Introduction
Cervical cancer, one of the most common
gynecological malignant diseases, ranked fourth in all
cancer-related mortalities in women, accounting for ~270,000 deaths
in USA (1,2). Remarkable advances in the diagnosis
and treatment of cervical cancer have been achieved, the prognosis
of cervical cancer patients especially for those in advanced stage
remains poor (3,4). Unfortunately, the exact molecular
mechanisms underlying the initiation and progression of cervical
cancer remain largely unknown. Therefore, investigating the
mechanisms for the initiation and progression of cervical cancer
will promote the identification of novel biomarkers and treatment
targets, which is critical for improving the prognosis of cervical
cancer patients.
Previous studies showed that the initiation and
progression of cervical cancer is a complex process in which
numerous proteins and non-coding RNAs are involved (5,6).
MicroRNAs (miRNAs), a group of small non-coding RNAs, have been
found to play important roles in human cancers including cervical
cancer (7,8). Some miRNAs have been regarded as the
biomarkers and therapeutic targets for cervical cancer patients
(9). Among numerous cancer-related
microRNAs, miR-187 was recently found to be a novel target in human
cancers including liver (10),
breast (11), ovarian (12), prostate (13), non-small cell lung (14) and colorectal cancer (15,16).
It was found to play an oncogenic role in breast (11), ovarian (12) and non-small cell lung cancer
(14). On the other hand, miR-187
was also found to play tumor suppressive roles in colorectal cancer
(15,16), hepatocellular carcinoma (10) and prostate cancer (13). However, the expression and function
of miR-187 in GC remain uninvestigated.
In the present study, we found that miR-187 was
significantly decreased in cervical cancer tissues and cell lines.
This study also confirmed that decreased expression of miR-187 was
associated with poor clinicopathological features and poor survival
of cervical cancer patients. miR-187 inhibited the proliferation
and increased the apoptosis of cervical cancer cells. Furthermore,
we identified that FGF9 was a downstream target of miR-187 in
cervical cancers. Inhibiting FGF9 is critical for the functional
influence of miR-187 in cervical cancer cells.
Materials and methods
Patients and tissue specimens
The collection of clinical samples were approved by
the Research Ethics Committee of Renmin Hospital of Wuhan
University. All patients enrolled in this study were pathologically
confirmed as cervical cancer and were provided with written consent
and agreed to donate their tissue samples for research use. These
clinical samples were stored in liquid nitrogen before extracting
RNAs.
Cell culture and transfection
Cervical cancer cell lines including C33A, HeLa,
Caski and SiHa were cultured in Dulbecco's modified Eagle's medium
(DMEM; Invitrogen, Carlsbad, CA, USA) supplemented with fetal
bovine serum (10%, FBS; HyClone Laboratories, Logan, UT, USA) and
penicillin/streptomycin (1%; Invitrogen). Human normal cervical
epithelial cells (NCEC) were derived from healthy female cervical
tissue and were cultured in serum-free medium (Invitrogen) along
with EGF, bovine pituitary extract and penicillin/streptomycin (1%;
Invitrogen). All cells were maintained in cell incubator at 37°C
with 5% CO2. miR-187 mimic and inhibitor were obtained
from Shanghai GenePharma Co., Ltd. (Shanghai, China). FGF9 vector
and FGF9 siRNA were from Guangzhou Ruibo Biological Technolog, Co.,
Ltd. (Guangzhou, China). All these vectors were transfected into
cervical cancer cells using Lipofectamine 2000 (Invitrogen) based
on the provided protocols. The transfection efficiency of miR-187
mimic and inhibitor were confirmed by evaluating miR-187 level with
qRT-PCR. The transfection efficiency of FGF9 vector and siRNA were
confirmed by evaluating FGF9 level with western blot analysis.
Reverse transcriptase-quantitative PCR
(RT-qPCR)
The RNA in cervical cancer tissues and cell lines
was isolated with TRIzol reagent (Invitrogen) according to the
manufacturer's instructions. Reverse transcription of miRNA and
mRNA were performed with miScript II RT kit (Qiagen, Hilden,
Germany) and QuantScript RT kit (Tiangen Biotech, Co., Ltd.,
Beijing, China), respectively. SYBR-Green PCR kit (Qiagen) was used
for RT-PCR quantification. Relative miR-187 level and FGF mRNA
level were calculated using 2−ΔΔCt method after
normalization to U6 and GAPDH, respectively.
Western blot analysis
Cellular proteins from cervical cancer cells were
extracted with RIPA buffer. Isolated proteins were subjected to
electrophoresis on 4–20% SDS-PAGE gels and transferred to PVDF
membranes. After blocked with 5% non-fat milk/TBST (Tris-buffered
saline Tween-20), the membranes were incubated with the primary
antibody including GAPDH (1:2,000; Santa Cruz Biotechnology, Santa
Cruz, CA, USA), FGF9 (1:1,000; Santa Cruz Biotechnology) and
caspase-3 (1:1,000; Santa Cruz Biotechnology) at 4°C overnight and
incubated with HRP-conjugated secondary antibody at room
temperature for 2 h. The protein bands were detected and visualized
with the ECL reagent (Beyotime Institute of Biotechnology, Haimen,
China).
Cell viability and proliferation
assay
For viability assay, cervical cancer cells
transfected with corresponding vectors were seeded in 96-well
plates (5×103 cells/well). After cell seeding (0, 24, 48
and 72 h) MTT reagent was added to each well and the cells were
incubated at 37°C for 4 h. After removing the culture medium,
cervical cancer cells were solubilized in 150 µl dimethyl sulfoxide
(DMSO) and subjected to colorimetric analysis (wavelength, 490 nm).
For proliferation assay, cervical cancer cells transfected with
corresponding vectors were seeded in 6-well plates and maintained
in cell incubators for 2 weeks. Two weeks later, the cell colonies
were stained with crystal violet solution. The number of cell
colonies were counted and compared between groups.
Apoptosis assays
To evaluate the apoptosis of cervical cancer cells,
the flow cytometry assay was performed to evaluate the percentage
of apoptotic cells. In brief, cervical cancer cells were harvested
and re-suspended in phosphate-buffered saline (PBS), and then
stained with Annexin V detection kit, and subjected to FACScan
analysis. Additionally, cellular level of caspase-3 which is a
marker of cell apoptosis was evaluated with western blot
analysis.
