Introduction
Breast cancer is the most frequently diagnosed
cancer, as well as, the leading cause of death in the female
population worldwide (1). The
aetiology of breast cancer remains largely unknown, probably due to
its multifactorial nature, but several risk factors have been
identified (2). These include
reproductive and hormonal factors, environmental factors (including
lifestyle changes with a higher consumption of animal fat, obesity
and limited physical activity) and hereditary factors.
Poor prognosis of breast cancer is strongly related
to advanced tumour stage and lymph node spread, although the
importance of specific molecular markers has also been investigated
(3–5). For a tumour to grow and metastasise it
needs to develop its own blood supply and this is achieved through
angiogenesis. The surrounding endothelial cells are stimulated to
form new blood vessels by growth factors secreted from both the
tumour and stroma. The process of angiogenesis is not the result of
one single growth factor but rather is dependent on the interaction
of multiple proteins with pro- or anti-angiogenic properties. In
breast cancer, like almost all other solid tumours, growth factors
such as vascular endothelial growth factor (VEGF), basic fibroblast
growth factor (FGF), hepatocyte growth factor (HGF), platelet
derived growth factor (PDGF), monocyte chemoattractant protein
(MCP)-1, macrophage inflammatory protein (MIP)-1β, interleukin
(IL)-8 and the regulated on activation, normal T cell expressed and
secreted (RANTES), are expressed and, depending on the subtype of
breast cancer, have been identified as negative prognostic factors
for patient survival (6–10). These growth factors may be
responsible for inducing angiogenesis as they are pro-angiogenic,
however, breast cancer cells and tissues can also produce
inhibitors of angiogenesis, including thrombospondin, angiostatin
and endostatin (11). Thus, it is
becoming clear that angiogenesis in cancer progression is the
result of a net balance between positive and negative regulators of
tubule formation. This evidence directs further investigation into
the importance of assessing the roles and levels of angiogenic
factors in plasma from women with and without breast cancer.
However, despite numerous studies into the detection of circulating
biomarkers in breast cancer patients (9,10,12,13),
the molecular mechanism(s) by which these circulating factors
present in breast cancer may contribute to, and influence
angiogenesis, are still poorly understood, suggesting a complex
mechanism of interactions. Therefore, the aim of the present study
was to assess the ability of circulating factors, present in the
plasma from women with breast cancer, to influence tubule formation
in an in vitro system (14)
and to identify key factors responsible for increased
angiogenesis.
Materials and methods
Materials
All chemicals and cell culture reagents were
purchased from Sigma-Aldrich Ltd. (Dorset, UK), unless otherwise
stated.
Cell culture
Primary human umbilical vein endothelial cells
(HUVEC) and primary normal human dermal fibroblasts (NHDF) were
obtained from Clonetics (Lonza, Slough, UK). HUVEC were maintained
in endothelial basal medium (EBM-2) supplemented with EGM-2
SingleQuots (i.e., EGM-2 medium) whilst NHDF were maintained in
fibroblast basal medium (FBM) supplemented with FGM-2 SingleQuots;
all from Clonetics; Lonza (i.e., FGM-2 medium).
The co-culture system was set up using HUVEC
(between passages 1–8) and NHDF (between passages 1–12) following
the protocol established by Bishop et al (15) and adapted by Barron et al
(14). HUVEC and NHDF were mixed
and seeded in 24-well plates (Thermo Fisher Scientific Nunc,
Loughborough, UK) in EGM-2 medium. Co-cultured cells were incubated
for up to 14 days at 37°C in a 5% CO2 in air humidified
incubator.
Study subjects
Participants in the study were recruited through the
Breast Clinic in Aberdeen Royal Infirmary, Scotland. The study was
approved by the Grampian Ethics Committee. Signed informed consent
was obtained from each study participant. Venous blood from 40
volunteers was used in this study: 20 women with breast cancer and
20 age- matched control individuals. The women with breast cancer
had a life-time risk estimated to be double that of the normal
female population, based on their history, and screen of no known
genetic basis for the development of their breast cancer (e.g.,
BRCA1 or BRCA2 mutations) (16). The control group included
individuals with the normal population risk of breast cancer who
did not have breast cancer but were undergoing other forms of
breast surgery (16).
Plasma isolation
Plasma was isolated from venous blood from all of
the participants using a Histopaque gradient (16). Briefly, 20 ml of blood was layered
on top of 20 ml Histopaque-1077 and centrifuged at 800 × g for 30
min at room temperature. The top layer containing the plasma was
collected and stored at −80°C until required.
Plasma samples (n=40) were assayed in duplicate to
measure the concentration of VEGF, TNFα, IL-6 and leptin (DuoSet
ELISA cat. no. DY293B, DY210, DY206 and DY398-05, respectively;
R&D Systems, Abingdon, UK) according to the manufacturers
instructions. These assays have a sensitivity of ~30, 15, 9 and 20
pg/ml, respectively, and quantify both the natural and recombinant
forms of each protein.
Co-culture treatment with human plasma
and combinations of cytokines
The co-culture system was incubated with EGM-2
medium to initiate tubule formation and, 7 days after co-culture
establishment, the medium was changed to EGM-2ӨFBS [i.e., EGM-2
medium minus VEGF SingleQuot and 2% (v/v) fetal bovine serum (FBS)
SingleQuot] or EGM-2ӨFBS supplemented with 2% plasma from a random
subset of women with breast cancer (n=8) or control individuals
(n=8). The medium was collected every 4 days and replaced with
respective treatments.
In order to examine the effects of inflammatory
cytokines, the co-culture system was incubated with EGM-2 medium to
initiate tubule formation and, 7 days after the co-culture
establishment, medium was changed to EGM-2+VEGF, EGM-2Ө,
EGM-2Ө+Leptin, or cytokine combinations: EGM-2Ө+TNFα+IL-6,
EGM-2Ө+TNFα+Leptin or EGM-2Ө+ IL-6+Leptin. The medium codes are
summarised in Table I. The medium
was collected every 4 days and was replaced with respective
treatments.
 | Table I.Details of medium employed for
co-culture treatments. |
Table I.
Details of medium employed for
co-culture treatments.
| Code | Details |
|---|
| EBM-2 | Endothelial basal
medium |
| EGM-2 | EBM-2 supplemented
with EGM-2 SingleQuots |
| EGM-2Ө | EGM-2 medium minus
VEGF SingleQuot |
| EBM-2ӨFBS | EGM-2 medium minus
VEGF and FBS SingleQuots |
| EGM-2Ө+Leptin | EGM-2Ө supplemented
with leptin |
| EGM-2+VEGF | EGM-2 supplemented
with VEGF |
|
EGM-2Ө+TNFα+IL-6 | EGM-2Ө supplemented
with TNFα and IL-6 |
|
EGM-2Ө+TNFα+Leptin | EGM-2Ө supplemented
with TNFα and leptin |
|
EGM-2Ө+IL-6+Leptin | EGM-2Ө supplemented
with IL-6 and leptin |
Immunostaining
After 14 days, co-cultures were fixed at room
temperature with ice-cold 70% (v/v) ethanol and stained with
myeloma cell adhesion molecule [MCAM (CD146); specific to HUVEC] to
assess the extent of tubule formation as previously described by
Barron et al (14).
