Introduction
Breast cancer is the most prevalent cancer and the
second most frequent cause of death in women (1). Approximately 10–30% of patients with
breast cancer are diagnosed with brain metastasis, which represents
a particularly devastating consequence of breast cancer due to its
high mortality (1–3). Although systemic therapy for breast
cancer patients has been improved and overall survival has been
prolonged, the incidence of brain metastasis is increasing. No
effective treatment for breast cancer brain metastasis (BCBM) is
available at present, and the overall survival is on the order of
months (mean 17 and median 15 months) (4). Therefore, there is an urgent need to
better understand the molecular basis of BCBM and explore new
therapeutic agents (3,5–8).
BCBM is a complex process that involves a series of
well-defined steps. To better understand BCBM, ‘brain-seeking’
clonal sublines have been generated (4,9,10).
Among them, MDA-MB-231-BR (231-BR), the brain-seeking clones of
MDA-MB-231, was derived by multiple selection rounds of brain
metastatic lesions after intracardiac injection of MDA-MB-231 cells
(9). Different from its parental
cell line MDA-MB-231 which is highly metastatic but has no organ
specificity, 231-BR metastasizes to the brain with 100% accuracy,
making it a widely used preclinical model for brain metastatic
breast cancer (11–16). On account of their syngeneic nature,
proteome of the brain-seeking cell line has been compared with its
parental cell lines, with the purpose of finding differentially
expressed proteins that may play roles in the establishment or
progression of BCBM (11). A few
genes or proteins have been discovered, while the possibility that
they can be served as potential biomarkers or targets for BCBM has
not been confirmed yet (4,11,17).
Among them is the multifunctional cytokine transforming growth
factor-β1 (TGF-β1), which has been extensively studied due to its
various effects on carcinoma cell populations (18).
Initially known as TGF-β1 stimulated clone 36
(TSC-36), Follistatin-like 1 (FSTL1) was first isolated from a
mouse osteoblast cell line as a TGF-β1-inducible gene (19). FSTL1 was reported as a regulator in
embryonic organogenesis (20), a
pro-inflammatory protein in rheumatoid arthritis (21), and a cardioprotective factor against
ischemic injury (22). In the past
decades, accumulating evidence has been obtained, suggesting a role
of FSTL1 in cancer (23–28), while its functions are still poorly
understood. Also, the signaling pathways involved in the expression
of FSTL1 remain to be determined. Various, sometimes contradictory
effects of FSTL1 have been demonstrated on cancer cell growth and
survival (23–28). To date, the expression and the
function of FSTL1 in breast cancer or BCBM has not been
investigated. Only one study mentioned that the number of
ALCAM+ cells was correlated with the amount of tumor
bone metastasis in a murine model, and ALCAM+ cells
correlatively increased in FSTL1+ tumor tissues of
patients with advanced breast cancer, which might suggest a
possible role of FSTL1 in breast cancer, while further validation
is required and signaling pathways involved need to be determined
(24).
To determine the expression of FSTL1 in breast
cancer, we detected its expression in four breast cancer cell
lines, as well as a brain metastatic breast cancer cell line
231-BR. A higher level of FSTL1 was detected in 231-BR, meanwhile
this protein was hardly detected in the other four breast cancer
cell lines including the 231-BR's parental cell line (MDA-MB-231).
These observations motivated us to investigate the possible role of
FSTL1 in breast cancer progression.
In the present study, the expression of FSTL1 was
determined in breast cancer cell lines. We reported for the first
time that the increased expression of FSTL1 in 231-BR is
accompanied with a decreased cell proliferation rate in MDA-MB-231.
Our data identified FSTL1 as an inhibitor of metastatic breast
cancer cell proliferation, which may provide new insights into the
development and management of breast cancer and BCBM.
Materials and methods
Cell culture
The human MDA-MB-231-BR ‘brain-seeking’ breast
cancer cell line (231-BR cells) was described previously (9). The 231-BR cell line transfected with
enhanced green fluorescent protein (EGFP) was kindly provided by Dr
Patricia S. Steeg (National Cancer Institute, Bethesda, MD, USA)
(9). MDA-MB-231, and MCF7 cell
lines were kindly provided by Dr Jun Wan (Hong Kong University of
Science and Technology, Hong Kong, China). ZR-75-1, HCC38 cell
lines were obtained from Dr Haili Qian (Chinese Academy of Medical
Sciences, Beijing, China). MDA-MB-231, 231-BR and MCF7 cells were
cultured in high-glucose Dulbecco's modified Eagle's medium (DMEM)
(Gibco, Grand Island, NY, USA) containing 10% fetal bovine serum
(Gibco) and 1% penicillin-streptomycin (Gibco). ZR-75-1 and HCC38
cells were cultured in RPMI-1640 (Gibco) with 10% fetal bovine
serum (Gibco) and 1% penicillin-streptomycin (Gibco). All cells
were cultured in 5% CO2 at 37°C.
Lentiviral construction
To stably overexpress FSTL1 in MDA-MB-231 cells,
lentiviral vector (Ubi-MCS-3flag-RFP-IRES-Puromycin) containing a
FSTL1 coding sequence (NM_007085) was constructed (GeneChem,
Shanghai, China). The empty vector was used as a control. To stably
knock down FSTL1 in 231-BR cells, lentiviral vector
(U6-MCS-Ubiquitin-Cherry-IRES-puromycin) containing the
short-hairpin RNA (shRNA) specifically targeting FSTL1 or a
negative control sequence was constructed (GeneChem). The following
shRNA sequence targeting FSTL1 was used: TAAGGAGCAAATCCAAGAT. The
following negative control sequence was used:
TTCTCCGAACGTGTCACGT.
MTT
Cells were seeded into 96-well plate with the
density of 2,000 cells per well at 37°C (day 0), allowed to adhere
overnight (day 1) or continue in culture for another 24 h, 48 h, 72
h or 96 h. The cell culture medium was removed, and cells were
incubated with 5 mg/ml 3-
(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)
(Sigma-Aldrich, St. Louis, MO, USA) for 3 h, and then 100 µl MTT
solution (10% Triton X-100, 0.1 N 37.5% HCl, 90% isopropanol) was
added per well. Absorbance was measured at 570 nm. Data were
normalized to day 1.
Real-time cellular analysis
(RTCA)
To examine the proliferation of cells, E-plates and
RTCA (RTCA-DP; ACEA Biosciences, USA) instrument was facilitated as
described previously (29).
Briefly, cells were seeded at a density of 2,000 cells per well in
E-plates (3 wells repeated). Cell index was recorded every 15 min
for 96 h.
Colony forming assay
Colony forming assay was performed to examine the
proliferation ability of cells. Cells were seeded into 6-well
plates at 400 cells per well (3 wells repeated), then cultured in
5% CO2 incubator. After 7 days, the plates were washed
with 0.01 M PBS and stained by 0.1% crystal violet (Sigma-Aldrich)
for 10 min. The plates were washed with 0.01 M PBS 3 times and
photographed using a microscope.