Luciferase reporter assay
Cervical cancer cells seeded in 24-well plates were
transfected with 200 ng of miR-187 mimic or inhibitor or the
corresponding control vector along with 50 ng of wild-type (WT) or
mutant (MT) 3′-UTR of FGF9 mRNA. Forty-eight hours after the
transfection, these cells were collected and luciferase activity
was detected with a Dual-luciferase reporter assay system following
the manufacturer's instructions (Promega, Madison, WI, USA).
Mouse xenograft model
To investigate the in vivo influence of
miR-187 on cervical cancer cells, C33A cells transfected with
miR-187 mimic or control vector were subcutaneously inoculated into
nude mice. Twenty-eight days after the cell inoculation, the mice
were sacrificed and the formed subcutaneous tumors were subjected
to Ki-67 staining. Tumor volumes were calculated every 7 days based
on the following formula: length × width2/2. The
protocols for in vivo experiments were approved by the
Animal Research Protection Committee of Renmin Hospital of Wuhan
University.
Statistical analysis
Statistical analyses were performed using the
GraphPad and SPSS 15.0 software. Comparisons between the groups
were performed using the t-test and the χ2 test. Overall
survival and progression-free survival analysis were performed
using the Kaplan-Meier method for plotting and the log-rank test
for comparison. All differences were regarded as statistically
significant at the level of P<0.05.
Results
miR-187 is downregulated in human
cervical cancer
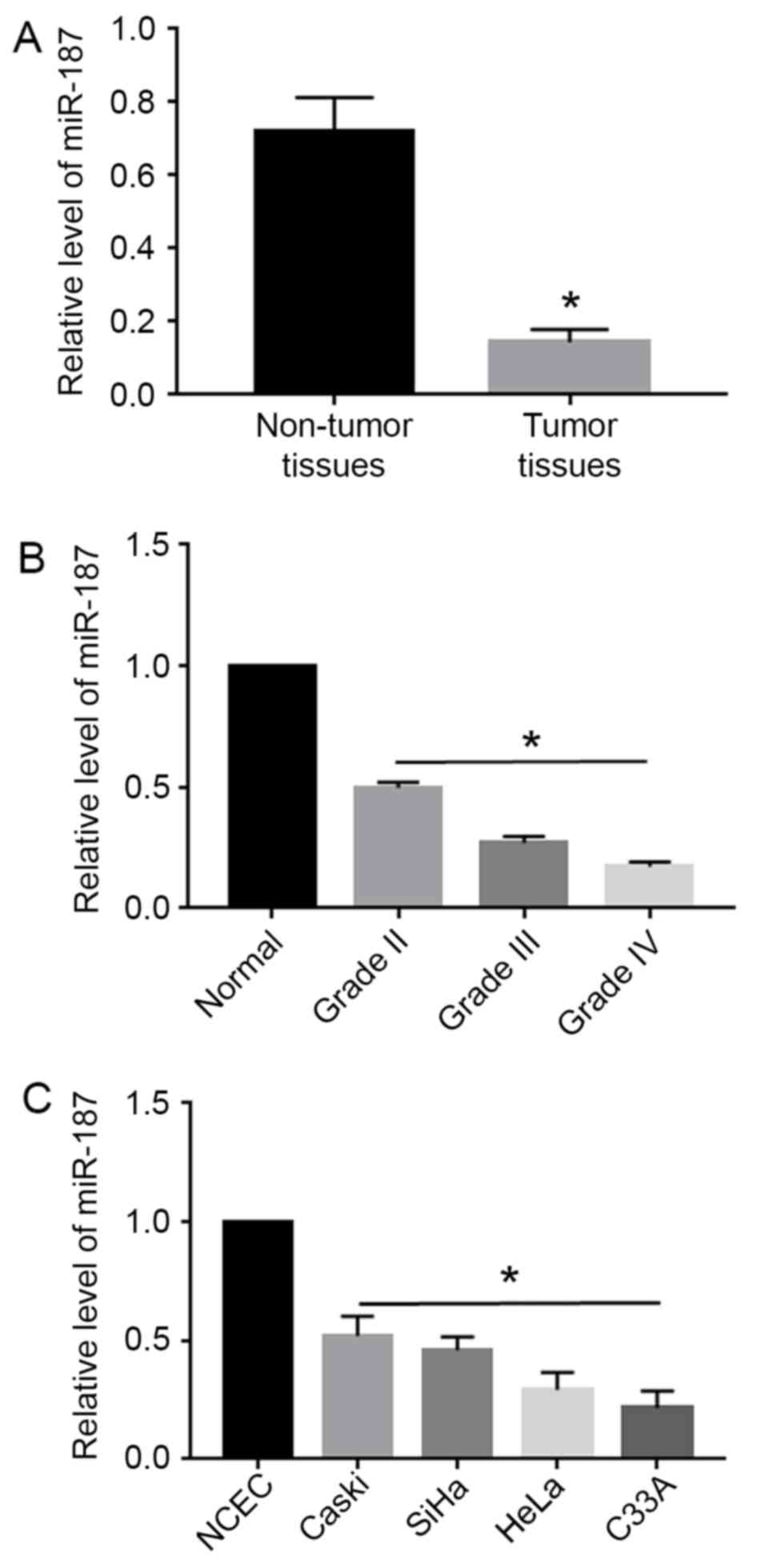
Using qRT-PCR, miR-187 expression levels were
evaluated in 60 pairs of cervical cancer tissues and the
corresponding non-tumor tissues, as well as in cervical cancer cell
lines. Compared with the non-tumor tissues, cervical cancer tissues
showed significantly decreased level of miR-187 (P<0.05;
Fig. 1A). Moreover, the level of
miR-187 was elevated with the progression of the stage of cervical
cancer (P<0.05; Fig. 1B).
Additionally, miR-187 level in all cervical cancer cell lines
including Caski, SiHa, HeLa and C33A was significantly decreased
compared with that in the normal cervical epithelial cells (NCEC)
(P<0.05; Fig. 1C).
