Introduction
Gastric cancer (GC) is one of the most common
malignancies, and is the second leading cause of cancer-related
death worldwide (1). Although rapid
advancements in chemotherapy, radiation therapy, and gastric
resection are widely used for GC patients, the 5-year overall
survival rate has remained at ~28% owing to tumor metastasis
(2,3).
The tumor microenvironment (TME) is comprised of
tumor cells, tumor stroma, blood vessels, infiltrating inflammatory
cells and a variety of associated tissue cells (4). Increasing evidence has indicated that
the TME plays an important role in cancer development and
metastasis (4,5). Macrophages infiltrated in the TME are
called tumor-associated macrophages (TAMs) and are key
orchestrators in TME (6,7). TAMs play a critical role in the
regulation of tumor growth and progression (8,9). The
high density of TAMs is correlated with a poor prognosis in various
types of cancer including GC (9–15).
However, the role of TAMs in GC and the underlying mechanism remain
elusive.
Epithelial-mesenchymal transition (EMT) is a process
by which epithelial cells lose their epithelial attributes and
acquire a mesenchymal cell phenotype, which is a key process in
promoting tumor cell metastasis (16). TAMs induce EMT in pancreatic and
non-small cell lung cancer (NSCLC), breast cancer,
cholangiocarcinoma and hepatocellular carcinoma (17–22).
Using literature review, we highlighted the role of TAMs in the
regulation of EMT during tumorigenesis (23). In GC, it has been reported that
infiltration of TAMs is associated with EMT-related proteins in
human GC tissues, but the exact mechanism has not been clarified
(14).
In the present study, we aimed to investigate the
role of TAMs in GC through the regulation of EMT. FOXQ1, a forkhead
box-containing transcription factor, promotes EMT and metastasis in
various types of cancer including GC (24–33).
The potential involvement of FOXQ1 in TAM-induced EMT and
metastasis was also investigated.
Materials and methods
Cell culture, reagent and plasmid
The THP-1 cell line and the human GC cell lines,
MKN45 and MKN74, were used in the present study. All cell lines
were obtained from the Cell Bank of Shanghai (Shanghai, China). The
cells were grown in RPMI-1640 medium (Gibco, Gaithersburg, MD, USA)
that was supplemented with 10% fetal bovine serum (FBS), penicillin
(100 U/ml) and streptomycin (100 mg/ml), and were incubated in a
humidified atmosphere containing 5% CO2 at 37°C and the
medium was replaced three times/week.
Rabbit anti-FOXQ1 (1:100; ab51340; Abcam, Cambridge,
MA, USA), rabbit anti-E-cadherin (1:100; AF0131; Affinity,
Sterling, VA, USA), rabbit anti-vimentin (1:100; 5741; Cell
Signaling Technology, Inc., Beverly, MA, USA) and mouse
anti-β-actin (1:100; T0022; Affinity) were used as primary
antibodies. The FOXQ1 shRNA lentiviral particle containing FOXQ1
shRNA sequences was purchased from Santa Cruz Biotechnology, Inc.
(Santa Cruz, CA, USA). MKN45 and MKN74 cells were infected with
shFOXQ1 lentiviral particles and a negative control for 48 h and
followed by 2 mg/ml of puromycin selection.
Co-culture of GC cells and
macrophage
GC MKN45 and MKN74 cells (105 cells/well)
were seeded into 24-well plates (BD Biosciences, Franklin Lakes,
NJ, USA) in RPMI-1640 medium supplemented with 10% FBS, penicillin
and streptomycin. THP-1 cells (3ⅹ105 cells/insert) were
seeded into the upper chamber of a Transwell insert with a pore
size of 8.0-µm (Corning Incorporated, Kennebunk, ME, USA) in
RPMI-1640 medium supplemented with 10% FBS, penicillin and
streptomycin. The cells were incubated in a humidified atmosphere
containing 5% CO2 at 37°C.
Quantitative real-time reverse
transcription PCR
Total RNA was extracted with TRIzol reagent
(Invitrogen, Carlsbad, CA, USA) according to the manufacturer's
instructions and reverse-transcribed into cDNA using a reverse
transcription kit from Takara Biotechnology, Ltd. (Dalian, China).
After adjusting the cDNA concentration in all groups, qRT-PCR was
performed using the CFX96 Real-Time PCR Detection System (Bio-Rad,
Hercules, CA, USA) with SYBR-Green. The PCR conditions were as
follows: pre-denaturation at 95°C for 30 sec; 35 cycles of
denaturation (95°C for 5 sec), annealing (55–60°C for 30 sec) and
extension (72°C for 1 min); and a final extension at 72°C for 10
min. The relative level of gene expression was calculated using the
ΔΔCt method with normalization to GAPDH. All experiments were
performed in triplicate. The primers used are listed in Table I.
 | Table I.Primer sequences for qRT-PCR
amplification of different genes. |
Table I.
Primer sequences for qRT-PCR
amplification of different genes.
| Gene | Primer |
|---|
| GAPDH | F
5′-CTTTGGTATCGTGGAAGGACTC-3′ |
|
| R
5′-GTAGAGGCAGGGATGATGTTCT-3′ |
| E-cadherin | F
5′-TGGCTTCCCTCTTTCATCTCC-3′ |
|
| R
5′-TCATAGTTCCGCTCTGTCTTTGG-3′ |
| Vimentin | F
5′-TCAATGTTAAGATGGCCCTTG-3′ |
|
| R
5′-TGAGTGGGTATCAACCAGAGG-3′ |
| FOXQ1 | F
5′-TGATTTCTTGCTATTGACCGATGC-3′ |
|
| R
5′-GCCCAAGGAGACCACAGTTAGAG-3′ |
Western blotting
Protein expression levels were analyzed by western
blotting standard protocols. The protein of MKN45 and MKN74 cells
was extracted using RIPA lysis buffer (Beyotime, Haimen, China)
with protease inhibitor PMSF (CWBiotech, Beijing, China) according
to the manufacturer's instructions. Briefly, 20 µl of total protein
extracts was resolved by denaturing sodium dodecyl
sulfate-polyacrylamide gel electrophoresis and transferred to
polyvinylidene difluoride membranes. The membranes were blocked
with 5% non-fat milk, and then incubated with primary antibodies.