Western blotting
Cells were washed with ice-cold 0.01 M PBS, lysed in
the RIPA buffer with protease inhibitor and protein phosphatase
inhibitor for 30 min on ice. Then cells were scraped and
centrifuged with 12,000 rcf for 15 min. Protein concentration was
determined with BCA Protein assay kit (Thermo Scientific, Rockford,
IL, USA). The proteins were separated by SDS-PAGE and transferred
onto PVDF membrane. The primary antibodies used were Human FSTL1
Antibody (1:200, AF1694; R&D Systems, Minneapolis, MN, USA),
GAPDH (1:3,000, #5174; Cell Signaling Technology Inc., Beverly, MA,
USA), Smad2/3 (D7G7) XP® Rabbit mAb (1:1,000, #8685;
Cell Signaling Technology Inc.), anti-phospho-Smad2/3 (pThr8)
(1:1,000, SAB4504208; Sigma-Aldrich), anti-Smad1/5/8 antibody
(1:200, sc-6031-R; Santa Cruz Biotechnology, Santa Cruz, CA, USA);
anti-p-Smad1/5/8 (Ser 463/Ser 465) antibody (1:200, sc-12353; Santa
Cruz Biotechnology); anti-Smad3 antibody (1:1,000, ab28379; Abcam,
Cambridge, MA, USA); anti-Smad3 (phospho S423+S425) antibody
[EP823Y] (1:2,000, ab52903; Abcam); caspase-3 antibody (1:1,000,
#9662; Cell Signaling Technology Inc.); cleaved caspase-3 (Asp175)
(5A1E) (1:1,000, #9664; Cell Signaling Technology Inc.).
Real time-PCR
Total cellular RNA was extracted using Tri reagent
(Sigma-Aldrich), and cDNA was generated using SuperRT cDNA kit
(CWBio, China) as previously described (30). Specific primers for human FSTL1
(forward, 5′-CCAGACCACGATGTGGAAAC-3′; reverse,
5′-GGCACAGATCTTGGATTTGC-3′) and β-actin (forward,
5′-ACCTTCTACAATGAGCTGCG-3′; reverse, 5′-CCTGGATAGCAACGTACATGG-3′)
were used.
Transfection of siRNAs
Specific siRNAs for silencing Smad3, the negative
control siRNA (si-NC) and transfection kit were purchased from
Ribobio (stQ0007005-1; Guangzhou, China). Cells were seeded in
24-well plate with the density of 105/well. After 24 h,
465.75 µl cell culture medium, 30 µl 1X ribo FECT™ CP buffer, 1.25
µl siRNA and 3 µl ribo FECT™ CP reagent were added into each well.
Cells were cultured in 5% CO2 at 37°C for 48 h and then
the expression of Smad3 was detected.
Xenograft tumor
Female immune-deficient BALB/C-nude mice (4 weeks
old, 15–25 g body weight) were kept under controlled temperature
and humidity with light-dark cycles in the animal room. A single
cell suspension of 8×107 cells were subcutaneously
inoculated into each mouse. The volume of tumors was calculated (VT
= length × width2 × 0.5) every other day from the tumor
observation. The nude mice were euthanized and the xenograft tumors
were harvested 12 days after implantation.
All animal experimental procedures were approved by
Laboratory animal ethics committee of Capital Medical University
(Beijing, China). The results of all animal experiments are
reported in accordance with the ARRIVE guidelines.
Immunohistochemistry (IHC)
staining
Human tumor samples from patients with breast cancer
or BCBM were obtained from Cancer Hospital of Huanxing (Beijing,
China). Tissues were harvested, fixed, paraffin-embedded,
sectioned, and photographed. The sections were stained with
hematoxylin. The expression of FSTL1 and Ki67 was determined using
IHC staining. The primary antibody used was polyclonal goat anti-
human FSTL1 antibody (5 µg/ml, AF1694; R&D Systems) and rabbit
anti-Ki67 antibody (1:250, ab16667; Abcam).
This study was approved by Cancer Hospital of
Huanxing review board, and informed consent was obtained from all
patients under the protocols prescribed by Cancer Hospital of
Huanxing ethics committee. All procedures performed involving human
participants were in accordance with the ethical standards of the
Institutional and National Research Committee and with the 1964
Helsinki Declaration and its later amendments or comparable ethical
standards.
Statistical analysis
Distributions of the data was tested for normality
using D'Agostino and Pearson omnibus normality test. Statistically
significant differences between two groups were determined by
Student's t-test, and data are presented as the mean ± SD. The
non-parametric Wilcoxon rank-sum test was used when the data were
not normally distributed, and data are presented as median ± the
range. All statistical analyses were performed in GraphPad Prism 5
(GraphPad Software, San Diego, CA, USA), and P<0.05 was
considered statistically significant.
Results
Expression of FSTL1 in breast cancer
cells and characterization of FSTL1 stable transfectants
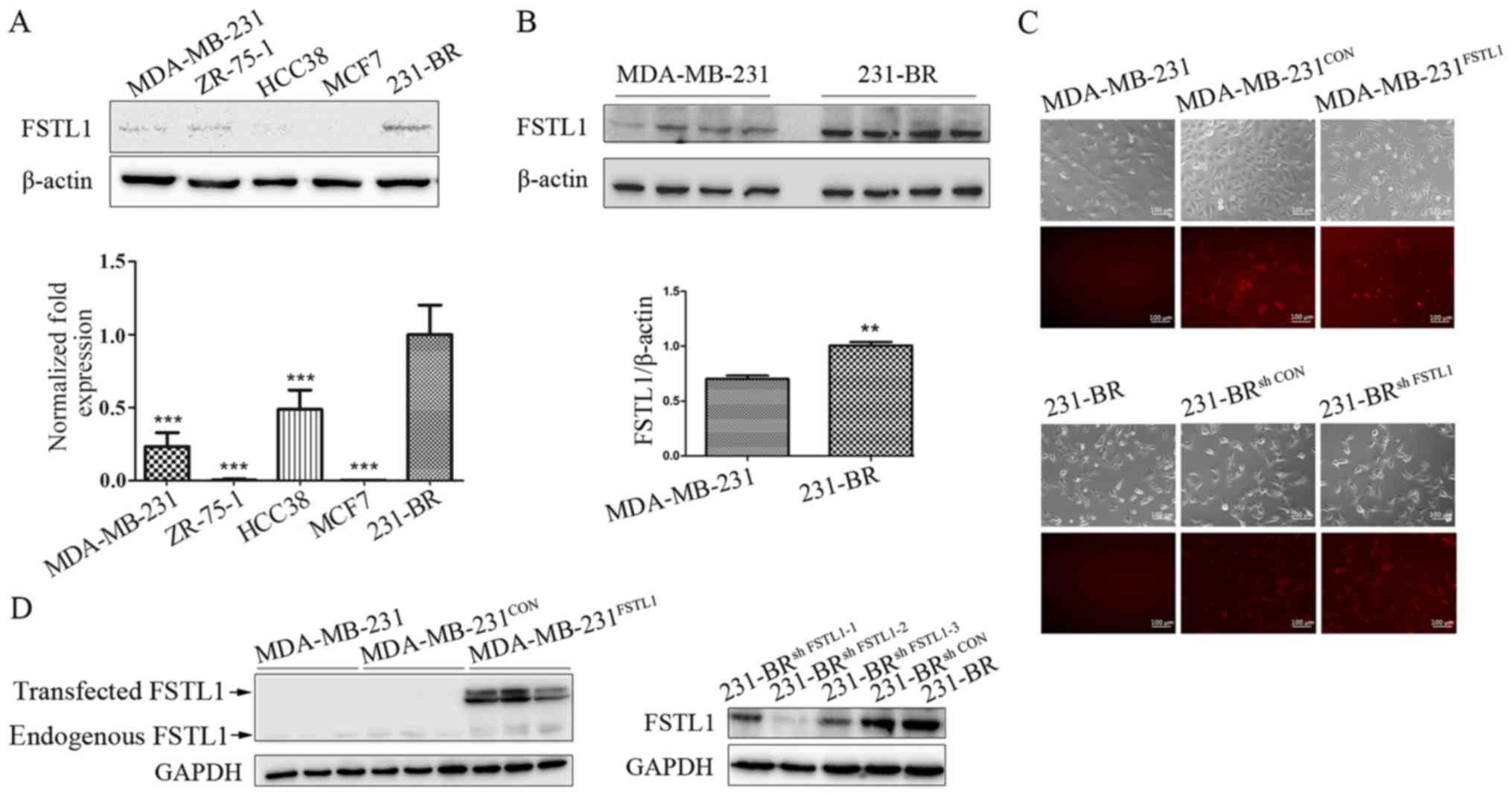
The protein and mRNA levels of FSTL1 in four breast
cancer cell lines MDA-MB-231, MCF7, ZR-75-1 and HCC38, and in the
specific brain metastatic cell line 231-BR were measured (Fig. 1A). FSTL1 was hardly detected in the
four breast cancer cell lines including MDA-MB-231, while it showed
an elevated expression in the 231-BR (Fig. 1B). To investigate the functional
role of FSTL1 in breast cancer cells, FSTL1 was overexpressed in
the MDA-MB-231 cells using red fluorescence labeled lentivirus, and
knocked down in 231-BR cells using Cherry fluorescence labeled
lentivirus. More than 90% of MDA-MB-231 control cells
(MDA-MB-231CON), FSTL1 overexpressed MDA-MB-231 cells
(MDA-MB-231FSTL1), 231-BR control cells (231-BRsh
CON) and FSTL1 knockdown 231-BR cells (231-BRsh
FSTL1) showed expression of RFP or Cherry fluorescence,
indicating a successful infection (Fig.