Decreased miR-187 level is associated
with poor survival of cervical cancer patients
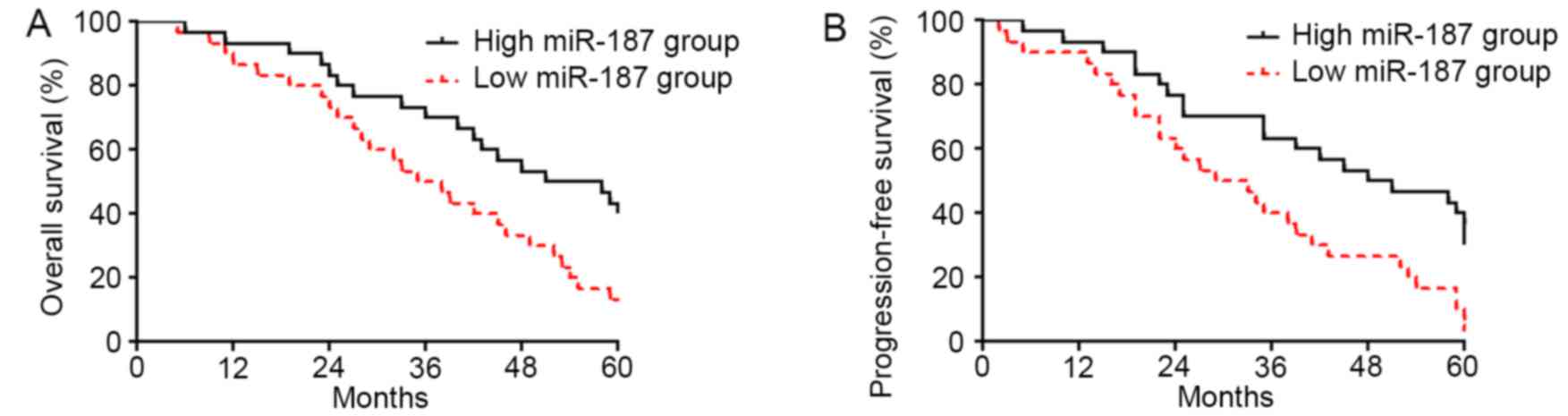
To explore the clinical significance of miR-187
level in cervical cancer, we investigated the correlation of
miR-187 expression level with the prognosis of cervical cancer
patients. As shown in Fig. 2A,
patients with low level of miR-187 had significantly decreased rate
of overall survival (P<0.05; Fig.
2A). The data of progression-free survival demonstrated that
low level of miR-187 was associated with decreased rate of
progression-free survival (P<0.05; Fig. 2B), thus indicating that miR-187
could serve as a biomarker for cervical cancer patients.
miR-187 inhibits the proliferation and
promotes apoptosis of cervical cancer cells
As the miR-187 level was highest in Caski cells and
lowest in C33A cells, we chose Caski cells for knockdown
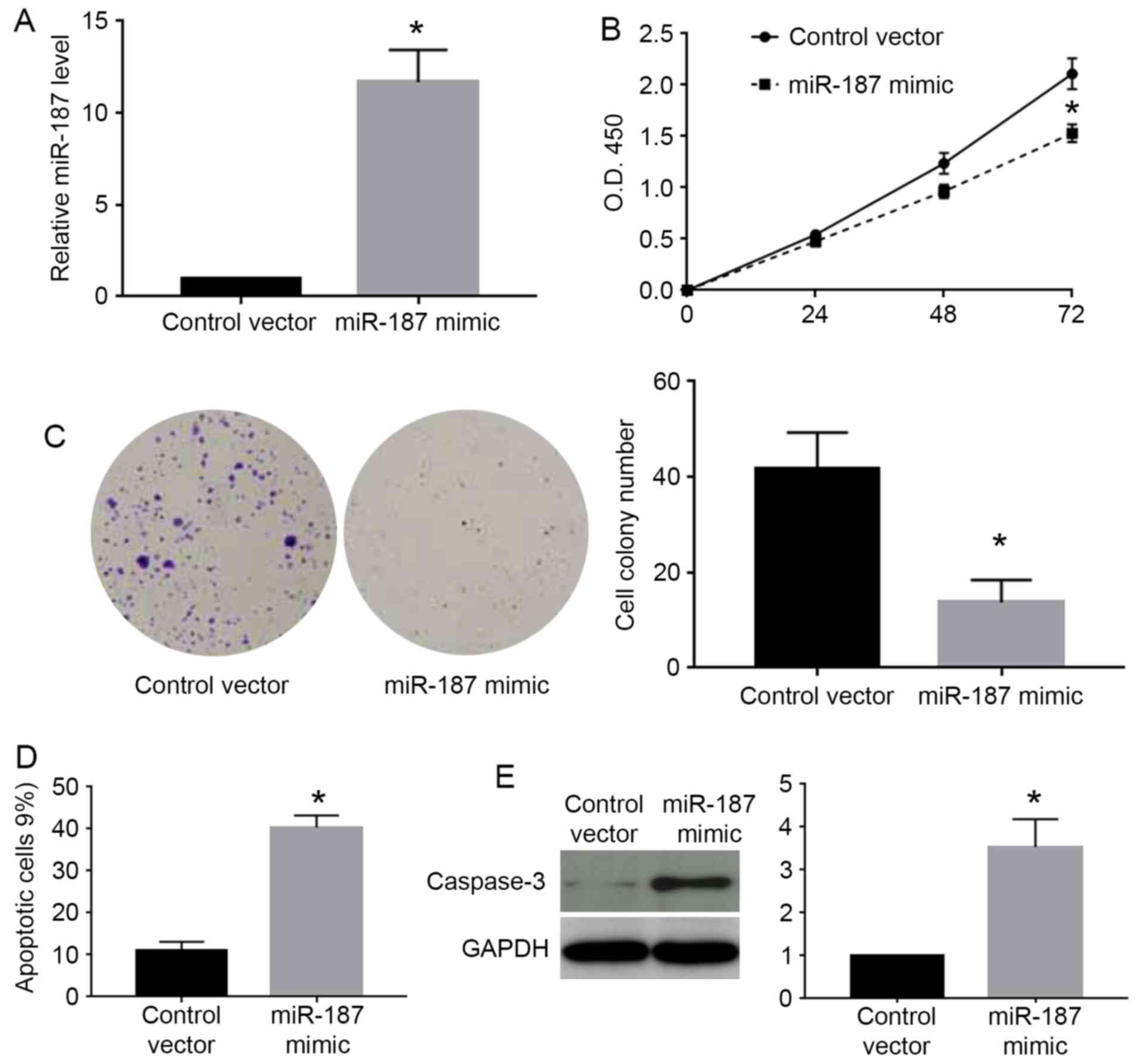
experiments and C33A for overexpression experiments. As shown in
Fig. 3A, miR-187 mimic
significantly increased the expression level of miR-187 in C33A