Then, the blots were washed and probed with the respective
secondary peroxidase-conjugated antibodies. Signals were detected
using a chemiluminescence solvent (Thermo Scientific, Rockford, IL,
USA).
Transwell invasion assay
THP-1 cells (105 cells/well) were seeded
in 24-well plates in RPMI-1640 medium supplemented with 10% FBS.
MKN45 and MKN74 cells (104 cells/insert) were seeded
into the upper chamber of a Transwell insert with a pore size of
8.0 µm precoated to a Matrigel Basement Membrane Matrix (BD
Biosciences) in RPMI-1640 medium supplemented with 1% FBS. After
incubated in a humidified atmosphere containing 5% CO2
at 37°C for 24 h, the cells migrated to the lower surface of the
membranes were fixed with 4% paraformaldehyde, stained with 0.1%
crystal violet and counted under a microscope. All these samples
were plated three times.
Wound healing assay
Approximately 5ⅹ104 GC cells from
different groups were seeded in 24-well plates and incubated for 24
h. Then, the monolayer cells were disrupted by scratching with a 10
µl microsterile pipette tips. Images were captured at 0 and 24 h in
a phase-contract microscope. The assays were performed in
triplicate, and four fields of each well were assessed.
Tissue specimens, immunohistochemistry
and assessment of CD68 and FOXQ1 expression
All specimens were obtained from the Department of
Surgical Oncology, The First Affiliated Hospital, Xi'an Jiaotong
University and the Department of Surgical Oncology, the 215th
Hospital of Shaanxi Province. Detailed information on the specimens
was previously provided (14). The
present study was approved by the Protection of Human Subjects
Committee of the First Affiliated Hospital, Xi'an Jiaotong
University and complied with the Helsinki Declaration. The tissues
specimens were fixed in neutral buffered formalin and embedded in
paraffin wax. The sections of 4-mm thickness were cut and mounted
on charged glass slides. Antigen retrieval was performed using
citrate buffer at pH 6.0. Immunohistochemical staining was
performed using mouse anti-human CD68 (1:100; ZM-0060; Beijing
Zhongshan Biotechnology, Beijing, China) and rabbit anti-human
FOXQ1 (1:100; bs-16175R; Beijing Bioss Biotechnology, Beijing,
China). The streptavidin-peroxidase technique (SP-9001 Golden
Bridge Int., Beijing, China) was used. An irrelevant rabbit
antiserum served as a negative control. The sections were stained
with 0.02% diaminobenzidine (DAB) solution followed by
counterstaining with hematoxylin. Staining results were classified
into high expression and low expression as previously described
(14,32).
Statistical analysis
Data analyses were performed using SPSS statistical
package 16.0 (SPSS Institute, Chicago, IL, USA) or Prism (GraphPad
Software, Inc., La Jolla, CA, USA). A P-value <0.05 was
considered to indicate a statistically significant result. The
χ2 test was used to analyze the correlation between CD68
and FOXQ1 expression.
Results
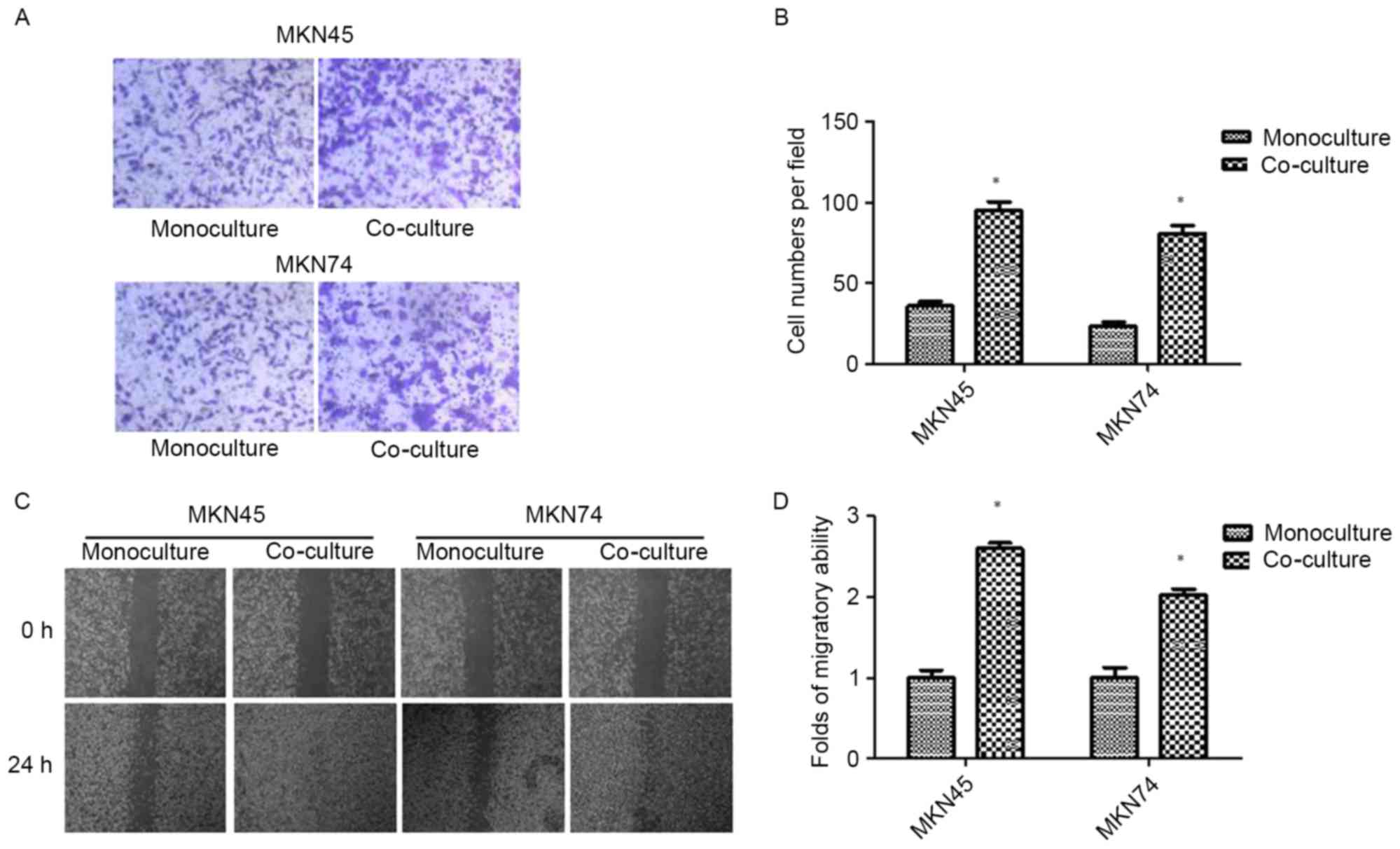
Co-culture with TAMs promotes invasion
and migration in GC cells
After 24 h of THP-1 co-culture, GC (MKN45 and MKN74)
cells were subjected to Transwell invasion assays. As shown in
Fig. 1A and B, THP-1 co-culture
resulted in an increase in the invasive ability of both MKN45 and
MKN74 cells. The result was confirmed by wound-healing assay.
