | 0.567 | 1.458±0.154 | 0.573 |
|
Female | 26 | 0.335±0.105 |
| 1.507±0.039 |
|
| Age (years) |
|
<60 | 40 | 0.349±0.162 | 0.803 | 1.528±0.132 | 0.453 |
|
≥60 | 28 | 0.341±0.189 |
| 1.469±0.087 |
|
|
Differentiation |
|
Well | 13 | 0.521±0.165 | 0.001a | 1.228±0.371 | 0.005a |
|
Moderate | 35 | 0.328±0.124 |
| 1.517±0.267 |
|
|
Poor | 20 | 0.279±0.058 |
| 1.584±0.158 |
|
| TNM stage |
| I | 27 | 0.447±0.116 | 0.001a | 1.307±0.166 | 0.001a |
| II | 31 | 0.295±0.034 |
| 1.589±0.203 |
|
|
III | 10 | 0.233±0.074 |
| 1.623±0.206 |
|
| Lymph node
metastasis |
|
Negative | 25 | 0.450±0.251 | 0.009a | 1.273±0.160 | 0.001a |
|
Positive | 43 | 0.312±0.279 |
| 1.597±0.206 |
|
RNA extraction and quantitative
real-time PCR
Total RNA was extracted from lung adenocarcinoma and
adjacent non-tumor tissue samples using TRIzol reagent (Invitrogen,
Carlsbad, CA, USA). RNA (1 µg) was used to synthesize cDNA. The
expression level of miR-125a-5p was determined using a
High-Specificity miR-125a-5p qRT-PCR Detection kit (Stratagene
Corp., La Jolla, CA, USA) using qRT-PCR (ABI 7500 Fast System;
Applied Biosystems, Foster City, CA, USA). All protocols were
performed according to the manufacturer's instructions. U6 small
nuclear RNA (U6 snRNA) was used as an endogenous control for
normalization. Relative expression level of miR-125a-5p was
calculated using the 2−ΔΔCt method. miR-125a-5p primers
were as follows: RT-primer,
5′-GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACAGGGACTC-3′;
forward, 5′-TCCGAAGTGTCCAATTTCCC-3′ and reverse,
5′-GTGCAGGGTCCGAGGT-3′; U6 primers were as follows: RT-primer,
5′-GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGA CAAAATATGGAACTGC-3′;
forward, 5′-CTCGCTTCGGCAGCACA-3′ and reverse,
5′-GTGCAGGGTCCGAGGT-3′.
Cell culture
Human lung adenocarcinoma cell lines (A549 and
H1299) were purchased from the Type Culture Collection of the
Chinese Academy of Sciences (Shanghai, China). Both cell lines were
maintained in Dulbecco's modified Eagle's medium supplemented with
10% fetal bovine serum (FBS; Gibco-BRL, Gaithersburg, MD, USA) and
incubated at 37°C/5% CO2.
miRNA transfection
The miR-125a-5p mimic (GMR-miR™ miRNA-125a-5p mimic)
and the scrambled oligonucleotide used in the present study were
synthesized by Shanghai GenePharma Co. Ltd. (Shanghai, China). For
cell transfection, lung adenocarcinoma cell lines A549 and H1299
were seeded into 6-well plates at a density of 1.5×105
cells/well. Once cells reached 50~80% confluency, transient
transfection was conducted using Lipofectamine™ 2000 (Invitrogen),
and the working concentration of miR-125a-5p mimic was 50 nM. Cells
from each cell line were subdivided into three groups: the blank
group (blank) that was non-transfected, scramble group (scramble)
that was transfected with the scrambled oligonucleotide, and the
miR-125a-5p mimic group (miR-125a-5p mimic) that was transfected
with the miR-125a-5p mimic.
Cell growth assay
Cells (blank, scramble and miR-125a-5p mimic) were
seeded into a 96-well plate at a density of 1×104
cells/well, with five replicate wells/group. The absorbance value
for each well was determined using Cell Counting Kit-8 (CCK-8;
Dojindo, Kumamoto, Japan). The optical density (OD) was detected
using CCK-8 reagents after 0, 24, 48 and 72 h of cultivation. The
OD was recorded using a microplate reader (ELx800; BioTek,
Winooski, VT, USA), at a wavelength of 450 nm (OD450).
The experiments were independently triplicated.
Transwell assay
Transwell filters (Costar, Cambridge, MA, USA) were
coated with Matrigel (3.9 µg/µl, 60–80 µl) on the upper surface of
a polycarbonic membrane (6.5-mm diameter, 8-µm pore size). After 30
min of incubation at 37°C, the Matrigel solidified and served as
the extracellular matrix for tumor cell invasion analysis. Three
groups of cells (blank, scramble and miR-125a-5p mimic) were
adjusted to 2×105 cells/ml, and resuspended in 200 µl of
serum-free medium in the upper chambers, and the lower chamber was
supplied with 500 µl of medium containing 10% FBS, followed by
incubation at 37°C in 5% CO2 for 48 h. After incubation,
the medium was removed from the upper chamber and cells in the
upper chamber were carefully removed with a cotton swab, and then
stained with Methylene Blue Staining solution (Beyotime, Haimen,
China). The number of cells invading the Matrigel was counted from
three randomly selected visual fields, using an inverted microscope
at a magnification of ×200. All experiments were performed in
triplicate.