cells (P<0.05). The MTT assay showed that miR-187 overexpression
significantly decreased the cell viability of C33A cells
(P<0.05; Fig. 3B). Colony
formation assay showed that miR-187 overexpression significantly
decreased the number of formed cell colonies of C33A cells
(P<0.05; Fig. 3C). On the other
hand, flow cytometry assay showed miR-187 overexpression increased
the percentage of apoptotic cells (P<0.05; Fig. 3D). The western blot analysis for
caspase-3 demonstrated that miR-187 overexpression significantly
increased the caspase-3 level in C33A cells (P<0.05; Fig. 3E). Furthermore, we performed miR-187
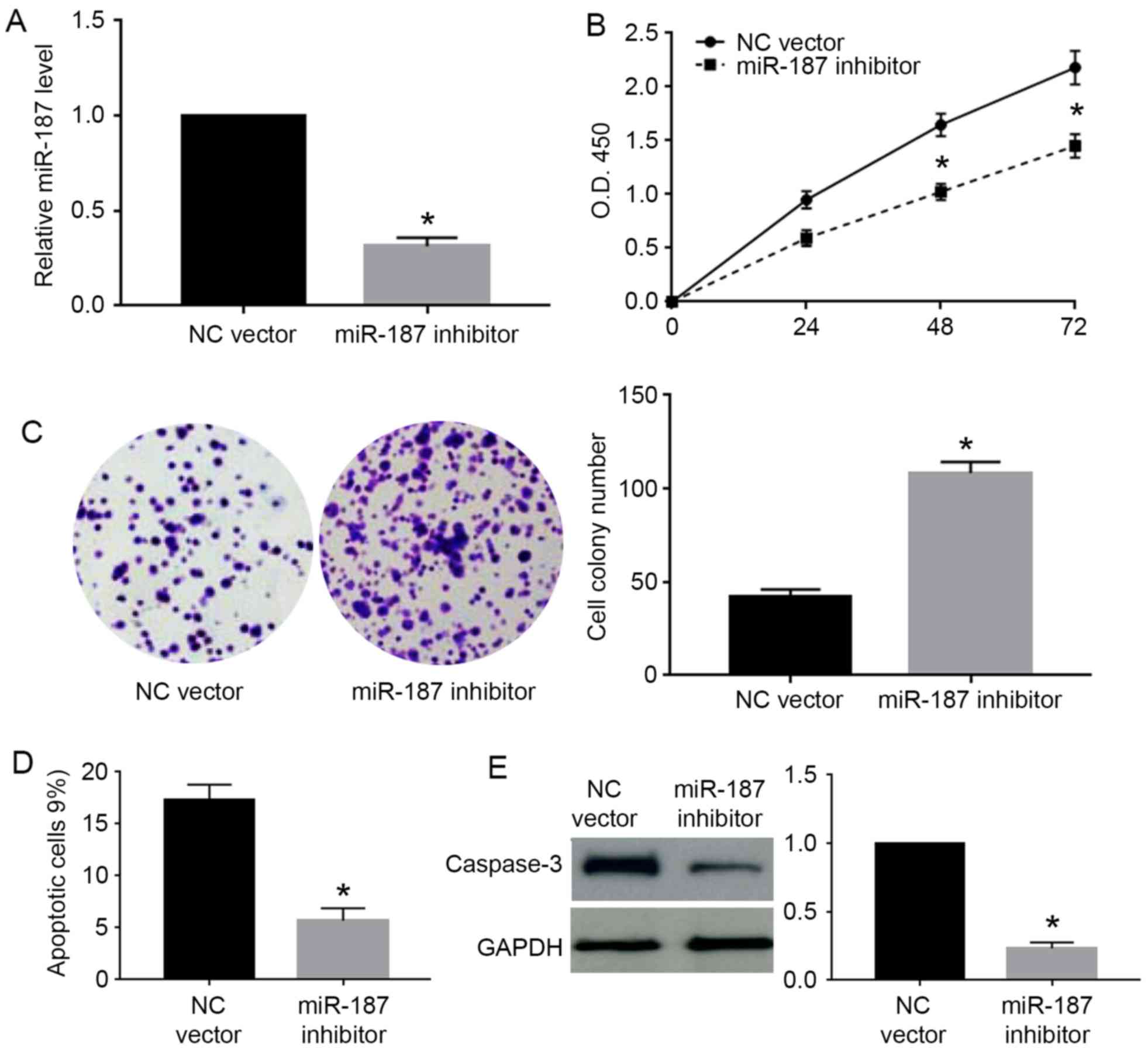
knockdown in Caski cells. miR-187 inhibitor significantly decreased
the level of miR-187 in Caski cells (P<0.05; Fig. 4A). miR-187 knockdown significantly
increased the cell viability (P<0.05; Fig. 4B) and cell colony number (P<0.05;
Fig. 4C) while decreased the
percentage of apoptotic cells (P<0.05; Fig. 4D) and caspase-3 level (P<0.05;
Fig. 4E) in Caski cells.