Compared with the control, GC cells co-cultured with THP-1 cells
exhibited a faster closure of the wound (Fig. 1C and D).
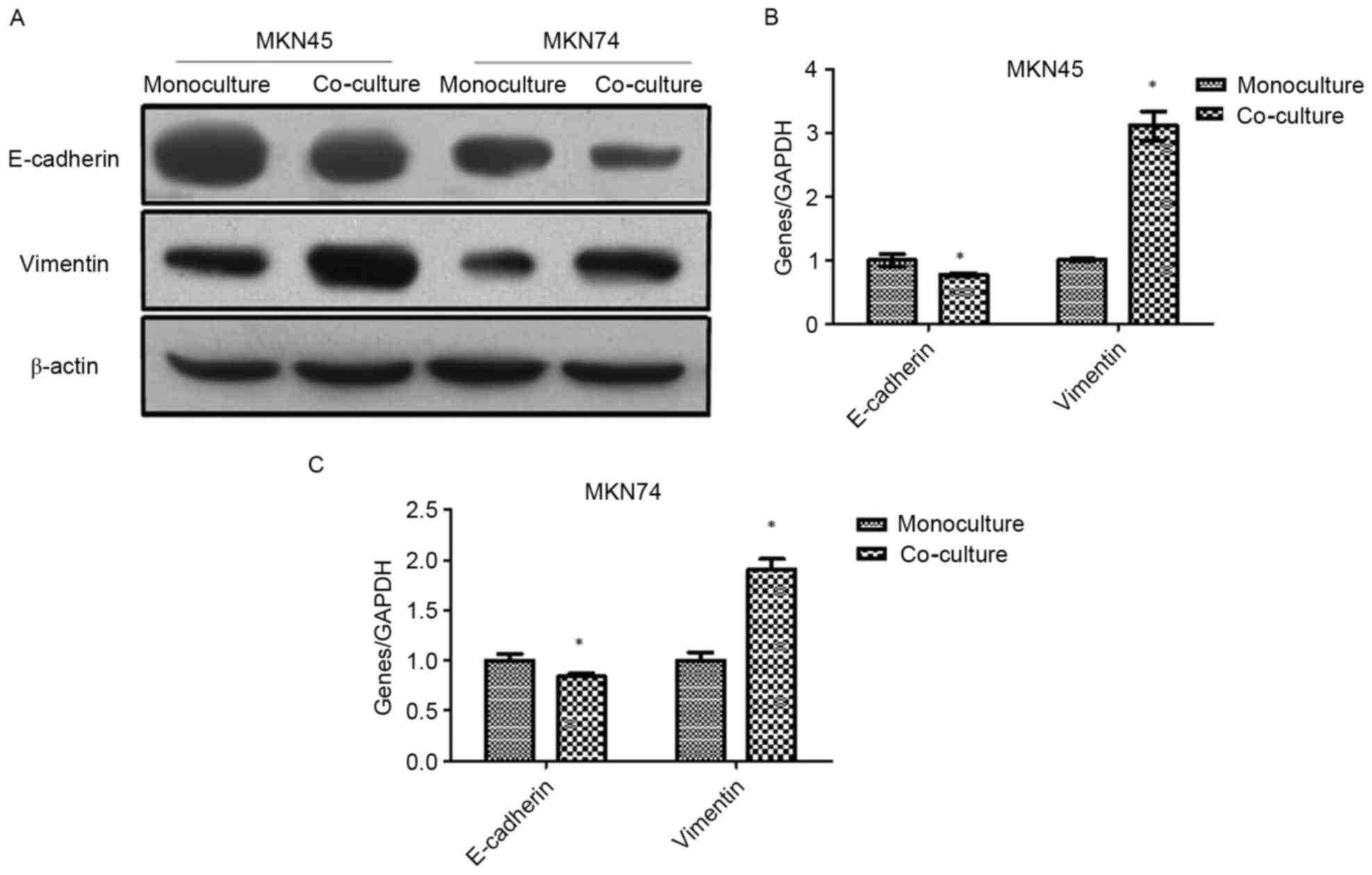
Co-culture with TAMs induces EMT in GC
cells
Western blotting and RT-PCR were used to analyze the
EMT markers in MKN45 and MKN74 cells after being co-cultured with
THP-1. As shown in Fig. 2, the
expression of epithelial marker E-cadherin was downregulated, while
the mesenchymal marker vimentin was upregulated.