Co-cultures were viewed using an Olympus 1XS1 microscope with
Olympus TL4 Light Box Infinity Capture Application version 5.0.0
(Lumenera Corp., Nepean, ON, Canada) with camera model Infinity
2–2c was used for image capture. Three images per well were
captured and saved as BMP images of 1616 × 1216 pixels.
Illumination was set to give a best contrast between stained
tubules and unstained fibroblasts. Tubule formation was defined by
total tubule length and quantified using AngioSys®
software version 1.0 (TCS Cellworks Ltd., Buckingham, UK).
Proteome Profiler™ Human Angiogenesis
Antibody array
The relative expression of 55 angiogenesis-related
proteins was determined in human plasma (4 women per group) using a
Proteome Profiler™ Human Angiogenesis Antibody Array according to
the manufacturers instructions (cat. no. ARY007; R&D Systems).
Briefly, after a 1-h membrane blocking step, human plasma (200 µl)
was pre-incubated with a cocktail of biotinylated detection
antibodies (15 µl) and added to the membrane before incubating
overnight at 4°C. After a series of washes, the membrane was
incubated with streptavidin-horseradish peroxidase (HRP; 2 ml) for
30 min, before chemiluminescence detection reagents were added in
equal volumes for 1 min. The signal was detected by exposing the
membrane to CL-XPosure X-ray film (Thermo Fisher Scientific) over a
variety of exposure times (1, 3, 5 and 10 min). The light produced
at each spot is proportional to the amount of analyte bound and the
mean pixel density of the duplicate spots produced on the film was
determined using a FUSION FX7™ imaging instrument with Fusion 1 and
BIO-1D™ imaging software (PeqLab; VWR International Ltd.,
Lutterworth, UK).
In addition to the 55 angiogenesis-related proteins,
each membrane contained three pairs of positive reference spots and
one pair of negative control spots. Following subtraction of the
mean pixel density of the negative control spots from all values,
the level of each protein was expressed as a ratio relative to the
mean of the positive reference spots which were assigned a value of
1. Relative expression ratios for all 55 angiogenesis-related
proteins were compared between plasma from women with breast cancer
(n=3) and plasma from control individuals (n=4).
Statistical analysis
Unless otherwise stated, all data are expressed as
mean ± SEM of three independent experiments from different cell
passage numbers. Statistical differences were determined by one-way
analysis of variance (ANOVA) with Dunnetts multiple comparison
t-test (GraphPad Prism). P<0.05 was considered to be
statistically significant.
Results
Effect of human plasma on tubule
formation
The effect of human plasma on tubule formation was
tested with EGM-2ӨFBS: human plasma replaced FBS (2% v/v) present
in EGM-2 medium minus VEGF. Co-cultures were initially incubated in
EGM-2 medium (day 0) and, after co-culture establishment (day 7):
EGM-2ӨFBS or EGM-2ӨFBS supplemented with either human plasma from
women with breast cancer (cancer group, n=8) or control (control
group, n=8) individuals was added. These conditions were used to
assess the ability of angiogenesis-related growth factors present
in human plasma to modulate tubule formation once the angiogenesis
process had already started in our in vitro co-culture
model.
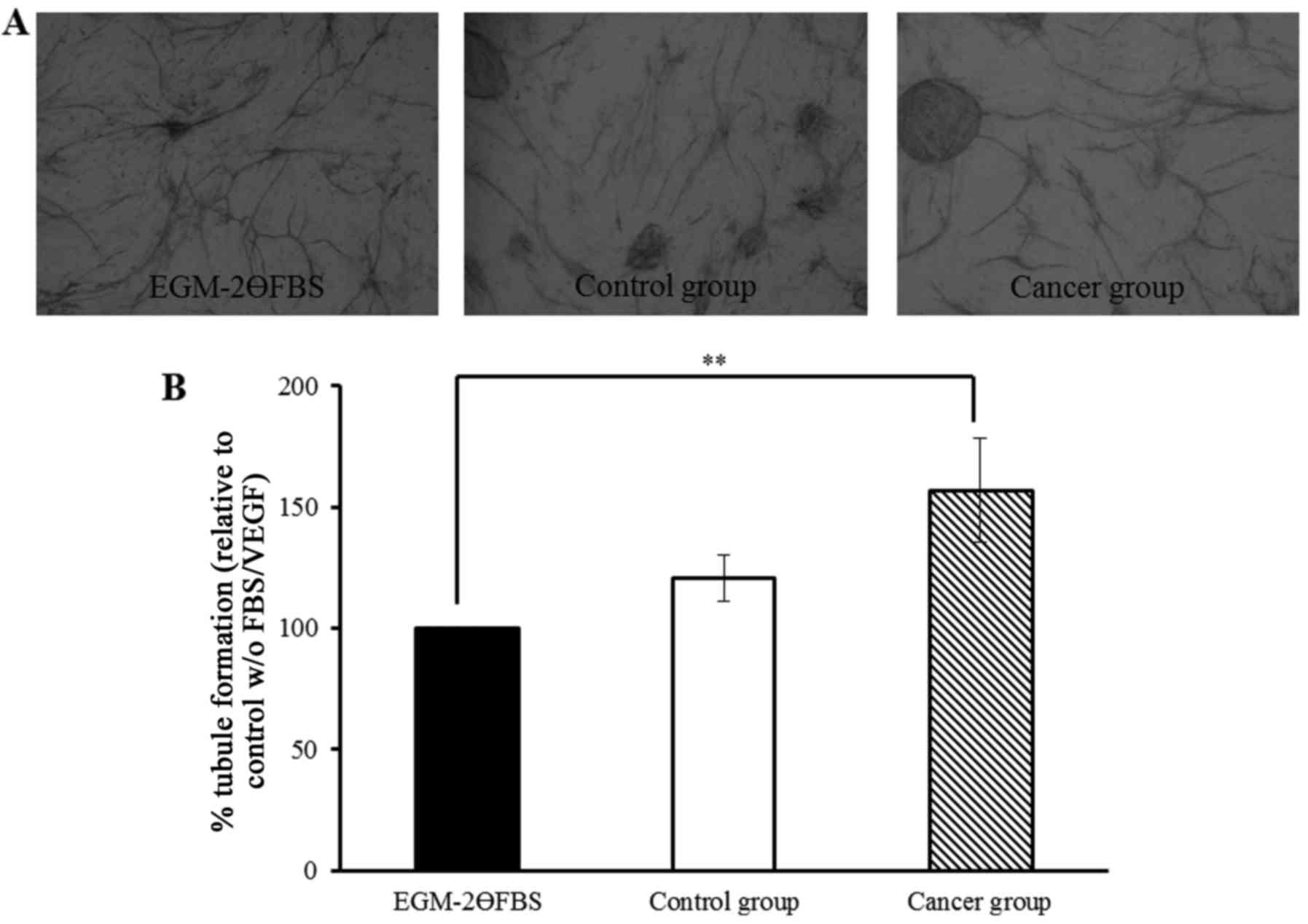
Tubules were clearly formed and visualised after 14
days (Fig. 1A) and similar results
were obtained in each experiment. The extent of tubule formation
was quantified by measuring total tubule length and the percentage
change was normalised to EGM-2ӨFBS (Fig. 1B). Tubule formation was
significantly increased by 57% (P<0.01) when plasma from women
with breast cancer was used in the co-culture system compared to
EGM-2ӨFBS. Although tubule formation was increased (by 21%) with
plasma from control individuals; no significant difference was
found compared to EGM-2ӨFBS.