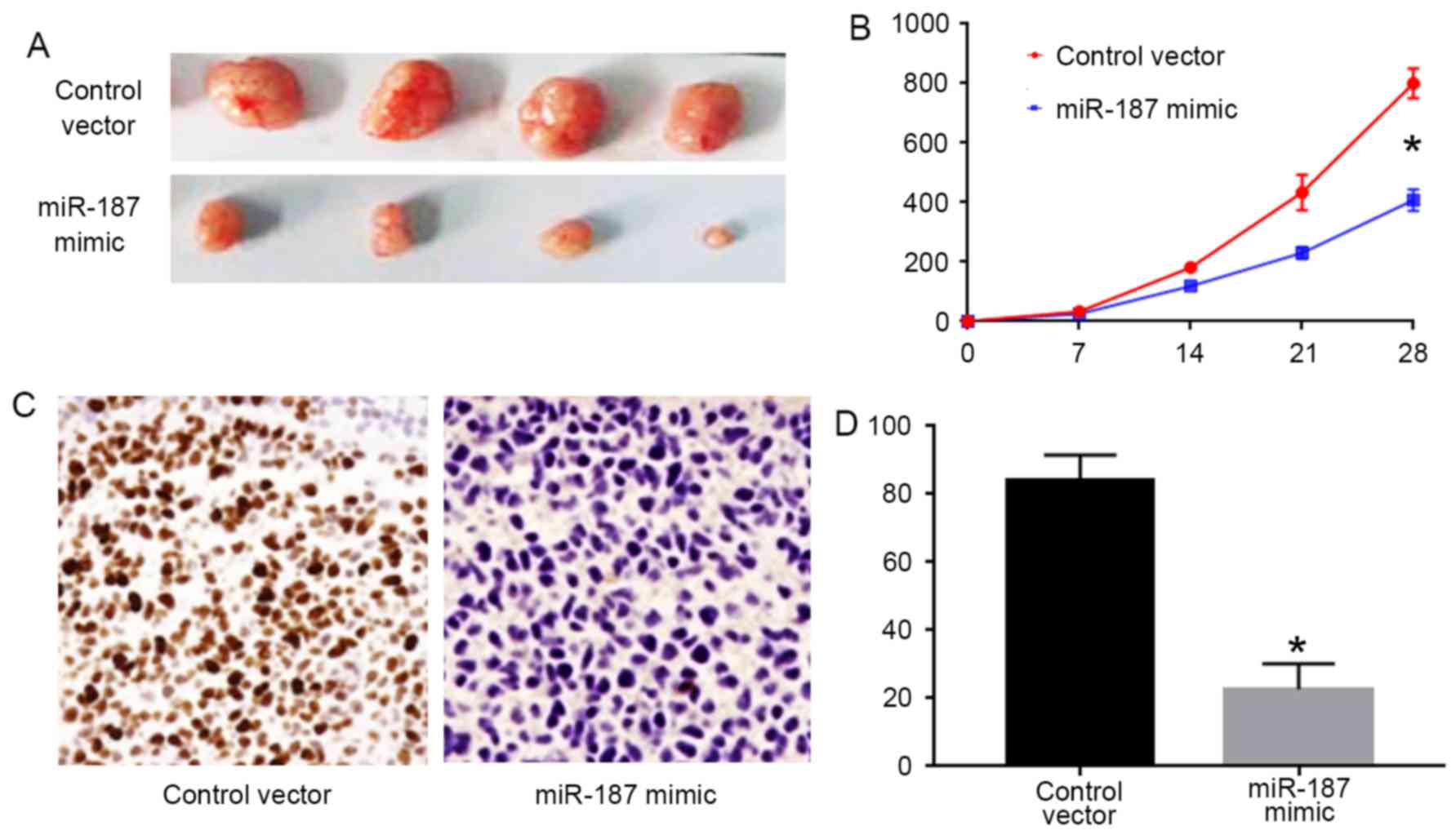
miR-187 inhibits the in vivo growth of
C33A cells
To confirm the in vitro effects of miR-187 on
C33A cells, we performed subcutaneous injection experiments for
C33A cells. As shown in Fig. 5A and
B, forced expression of miR-187 in C33A cells inhibited the
tumor growth in nude mice (P<0.05). Furthermore, we performed
Ki-67 staining for the formed tumors. The results of Ki-67 staining
showed that miR-187 overexpression significantly decreased the
number of Ki-67 positive cells in the tumors (P<0.05; Fig. 5C and D).
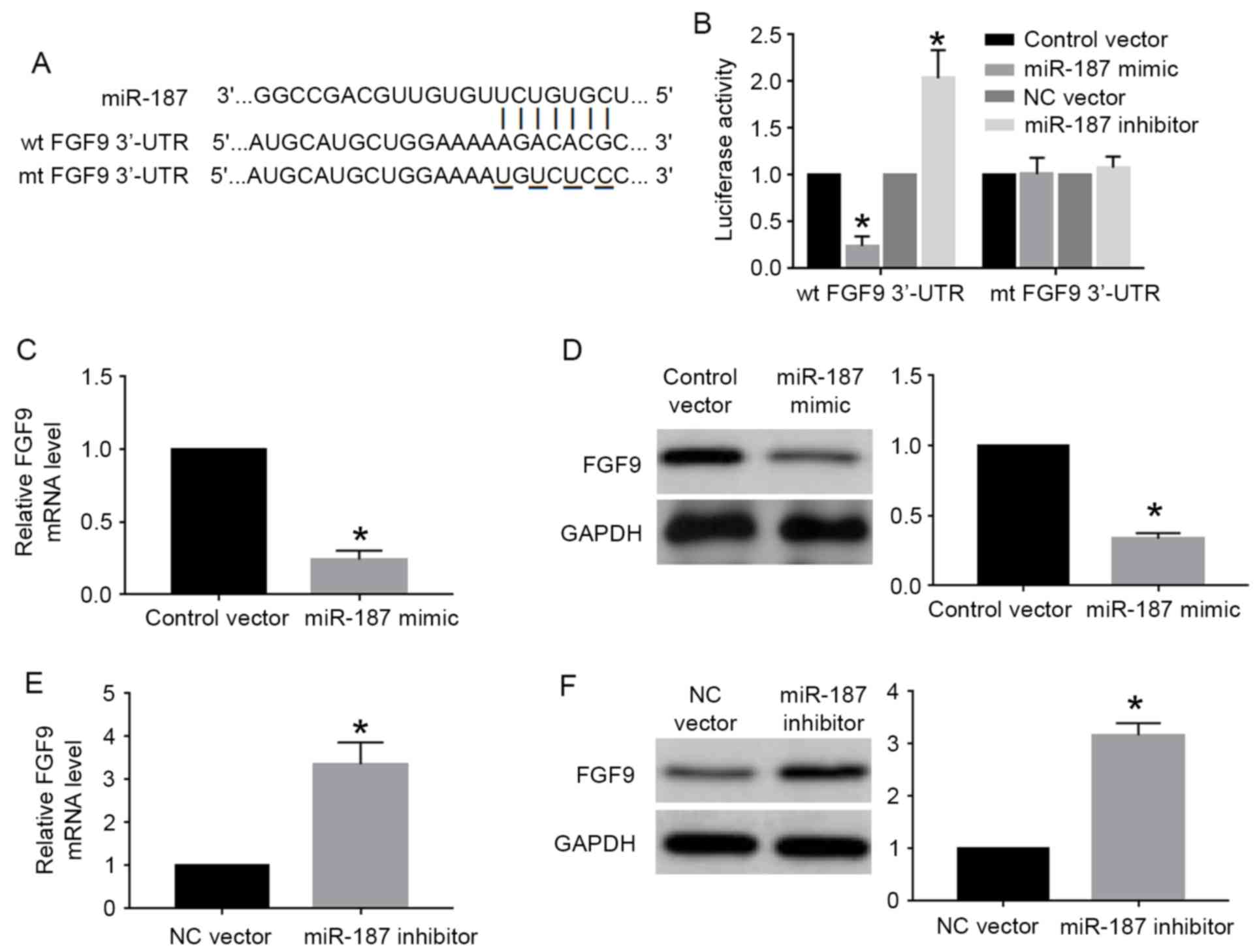
FGF9 is the downstream target of
miR-187 in cervical cancer cells
miRNA regulated cell proliferation, apoptosis and
other cellular processes by interacting with the 3′-UTR site of
targeted genes. FGF9, which had the complementary sequence for
miR-187 binding (Fig. 6A), was one
of the predicted genes of miR-187 based on the data of online
database. We then performed luciferase assay for wild-type and
mutant FGF9 3′-UTR to investigate whether miR-187 could interact
with FGF9 3′-UTR. The results of luciferase assay showed that
miR-187 overexpression significantly decreased the luciferase
activity of wild-type FGF9 3′-UTR while miR-187 knockdown
significantly increased that of wild-type FGF9 3′-UTR (P<0.05;
Fig. 6B). Altering miR-187 level
did not affect the luciferase activity of mutant FGF9 3′-UTR
(Fig. 6B). Furthermore, qRT-PCR and
western blot analysis showed miR-187 overexpression significantly
reduced the mRNA (P<0.05; Fig.