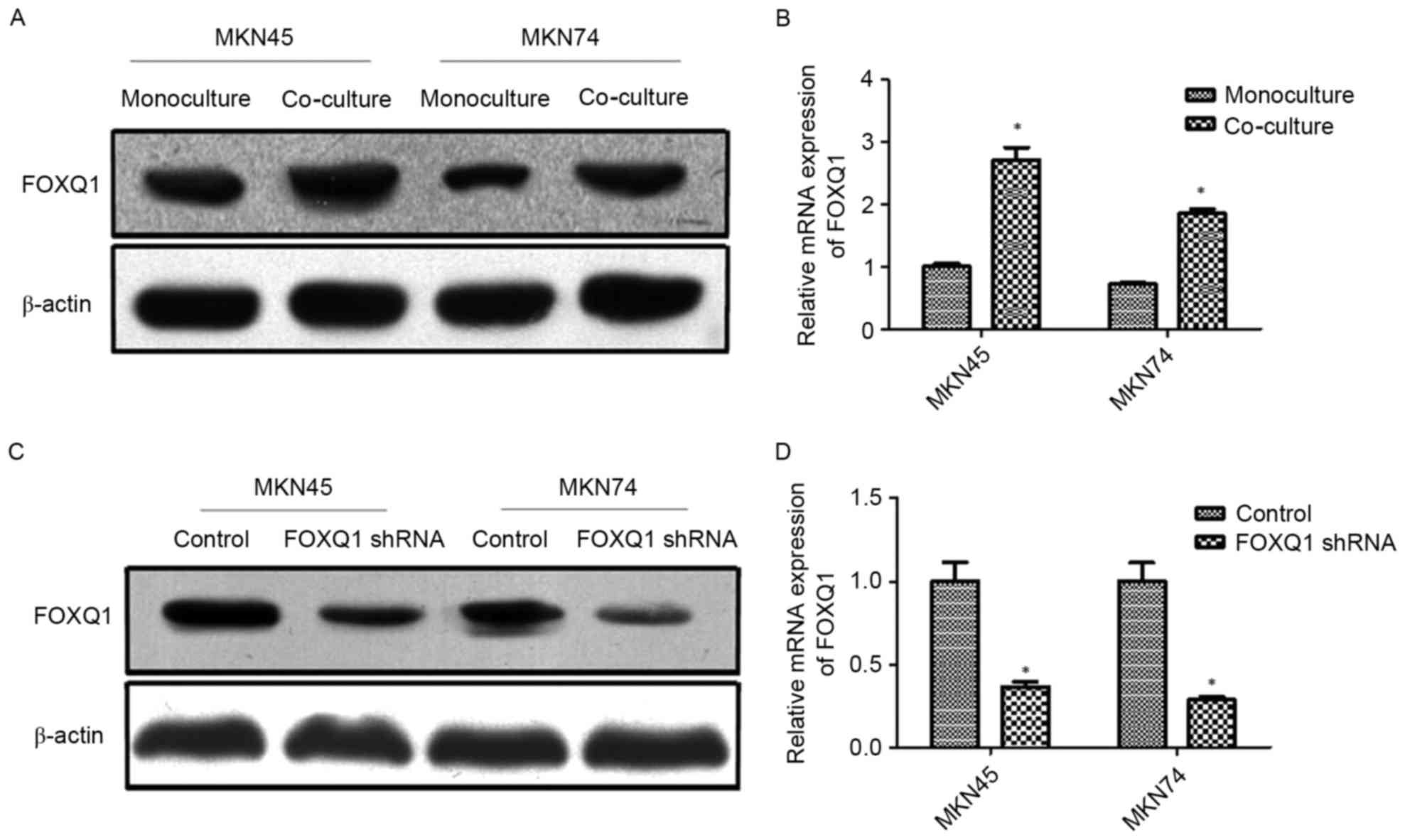
FOXQ1 is involved in the EMT of GC
cells induced by co-culture with TAMs
A previous study revealed that FOXQ1 promoted EMT
and metastasis in GC (32). The
potential involvement of FOXQ1 in TAM-induced EMT and metastasis is
still unknown. In the present study, MKN45 and MKN74 cells were
co-cultured with THP-1. The results revealed that co-culture with
THP-1 significantly increased the expression of FOXQ1 at both the
mRNA and protein levels of MKN45 and MKN74 cells (Fig. 3A and B). Thereafter, transfection of
shRNA targeting FOXQ1 resulted in pronounced knockdown of mRNA and
protein levels in MKN45 and MKN74 cells (Fig. 3C and D).
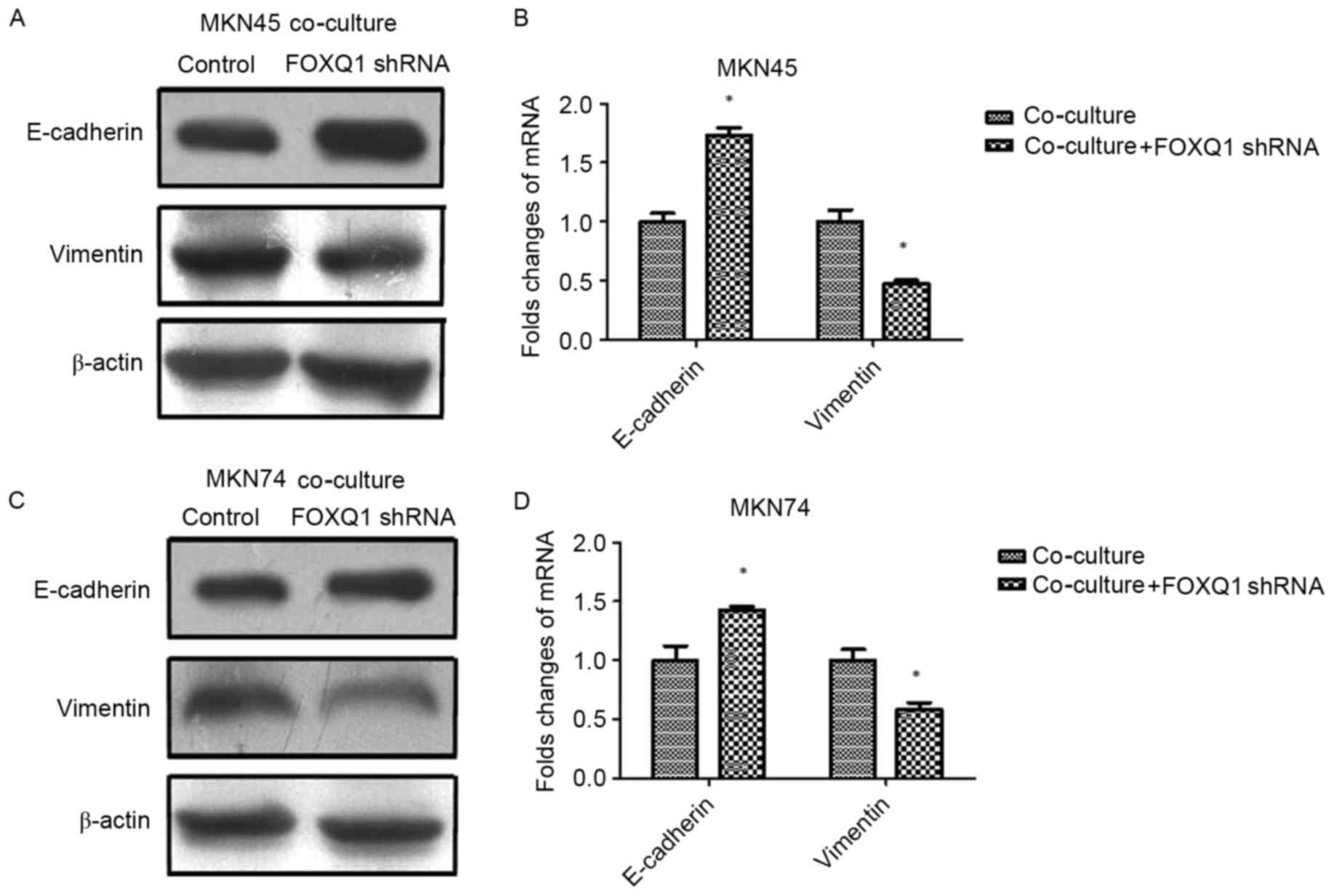
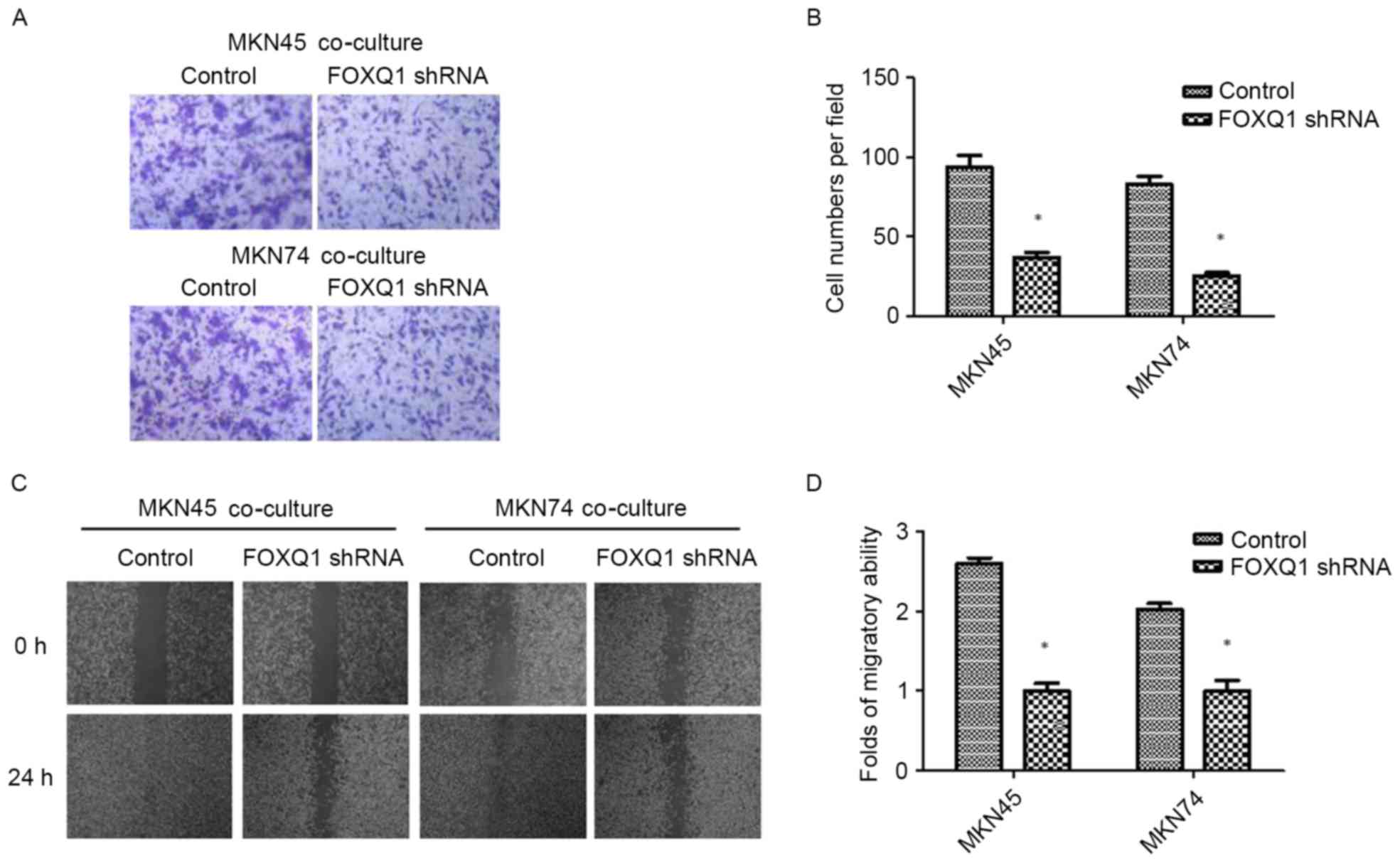
FOXQ1 is essential for TAM-induced EMT
and metastasis in GC cells
To confirm the role of FOXQ1 in TAM-induced EMT and
invasion in GC cells, we silenced FOXQ1 expression in MKN45 and
MKN74 cells with shRNA before being co-cultured with THP-1 cells.
As shown in Fig. 4, the expression
of mesenchymal marker vimentin was decreased, while the epithelial
marker E-cadherin was increased in both MKN45-FOXQ1-shRNA and
MKN74-FOXQ1-shRNA cells co-cultured with THP-1 cells. Transwell
invasion assays revealed that silencing of FOXQ1 in MKN45 and MKN74
cells decreased their invasive ability after being co-cultured with
THP-1 (Fig. 5A and B).
Wound-healing assay indicated that silencing of FOXQ1 in GC MKN45
and MKN74 cells co-cultured with THP-1 cells exhibited a slower
closure of the wound (Fig. 5C and
D). These results clearly revealed that silencing of FOXQ1
blocked the effect of TAM-enhanced EMT and metastasis of GC
cells.