Apoptosis assay
Three groups of cells (blank, scramble and
miR-125a-5p mimic) were harvested at 48 h post-transfection by
trypsinization, and were adjusted to 1×106 cells/ml in
1X binding buffer. After staining with FITC-Annexin V and propidium
iodide (PI) using the FITC-Annexin V Apoptosis Detection Kit I
(BestBio, Shanghai, China), cells were analyzed using a
FACScan® flow cytometer (BD Biosciences, Franklin Lakes,
NJ, USA).
Caspase activity assay
Cells from each treatment group (blank, scramble and
miR-125a-5p mimic) were harvested at 48 h post-transfection. Then
caspase activity was conducted using a caspase activity assay kit
(Beyotime) according to the manufacturers instructions. Cellular
extracts and substrates (Ac-DEVD-pNA) were kept into 96-well plates
for 2 h at 37°C. Absorbance values were recorded using a microplate
reader (ELx800), at a wavelength of 405 nm.
Dual-luciferase assay
The human NEDD9 3′ UTR fragments containing putative
binding sites for miR-125a-5p were amplified by PCR from human
genomic DNA. Overlap extension PCR was used to obtain the mutant
NEDD9 3′ UTRs. These DNA fragments were cloned into a pmir-GLO
reporter vector (Promega, Madison, WI, USA), downstream of the
luciferase gene, to generate the recombinant vectors, wild-type-3′
UTR and mutant type 3′ UTR. For the luciferase reporter assay, A549
cells were transiently co-transfected with miRNA (miR-125a-5p mimic
or scrambled-125a-5p) and reporter vectors (wild-type-3′ UTR or
mutant type 3′ UTR), using Lipofectamine™ 2000. Luciferase
activities were measured 48 h post-transfection using the
Dual-Luciferase Assay kit (Promega), according to the
manufacturer's instructions.
Western blotting
Total proteins of the cultured cells were extracted
using RIPA buffer containing phenylmethanesulfonyl fluoride (PMSF)
48 h post-transfection. The protein concentrations were determined
using BCA protein assay kit (Beyotime). Proteins (30 µg) were
subjected to sodium dodecyl sulfate-polyacrylamide gel
electrophoresis (SDS-PAGE) and transferred onto polyvinylidene
fluoride (PVDF) membranes. After blocking with 5% BSA in 0.05%
Tween-20-TBS for 1 h, the membranes were incubated overnight at 4°C
with diluted (1:1,000) primary antibodies (polyclonal rabbit
anti-NEDD9; Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA).
After extensive washing with TBST, the membranes were incubated
with diluted (1:3,000) anti-rabbit IgG-HRP secondary antibody
(Santa Cruz Biotechnology, Inc.). Signals were detected using a
chemiluminescence detection kit (Amersham Pharmacia Biotech,
Piscataway, NJ, USA). GAPDH (Santa Cruz Biotechnology, Inc.) was
used as an endogenous reference.
Statistical analysis
All analyses were performed using SPSS 17.0
software. All data are expressed as the mean ± standard deviation
(SD). One-way analysis of variance (ANOVA) was used to analyze the
multiple groups data. A value of P<0.05 was considered
statistically significant.
Results
Downregulation of miR-125a-5p and
upregulation of NEDD9 in lung adenocarcinoma
The clinicopathological characteristics of the 68
lung adenocarcinoma cases included in the present study are
presented in Table I. We found that
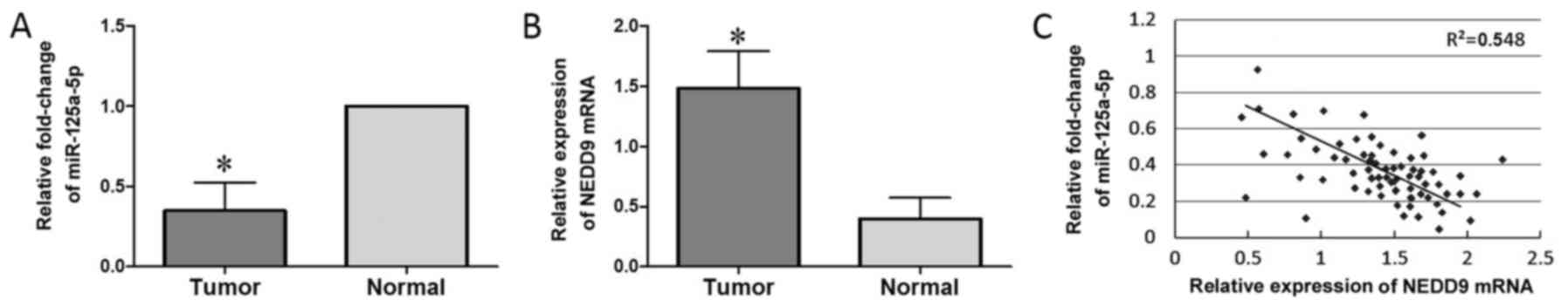
the miR-125a-5p expression level in lung adenocarcinoma tissues was
associated with differentiation status, tumor-node-metastasis (TNM)
stage and lymph node metastasis (P<0.05; Table I). No significant differences were
observed between miR-125a-5p expression and sex or age. Using
adjacent non-tumor tissues as reference, miR-125a-5p expression in
lung adenocarcinoma tissues was found to be significantly reduced
(P<0.05; Fig. 1A). Compared to
adjacent non-cancerous tissues, the expression level of NEDD9 in
lung adenocarcinoma tissues was higher (P<0.05; Fig. 1B). To investigate the correlation
between miR-125a-5p and NEDD9, we examined their expression levels
in primary human lung adenocarcinoma tissues. Unlike the matched
normal lung tissues, in the tumor tissues from the 68 patients with
lung adenocarcinoma, miR-125a-5p was reduced, whereas NEDD9 protein
was increased, which demonstrated a significant negative
correlation (R2=0.632; P<0.01; Fig. 1C). The data suggest that the
expression of NEDD9 and miR-125a-5p have an inverse correlation in
lung adenocarcinoma tissues.