6C) and protein (P<0.05; Fig.
6D) level of FGF9 in C33A cells. miR-187 knockdown
significantly increased the mRNA (P<0.05; Fig. 6E) and protein (P<0.05; Fig. 6F) level of FGF9 in Caski cells.
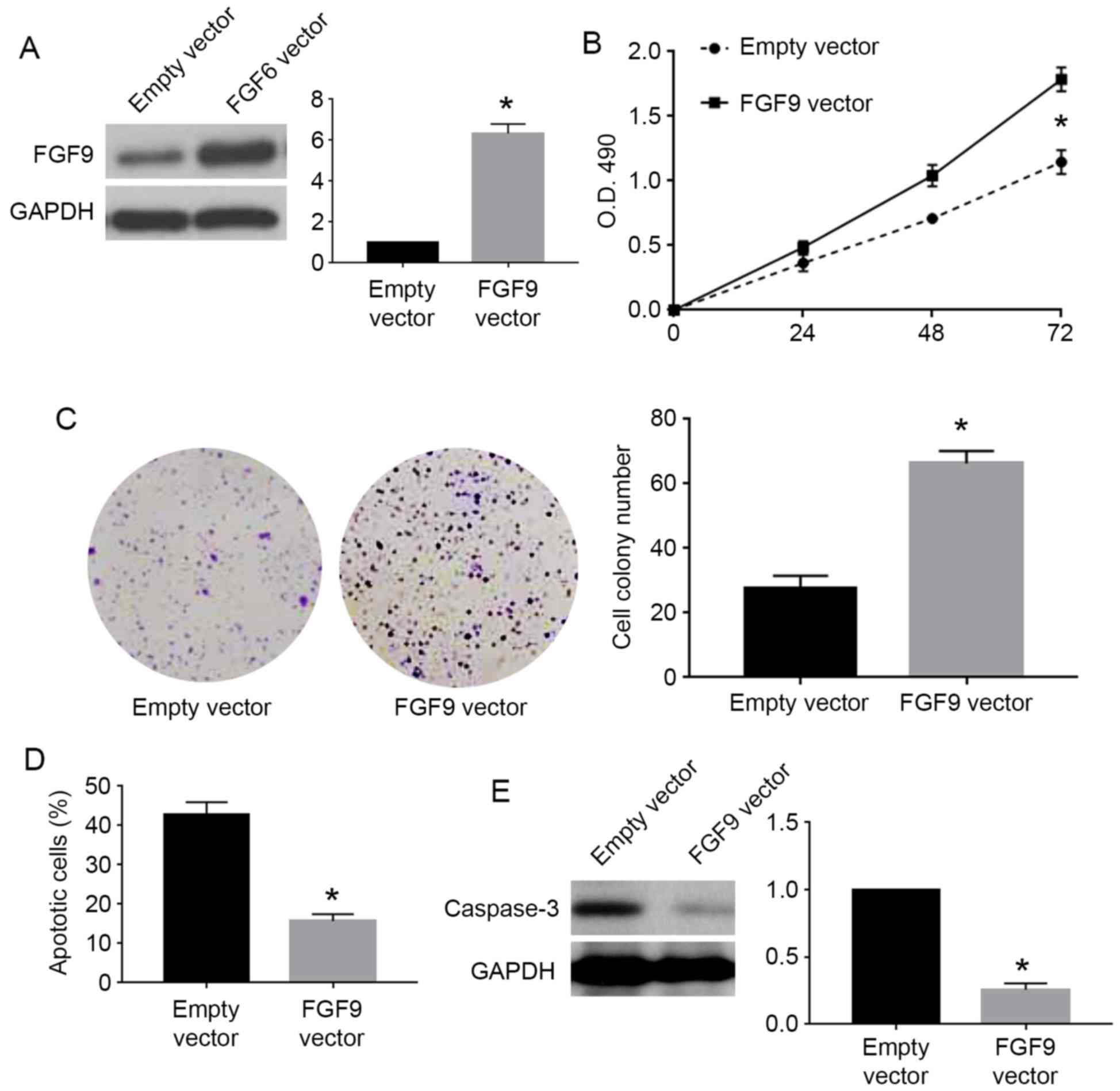
Inhibiting FGF9 was critical for the
biological function of miR-187 in cervical cancer cells
To confirm whether FGF9 was involved in the
biological function of miR-187 in cervical cancer cells, we
performed rescue experiments for FGF9 in cervical cancer cells. For
C33A cells overexpressing miR-187, we used FGF9 vector to
overexpress FGF9 in these cells. FGF9 vector significantly
increased FGF9 expression in C33A cells overexpressing miR-187
(P<0.05; Fig. 7A). FGF9
overexpression reversed the inhibitory effects of miR-187 on cell
viability (P<0.05; Fig. 7B) and
colony formation (P<0.05; Fig.