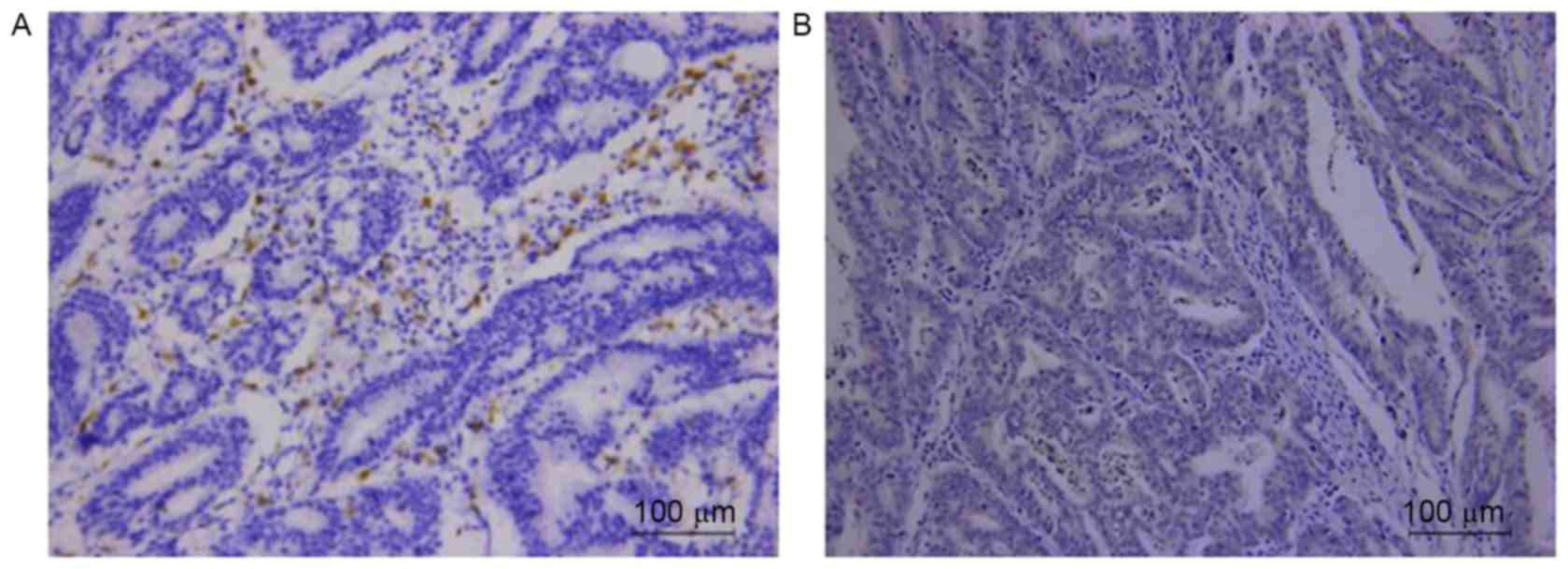
Correlation of TAM marker CD68 and
FOXQ1 expression in GC tissues
Respective photomicrographs of immunohistochemical
staining of CD68 and FOXQ1 are shown in Fig. 6. The relationship between the
expression of CD68 and FOXQ1 was calculated and has been outlined
in Table II. The result revealed
that high expression of CD68 was correlated with positive FOXQ1
expression (r=0.613; P<0.001) in clinical GC samples.
| Table II.The association between CD68 and
FOXQ1. |
Table II.
The association between CD68 and
FOXQ1.
|
| CD68 |
|
|
|---|
|
|
|
|
|
|---|
|
| High | Low | r | P-value |
|---|
| FOXQ1 |
|
|
|
|
|
High | 79 | 24 | 0.613 | 0.000 |
|
Low | 11 | 64 |
|
|
Discussion
The tumor microenvironment (TME) is comprised of
macrophages, fibroblasts, lymphocytes, endothelial cells,
adipocytes, perivascular cells, neurons and extracellular matrix
(ECM) components, and plays an important role in cancer invasion
and metastasis (4). TAMs are key
orchestrators and a set of macrophages of the TME (6,7). TAMs
play a critical role in regulating tumor growth and progression
(8,9). The high density of TAMs is correlated
with a poor prognosis in various types of cancer including gastric
cancer (GC) (9–15). However, the role of TAMs in GC and
the underlying mechanism remain elusive.
In the present study, we used THP-1 cells as a
substitute to investigate the impact of TAMs on GC cells. Transwell
invasion and wound healing assays indicated that co-cultured GC
cells with TAMs significantly promoted GC cell invasion and
migration. Loss of epithelial marker E-cadherin expression and gain
of mesenchymal marker vimentin expression is a major hallmark of
EMT (34). Aberrant reactivation of
EMT promoted tumor cell migration and invasion (35). In the present study, E-cadherin
expression in GC cells co-cultured with TAMs was decreased, while
vimentin expression in GC cells co-cultured with TAMs was
increased. This indicated that GC cells co-cultured with TAMs
underwent EMT. These results revealed that TAMs promoted GC cell
invasion and migration through EMT.
FOXQ1 is a member of the large forkhead (FOX)
transcription factor family (36).
It is expressed in different tissues and plays an important role in
development, metabolism, aging and cancer (37–39).
It is an ~42 kDa protein initially shown to be involved in hair
follicle differentiation, gastrulation and mucin production in mice
(40–42). Several recent studies demonstrated
that increased FOXQ1 expression in many human types of cancer,
including esophageal, breast and colorectal cancer, hepatocellular
carcinoma and non-small cell lung cancer, was correlated with
metastasis and poor prognosis (25–27,29,30,43–48).
FOXQ1 has been shown to be involved in the regulation of EMT
(37). In breast cancer and
non-small cell lung cancer, FOXQ1 has been shown to promote EMT by
regulating the expression of E-cadherin, β-catenin and vimentin
(26,27,30).
In hepatocellular carcinoma, FOXQ1 was reported to induce EMT and
enhance the invasive capability by activating transcription ZEB2
and Versican V1 (45). In bladder
cancer, knockdown of FOXQ1 inhibited invasion and metastasis via
the reversal of EMT (28). These
studies indicated that FOXQ1 plays an important role in EMT, and
subsequently in the invasion and metastasis of many types of
cancer. A recent study reported that FOXQ1 promoted GC metastasis
through upregulation of Snail (32).
We hypothesized that TAMs promote GC cell EMT,
invasion and migration through the FOXQ1 pathway. RT-PCR and
western blot results revealed that co-culture with TAMs
significantly increased FOXQ1 expression at both the mRNA and
protein levels of GC cells, which indicated that FOXQ1 was involved
in the EMT of GC cells induced by TAMs. Then, FOXQ1 shRNA was used
to silence the expression of FOXQ1. The results revealed that
promotion of TAMs in EMT was suppressed. Meanwhile, the promotion
of TAMs on the abilities of migration and invasion in GC cells were
also inhibited.