1C). The protein level of FSTL1 was obviously upregulated in
MDA-MB-231FSTL1 cells compared with control
(MDA-MB-231CON) (Fig.
1D, left panel), and downregulated in 231-BRsh FSTL1
cells compared with control (231-BRsh CON) (Fig. 1D, right panel).
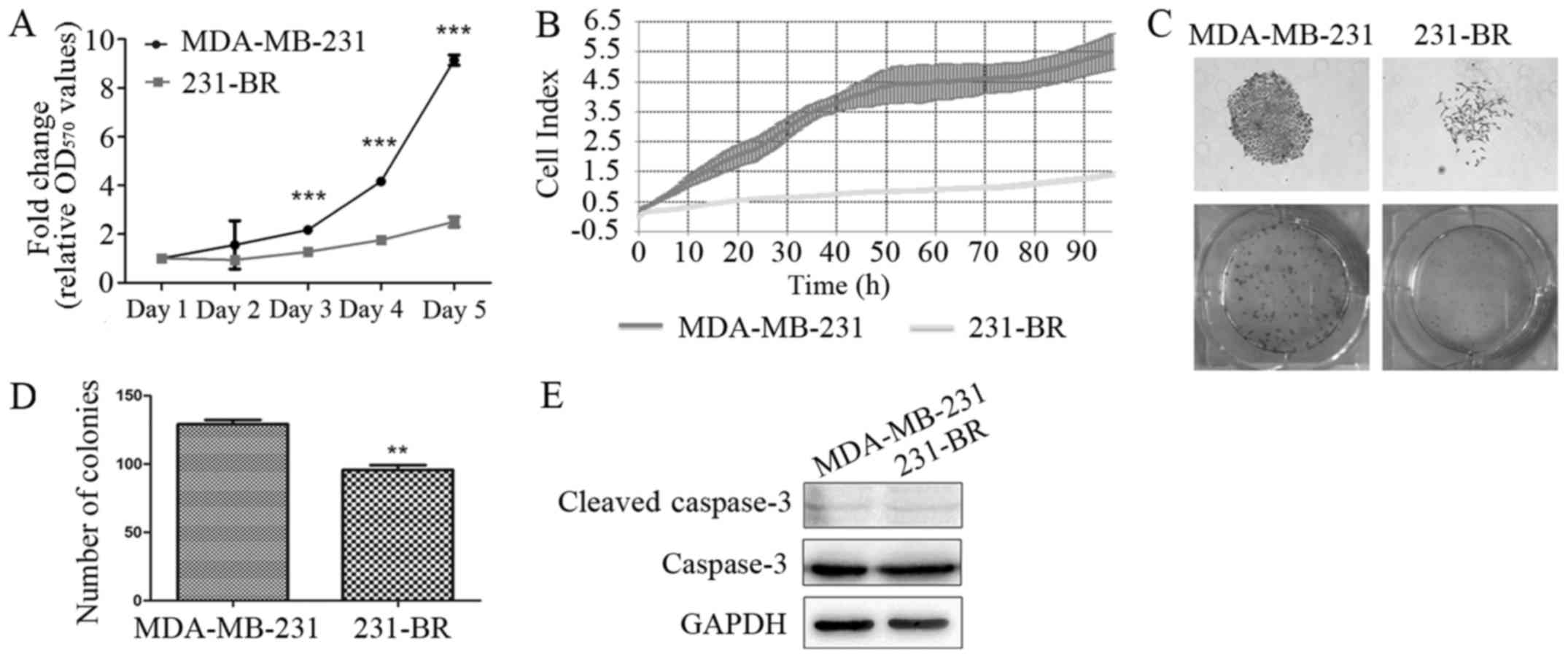
MDA-MB-231 cells show a higher
proliferation rate than 231-BR cells
To compare the proliferation rates between
MDA-MB-231 and 231-BR cells, MTT assay, RTCA and colony forming
assay were performed. As shown in Fig.
2A, MDA-MB-231 cells grew faster compared with 231-BR cells as
detected by MTT assay (3.65-fold on day 5, P<0.001). Similar
result was obtained in RTCA, that the cell growth of MDA-MB-231 was
3.9-fold faster than 231-BR on 96 h (P<0.001) (Fig. 2B). Furthermore, in colony forming
assay, more colonies were observed in MDA-MB-231 cells compared
with 231-BR (1.66-fold, P<0.01) (Fig. 2C and D).
In addition, apoptosis was investigated by measuring
protein level of cleaved caspase-3 through western blotting.
Results showed that the expression levels of cleaved caspase-3 in
MDA-MB-231 and 231-BR cells were comparable, indicating that the
inhibited cell growth in 231-BR cells was caused by suppressed cell
proliferation (Fig. 2E).