Upregulation of miR-125a-5p inhibits
the proliferation of A549 and H1299 cells
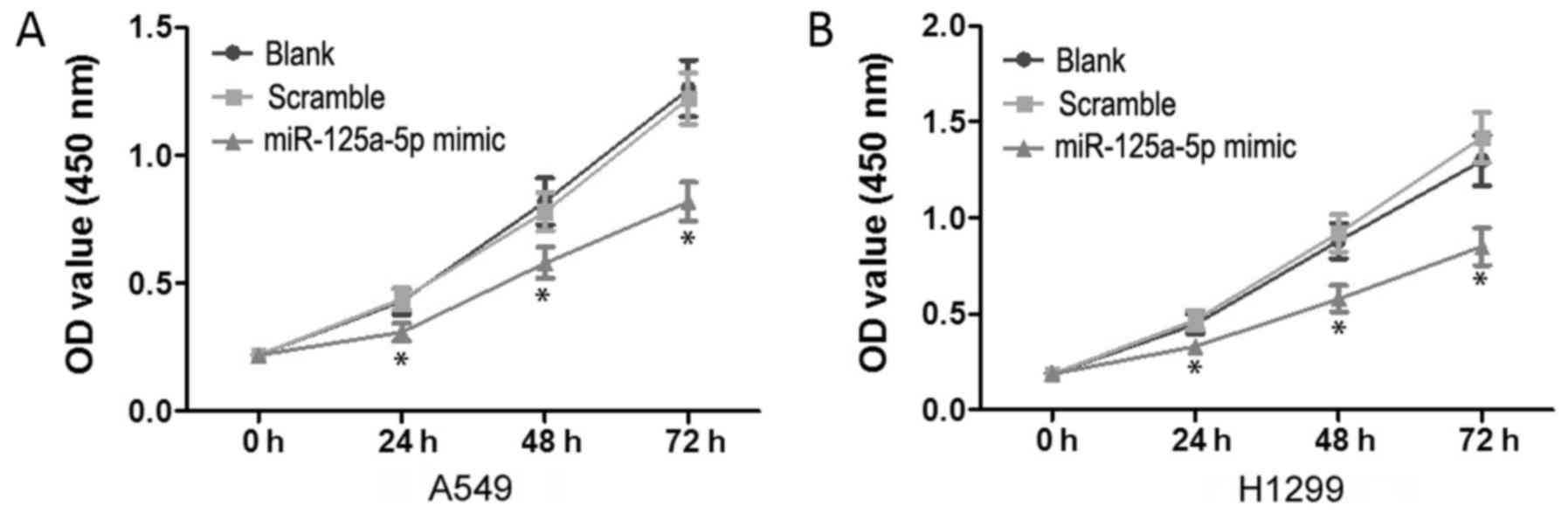
To test the proliferation effects of miR-125a-5p on
lung adenocarcinoma cells, we performed CCK-8 assay. The
corresponding cell growth curves are presented in Fig. 2. There were no significant
differences in OD450 values between the blank and
scramble groups (P>0.05). However, compared to the blank and
scramble groups, the OD450 values for the miR-125a-5p
mimic group on 24, 48 and 72 h were significantly decreased
(P<0.05) in both the A549 (Fig.
2A) and H1299 (Fig. 2B) cell
lines.
Upregulation of miR-125a-5p inhibits
the invasive ability of A549 and H1299 cells
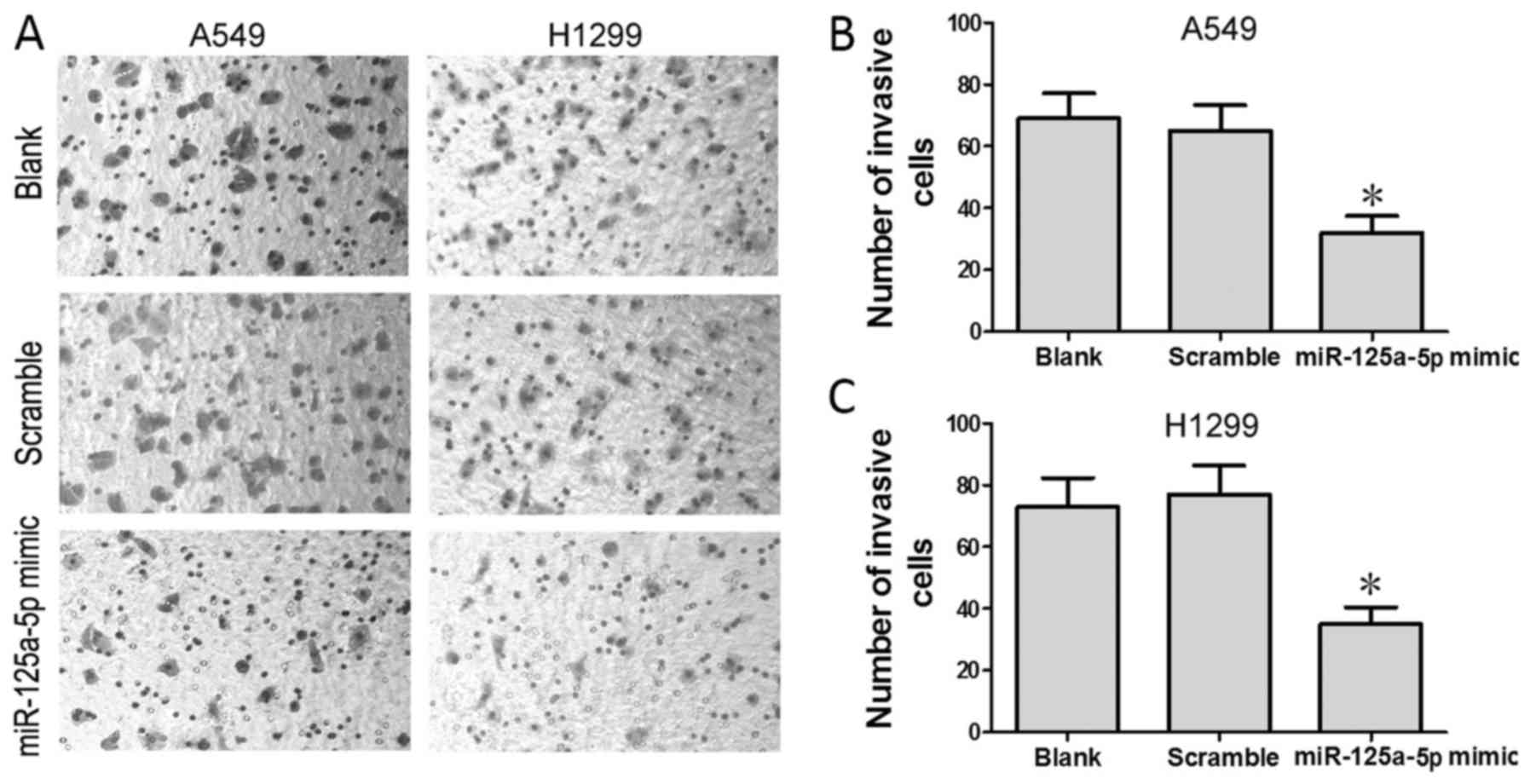
To test the effect of miR-125a-5p on the invasive
ability of lung adenocarcinoma cells, we performed Transwell assay.