7C) while blocked the promoting effects of miR-187 on apoptosis
(P<0.05; Fig. 7D) and caspase-3
level (P<0.05; Fig. 7E). For
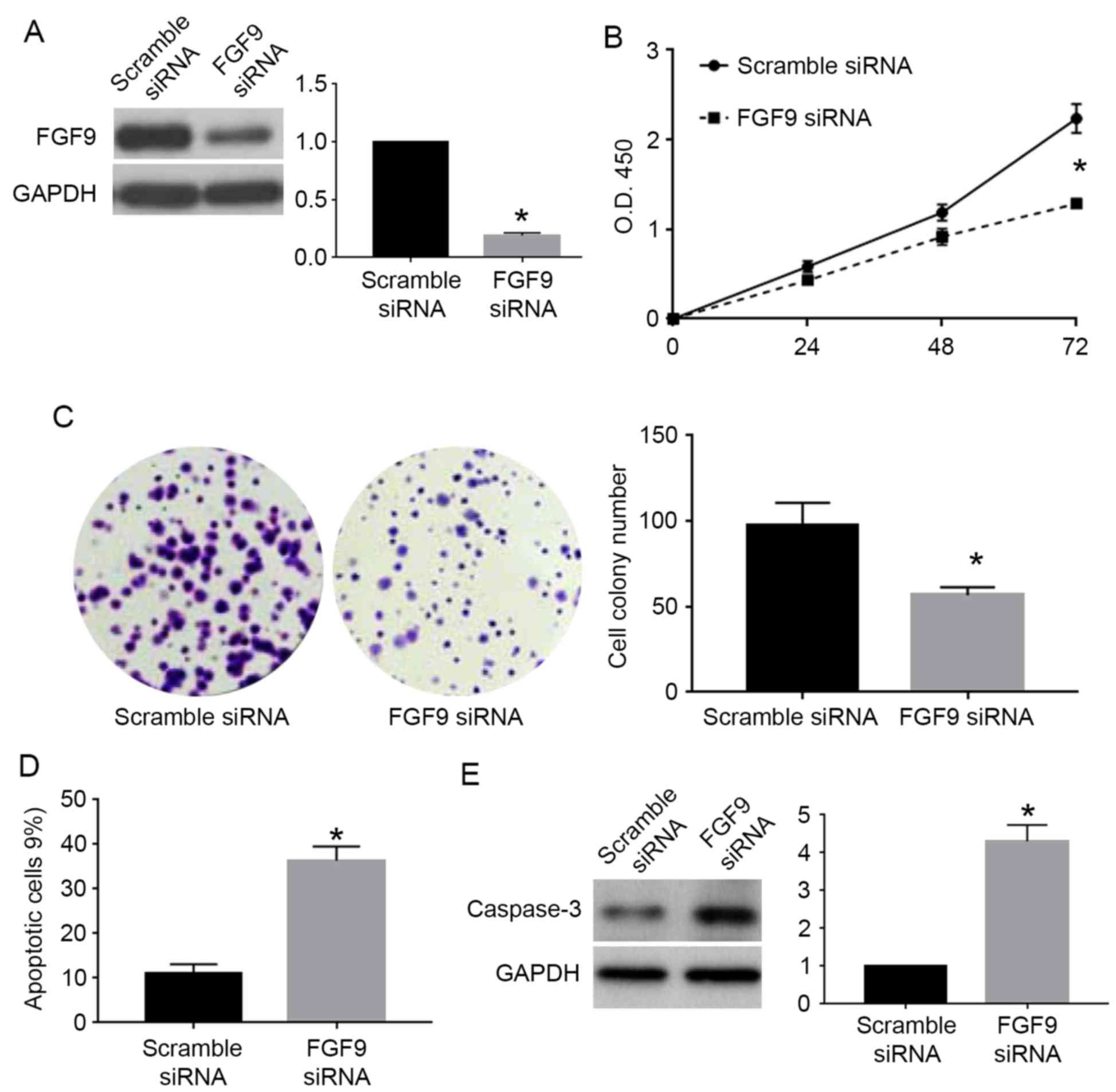
Caski cells with miR-187 knockdown, we used FGF9 siRNA to inhibit
FGF9 expression in these cells. FGF9 siRNA significantly reduced
the expression of FGF9 in Caski cells with miR-187 knockdown
(P<0.05; Fig. 8A). FGF9
knockdown in these cells significantly blocked the promoting
effects of miR-187 knockdown on cell viability (P<0.05; Fig. 8B) and colony formation (P<0.05;
Fig. 8C) while reversed the
inhibitory effects of miR-187 knockdown on apoptosis (P<0.05;
Fig. 8D) and caspase-3 level
(P<0.05; Fig. 8E).
Discussion
Many studies have confirmed that miRNAs were
critical players in the initiation and progression of human cancers
(17–19). Accumulating evidence shows that
miRNAs are promising biomarkers and therapeutic targets in cervical
cancers (9). miR-187 was recently
identified as a novel cancer-related miRNAs. Its expression status
has been confirmed in liver (10),
breast (11), ovarian (12), prostate (13), non-small cell lung (14) and colorectal cancer (15,16).
Study of liver (10), ovarian
(12) and colorectal cancer
(15,16) suggested that miR-187 acted as a
tumor suppressive factor. However, study of breast cancer and
non-small cell lung cancer showed that miR-187 played oncogenic
roles in these tumors (11,14). In the present study, we demonstrated
for the first time that miR-187 expression level was downregulated
in cervical cancer tissues and cell lines. Importantly, decreased
level of miR-187 in cervical cancer patients was associated with
poor prognosis of the patients. These data suggest that miR-187 has
a tumor suppressive role in cervical cancer and can serve as a
biomarker for cervical cancer patients.
Previous studies regarding miR-187 showed that
miR-187 had different roles in different types of human cancers.
miR-187 was found to inhibit the metastasis and
epithelial-mesenchymal transition of liver cancer cells (10) and colorectal cancers (15,16). A
study of colorectal cancer cells showed that miR-187 could also
inhibit the growth of colorectal cancer cells (16). In this study, both overexpression
and knockdown methods confirmed that miR-187 exerted its tumor
suppressive roles in cervical cancer cells by inhibiting
proliferation and prompting apoptosis of cervical cancer cells.