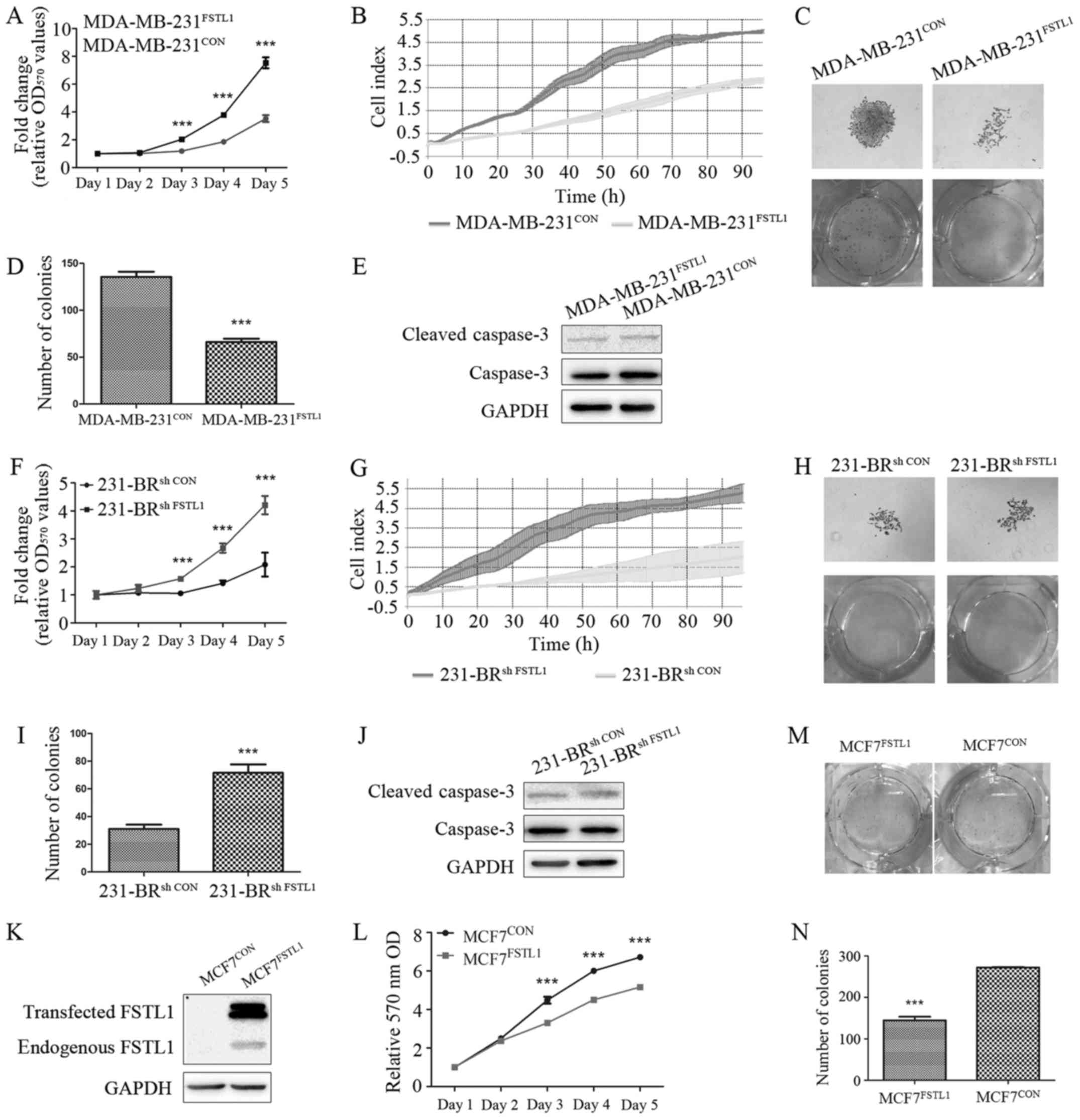
FSTL1 overexpression inhibits cell
proliferation of MDA-MB-231 and MCF7 cells in vitro
In order to elucidate the effect of FSTL1
overexpression on cell proliferation of MDA-MB-231 cells, MTT, RTCA
and colony forming assay were carried out for the transfected
MDA-MB-231 cells. After overexpression of FSTL1, the growth rate of
MDA-MB-231FSTL1 was reduced by 53.21% (P<0.001)
compared with MDA-MB-231CON as detected by MTT assay on
day 5 (Fig. 3A). Suppressed
proliferation of MDA-MB-231FSTL1 cells was also detected
in RTCA (42.29%, P<0.001) (Fig.
3B). In addition, in colony forming assay, the number of
MDA-MB-231FSTL1 cell colonies was reduced by 51.29%
compared with control (P<0.001) (Fig. 3C and D).
The protein level of cleaved caspase-3 was measured
to detect cell apoptosis, and no significant difference was
observed (Fig. 3E). To validate the
effect of FSTL1 on breast cancer cell proliferation, FSTL1 was also
overexpressed in MCF7 cells (Fig.
3K). Cell proliferation was evaluated by MTT and colony forming
assay and similar results were observed (Fig. 3L-N).
FSTL1 knockdown promotes cell
proliferation of 231-BR cells in vitro
MTT, RTCA and colony forming assay were also carried
out for the transfected 231-BR cells to explore the effect of FSTL1
knockdown on cell proliferation in 231-BR cells. As expected, the
knockdown of FSTL1 in 231-BR cells increased cell proliferation by
1.67-fold on day 5 (P<0.001), as detected by MTT assay (Fig. 3F). Consistently, RTCA showed that
the growth of 231-BRsh FSTL1 was 2.63-fold (P<0.001)
faster than 231-BRsh CON (Fig. 3G). In addition, knockdown of FSTL1
in 231-BR cells increased colony size and number in colony forming
assay (1.98-fold, P<0.001) (Fig. 3H
and I). Expression level of cleaved caspase-3 was also detected
in these transfected cell lines, and no significant difference was
observed (Fig. 3J).
FSTL1 inhibits the proliferation of
MDA-MB-231 and 231-BR cells by regulating TGF-β1 signaling pathway
through Smad3
TGF-β superfamily signaling pathway plays critical
roles in the regulation of cancer cell growth (31). In addition, FSTL1 has been showed to
activate TGF-β1-Smad2/3 signaling pathway in pulmonary fibrogenesis
(32) and myocardium (22), and inhibit BMP-Smad1/5/8 signaling
pathway in controlling mouse lung development (33). These observations motivated us to
question if there is an interaction between FSTL1 and TGF-β
signaling pathways that may contribute to the modification of cell
proliferation.