Levels of VEGF, TNFα, IL-6 and leptin
in human plasma
Higher levels of VEGF (~6-fold increase, P=0.053),
TNFα (~3-fold increase) and IL-6 (~3.5-fold increase) were present
in plasma samples of the breast cancer group compared to the
control group (n=20 in each group), but differences were not
statistically significant (Table
II) (P>0.05). No changes in leptin levels were observed
between groups (Table II).
| Table II.Baseline characteristics of control
and cancer groups. |
Table II.
Baseline characteristics of control
and cancer groups.
|
Characteristics | Control group
(n=20) | Cancer group
(n=20) |
|---|
| Age (years) |
41.9±1.3 |
41.1±1.2 |
| VEGF (pg/ml) |
23.7±12.2 |
142.3±58.2 |
| TNFα (pg/ml) |
159.9±90.6 |
491.4±225.8 |
| IL-6 (pg/ml) |
43.1±31.2 |
153.1±79.5 |
| Leptin (ng/ml) |
15.9±1.5 |
17.8±1.7 |
Effect of the adipokine leptin and of
a combination of inflammatory cytokines/adipokine on tubule
formation
We have previously shown that angiogenic factors and
inflammatory markers such as VEGF, TNFα and IL-6 play an important
role in modulating angiogenesis (14); however, their effects on modulating
angiogenesis when used in combination with the adipokine leptin or
other inflammatory cytokines have not been previously reported.
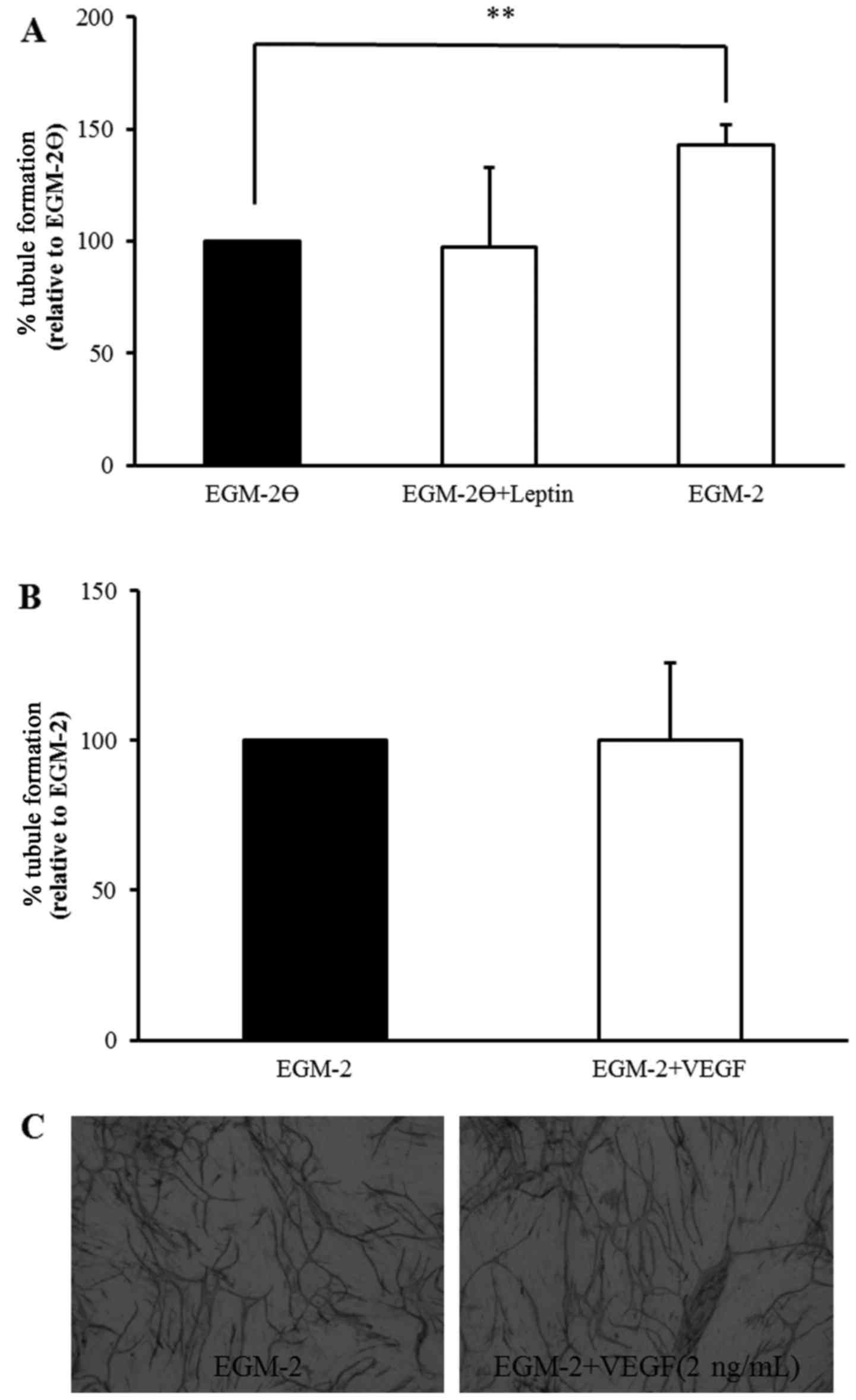
The effect of leptin on tubule formation was first
examined in the absence of VEGF (i.e., EGM-2Ө medium) in a similar
manner to Barron et al (14). Tubule formation was quantified by
measuring total tubule length and the percentage change was
normalised to EGM-2Ө (Fig. 2A).