Furthermore, we investigated the correlation of TAM
marker CD68 and FOXQ1 expression in GC tissues. In accordance with
the present in vitro study, the result revealed that high
expression of CD68 was correlated with positive FOXQ1 expression
(r=0.613; P<0.001) in clinical GC samples.
In conclusion, our data provided evidence that TAMs
promote EMT, invasion and migration of GC cells via FOXQ1.
Therefore, the TAM/FOXQ1 axis may represent a novel target for GC
cells.
Acknowledgements
The present study was supported by the National
Natural Science Foundation of China (no. 81301981), and the
Fundamental Research Funds for the Central Universities.
References
|
1
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
DeSantis CE, Lin CC, Mariotto AB, Siegel
RL, Stein KD, Kramer JL, Alteri R, Robbins AS and Jemal A: Cancer
treatment and survivorship statistics, 2014. CA Cancer J Clin.
64:252–271. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zeng H, Zheng R, Guo Y, Zhang S, Zou X,
Wang N, Zhang L, Tang J, Chen J, Wei K, et al: Cancer survival in
China, 2003–2005: A population-based study. Int J Cancer.
136:1921–1930. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Whiteside TL: The tumor microenvironment
and its role in promoting tumor growth. Oncogene. 27:5904–5912.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lee CH, Liu SY, Chou KC, Yeh CT, Shiah SG,
Huang RY, Cheng JC, Yen CY and Shieh YS: Tumor-associated
macrophages promote oral cancer progression through activation of
the Axl signaling pathway. Ann Surg Oncol. 21:1031–1037. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Quail DF and Joyce JA: Microenvironmental
regulation of tumor progression and metastasis. Nat Med.
19:1423–1437. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
7
|
Joyce JA and Pollard JW:
Microenvironmental regulation of metastasis. Nat Rev Cancer.
9:239–252. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Fan QM, Jing YY, Yu GF, Kou XR, Ye F, Gao
L, Li R, Zhao QD, Yang Y, Lu ZH, et al: Tumor-associated
macrophages promote cancer stem cell-like properties via
transforming growth factor-beta1-induced epithelial-mesenchymal
transition in hepatocellular carcinoma. Cancer Lett. 352:160–168.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hu Y, He MY, Zhu LF, Yang CC, Zhou ML,
Wang Q, Zhang W, Zheng YY, Wang DM, Xu ZQ, et al: Tumor-associated
macrophages correlate with the clinicopathological features and
poor outcomes via inducing epithelial to mesenchymal transition in
oral squamous cell carcinoma. J Exp Clin Cancer Res. 35:122016.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhang BC, Gao J, Wang J, Rao ZG, Wang BC
and Gao JF: Tumor-associated macrophages infiltration is associated
with peritumoral lymphangiogenesis and poor prognosis in lung
adenocarcinoma. Med Oncol. 28:1447–1452. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Suriano F, Santini D, Perrone G, Amato M,
Vincenzi B, Tonini G, Muda A, Boggia S, Buscarini M and Pantano F:
Tumor associated macrophages polarization dictates the efficacy of
BCG instillation in non-muscle invasive urothelial bladder cancer.
J Exp Clin Cancer Res. 32:872013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yan Y, Zhang J, Li JH, Liu X, Wang JZ, Qu
HY, Wang JS and Duan XY: High tumor-associated macrophages
infiltration is associated with poor prognosis and may contribute
to the phenomenon of epithelial-mesenchymal transition in gastric
cancer. Onco Targets Ther. 9:3975–3983. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Mei J, Xiao Z, Guo C, Pu Q, Ma L, Liu C,
Lin F, Liao H, You Z and Liu L: Prognostic impact of
tumor-associated macrophage infiltration in non-small cell lung
cancer: A systemic review and meta-analysis. Oncotarget.
7:34217–34228. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhang J, Yan Y, Yang Y, Wang L, Li M and
Wang J, Liu X, Duan X and Wang J: High infiltration of
tumor-associated macrophages influences poor prognosis in human
gastric cancer patients, associates with the phenomenon of EMT.
Medicine. 95:e26362016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhang H, Wang X, Shen Z, Xu J, Qin J and
Sun Y: Infiltration of diametrically polarized macrophages predicts
overall survival of patients with gastric cancer after surgical
resection. Gastric Cancer. 18:740–750. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Thiery JP: Epithelial-mesenchymal
transitions in tumour progression. Nat Rev Cancer. 2:442–454. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Bonde AK, Tischler V, Kumar S, Soltermann
A and Schwendener RA: Intratumoral macrophages contribute to
epithelial-mesenchymal transition in solid tumors. BMC Cancer.
12:352012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Liu CY, Xu JY, Shi XY, Huang W, Ruan TY,
Xie P and Ding JL: M2-polarized tumor-associated macrophages
promoted epithelial-mesenchymal transition in pancreatic cancer
cells, partially through TLR4/IL-10 signaling pathway. Lab Invest.
93:844–854. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ravi J, Elbaz M, Wani NA, Nasser MW and
Ganju RK: Cannabinoid receptor-2 agonist inhibits macrophage
induced EMT in non-small cell lung cancer by downregulation of EGFR
pathway. Mol Carcinog. 55:2063–2076. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Deng YR, Liu WB, Lian ZX, Li X and Hou X:
Sorafenib inhibits macrophage-mediated epithelial-mesenchymal
transition in hepatocellular carcinoma. Oncotarget. 7:38292–38305.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Su S, Liu Q, Chen J, Chen J, Chen F, He C,
Huang D, Wu W, Lin L, Huang W, et al: A positive feedback loop
between mesenchymal-like cancer cells and macrophages is essential
to breast cancer metastasis. Cancer Cell. 25:605–620. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Techasen A, Loilome W, Namwat N, Dokduang
H, Jongthawin J and Yongvanit P: Cytokines released from activated
human macrophages induce epithelial mesenchymal transition markers
of cholangiocarcinoma cells. Asian Pac J Cancer Prev. 13
(Suppl):115–118. 2012.PubMed/NCBI
|
|
23
|
Zhang J, Yao H, Song G, Liao X, Xian Y and
Li W: Regulation of epithelial-mesenchymal transition by
tumor-associated macrophages in cancer. Am J Transl Res.