We found that the mean number of cells that penetrated the
Transwell membrane was significantly lower in the miR-125a-5p mimic
group than these numbers in the blank and scramble groups
(P<0.05) in both the A549 (Fig.
3B) and H1299 (Fig. 3C) cell
lines.
Upregulation of miR-125a-5p induces
the apoptosis of A549 and H1299 cells
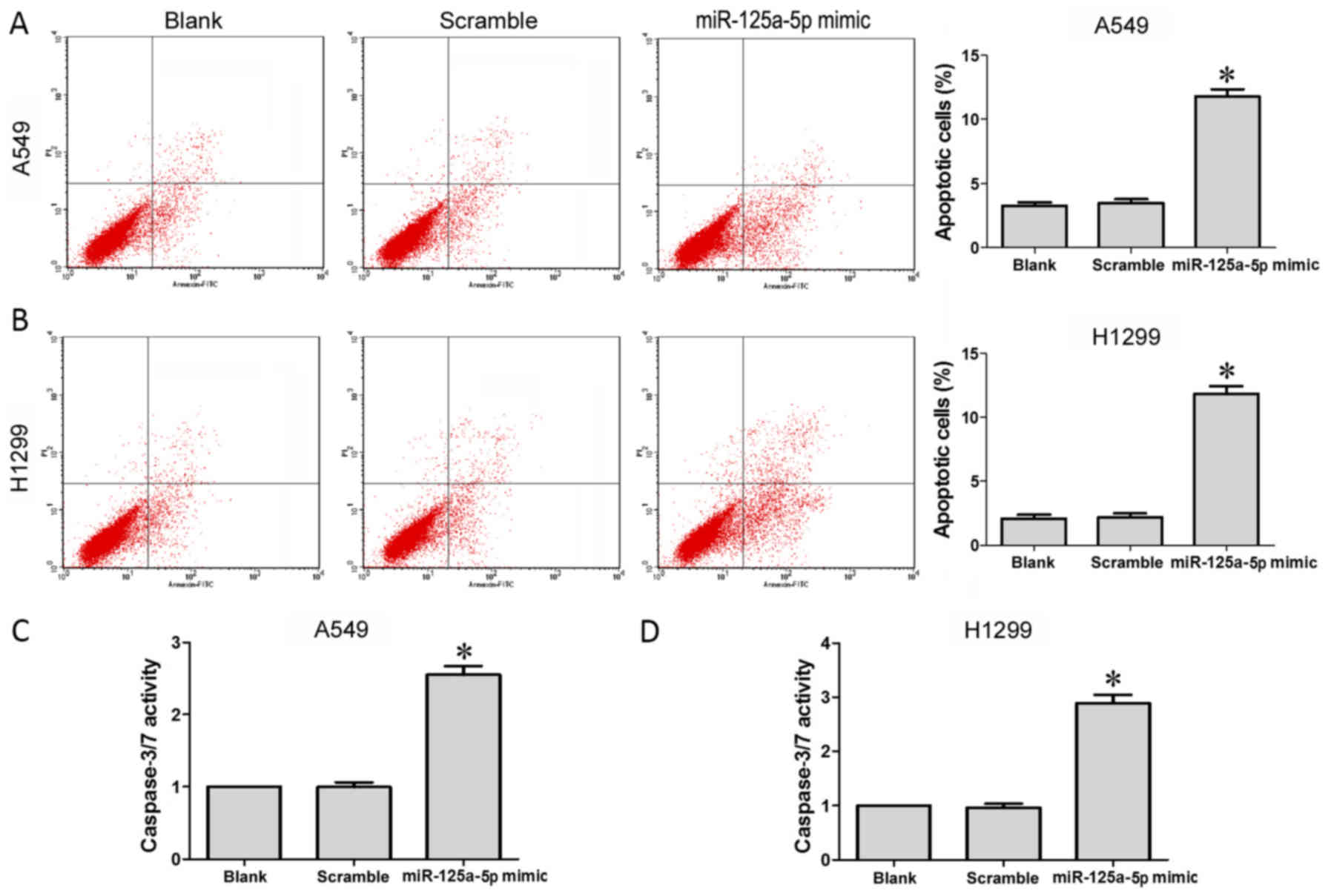
Our FCM results indicated that the apoptosis level
of cells transfected with the miR-125a-5p mimic was significantly
enhanced compared to these levels in the cells in the blank and
scramble groups (P<0.05) in both the A549 (Fig. 4A) and H1299 (Fig. 4B) cell lines. Similarly,
transfection of the A549 and H1299 cells with the miR-125a-5p mimic
was found to significantly increase caspase-3/−7 activity compared
to cells in the blank and scramble groups (P<0.05) in both the
A549 (Fig. 4C) and H1299 (Fig. 4D) cell lines.