Incubating co-cultures with leptin (10 nM, added on day 7;
EGM-2Ө+leptin) did not alter tubule formation compared to EGM-2Ө.
Whereas, tubule formation was significantly increased (43%) when
complete EGM-2 medium was added at day 7 compared to EGM-2Ө,
confirming the ability of the co-culture system to further produce
tubules with addition of VEGF. However, addition of VEGF (2 ng/ml;
EGM-2+VEGF) to complete EGM-2 medium after 7 days did not induce
further tubule formation (Fig. 2B),
suggesting an inability of the in vitro system to create
more new tubules than were normally formed with complete EGM-2
medium as clearly visualised after staining with MCAM on day 14
(Fig. 2C).
Further studies were undertaken to determine the
effect of cytokine/adipokine combinations on tubule formation.
Co-cultures were initially incubated with EGM-2 medium (day 0) and,
after co-culture establishment (day 7), EGM-2Ө, EGM-2Ө+TNFα+IL-6,
EGM-2Ө+TNFα+Leptin or EGM-2Ө+IL-6+Leptin were added. As combination
studies in which cytokines and adipokines are tested together in
equal or different concentrations have not been previously carried
out, these studies, which use an in vitro system reflecting
an in vivo situation where cytokines and adipokines do not
exist alone, are therefore very important to identify factors
responsible for angiogenesis.
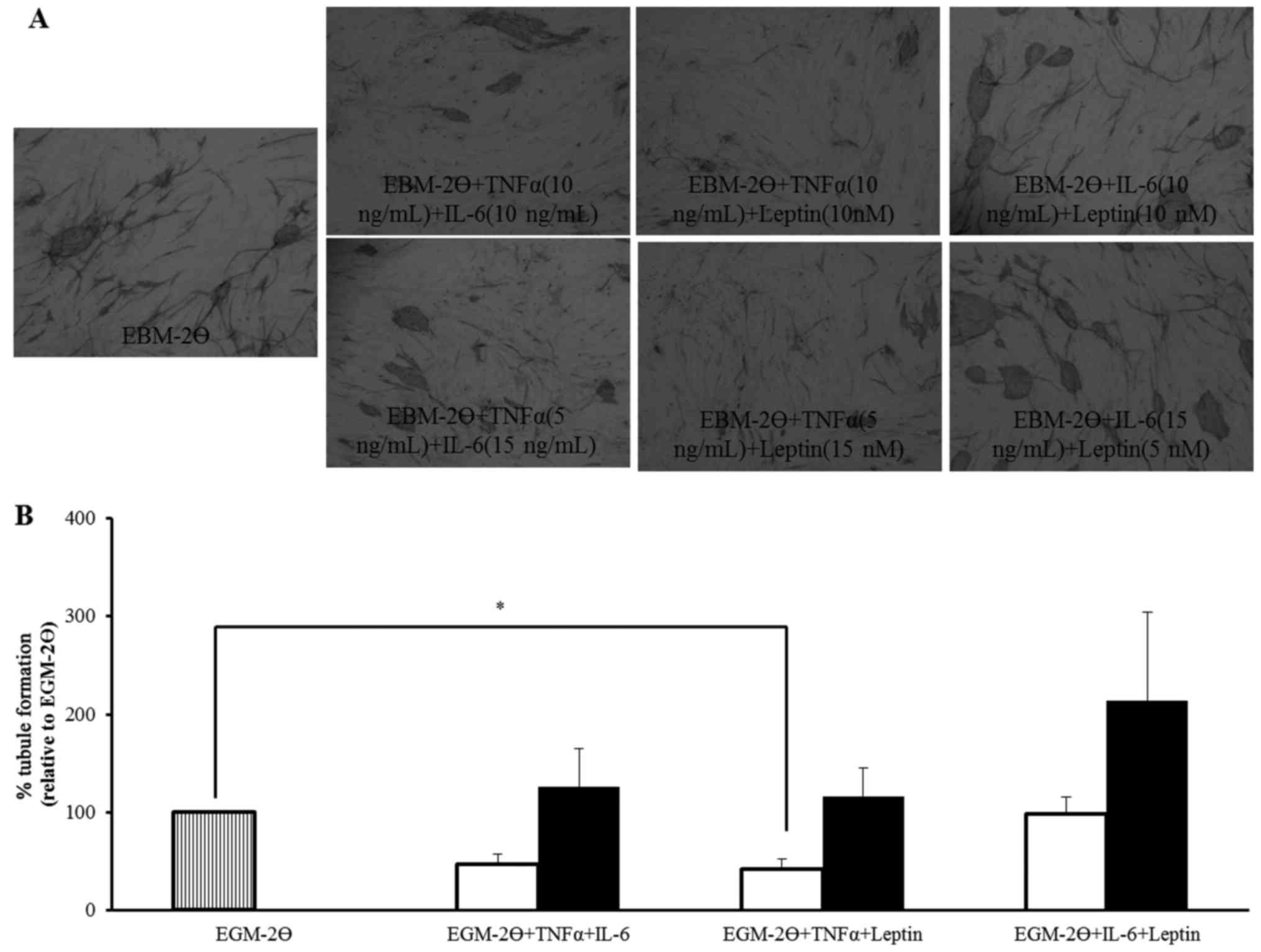
Tubules were clearly formed and visualised after 14
days (Fig. 3A) and similar results
were obtained in each experiment. The extent of tubule formation
was quantified by measuring total tubule length and the percentage
change was normalised to EGM-2Ө (Fig.
3B). TNFα (10 ng/ml) combined with IL-6 (10 ng/ml) or TNFα (10
ng/ml) combined with leptin (10 nM) (P<0.05), reduced tubule
formation by 53 and 58%, respectively. IL-6 (10 ng/ml) combined
with leptin (10 nM) did not alter tubule formation compared to
EGM-2Ө but, tubule formation was increased when a higher
concentration of IL-6 (15 ng/ml, increased by 26%) or leptin (15
nM, increased by 16%) were combined with TNFα (5 ng/ml) but, these
changes were not statistically significant when compared to EGM-2Ө.
A 114% increase in tubule formation was observed with IL-6 (15
ng/ml) combined with leptin (5 nM) but again, this was not
statistically significant when compared to EGM-2Ө. No statistically
significant differences were observed between the equivalent (white
bars) and different (black bars) concentrations of each combination
(Fig. 3B). These results indicate
that TNFα, IL-6 and leptin may differentially modulate tubule
formation either added individually or in combination, but the
limitation of the co-culture system does not allow this or,
alternatively, additional key factors are required.