7:1699–1711. 2015.PubMed/NCBI
|
|
24
|
Zhang M, Xu Q, Yan S, Li Z, Yan W and Jia
X: Suppression of forkhead box Q1 by microRNA-506 represses the
proliferation and epithelial-mesenchymal transition of cervical
cancer cells. Oncol Rep. 35:3106–3114. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Qiao Y, Jiang X, Lee ST, Karuturi RK, Hooi
SC and Yu Q: FOXQ1 regulates epithelial-mesenchymal transition in
human cancers. Cancer Res. 71:3076–3086. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhang H, Meng F, Liu G, Zhang B, Zhu J, Wu
F, Ethier SP, Miller F and Wu G: Forkhead transcription factor
foxq1 promotes epithelial-mesenchymal transition and breast
cancer metastasis. Cancer Res. 71:1292–1301. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Feng J, Zhang X, Zhu H, Wang X, Ni S and
Huang J: FoxQ1 overexpression influences poor prognosis in
non-small cell lung cancer, associates with the phenomenon of EMT.
PLoS One. 7:e399372012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhu Z, Zhu Z, Pang Z, Xing Y, Wan F, Lan D
and Wang H: Short hairpin RNA targeting FOXQ1 inhibits invasion and
metastasis via the reversal of epithelial-mesenchymal transition in
bladder cancer. Int J Oncol. 42:1271–1278. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Sehrawat A, Kim SH, Vogt A and Singh SV:
Suppression of FOXQ1 in benzyl isothiocyanate-mediated inhibition
of epithelial-mesenchymal transition in human breast cancer cells.
Carcinogenesis. 34:864–873. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Feng J, Xu L, Ni S, Gu J, Zhu H, Wang H,
Zhang S, Zhang W and Huang J: Involvement of FoxQ1 in NSCLC through
regulating EMT and increasing chemosensitivity. Oncotarget.
5:9689–9702. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Fan DM, Feng XS, Qi PW and Chen YW:
Forkhead factor FOXQ1 promotes TGF-β1 expression and induces
epithelial-mesenchymal transition. Mol Cell Biochem. 397:179–186.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhang J, Liu Y, Zhang J, Cui X, Li G, Wang
J, Ren H and Zhang Y: FOXQ1 promotes gastric cancer metastasis
through upregulation of Snail. Oncol Rep. 35:3607–3613. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Xiang XJ, Deng J, Liu YW, Wan LY, Feng M,
Chen J and Xiong JP: MiR-1271 inhibits cell proliferation, invasion
and EMT in gastric cancer by targeting FOXQ1. Cell Physiol Biochem.
36:1382–1394. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Thiery JP, Acloque H, Huang RY and Nieto
MA: Epithelial-mesenchymal transitions in development and disease.
Cell. 139:871–890. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Thiery JP and Sleeman JP: Complex networks
orchestrate epithelial-mesenchymal transitions. Nat Rev Mol Cell
Biol. 7:131–142. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
36
|
Myatt SS and Lam EW: The emerging roles of
forkhead box (Fox) proteins in cancer. Nat Rev Cancer. 7:847–859.
2007. View
Article : Google Scholar : PubMed/NCBI
|
|
37
|
Feuerborn A, Srivastava PK, Küffer S,
Grandy WA, Sijmonsma TP, Gretz N, Brors B and Gröne HJ: The
Forkhead factor FoxQ1 influences epithelial differentiation. J Cell
Physiol. 226:710–719. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Eijkelenboom A and Burgering BM: FOXOs:
Signalling integrators for homeostasis maintenance. Nat Rev Mol
Cell Biol. 14:83–97. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Jonsson H and Peng SL: Forkhead
transcription factors in immunology. Cell Mol Life Sci. 62:397–409.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Goering W, Adham IM, Pasche B, Manner J,
Ochs M, Engel W and Zoll B: Impairment of gastric acid secretion
and increase of embryonic lethality in Foxq1-deficient mice.
Cytogenet Genome Res. 121:88–95. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Hong HK, Noveroske JK, Headon DJ, Liu T,
Sy MS, Justice MJ and Chakravarti A: The winged helix/forkhead
transcription factor Foxq1 regulates differentiation of hair
in satin mice. Genesis. 29:163–171. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Potter CS, Peterson RL, Barth JL, Pruett
ND, Jacobs DF, Kern MJ, Argraves WS, Sundberg JP and Awgulewitsch
A: Evidence that the satin hair mutant gene Foxq1 is among
multiple and functionally diverse regulatory targets for
Hoxc13 during hair follicle differentiation. J Biol Chem.
281:29245–29255. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Kaneda H, Arao T, Tanaka K, Tamura D,
Aomatsu K, Kudo K, Sakai K, De Velasco MA, Matsumoto K, Fujita Y,
et al: FOXQ1 is overexpressed in colorectal cancer and enhances
tumorigenicity and tumor growth. Cancer Res. 70:2053–2063. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Wang W, He S, Ji J, Huang J, Zhang S and
Zhang Y: The prognostic significance of FOXQ1 oncogene
overexpression in human hepatocellular carcinoma. Pathol Res Pract.
209:353–358. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Xia L, Huang W, Tian D, Zhang L, Qi X,
Chen Z, Shang X, Nie Y and Wu K: Forkhead box Q1 promotes
hepatocellular carcinoma metastasis by transactivating ZEB2 and
VersicanV1 expression. Hepatology. 59:958–973. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Pei Y, Wang P, Liu H, He F and Ming L:
FOXQ1 promotes esophageal cancer proliferation and metastasis by
negatively modulating CDH1. Biomed Pharmacother. 74:89–94. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Peng X, Luo Z, Kang Q, Deng D, Wang Q,
Peng H, Wang S and Wei Z: FOXQ1 mediates the crosstalk between
TGF-β and Wnt signaling pathways in the progression of colorectal
cancer. Cancer Biol Ther. 16:1099–1109. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Liang SH, Yan XZ, Wang BL, Jin HF, Yao LP,
Li YN, Chen M, Nie YZ, Wang X, Guo XG, et al: Increased expression
of FOXQ1 is a prognostic marker for patients with gastric cancer.
Tumour Biol. 34:2605–2609. 2013. View Article : Google Scholar : PubMed/NCBI
|