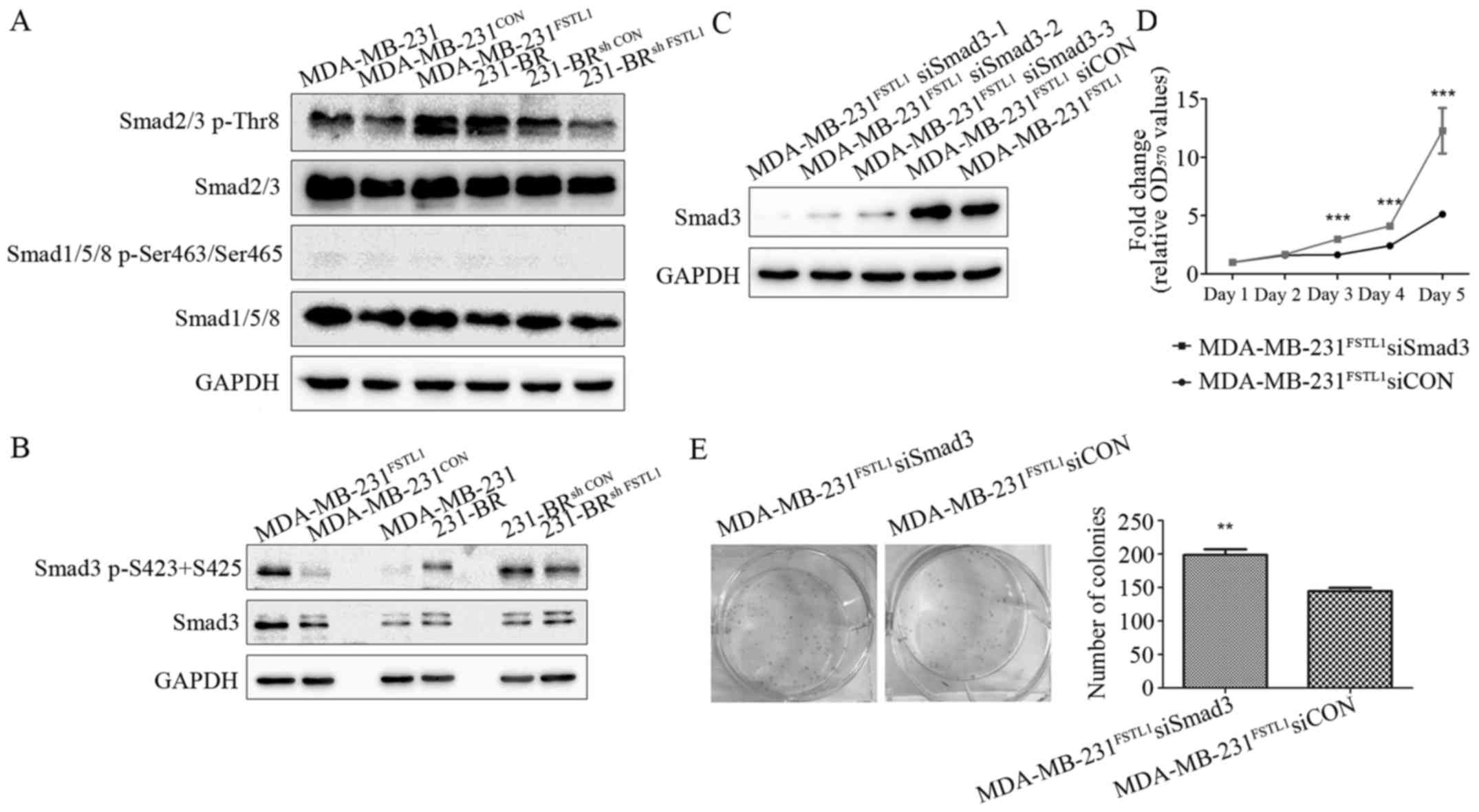
As shown in Fig. 4A,
lower phosphorylation of Smad2/3 was detected in MDA-MB-231 cells
than 231-BR cells. In the transfected cell lines, the
phosphorylation of Smad2/3 was increased after FSTL1 overexpression
in MDA-MB-231FSTL1 cells compared with
MDA-MB-231CON (Fig. 4A),
and decreased after FSTL1 knockdown in 231-BRsh FSTL1
cells compared with 231-BRsh CON cells (Fig. 4A). The phosphorylated Smad1/5/8 was
hardly detected in these cell lines, suggesting an inactivation of
BMP-Smad1/5/8 signaling pathway (Fig.
4A). These observations indicated that FSTL1 plays a role in
the regulation of TGF-β1-Smad2/3 signaling pathway, but not
BMP-Smad1/5/8, in MDA-MB-231 and 231-BR cells.
Considering that Smad3 plays a central role in TGF-β
signaling pathway, we performed further studies to test if FSTL1
regulated cell proliferation through the TGF-β pathway by
regulating Smad3. As shown in Fig.
4B, phosphorylation level of Smad3 was higher in 231-BR cells
than MDA-MB-231 cells. After transfection, the phosphorylation of
Smad3 was increased after FSTL1 overexpression (in
MDA-MB-231FSTL1 cells) and decreased after FSTL1
knockdown (in 231-BRsh FSTL1 cells), compared with their
corresponding controls. Next, Smad3 was knocked down by siRNAs in
MDA-MB-231FSTL1 cells. Expression levels of Smad3 were
measured using western blotting and the siRNA with highest silence
efficiency (siSmad3-1) was used for the following experiments
(Fig. 4C). As evaluated by MTT
assay, knockdown of Smad3 in MDA-MB-231FSTL1
significantly enhanced cell proliferation by 2.93-fold (P<0.001)
on day 5 compare with control (Fig.
4D). Colony forming assay was also carried out, showing that
colony number of MDA-MB-231FSTL1 cells was 1.37-fold
increased (P<0.01) after Smad3 knockdown (Fig. 4E). These results revealed that
knockdown of Smad3 restored the cell proliferation, which was
inhibited by FSTL1 overexpression in MDA-MB-231FSTL1
cells.
FSTL1 overexpression inhibits the
proliferation of MDA-MB-231 cells in vivo
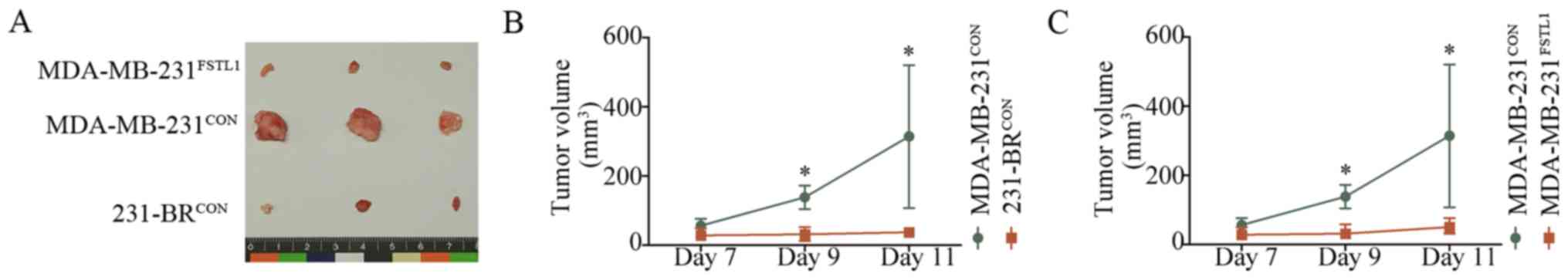
To investigate the effect of FSTL1 on tumor growth
in vivo, MDA-MB-231FSTL1,
MDA-MB-231CON and 231-BRCON cells were
inoculated subcutaneously into female nude mice. Tumor volume was
measured every other day from one week after cell injection. Tumors
were harvested and photographed on day 12 (Fig. 5A). Results showed that the tumor
volume in 231-BRCON cell-bearing mice was significantly
smaller than the xenografts formed by MDA-MB-231CON
cells (n=5, P<0.05) (Fig. 5B),
and reduction in tumor volume could be detected in
MDA-MB-231FSTL1 group compared with the control group
(n=5, P<0.05) (Fig. 5C). These
results indicated that MDA-MB-231 had a higher proliferation rate
than 231-BR cells and FSTL1 overexpression inhibited the
proliferation of MDA-MB-231 in vivo, which was in agreement
with the study in vitro.