Comparison of angiogenesis-related
protein expression between plasma from women with breast cancer and
control group
Although higher levels of VEGF, TNFα, IL-6 or leptin
were observed in the breast cancer group (Table II), the addition of exogenous VEGF,
TNFα, IL-6 or leptin alone (Fig. 2)
and Barron et al (14) or in
combination (Fig. 3) showed that
these molecules are not solely responsible for the increase in
tubule formation observed in the breast cancer group (Fig. 1). Proteome profiler human
angiogenesis antibody arrays were, therefore, used to identify
differences in protein expression that could be responsible for the
increase in tubule formation. Angiogenesis protein profiles in
plasma from women with breast cancer (n=3) were compared to
profiles from control individuals (n=4) (plasma from one breast
cancer individual was not acceptable for protein array analysis and
was excluded).
Levels of proteins expressed in plasma from the
control group were compared to those in plasma from women with
breast cancer, irrespective of whether the plasma sample increased,
or not, tubule formation in the in vitro co-culture system.
A cut-off of 1.5-fold increase difference and a cut-off of 0.5-fold
decrease difference was used as considered threshold for meaningful
physiological changes in protein arrays studies. Thirty-seven
proteins in the array showed no change in expression relative to
the positive reference spots (value of 1) and were not further
examined (Table III). Eighteen
angiogenic-related factors demonstrated a level of expression that
was 1.5-fold different in either plasma obtained from control or
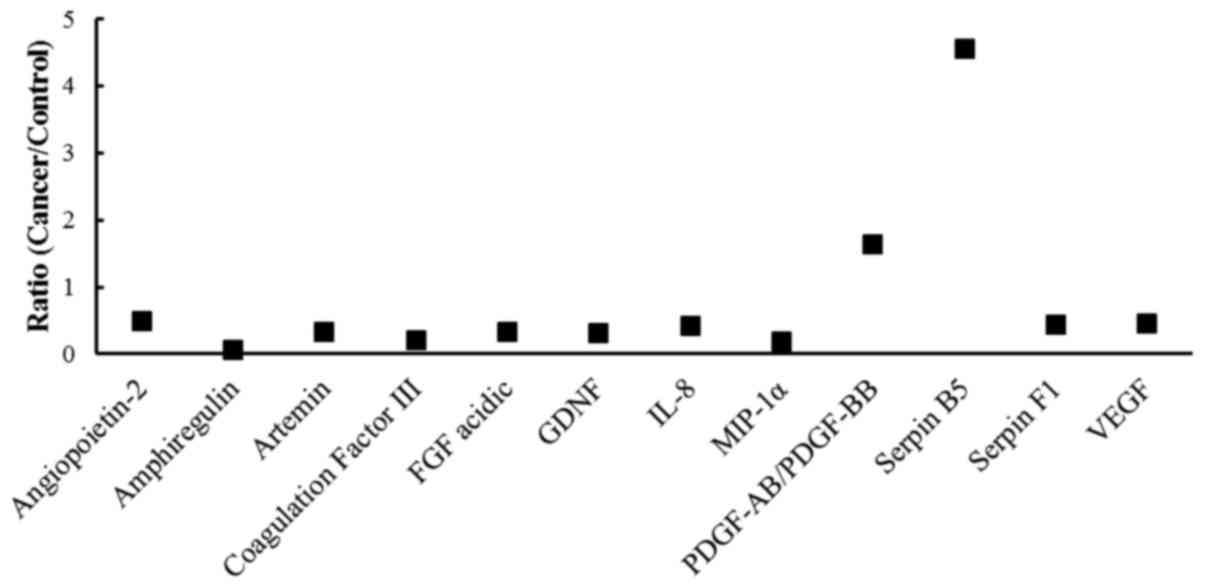
women with cancer. For these 18 proteins, cancer-to-control ratios
were calculated. Overall, 6 proteins were excluded that had ratios
between 0.5–1.5 (i.e. pro-angiogenic: FGF basic, persephin, uPA and
vasohibin; anti-angiogenic: serpin E1 and TIMP-1), 10 proteins
showed reduced expression (ratio <0.5) (pro-angiogenic:
amphiregulin, artemin, coagulation factor III, FGF acidic, GDNF,
IL-8, MIP-1α and VEGF; anti-angiogenic: angiopoietin-2 and serpin
F1) (Fig. 4); whilst, 2 proteins
showed increased expression (ratio >1.5): PDGF-AB/PDGF-BB (ratio
of 1.64, pro-angiogenic) and serpin B5 (ratio of 4.56,
anti-angiogenic) (Fig. 4).
| Table III.Relative expression of angiogenic
factors which demonstrated no change in protein expression. |
Table III.
Relative expression of angiogenic
factors which demonstrated no change in protein expression.
| Angiogenic
factor | Control group
(n=4) | Cancer group
(n=3) |
|---|
| Activin A | 0.07 | 0.07 |
| ADAMTS-1 | 0.08 | 0.10 |
| Angiogenin | 1.14 | 1.23 |
| Angiopoietin-1 | 0.50 | 0.65 |
|
Angiostatin/plasminogen | 0.14 | 0.13 |
| CXCL16 | 1.01 | 1.08 |
| DPPIV | 1.07 | 1.04 |
| EGF | 0.14 | 0.12 |
| EG-VEGF | 0.19 | 0.15 |
| Endoglin | 0.97 | 0.89 |
| Endostatin/collagen
XVIII | 1.05 | 1.15 |
| Endothelin-1 | 0.54 | 0.67 |
| FGF-4 | 0.10 | 0.09 |
| FGF-7 | 0.06 | 0.07 |
| GM-CSF | 0.12 | 0.09 |
| HB-EGF | 0.19 | 0.16 |
| HGF | 0.14 | 0.12 |
| IGFBP-1 | 1.01 | 1.15 |
| IGFBP-2 | 1.03 | 1.15 |
| IGFBP-3 | 1.02 | 1.12 |
| IL-1β | 0.09 | 0.07 |
| LAP (TGF-β1) | 0.10 | 0.08 |
| Leptin | 1.00 | 1.07 |
| MCP-1 | 0.06 | 0.06 |
| MMP-8 | 0.97 | 1.06 |
| MMP-9 | 1.03 | 1.04 |
| NRG1-β1 | 0.15 | 0.15 |
| Pentraxin 3
(PTX3) | 0.45 | 0.59 |
| PD-ECGF | 0.21 | 0.22 |
| PDGF-AA | 0.60 | 0.78 |
| Platelet factor 4
(PF4) | 1.02 | 1.09 |
| PIGF | 0.11 | 0.11 |
| Prolactin | 0.86 | 0.99 |
| TIMP-4 | 1.07 | 0.73 |
|
Thrombospondin-1 | 1.05 | 0.73 |
|
Thrombospondin-2 | 0.05 | 0.05 |
| VEGF-C | 0.04 | 0.03 |
Furthermore, plasma samples, from both the normal
and cancer groups, which induced an increase in tubule formation in
the in vitro co-culture model, were compared to identify
which angiogenesis-related proteins could be responsible for tubule
formation. For array analysis, a cut-off of 1.5-fold difference was
set as before. In the normal group, 42 out of the 55
angiogenesis-related proteins showed no change in protein
expression and were not further examined. Thirteen of the 55
angiogenic-related factors had levels of expression that were
1.5-fold different. In relation to the expression of these 13
proteins, further analysis was carried out by comparing their
expression and calculating a ratio between the same proteins in
plasma samples which induced an increase-to-no change in tubule
formation when compared to EGM-2ӨFBS (Fig. 1). After exclusion of 6 proteins that
had ratios between 0.5–1.5 (i.e., pro-angiogenic: angiopoietin-1,
artemin, IL-8, PDGF-AA, uPA and VEGF-C), 1 angiogenesis-related
protein (i.e. PDGF-AB/PDGF-BB, pro-angiogenic) showed reduced
expression (ratio <0.5) whereas 6 angiogenesis-related proteins
(i.e. pro-angiogenic: FGF acidic, GDNF, GM-CSF and MIP-1α;
anti-angiogenic: activin A and thrombospondin-2), had increased
expression (ratio >1.5) (Table
IV).