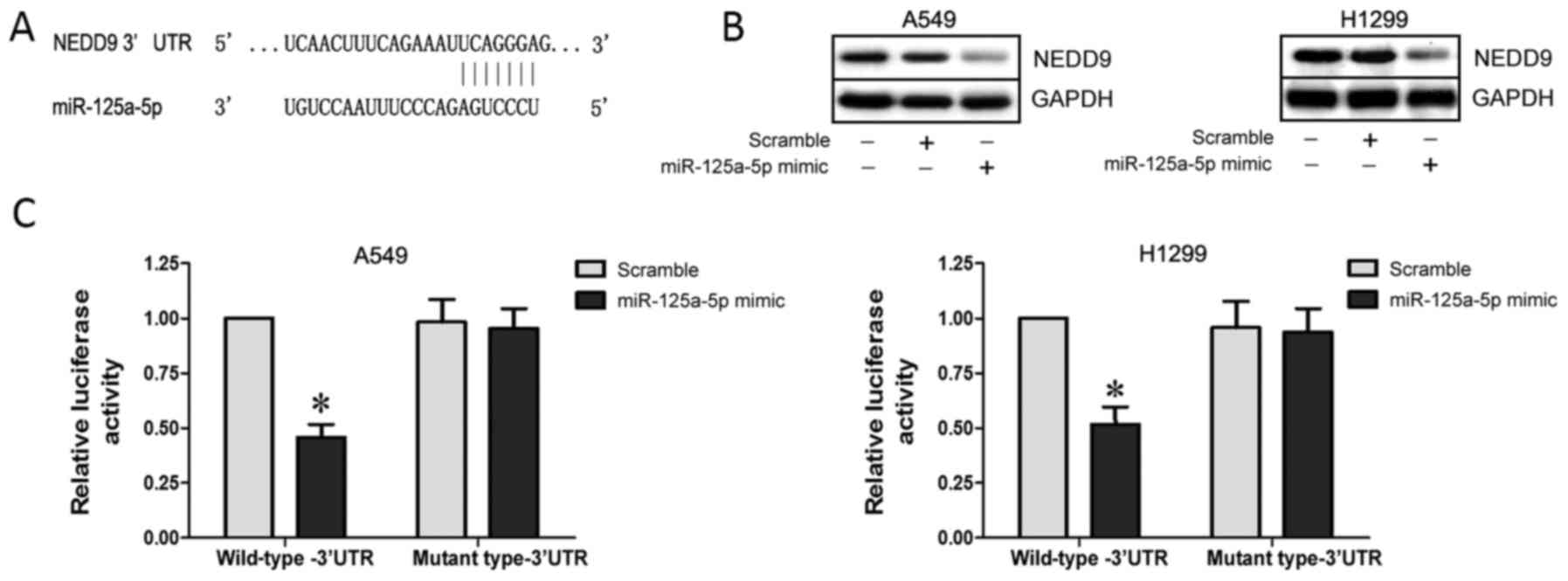
NEDD9 is a direct target of
miR-125a-5p
Bioinformatic analyses using TargetScan (www.targetscan.org) and miRanda (www.microrna.org) predicted that the 3′ UTR of NEDD9
contains binding sites for miR-125a-5p (Fig. 5A). Subsequent western blot analysis
indeed showed that NEDD9 expression was downregulated in the A549
and H1299 cells following transfection with the miR-125a-5p mimic
(P<0.05; Fig. 5B). To verify
whether NEDD9 is a direct target of miR-125a-5p, we used a
dual-luciferase reporter system containing either wild-type or
mutant 3′ UTR of NEDD9, respectively. Co-transfection with
miR-125a-5p and the reporter vector containing the wild-type-3′ UTR
significantly suppressed luciferase activity (P<0.05; Fig. 5C). These results indicate that
miR-125a-5p negatively regulates NEDD9 expression by directly
binding to putative binding sites in the 3′ UTR.