In vivo experiments proved that miR-187 could inhibit the
in vivo growth of cervical cancer cells in nude mice.
miRNAs usually have multiple downstream targets in
different types of cells. Previously identified downstream targets
of miRNAs include S100A4 (10),
Dab2 (12), aldehyde dehydrogenase
1A3 (13) and CD276 (16). In this study, we identified that
FGF9 was a novel downstream target of miR-187 in cervical cancers
based on the data of luciferase assay, qRT-PCR and western blot
analysis. FGF9 is well known oncogenic protein in human cancers
including colon (20), ovarian
(21) and lung cancer (22). Our data demonstrated that inhibiting
FGF9 was critical for miR-187 exerting its suppressive effects on
the growth of cervical cancer cells.
In conclusion, the present study demonstrated that
miR-187 level was decreased in cervical cancer tissues and cell
lines. Decreased miR-187 level in cervical cancer patients was
associated with poor overall survival and progression-free
survival. miR-187 inhibited the proliferation and promoted
apoptosis of cervical cancer cells. In vivo experiment
confirmed that miR-187 inhibited the growth of cervical cancer
cells in nude mice. Furthermore, FGF9 was identified to be the
downstream target of miR-187 in cervical cancer cells. Inhibition
of FGF9 was required for miR-187 to exert tumor suppressive role in
cervical cancer cells.
References
|
1
|
DeSantis CE, Lin CC, Mariotto AB, Siegel
RL, Stein KD, Kramer JL, Alteri R, Robbins AS and Jemal A: Cancer
treatment and survivorship statistics, 2014. CA Cancer J Clin.
64:252–271. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Hildesheim A and Wang SS: Host and viral
genetics and risk of cervical cancer: A review. Virus Res.
89:229–240. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Bosch FX and de Sanjosé S: Chapter 1:
Human papillomavirus and cervical cancer - burden and assessment of
causality. J Natl Cancer Inst Monogr. 2003:3–13. 2003. View Article : Google Scholar
|
|
4
|
Smith JS, Lindsay L, Hoots B, Keys J,
Franceschi S, Winer R and Clifford GM: Human papillomavirus type
distribution in invasive cervical cancer and high-grade cervical
lesions: A meta-analysis update. Int J Cancer. 121:621–632. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Gutschner T and Diederichs S: The
hallmarks of cancer: A long non-coding RNA point of view. RNA Biol.
9:703–719. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Esquela-Kerscher A and Slack FJ: Oncomirs
- microRNAs with a role in cancer. Nat Rev Cancer. 6:259–269. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Hu X, Schwarz JK, Lewis JS Jr, Huettner
PC, Rader JS, Deasy JO, Grigsby PW and Wang X: A microRNA
expression signature for cervical cancer prognosis. Cancer Res.
70:1441–1448. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Farazi TA, Hoell JI, Morozov P and Tuschl
T: MicroRNAs in human cancer. Adv Exp Med Biol. 774:1–20. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhao S, Yao D, Chen J and Ding N:
Circulating miRNA-20a and miRNA-203 for screening lymph node
metastasis in early stage cervical cancer. Genet Test Mol
Biomarkers. 17:631–636. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Dou C, Liu Z, Xu M, Jia Y, Wang Y, Li Q,
Yang W, Zheng X, Tu K and Liu Q: miR-187-3p inhibits the metastasis
and epithelial-mesenchymal transition of hepatocellular carcinoma
by targeting S100A4. Cancer Lett. 381:380–390. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Mulrane L, Madden SF, Brennan DJ, Gremel
G, McGee SF, McNally S, Martin F, Crown JP, Jirström K, Higgins DG,
et al: miR-187 is an independent prognostic factor in breast cancer
and confers increased invasive potential in vitro. Clin Cancer Res.
18:6702–6713. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chao A, Lin CY, Lee YS, Tsai CL, Wei PC,
Hsueh S, Wu TI, Tsai CN, Wang CJ, Chao AS, et al: Regulation of
ovarian cancer progression by microRNA-187 through targeting
Disabled homolog-2. Oncogene. 31:764–775. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Casanova-Salas I, Masiá E, Armiñán A,
Calatrava A, Mancarella C, Rubio-Briones J, Scotlandi K, Vicent MJ
and López-Guerrero JA: MiR-187 targets the androgen-regulated gene
ALDH1A3 in prostate cancer. PLoS One. 10:e01255762015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Sun C, Li S, Yang C, Xi Y, Wang L, Zhang F
and Li D: MicroRNA-187-3p mitigates non-small cell lung cancer
(NSCLC) development through down-regulation of BCL6. Biochem
Biophys Res Commun. 471:82–88. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhang F, Luo Y, Shao Z, Xu L, Liu X, Niu
Y, Shi J, Sun X, Liu Y, Ding Y, et al: MicroRNA-187, a downstream
effector of TGFβ pathway, suppresses Smad-mediated
epithelial-mesenchymal transition in colorectal cancer. Cancer
Lett. 373:203–213. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wang ZS, Zhong M, Bian YH, Mu YF, Qin SL,
Yu MH and Qin J: MicroRNA-187 inhibits tumor growth and invasion by
directly targeting CD276 in colorectal cancer. Oncotarget.
7:44266–44276. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Gregory RI and Shiekhattar R: MicroRNA
biogenesis and cancer. Cancer Res. 65:3509–3512. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Calin GA and Croce CM: MicroRNA signatures
in human cancers. Nat Rev Cancer. 6:857–866. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hampton T: MicroRNA and metastasis. JAMA.
298:1998. 2007. View Article : Google Scholar
|
|
20
|
Fearon ER: Molecular genetics of
colorectal cancer. Annu Rev Pathol. 6:479–507. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hendrix ND, Wu R, Kuick R, Schwartz DR,
Fearon ER and Cho KR: Fibroblast growth factor 9 has oncogenic
activity and is a downstream target of Wnt signaling in ovarian
endometrioid adenocarcinomas. Cancer Res. 66:1354–1362. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
White AC, Xu J, Yin Y, Smith C, Schmid G
and Ornitz DM: FGF9 and SHH signaling coordinate lung growth and
development through regulation of distinct mesenchymal domains.
Development. 133:1507–1517. 2006. View Article : Google Scholar : PubMed/NCBI
|