| Table IV.Relative expression of angiogenic
factors which demonstrated an increase in tubule formation when
plasma from both the control and breast cancer groups was used in
the in vitro co-culture system. |
Table IV.
Relative expression of angiogenic
factors which demonstrated an increase in tubule formation when
plasma from both the control and breast cancer groups was used in
the in vitro co-culture system.
| Angiogenic
factor | Increase in tubule
formation Control group (n=2) | Increase in tubule
formation Cancer group (n=2) |
|---|
| Activin A | 2.28c | 0.36b |
| ADAMTS-1 | – | 0.60a |
| Angiopoietin-1 | 0.64a | 3.54c |
|
Angiostatin/plasminogen | – | 3.10c |
| Artemin | 1.45a | 3.23c |
| EGF | – | 0.68a |
| EG-VEGF | – | 0.64a |
| Endoglin | – | 0.61a |
| FGF-4 | – | 2.02c |
| FGF acidic | 1.78c | 49.39c |
| GDNF | 2.07c | – |
| GM-CSF | 1.56c | 0.48b |
| HGF | – | 0.62a |
| IL-1β | – | 2.22c |
| IL-8 | 0.61a | 1.55c |
| LAP (TGF-β1) | – | 1.49a |
| MIP-1α | 1.54c | – |
| Pentraxin 3
(PTX3) | – | 0.43b |
| PD-ECGF | – | 0.61a |
| PDGF-AA | 0.61a | 2.54c |
|
PDGF-AB/PDGF-BB | 0.37b | 4.31c |
| Persephin | – | 2.02c |
| PIGF | – | 1.63c |
| Serpin B5 | – | 30.86c |
| TIMP-1 | – | 0.01b |
| TIMP-4 | – | 0.04b |
|
Thrombospondin-2 | 8.12c | – |
| uPA | 0.61a | 0.11b |
| VEGF | – | 0.24b |
| VEGF-C | 0.57a | 1.54c |
Conversely, in the cancer group, 28 of the 55
angiogenesis-related proteins showed no change in protein
expression and were not examined further. Twenty-seven of the 55
angiogenesis-related factors identified had levels of expression
that were 1.5-fold different (Table
IV) and, of these 27 remaining angiogenesis-related proteins,
the increase in tubule formation-to-no change in tubule formation
ratios was calculated. Overall, after exclusion of 7
angiogenesis-related proteins that had ratios between 0.5–1.5 [i.e.
pro-angiogenic: EGF, EG-VEGF, endoglin, HGF, LAP (TGF-β1), and
PD-ECGF; anti-angiogenic: ADAMTS-1], 20 showed differences in
levels: the level of expression was reduced (ratio <0.5) for 7
proteins (i.e. pro-angiogenic: GM-CSF, pentraxin 3, uPA and VEGF;
anti-angiogenic: activin A, TIMP-1 and TIMP-4) whereas 13 protein
levels were increased (ratio >1.5) (i.e. pro-angiogenic:
angiopoietin-1, artemin, FGF acidic, FGF-4, IL-1β, IL-8, PDGF-AA,
PDGF-AB/PGDF-BB, persephin, PIGF and VEGF-C; anti-angiogenic:
angiostatin/plasminogen, and serpin B5) in the cancer group
(Table IV).
Discussion
Angiogenesis is one of the major causes for cancer
metastasis. The formation of new blood vessels is essential for
cancer growth and metastasis; therefore, the development of
angiogenic characteristics is vital in a number of tumour types
including breast cancer (17,18).
Many studies have measured the production of angiogenesis-related
factors in serum from women with breast cancer via ELISA
analysis; however, the majority of these have only examined one or
two angiogenesis-related factors, including VEGF (6,8,19,20),
basic FGF (6), IL-6 (21), leptin and prolactin (8) and thrombospondin-1 (19). To date, there have only been three
studies which have looked at the expression of multiple angiogenic
factors in serum from women with breast cancer (10,12,22).
The present study evaluated the effect of factors
present in plasma obtained from women with breast cancer compared
to age-matched control women in relation to tubule formation, using
a versatile in vitro co-culture model established in our
laboratory (14) in addition to
testing the effect of cytokines/adipokines added singularly or in
combination to the system. Finally it examined the relative
expression of 55 angiogenesis-related proteins in a subset of
plasma samples from cancer and control individuals in relation to
tubule formation. This study therefore provides additional
information on proteins differentially expressed and on the
importance of a balance between pro-angiogenic/anti-angiogenic
factors in tubule formation.
The in vitro co-culture system used to study
tubule formation has been optimised so that molecules of interest
could be added 7 days after initiation of tubule formation to make
it relevant when considering breast tumour progression and
metastasis. By using plasma samples, it was found that tubule
formation in vitro was significantly increased (P<0.01)
after incubation with plasma from women with breast cancer (n=8).