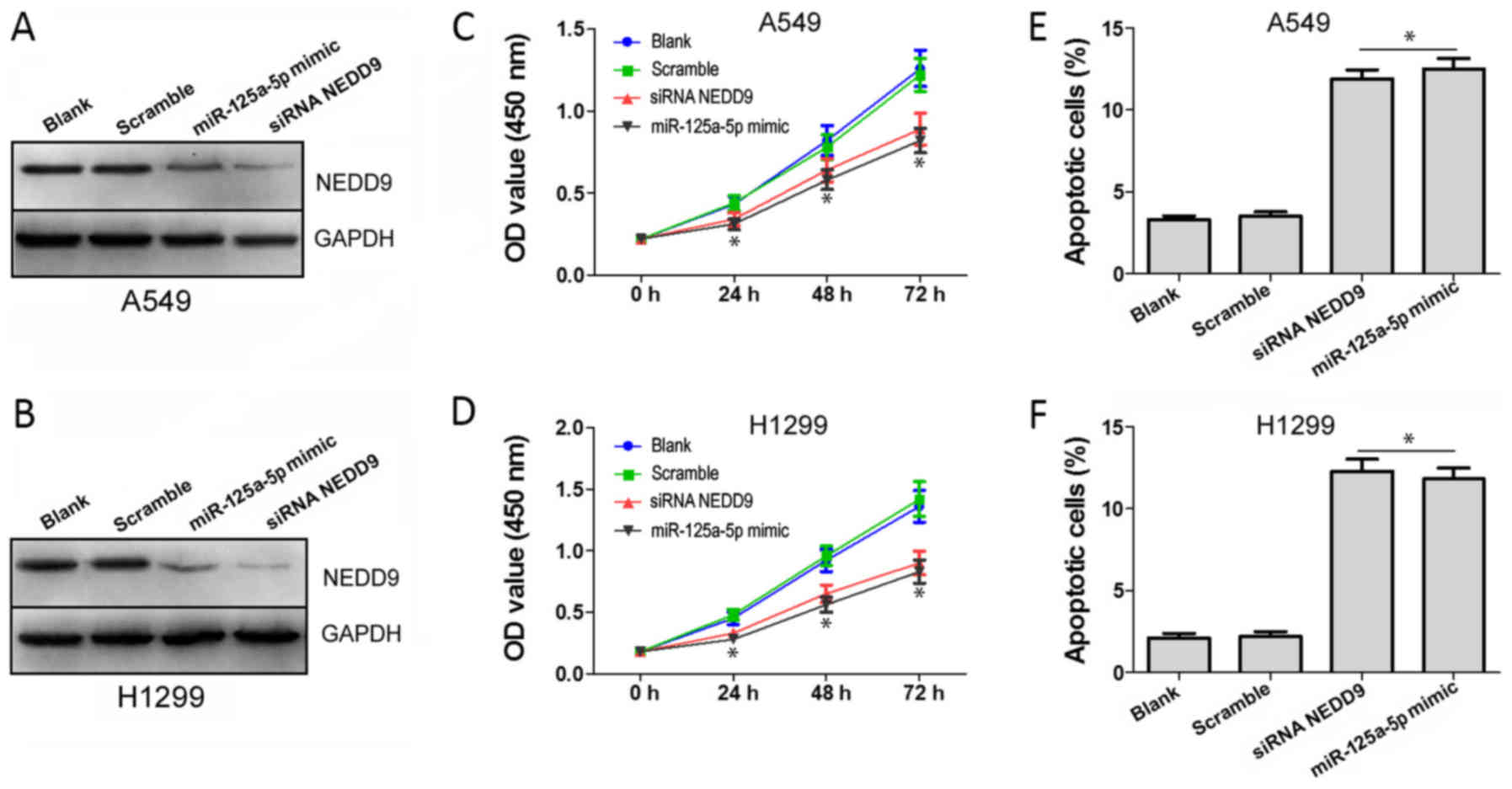
NEDD9 silencing and miR-125a-5p
overexpression exert antiproliferative and pro-apoptotic effects on
A549 and H1299 cells
In order to verify whether miR-125a-5p functions by
targeting NEDD9 in lung adenocarcinoma cells, we divided cells into
four groups (blank, scramble, miR-125a-5p mimic and siRNA NEDD9),
and conducted CCK-8 and FCM assays. Western blot analysis indeed
showed that NEDD9 expression of the miR-125a-5p mimic and siRNA
NEDD9 groups were downregulated (P<0.05) in the A549 (Fig. 6A) and H1299 cells (Fig. 6B). CCK-8 assay results showed that
the OD450 values of miR-125a-5p mimic and siRNA NEDD9
groups on days 24, 48 and 72 h were significantly decreased
(P<0.05) in both the A549 (Fig.
6C) and H1299 (Fig. 6D) cells,
compared to the blank and scramble groups. Results of FCM indicated
that the apoptosis levels in the miR-125a-5p mimic and siRNA NEDD9
groups were significantly enhanced compared to those of the cells
in the blank and scramble groups (P<0.05) in both the A549
(Fig. 6E) and H1299 (Fig. 6F) cell lines. These results indicate
that miR-125a-5p suppresses proliferation and induces cell
apoptosis by targeting NEDD9 in A549 and H1299 cells.