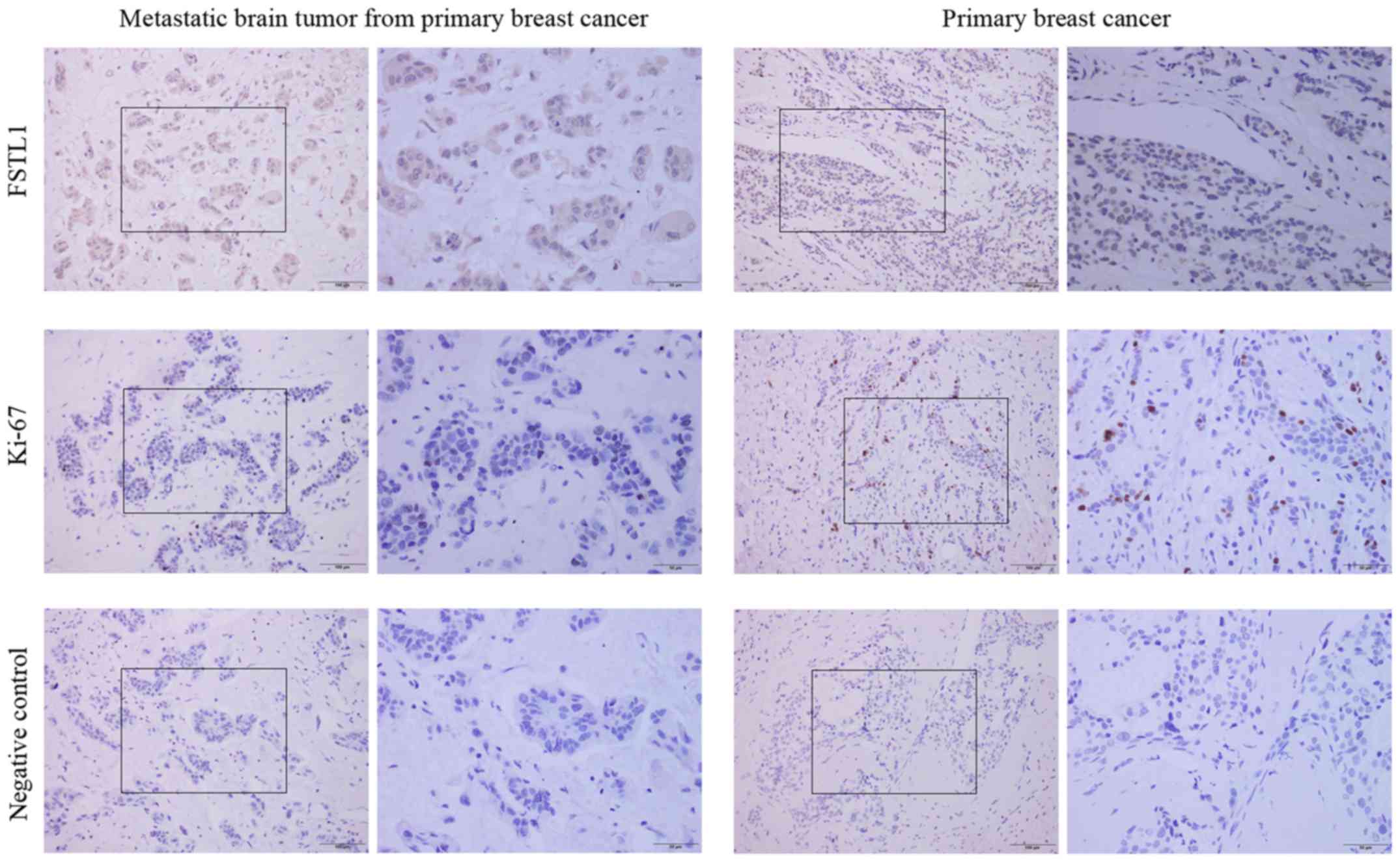
One metastatic brain tumor from
primary breast cancer of BCBM patient showed a high-level of FSTL1,
while low-level of this protein was detected in primary breast
cancer specimens
We wondered whether the correlation of FSTL1 and
proliferation observed in vitro and in vivo also
existed in human specimens, the FSTL1 expression was detected in
primary breast cancer specimens (n=5) and one metastatic brain
tumor (n=1) from primary breast cancer. As IHC staining
demonstrated (Fig. 6), the resected
metastatic brain tumor from primary breast cancer showed a higher
expression level of FSTL1; whereas the Ki67 expression was lower
than that in primary breast cancer. On the other hand, the five
specimens of primary breast cancer all showed a lower expression
level of FSTL1 and a higher expression of Ki67 (Fig. 6). This result showed a trend similar
to the results in vitro and in vivo.
The collection of brain metastatic tumors from BCBM
patients is difficult because only a small part of BCBM patients
with a single brain metastasis will undergo neurosurgical
resection. In this study, only one metastatic brain tumor from
primary breast cancer was obtained so that this result only showed
a trend which may not be significant. More evidence is required to
confirm this result.
Discussion
Aberrant expression and various functions of FSTL1
have been observed in tumor cell lines and clinical tumor tissues,
suggesting a role of FSTL1 in cancer. However, the underlying
mechanisms have rarely been studied and remain largely unknown. In
some studies, FSTL1 was identified as a tumor-suppressive protein.
It was found to be downregulated in metastatic clear-cell
renal-cell carcinoma and considered to be related to metastasis
(34). In ovarian and endometrial
carcinogenesis, downregulation of FSTL1 was detected and a tumor
suppressor role was demonstrated (25). In contrast, FSTL1 overexpression was
observed in glioblastoma as a hallmark of poor prognosis by Reddy
et al in 2010 (23). In that
study, analysis of retrospective GBM cases with known survival data
revealed that the coexpression of FSTL1 with p53 was associated
with poor survival. However, the functional role of FSTL1 in
astrocytomas was not investigated, and the signaling pathways
involved in the expression of FSTL1 remain to be determined
(23). To date, the functional
roles of FSTL1 in cancer remain controversial and unclear. The role
of FSTL1 in breast cancer or BCBM has not been investigated. Our
study is the first report documenting the increased level of FSTL1
in 231-BR cell line and linking the possible functions of FSTL1 in
breast cancer progression.
The signaling pathways of FSTL1 involved in cancer
are poorly defined. The following lines of evidence motivated us to
investigate the correlation between FSTL1 and TGF-β signaling
pathway in breast cancer cells. Firstly, as a TGF-β1-inducible
gene, FSTL1 encodes a secreted glycoprotein belonging to a
group of matricellular proteins (19). Two recent studies showed that it
activated TGF-β1-Smad2/3 signaling in pulmonary fibrogenesis
(32) and myocardium (22), respectively. Also, in lung
development, it can reduce the activity of TGF-β/BMP signaling
(33). These studies indicated a
role of FSTL1 in regulating TGF-β signaling pathways. Secondly, as
a pleiotropic cytokine, TGF-β signaling pathways regulates diverse
cellular processes in cancer, including apoptosis, cell growth and
epithelial-mesenchymal transition (35). A direct influence on breast cancer
pathophysiology by TGF-β1 was documented (18). It inhibits breast cancer cell growth
and promotes apoptosis in early stages, while it is related to
increased tumor progression in late stages. Moreover, TGF-β1 has
already been shown to inhibit the anchorage-independent growth of
MDA-MB-231 and 231-BR (9). Taken
these two points into consideration, we investigated the role of
FSTL1 in the regulation of TGF-β signaling pathways. We detected
Smad2/3-mediated TGF-β1 and Smad1/5/8-mediated TGF-β/BMP signaling
pathway in wild-type and transfected MDA-MB-231 and 231-BR cells.
We clarified that the TGF-β1-Smad2/3 signaling pathway was, at
least partly, the molecular mechanism whereby FSTL1 modulates the
cell proliferation rate. To date, it is still not clear if FSTL1
exerts its effects in autocrine and paracrine manner as a secreted
protein in cancer. In 2010, DIP2A was suggested to be a potential
receptor of FSTL1 that mediates the protective roles of
cardiomyocytes, while the signaling pathways involved in this
process were not clarified (36,37).