Indeed, a higher level of VEGF was present in the breast cancer
group compared to the control group. However, not all individual
plasma samples increased tubule formation, which could be due to
low/absent levels of VEGF but also, to higher levels of the
anti-angiogenic growth factor, TNFα. As inflammatory cytokines such
as TNFα (23) and IL-6 (unpublished
data) and, the adipokine leptin have been shown, in vitro,
to play a putative role in breast cancer progression (24), it is important to investigate the
effect that these exogenous cytokines/adipokines might have on
angiogenesis in isolation, but also to study their effect when
present in combination as these growth factors do not exist in
isolation in human plasma. Using our in vitro co-culture
cell model, it was found that leptin did not alter tubule formation
when VEGF was lacking but, addition of complete EGM-2 medium
supplemented with VEGF did further increase tubule formation. These
results are in contrast with the pro-angiogenic effect of leptin
(10, 100 and 1000 ng/ml) on endothelial tube formation previously
described in single-cell conditions using HUVECs and a matrigel
matrix (25–27) or, HUVECs and a collagen I matrix
(28); suggesting that the system
used for studying angiogenesis in vitro might be critical
even if the use of in vitro co-culture systems are more
closely related to in vivo conditions.
This study is also the first to use an in
vitro endothelial cell/fibroblast co-culture cell system to
investigate the effect of combining exogenous cytokines/adipokines.
Results from this study revealed that when TNFα was combined with
either IL-6 or leptin, tubule formation was reduced but, not to the
same extent as TNFα alone as it stopped tubule formation (14); therefore, pro-angiogenic IL-6 and
leptin may counteract the anti-angiogenic effect of TNFα, but not
completely. An increase in tubule formation was observed when IL-6
was combined with leptin. These observations further indicate that
angiogenesis is modulated by specific pro- and anti-angiogenic
inflammatory markers. More specifically, TNFα, IL-6 and leptin
differentially affect tubule formation either individually or in
combination. Thus, other angiogenesis-related proteins may be
involved. Since angiogenesis is a complex process that involves
many proteins; angiogenesis protein arrays were used to assess the
expression of a large group of proteins. Several
angiogenesis-related proteins (both pro- and anti-angiogenic) were
observed to have altered expression in the plasma from women with
breast cancer which may correlate with the increase in tubule
formation and therefore, may be involved in breast tumour
progression and metastasis. Accordingly, angiogenesis in breast
cancer is regulated by a net balance between pro- and
anti-angiogenic proteins. In the present study, 12 out of 55
angiogenesis-related proteins analysed had altered expression in
the breast cancer group compared to the control group; with 10
proteins observed to have reduced expression (pro-angiogenic:
amphiregulin, artemin, coagulation factor III, FGF acidic, GDNF,
IL-8, MIP-1α and VEGF; anti-angiogenic: angiopoietin-2 and serpin
F1); whilst, 2 proteins showed increased expression:
PDGF-AB/PDGF-BB (ratio of 1.64, pro-angiogenic) and serpin B5
(ratio of 4.56, anti-angiogenic). To the authors knowledge, this is
the first study to directly examine angiogenesis-related protein
expression and tubule formation via a co-culture cell system
in vitro using human plasma samples. However, a study by
Morelli et al (19),
investigated the in vitro effect of sera from patients with
breast or gastrointestinal cancers on HUVEC proliferation using a
colourimetric assay. Morelli et al (19) observed that sera from 19% (15 of 78)
of breast cancer patients induced growth of HUVECs whilst, sera
from 5% (4 of 78) of breast cancer patients inhibited endothelial
cell proliferation. Furthermore, when sera stimulated endothelial
growth, it was correlated with high levels of VEGF detected by
ELISA whilst, when sera inhibited endothelial cell growth, it was
correlated with high levels of soluble thrombospondin revealed by
western blot analysis (19). These
results show that individual variations of angiogenesis-related
proteins are evident in specific cohorts of study volunteers and
could depend on disease stage, subtype and treatment. In relation
to this, we recognise a limitation of this study to analyse the
effects on tubule formation in the co-culture stystem in function
of disease stage and subtype: no treatment differences were present
as samples were collected just after diagnosis of breast cancer and
before treatment commenced.
The present study focused on the identification of
factors which drive angiogenesis, however, it also highlighted the
importance of the fine balance between pro- and anti-angiogenic
factors present in cancer patients plasma and driving angiogenesis.
To date, anti-angiogenic drugs for cancer treatment have been of
relative success and have predominantly targeted the
expression/binding of pro-angiogenic factors such as VEGF (29). Our data emphasise the need for
development of drug therapies which target, at the same time, not
only the expression of pro-angiogenic factors but also affect the
expression of some anti-angiogenic factors as it is the result of
their differential expression which affects angiogenesis and cell
proliferation.
In conclusion, many angiogenesis-related proteins
may be involved in the growth, spread and progression of breast
tumours. This study identified potential key factors which might be
responsible to drive angiogenesis in breast cancer and, once their
role has been defined further, they might provide targets for
future clinical therapies. The identified factors have been shown
to be relevant in other malignancies including gastrointestinal
(19) and laryngeal cancer
(30), confirming their potential
importance in breast cancer.
Acknowledgements
The authors would like to thank Tenovus Scotland and
Robert Gordon University (Faculty of Health and Social Care) for
financial support. The authors would like to thank Professor S.D.
Heys from the University of Aberdeen for facilitating collection of
the volunteers samples.
Glossary
Abbreviations
Abbreviations:
|
EC
|
endothelial cells
|
|
HUVEC
|
human umbilical vein endothelial
cells
|
|
IL-6
|
interleukin-6
|
|
MMP
|
matrix metalloproteinases
|
|
NHDF
|
normal human dermal fibroblasts
|
|
TNFα
|
tumour necrosis factor-α
|
|
VEGF
|
vascular endothelial growth factor
|
References
|
1
|
DeSantis CE, Lin CC, Mariotto AB, Siegel
RL, Stein KD, Kramer JL, Alteri R, Robbins AS and Jemal A: Cancer
treatment and survivorship statistics, 2014. CA Cancer J Clin.
64:252–271. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
DeBruin LS and Josephy PD: Perspectives on
the chemical etiology of breast cancer. Environ Health Perspect.
110:(Suppl 1). 119–128. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Weigelt B, Peterse JL and van t Veer LJ:
Breast cancer metastasis: Markers and models. Nat Rev Cancer.
5:591–602. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
4
|
Taneja P, Maglic D, Kai F, Zhu S, Kendig
RD, Fry EA and Inoue K: Classical and novel prognostic markers for
breast cancer and their clinical significance. Clin Med Insights
Oncol. 4:15–34. 2010.PubMed/NCBI
|
|
5
|
Yersal O and Barutca S: Biological
subtypes of breast cancer: Prognostic and therapeutic implications.