Discussion
Lung cancer is regarded as one of the leading causes
of cancer-related death worldwide. Non-small cell lung cancer
(NSCLC) accounts for ~80% of all lung cancer cases with a 15%
5-year survival. Numerous studies have shown that expression of
miRNAs is altered in lung adenocarcinoma (19–21),
indicating that deregulation of miRNAs may play a role in the
carcinogenesis of lung adenocarcinoma. Some research has estimated
that miRNAs may regulate up to 30% of all human genes and control a
variety of cellular processes (27–29).
miRNAs have been shown to be deregulated in various cancers, and
their expression levels are related with the diagnosis and
prognosis of different types of tumors (29–31).
In the present study, we found that miR-125a-5p
expression was downregulated in human lung adenocarcinoma tissues,
whereas NEDD9 expression was upregulated. In addition, the
expression levels of NEDD9 and miR-125a-5p were found to have an
inverse correlation in lung adenocarcinoma tissues. In addition, we
also found that the expression level of miR-125a-5p is associated
with lymph node metastasis, TNM stage and the differentiation
status in lung adenocarcinoma. Upregulation of miR-125a-5p in A549
and H1299 cells was found to suppress the proliferative and
invasive capacities of lung adenocarcinoma cells, and promote
apoptosis at the same time. These results suggest that the
expression of miR-125a-5p has a close correlation with lung
adenocarcinoma development and progression.
NEDD9 is a non-catalytic scaffolding protein, which
belongs to the Crk-associated substrate (CAS) family. Large studies
have shown that the NEDD9 protein could act as a biomarker of
invasive capacity and an essential key for pro-metastatic behavior
in many types of cancer, including breast cancer and melanoma
(32–34). The interaction of NEDD9 with FAK and
Src leads to the tyrosine phosphorylation of NEDD9, and regulates
and activates transcription pathways in the end (35). However, the function of NEDD9 in
lung adenocarcinoma is not clear, and to date no studies have
investigated whether NEDD9 expression is regulated by miR-125a-5p
in lung adenocarcinoma. In the present study, we confirmed that
miR-125a-5p upregulation negatively regulates the expression of
NEDD9. miR-125a-5p was found to suppress the metastatic and the
invasive abilities of lung adenocarcinoma cell lines A549 and
H1299. NEDD9 was shown to act as a direct functional target of
miR-125a-5p using western blot and luciferase reporter assays.
Moreover, CCK-8 and FCM assay results showed that miR-125a-5p
suppressed proliferation and induced cell apoptosis by targeting
NEDD9 in A549 and H1299 cells. Taken together, our findings
indicate that miR-125a-5p may act as a tumor-suppressor by
targeting NEDD9.
In conclusion, we demonstrated that miR-125a-5p is
downregulated in lung adenocarcinoma, and upregulation of
miR-125a-5p inhibits the proliferative and invasive capacities and
promotes apoptosis in lung adenocarcinoma cell lines A549 and
H1299. The present study established an experimental and
theoretical foundation for individualized chemotherapy for lung
adenocarcinoma patients.
Acknowledgements
We thank all staff at the Research Centre who
contributed to the present study. The present study was supported
by a grant from the Ministry of Major Science and Technology of
Henan (201401005).
References
|
1
|
Abdelrahim M, Smith R III, Burghardt R and
Safe S: Role of Sp proteins in regulation of vascular endothelial
growth factor expression and proliferation of pancreatic cancer
cells. Cancer Res. 64:6740–6749. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Brennecke J and Cohen SM: Towards a
complete description of the microRNA complement of animal genomes.
Genome Biol. 4:2282003. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Calin GA and Croce CM: MicroRNA signatures
in human cancers. Nat Rev Cancer. 6:857–866. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Chan JA, Krichevsky AM and Kosik KS:
MicroRNA-21 is an antiapoptotic factor in human glioblastoma cells.
Cancer Res. 65:6029–6033. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Dynan WS and Tjian R: The
promoter-specific transcription factor Sp1 binds to upstream
sequences in the SV40 early promoter. Cell. 35:79–87. 1983.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Enzinger PC and Mayer RJ: Esophageal
cancer. N Engl J Med. 349:2241–2252. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Esteller M: Non-coding RNAs in human
disease. Nat Rev Genet. 12:861–874. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Guo Y, Chen Z, Zhang L, Zhou F, Shi S,
Feng X, Li B, Meng X, Ma X, Luo M, et al: Distinctive microRNA
profiles relating to patient survival in esophageal squamous cell
carcinoma. Cancer Res. 68:26–33. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Han Y, Chen J, Zhao X, Liang C, Wang Y,
Sun L, Jiang Z, Zhang Z, Yang R, Chen J, et al: MicroRNA expression
signatures of bladder cancer revealed by deep sequencing. PLoS One.
6:e182862011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
He H, Jazdzewski K, Li W, Liyanarachchi S,
Nagy R, Volinia S, Calin GA, Liu CG, Franssila K, Suster S, et al:
The role of microRNA genes in papillary thyroid carcinoma. Proc
Natl Acad Sci USA. 102:19075–19080. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Iorio MV, Ferracin M, Liu CG, Veronese A,
Spizzo R, Sabbioni S, Magri E, Pedriali M, Fabbri M, Campiglio M,
et al: MicroRNA gene expression deregulation in human breast
cancer. Cancer Res. 65:7065–7070. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Iorio MV, Visone R, Di Leva G, Donati V,
Petrocca F, Casalini P, Taccioli C, Volinia S, Liu CG, Alder H, et
al: MicroRNA signatures in human ovarian cancer. Cancer Res.