In addition, the roles of DIP2A in cancers have not been
investigated yet. Therefore, to detect the expression of DIP2A in
breast cancer cells and patient tissues, and to find out the
signaling pathways involved may also help to study the effects of
FSTL1 on breast cancer cell proliferation. This will be
investigated in a future study.
The metastatic cascade of BCBM involves a series of
well-defined steps including local invasion, intravasation,
survival in the circulation, extravasation, colonization and
proliferation, while the mechanisms underlying this complex process
are largely unknown. To date, majority of the preclinical studies
focuses on early stages of BCBM, to find the possible risk factors
for the development of brain metastases. However, the present study
may provide another view and demonstrate a likely role of FSTL1 in
the step of ‘proliferation’, which is the last step of BCBM cells
after they entered the brain. However, the effects of brain
microenvironment and its mechanical properties on breast cancer
cells should also be investigated.
Although FSTL1 showed an anti-proliferative effect
on breast cancer cells, it is too early to say if it is an
antitumor factor, and there is still a long way to go to clarify
the role of FSTL1 in breast cancer prognosis and to design drug for
breast cancer patient. In the present study, FSTL1 was suggested to
exert its effect through regulating TGF-β signaling, which could
inhibit cell growth in early stages and increased tumor progression
in late stages and has ‘two faces’ in cancer (18). Also, added to the inhibitory effect
of FSTL1 on cell proliferation and the overexpression of FSTL1 in
metastatic 231-BR cells observed in the present study, FSTL1 may
have dynamic roles in breast cancer. The effects of FSTL1 on cell
invasion, and cell migration, should also be taken into
consideration, which may affect the tumor malignant process.
Additionally, the association of FSTL1 level with BCBM clinical
prognosis needs to be evaluated. With the rapid development of the
molecular biology techniques, the use of genetic screens that
delineate the tumor-suppressive versus the possible tumor-promoting
roles of FSTL1 may provide a basis for new study that will enable
the targeting of its specific oncogenic sub-arms. At present,
studies focusing on the effect of FSTL1 on invasion and metastasis
of breast cancer cells are undergoing in our laboratory.
As the expression of FSTL1 in clinical biopsy
specimens of breast cancer patients has rarely been documented, we
tried to detect the expression of FSTL1 in human tissue. However,
patients with BCBM often bear multiple brain metastases that are
not suitable to undergo surgical resection. Moreover, the whole
brain radiation remains standard in the management of BCBM. Only
for a small part of BCBM patients with a single brain metastasis,
neurosurgical resection will be carried out followed by WBR
(38). It makes the collection of
brain metastatic tumors from BCBM patients difficult. In this
study, only one metastatic brain tumor from primary breast cancer
was obtained (Fig. 6) from a
patient with a single brain metastasis, and a higher expression
level of FSTL1 was observed (n=1). Meanwhile, the expression of
FSTL1 was much lower in breast cancer in situ (n=5). More
evidence is required from clinical data, and the specimens are
being collected continually.
In the present study, a high-level expression of
FSTL1 was first detected in the brain metastatic breast cancer cell
line 231-BR. An inhibitory effect of FSTL1 on cell proliferation
was determined in 231-BR and MDA-MB-231 cells in vitro and
in vivo. This anti-proliferative effect was further explored
to be associated with regulation of TGF-β signaling pathway through
Smad3. Our data define FSTL1 as an inhibitor of proliferation of
breast cancer cells, which may provide new insight into the
mechanism underlying BCBM.
Acknowledgements
We thank Dr Patricia S. Steeg (National Cancer
Institute, Bethesda, MD, USA) for the 231-BR cell line, Dr Jun Wan
(Hong Kong University of Science and Technology, Hong Kong, China)
for the MDA-MB-231 and MCF7 cell lines, and Dr Haili Qian (Cancer
Hospital Chinese Academy of Medical Sciences, Beijing, China) for
the ZR-75-1 and HCC38 cell lines. This study was supported by
National Natural Science Foundation of China, grant no. 81602532;
Beijing Natural Science Foundation, grant nos. 7164238 and 7152016;
Beijing Municipal Commission of Education, grant no.
KM201410025002; Beijing Municipal Organization Department Talents
Project, grant no. 2015000020124G113; Support Project of High-level
Teachers in Beijing Municipal Universities in the Period of 13th
Five-year Plan, grant no. IDHT20170516.
Glossary
Abbreviations
Abbreviations:
|
231-BR
|
MDA-MB-231-BR
|
|
BCBM
|
breast cancer brain metastases
|
|
BMP
|
bone morphogenetic protein
|
|
FSTL1
|
Follistatin-like 1
|
|
FRP
|
follistatin related protein
|
|
IHC
|
immunohistochemistry
|
|
RTCA
|
real-time cellular analysis
|
|
shRNA
|
short-hairpin RNA
|
|
TGF-β
|
transforming growth factor-β
|
|
TSC-36
|
TGF-β1 stimulated clone 36
|
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2016. CA Cancer J Clin. 66:7–30. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kienast Y, von Baumgarten L, Fuhrmann M,
Klinkert WE, Goldbrunner R, Herms J and Winkler F: Real-time
imaging reveals the single steps of brain metastasis formation. Nat
Med. 16:116–122. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Palmieri D, Smith QR, Lockman PR, Bronder
J, Gril B, Chambers AF, Weil RJ and Steeg PS: Brain metastases of
breast cancer. Breast Dis. 26:139–147. 2006-2007. View Article : Google Scholar
|
|
4
|
Kodack DP, Askoxylakis V, Ferraro GB,
Fukumura D and Jain RK: Emerging strategies for treating brain
metastases from breast cancer. Cancer Cell. 27:163–175. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Rostami R, Mittal S, Rostami P, Tavassoli
F and Jabbari B: Brain metastasis in breast cancer: A comprehensive
literature review. J Neurooncol. 127:407–414. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Sihto H, Lundin J, Lundin M, Lehtimäki T,
Ristimäki A, Holli K, Sailas L, Kataja V, Turpeenniemi-Hujanen T,
Isola J, et al: Breast cancer biological subtypes and protein
expression predict for the preferential distant metastasis sites: A
nationwide cohort study. Breast Cancer Res. 13:R872011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Gil-Gil MJ, Martinez-Garcia M, Sierra A,
Conesa G, Del Barco S, González-Jimenez S and Villà S: Breast
cancer brain metastases: A review of the literature and a current
multidisciplinary management guideline. Clin Transl Oncol.
16:436–446. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lam SW, Jimenez CR and Boven E: Breast
cancer classification by proteomic technologies: Current state of
knowledge. Cancer Treat Rev. 40:129–138. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yoneda T, Williams PJ, Hiraga T, Niewolna
M and Nishimura R: A bone-seeking clone exhibits different
biological properties from the MDA-MB-231 parental human breast
cancer cells and a brain-seeking clone in vivo and in vitro. J Bone
Miner Res. 16:1486–1495. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Stark AM, Anuszkiewicz B, Mentlein R,
Yoneda T, Mehdorn HM and Held-Feindt J: Differential expression of
matrix metalloproteinases in brain- and bone-seeking clones of
metastatic MDA-MB-231 breast cancer cells. J Neurooncol. 81:39–48.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Dun MD, Chalkley RJ, Faulkner S, Keene S,
Avery-Kiejda KA, Scott RJ, Falkenby LG, Cairns MJ, Larsen MR,
Bradshaw RA, et al: Proteotranscriptomic profiling of 231-BR breast
cancer cells: Identification of potential biomarkers and
therapeutic targets for brain metastasis. Mol Cell Proteomics.