World J Clin Oncol. 5:412–424. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Dirix LY, Vermeulen PB, Pawinski A, Prové
A, Benoy I, De Pooter C, Martin M and Van Oosterom AT: Elevated
levels of the angiogenic cytokines basic fibroblast growth factor
and vascular endothelial growth factor in sera of cancer patients.
Br J Cancer. 76:238–243. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Relf M, LeJeune S, Scott PA, Fox S, Smith
K, Leek R, Moghaddam A, Whitehouse R, Bicknell R and Harris AL:
Expression of the angiogenic factors vascular endothelial cell
growth factor, acidic and basic fibroblast growth factor, tumor
growth factor β-1, platelet-derived endothelial cell growth factor,
placenta growth factor, and pleiotrophin in human primary breast
cancer and its relation to angiogenesis. Cancer Res. 57:963–969.
1997.PubMed/NCBI
|
|
8
|
Coskun U, Günel N, Toruner FB, Sancak B,
Onuk E, Bayram O, Cengiz O, Yilmaz E, Elbeg S and Ozkan S: Serum
leptin, prolactin and vascular endothelial growth factor (VEGF)
levels in patients with breast cancer. Neoplasma. 50:41–46.
2003.PubMed/NCBI
|
|
9
|
Chavey C, Bibeau F, Gourgou-Bourgade S,
Burlinchon S, Boissière F, Laune D, Roques S and Lazennec G:
Oestrogen receptor negative breast cancers exhibit high cytokine
content. Breast Cancer Res. 9:R15–R26. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Gonzalez RM, Daly DS, Tan R, Marks JR and
Zangar RC: Plasma biomarker profiles differ depending on breast
cancer subtype but RANTES is consistently increased. Cancer
Epidemiol Biomarkers Prev. 20:1543–1551. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Castañeda-Gill JM and Vishwanatha JK:
Antiangiogenic mechanisms and factors in breast cancer treatment. J
Carcinog. 15:1–39. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Li L, Chen L, Zhang W, Liao Y, Chen J, Shi
Y and Luo S: Serum cytokine profile in patients with breast cancer.
Cytokine. 89:173–178. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Rovati B, Mariucci S, Delfanti S, Grasso
D, Tinelli C, Torre C, De Amici M and Pedrazzoli P: Simultaneous
detection of circulating immunological parameters and tumor
biomarkers in early stage breast cancer patients during adjuvant
chemotherapy. Cell Oncol (Dordr). 39:211–228. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Barron GA, Bordet E, Goua M and Bermano G:
Modulation of angiogenesis by inflammatory markers and the role of
matrix metalloproteinases in an endothelial cell/fibroblast
co-culture system. Curr Angiogenes. 3:152–163. 2014. View Article : Google Scholar
|
|
15
|
Bishop ET, Bell GT, Bloor S, Broom IJ,
Hendry NF and Wheatley DN: An in vitro model of angiogenesis: Basic
features. Angiogenesis. 3:335–344. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Bermano G, Smyth E, Goua M, Heys SD and
Wahle KW: Impaired expression of glutathione peroxidase-4 gene in
peripheral blood mononuclear cells: A biomarker of increased breast
cancer risk. Cancer Biomark. 7:39–46. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Schneider BP and Miller KD: Angiogenesis
of breast cancer. J Clin Oncol. 23:1782–1790. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Carmeliet P and Jain RK: Molecular
mechanisms and clinical applications of angiogenesis. Nature.
473:298–307. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Morelli D, Lazzerini D, Cazzaniga S,
Squicciarini P, Bignami P, Maier JA, Sfondrini L, Ménard S,
Colnaghi MI and Balsari A: Evaluation of the balance between
angiogenic and antiangiogenic circulating factors in patients with
breast and gastrointestinal cancers. Clin Cancer Res. 4:1221–1225.
1998.PubMed/NCBI
|
|
20
|
Gisterek I, Matkowski R, Lacko A,
Sedlaczek P, Szewczyk K, Biecek P, Halon A, Staszek U, Szelachowska
J, Pudelko M, et al: Serum vascular endothelial growth factors a, C
and d in human breast tumors. Pathol Oncol Res. 16:337–344. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhang GJ and Adachi I: Serum interleukin-6
levels correlate to tumor progression and prognosis in metastatic
breast carcinoma. Anticancer Res. 19:(2B). 1427–1432.
1999.PubMed/NCBI
|
|
22
|
Georgiou GK, Igglezou M, Sainis I, Vareli
K, Batsis H, Briasoulis E and Fatouros M: Impact of breast cancer
surgery on angiogenesis circulating biomarkers: A prospective
longitudinal study. World J Surg Oncol. 11:213–221. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Weichhaus M, Broom I and Bermano G: The
molecular contribution of TNF-α in the link between obesity and
breast cancer. Oncol Rep. 25:477–483. 2011.PubMed/NCBI
|
|
24
|
Weichhaus M, Broom J, Wahle K and Bermano
G: Leptin inhibits proliferation of breast cancer cells at
supraphysiological concentrations by inhibiting mitogen-activated
protein kinase signaling. Oncol Lett. 8:374–378. 2014.PubMed/NCBI
|
|
25
|
Rodrigues S, Van Aken E, Van Bocxlaer S,
Attoub S, Nguyen QD, Bruyneel E, Westley BR, May FE, Thim L, Mareel
M, et al: Trefoil peptides as proangiogenic factors in vivo and in
vitro: Implication of cyclooxygenase-2 and EGF receptor signaling.
FASEB J. 17:7–16. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Garonna E, Botham KM, Birdsey GM, Randi
AM, Gonzalez-Perez RR and Wheeler-Jones CP: Vascular endothelial
growth factor receptor-2 couples cyclo-oxygenase-2 with
pro-angiogenic actions of leptin on human endothelial cells. PLoS
One. 6:e188232011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Dubois V, Delort L, Billard H, Vasson MP
and Caldefie-Chezet F: Breast cancer and obesity: In vitro
interferences between adipokines and proangiogenic features and/or
antitumor therapies? PLoS One. 8:e585412013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ferla R, Bonomi M, Otvos L Jr and Surmacz
E: Glioblastoma-derived leptin induces tube formation and growth of
endothelial cells: Comparison with VEGF effects. BMC Cancer.
11:303–314. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Nielsen DL, Andersson M, Andersen JL and
Kamby C: Antiangiogenic therapy for breast cancer. Breast Cancer
Res. 12:209–224. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
30
|
Korampalli TS, Green V, Greenman J and
Stafford ND: Protein profiling of angiogenesis-related growth
factors in laryngeal carcinoma: pattern of protein expression in
relation to tumour progression. Int J Oncol. 39:1033–1039.
2011.PubMed/NCBI
|