67:8699–8707. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Jensen RH, Tiirikainen M, You L, Ginzinger
D, He B, Uematsu K, Xu Z, Treseler P, McCormick F and Jablons DM:
Genomic alterations in human mesothelioma including high resolution
mapping of common regions of DNA loss in chromosome arm 6q.
Anticancer Res. 23:2281–2289. 2003.PubMed/NCBI
|
|
14
|
Kim IK and Jung YK, Noh DY, Song YS, Choi
CH, Oh BH, Masuda ES and Jung YK: Functional screening of genes
suppressing TRAIL-induced apoptosis: Distinct inhibitory activities
of Bcl-XL and Bcl-2. Br J Cancer. 88:910–917. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kong LM, Liao CG, Fei F, Guo X, Xing JL
and Chen ZN: Transcription factor Sp1 regulates expression of
cancer-associated molecule CD147 in human lung cancer. Cancer Sci.
101:1463–1470. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kozomara A and Griffiths-Jones S: miRBase:
Integrating microRNA annotation and deep-sequencing data. Nucleic
Acids Res. 39:D152–D157. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chen WQ, Zhang SW, Zou XN and Zhao P:
Cancer incidence and mortality in china, 2006. Chin J Cancer Res.
23:3–9. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Shi L, Zhang B, Sun X, Lu S, Liu Z, Liu Y,
Li H, Wang L, Wang X and Zhao C: MiR-204 inhibits human NSCLC
metastasis through suppression of NUAK1. Br J Cancer.
111:2316–2327. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Guo H, Li W, Zheng T and Liu Z: MiR-195
targets HDGF to inhibit proliferation and invasion of NSCLC cells.
Tumour Biol. 35:8861–8866. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wang X, Wang Y, Lan H and Li J: MiR-195
inhibits the growth and metastasis of NSCLC cells by targeting
IGF1R. Tumour Biol. 35:8765–8770. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Bi Q, Tang S, Xia L, Du R, Fan R, Gao L,
Jin J, Liang S, Chen Z, Xu G, et al: Ectopic expression of MiR-125a
inhibits the proliferation and metastasis of hepatocellular
carcinoma by targeting MMP11 and VEGF. PLoS One. 7:e401692012.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Tiwari A, Shivananda S, Gopinath KS and
Kumar A: MicroRNA-125a reduces proliferation and invasion of oral
squamous cell carcinoma cells by targeting estrogen-related
receptor α: Implications for cancer therapeutics. J Biol Chem.
289:32276–32290. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Fan Z, Cui H, Xu X, Lin Z, Zhang X, Kang
L, Han B, Meng J, Yan Z, Yan X, et al: MiR-125a suppresses tumor
growth, invasion and metastasis in cervical cancer by targeting
STAT3. Oncotarget. 6:25266–25280. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Jiang L, Huang Q, Zhang S, Zhang Q, Chang
J, Qiu X and Wang E: Hsa-miR-125a-3p and hsa-miR-125a-5p are
downregulated in non-small cell lung cancer and have inverse
effects on invasion and migration of lung cancer cells. BMC Cancer.
10:3182010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Seo S, Ichikawa M and Kurokawa M:
Structure and function of cas-L and integrin-mediated signaling.
Crit Rev Immunol. 26:391–406. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Sun T, Wang C, Xing J and Wu D: miR-429
modulates the expression of c-myc in human gastric carcinoma cells.
Eur J Cancer. 47:2552–2559. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Suske G, Bruford E and Philipsen S:
Mammalian SP/KLF transcription factors: Bring in the family.
Genomics. 85:551–556. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Tavazoie SF, Alarcón C, Oskarsson T, Padua
D, Wang Q, Bos PD, Gerald WL and Massagué J: Endogenous human
microRNAs that suppress breast cancer metastasis. Nature.
451:147–152. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wang Y, Zang W, Du Y, Ma Y, Li M, Li P,
Chen X, Wang T, Dong Z and Zhao G: Mir-655 up-regulation suppresses
cell invasion by targeting pituitary tumor-transforming gene-1 in
esophageal squamous cell carcinoma. J Transl Med. 11:3012013.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wang Y, Li M, Zang W, Ma Y, Wang N, Li P,
Wang T and Zhao G: MiR-429 up-regulation induces apoptosis and
suppresses invasion by targeting Bcl-2 and SP-1 in esophageal
carcinoma. Cell Oncol. 36:385–394. 2013. View Article : Google Scholar
|
|
32
|
Singh MK, Izumchenko E, Klein-Szanto AJ,
Egleston BL, Wolfson M and Golemis EA: Enhanced genetic instability
and dasatinib sensitivity in mammary tumor cells lacking NEDD9.
Cancer Res. 70:8907–8916. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Chang JX, Gao F, Zhao GQ and Zhang GJ:
Expression and clinical significance of NEDD9 in lung tissues. Med
Oncol. 29:2654–2660. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Kim M, Gans JD, Nogueira C, Wang A, Paik
JH, Feng B, Brennan C, Hahn WC, Cordon-Cardo C, Wagner SN, et al:
Comparative oncogenomics identifies NEDD9 as a melanoma metastasis
gene. Cell. 125:1269–1281. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
O'Neill GM, Seo S, Serebriiskii IG, Lessin
SR and Golemis EA: A new central scaffold for metastasis: Parsing
HEF1/Cas-L/NEDD9. Cancer Res. 67:8975–8979. 2007. View Article : Google Scholar : PubMed/NCBI
|