14:2316–2330. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Fitzgerald DP, Subramanian P, Deshpande M,
Graves C, Gordon I, Qian Y, Snitkovsky Y, Liewehr DJ, Steinberg SM,
Paltán-Ortiz JD, et al: Opposing effects of pigment
epithelium-derived factor on breast cancer cell versus neuronal
survival: Implication for brain metastasis and metastasis-induced
brain damage. Cancer Res. 72:144–153. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Gril B, Palmieri D, Bronder JL, Herring
JM, Vega-Valle E, Feigenbaum L, Liewehr DJ, Steinberg SM, Merino
MJ, Rubin SD, et al: Effect of lapatinib on the outgrowth of
metastatic breast cancer cells to the brain. J Natl Cancer Inst.
100:1092–1103. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lockman PR, Mittapalli RK, Taskar KS,
Rudraraju V, Gril B, Bohn KA, Adkins CE, Roberts A, Thorsheim HR,
Gaasch JA, et al: Heterogeneous blood-tumor barrier permeability
determines drug efficacy in experimental brain metastases of breast
cancer. Clin Cancer Res. 16:5664–5678. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Palmieri D, Bronder JL, Herring JM, Yoneda
T, Weil RJ, Stark AM, Kurek R, Vega-Valle E, Feigenbaum L,
Halverson D, et al: Her-2 overexpression increases the metastatic
outgrowth of breast cancer cells in the brain. Cancer Res.
67:4190–4198. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Percy DB, Ribot EJ, Chen Y, McFadden C,
Simedrea C, Steeg PS, Chambers AF and Foster PJ: In vivo
characterization of changing blood-tumor barrier permeability in a
mouse model of breast cancer metastasis: A complementary magnetic
resonance imaging approach. Invest Radiol. 46:718–725.
2011.PubMed/NCBI
|
|
17
|
Chen G and Davies MA: Emerging insights
into the molecular biology of brain metastases. Biochem Pharmacol.
83:305–314. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zarzynska JM: Two faces of TGF-beta1 in
breast cancer. Mediators Inflamm. 2014:1417472014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Shibanuma M, Mashimo J, Mita A, Kuroki T
and Nose K: Cloning from a mouse osteoblastic cell line of a set of
transforming-growth-factor-beta 1-regulated genes, one of which
seems to encode a follistatin-related polypeptide. Eur J Biochem.
217:13–19. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Adams D, Larman B and Oxburgh L:
Developmental expression of mouse Follistatin-like 1 (Fstl1):
Dynamic regulation during organogenesis of the kidney and lung.
Gene Expr Patterns. 7:491–500. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kawabata D, Tanaka M, Fujii T, Umehara H,
Fujita Y, Yoshifuji H, Mimori T and Ozaki S: Ameliorative effects
of follistatin-related protein/TSC-36/FSTL1 on joint inflammation
in a mouse model of arthritis. Arthritis Rheum. 50:660–668. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Xi Y, Gong DW and Tian Z: FSTL1 as a
potential mediator of exercise-induced cardioprotection in
post-myocardial infarction rats. Sci Rep. 6:324242016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Reddy SP, Britto R, Vinnakota K, Aparna H,
Sreepathi HK, Thota B, Kumari A, Shilpa BM, Vrinda M, Umesh S, et
al: Novel glioblastoma markers with diagnostic and prognostic value
identified through transcriptome analysis. Clin Cancer Res.
14:2978–2987. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kudo-Saito C, Fuwa T, Murakami K and
Kawakami Y: Targeting FSTL1 prevents tumor bone metastasis and
consequent immune dysfunction. Cancer Res. 73:6185–6193. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chan QK, Ngan HY, Ip PP, Liu VW, Xue WC
and Cheung AN: Tumor suppressor effect of follistatin-like 1 in
ovarian and endometrial carcinogenesis: A differential expression
and functional analysis. Carcinogenesis. 30:114–121. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Chaly Y, Hostager B, Smith S and Hirsch R:
Follistatin-like protein 1 and its role in inflammation and
inflammatory diseases. Immunol Res. 59:266–272. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Chen SX, Xu XE, Wang XQ, Cui SJ, Xu LL,
Jiang YH, Zhang Y, Yan HB, Zhang Q, Qiao J, et al: Identification
of colonic fibroblast secretomes reveals secretory factors
regulating colon cancer cell proliferation. J Proteomics.
110:155–171. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Bae K, Park KE, Han J, Kim J, Kim K and
Yoon KA: Mitotic cell death caused by follistatin-like 1 inhibition
is associated with up-regulated Bim by inactivated Erk1/2 in human
lung cancer cells. Oncotarget. 7:18076–18084. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Tan J, Yang S, Shen P, Sun H, Xiao J, Wang
Y, Wu B, Ji F, Yan J, Xue H, et al: C-kit signaling promotes
proliferation and invasion of colorectal mucinous adenocarcinoma in
a murine model. Oncotarget. 6:27037–27048. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Meng X, Zheng R, Zhang Y, Qiao M, Liu L,
Jing P, Wang L, Liu J and Gao Y: An activated sympathetic nervous
system affects white adipocyte differentiation and lipolysis in a
rat model of Parkinson's disease. J Neurosci Res. 93:350–360. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wieser R: The transforming growth
factor-beta signaling pathway in tumorigenesis. Curr Opin Oncol.
13:70–77. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Dong Y, Geng Y, Li L, Li X, Yan X, Fang Y,
Li X, Dong S, Liu X, Li X, et al: Blocking follistatin-like 1
attenuates bleomycin-induced pulmonary fibrosis in mice. J Exp Med.
212:235–252. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Geng Y, Dong Y, Yu M, Zhang L, Yan X, Sun
J, Qiao L, Geng H, Nakajima M, Furuichi T, et al: Follistatin-like
1 (Fstl1) is a bone morphogenetic protein (BMP) 4 signaling
antagonist in controlling mouse lung development. Proc Natl Acad
Sci USA. 108:pp. 7058–7063. 2011; View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Tan X, Zhai Y, Chang W, Hou J, He S, Lin
L, Yu Y, Xu D, Xiao J, Ma L, et al: Global analysis of
metastasis-associated gene expression in primary cultures from
clinical specimens of clear-cell renal-cell carcinoma. Int J
Cancer. 123:1080–1088. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Massagué J: TGFbeta in cancer. Cell.
134:215–230. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Tanaka M, Murakami K, Ozaki S, Imura Y,
Tong XP, Watanabe T, Sawaki T, Kawanami T, Kawabata D, Fujii T, et
al: DIP2 disco-interacting protein 2 homolog A (Drosophila) is a
candidate receptor for follistatin-related protein/follistatin-like
1 - analysis of their binding with TGF-β superfamily proteins. FEBS
J. 277:4278–4289. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Ouchi N, Asaumi Y, Ohashi K, Higuchi A,
Sono-Romanelli S, Oshima Y and Walsh K: DIP2A functions as a FSTL1
receptor. J Biol Chem. 285:7127–7134. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Soffietti R, Rudà R and Trevisan E: Brain
metastases: Current management and new developments. Curr Opin
Oncol. 20:676–684. 2008. View Article : Google Scholar : PubMed/NCBI
|