Introduction
Breast cancer is the most common type of cancer and
a leading cause of mortality among women (1). Various molecular-targeted therapies of
breast cancer have been developed in the past decade (2–4) and
treatments have been diversified. However, development of the
resistance of cancer cells to chemotherapeutic agents remains an
important clinical issue. Since the major cause of recurrence is
chemoresistance to systemic therapies (5,6), the
mechanisms underlying chemoresistance need to be elucidated in more
detail.
The X-linked inhibitor of apoptosis (XIAP) is a
potent inhibitor of apoptosis proteins (7). XIAP binds to and inhibits caspases,
which are the principal inducers of apoptosis (8–11). In
addition, XIAP has been associated with drug resistance in various
types of cancer such as breast (12,13),
bladder (14) and ovarian cancer
(15). In breast cancer cells, XIAP
is significantly upregulated in cells that are resistant to
anthracycline or taxane (12),
which are mainly used in adjuvant chemotherapies for breast cancer.
Previous studies have revealed that XIAP was overexpressed at the
mRNA and immunohistochemical levels in the breast cancer specimens
of patients (12,16,17).
Although the nuclear factor-κB (NF-κB) upregulates the expression
of XIAP (18), the mechanisms
underlying its overexpression remain currently unclear.
On the other hand, ribosomes are composed of small
and large subunits and synthesize proteins from transcribed mRNAs.
However, several ribosomal proteins have been identified to have
extraribosomal functions (19,20).
For instance, the ribosomal protein S3 (RPS3) is a component of the
small 40S ribosomal subunit, but it is also a subunit in the
transcription factor NF-κB, which controls inflammatory gene
regulation (21). RPS3 activates
the NF-κB signaling by binding to the p65 subunit of NF-κB.
Moreover, RPS3 is known to function as a DNA repair endonuclease,
which is also an extraribosomal function (22).
In order to clarify the mechanisms responsible for
the overexpression of XIAP in breast cancer, we initially examined
the molecular actions of the natural product curcumin which
downregulates the expression of XIAP (23,24),
since we considered the possibility that curcumin may target the
proteins related to the expression of XIAP. We fixed curcumin onto
magnetic FG beads as previously described (25) and RPS3 was identified as one of the
curcumin-binding proteins using curcumin-fixed beads. Moreover, the
knockdown of RPS3 reduced the expression of XIAP independently of
the NF-κB pathway. In the present study, we identified a novel
function for RPS3, which regulates the expression of XIAP in breast
cancer cells.
Materials and methods
Reagents
Curcumin (Nagara Science Co., Ltd., Gifu, Japan) was
purchased and dissolved in dimethyl sulfoxide (DMSO).
Cell culture
Human breast cancer MCF-7 (estrogen and progesterone
receptor-positive, luminal type), MDA-MB-231 (triple-negative,
basal type) and human prostate cancer PC-3 cells were obtained from
the NCI-60 cancer cell line panel of the National Cancer Institute
Developmental Therapeutics Program (NCI DTP). MCF-7 cells were
maintained in Dulbecco's modified Eagle's medium (DMEM),
supplemented with 10% fetal bovine serum (FBS), 4 mM glutamine, 50
U/ml penicillin and 100 µg/ml streptomycin at 37°C in 5%
CO2. The MDA-MB-231 and PC-3 cells were cultured in
RPMI-1640 medium with 10% FBS, 2 mM glutamine, 50 U/ml penicillin
and 100 µg/ml streptomycin at 37°C in 5% CO2. It has
been identified that the activity of NF-κB in MDA-MB-231 cells is
higher than that in MCF-7 cells (26).
Cell viability assay
The cell growth was evaluated using the Cell
Counting Kit-8 (CCK-8; Dojindo Molecular Technologies, Inc.,
Kumamoto, Japan). The CCK-8 solution was added to the medium of
curcumin-treated or siRNA-transfected MCF-7 and MDA-MB-231 cells.
The absorbance (450 nm) of the medium was determined using
Multiskan FC (Thermo Fisher Scientific, Waltham, MA, USA) after 4 h
of incubation.
Detection of apoptosis
The MCF-7 and MDA-MB-231 breast cancer cells treated
with curcumin or transfected with each siRNA for RPS3 were
harvested by trypsinization. The cells were washed with
phosphate-buffered saline (PBS) and suspended in PBS containing
0.1% Triton X-100, 150 µg/ml RNase A and 50 µg/ml propidium iodide
to stain the nuclei. The DNA content in the stained nuclei was
analyzed by FACSCalibur (Becton-Dickinson, Franklin Lakes, NJ, USA)
and the hypodiploid population (sub-G1) was detected and
quantified.
Western blot analysis
The MCF-7 and MDA-MB-231 breast cancer cells were
treated with curcumin or transfected with each siRNA and lysed with
RIPA buffer [50 mM Tris-HCl (pH 8.0), 150 mM NaCl, 1% NP-40, 0.5%
deoxycholic acid, 0.1% SDS, 1 mM DTT and 0.5 mM PMSF] at 4°C for 30
min and centrifuged. The supernatants were separated by 12%
SDS-PAGE and transferred to Immobilon-P membranes (Millipore,
Billerica, MA, USA). The membranes were blocked in Tris-buffered
saline containing 5% skim milk and incubated with primary
antibodies at room temperature for 1 h. The primary antibodies were
a mouse monoclonal anti-XIAP antibody (MAB822; R&D Systems,
Minneapolis, MN, USA), a rabbit monoclonal anti-RPS3 antibody
(#9538; Cell Signaling Technology, Danvers, MA, USA) and a mouse
monoclonal anti-β-actin antibody (A5441; Sigma-Aldrich, St. Louis,
MO, USA). Subsequently, the membranes were incubated with the
secondary antibodies for 1 h at room temperature. Each protein was
visualized on BioMax XAR film (Carestream Health, Inc., Rochester,
NY, USA) using Chemi-Lumi One L (Nacalai Tesque, Kyoto, Japan) or
Immobilon Western (Millipore).
Preparation of the curcumin-fixed
beads
The magnetic FG beads with epoxy linkers were
purchased from Tamagawa Seiki Co., Ltd. (Nagano, Japan). The beads
were mixed with curcumin in dimethylformamide (DMF) containing
potassium carbonate (K2CO3) at 37°C
overnight. The resulting beads were washed three times with
deionized water and stored at 4°C.
Purification and identification of the
curcumin-binding proteins
The MCF-7 and PC-3 cells were lysed with binding
buffer [50 mM Tris-HCl (pH 8.0), 150 mM NaCl, 1% NP-40, 1 mM DTT
and 0.5 mM PMSF] for 30 min at 4°C and centrifuged. The collected
supernatants were used as whole cell extracts. The extracts were
incubated with curcumin-fixed or empty beads for 4 h at 4°C. After
the beads were washed three times with the binding buffer, the
proteins binding to these beads were eluted with Laemmli dye. The
binding proteins were subjected to SDS-PAGE and stained by silver
staining. Subsequently these proteins were subjected to in-gel
digestion with Sequencing Grade Modified Trypsin (Promega, Madison,
WI, USA) and the resulting peptide fragments were analyzed by
Autoflex II (Bruker Daltonics, Billerica, MA, USA).
Quantitative real-time RT-PCR
The total cellular RNA was extracted from the MCF-7
and MDA-MB-231 cells transfected with each siRNA using Sepasol-RNA
I Super G (Nacalai Tesque, Kyoto, Japan) and complementary DNA
(cDNA) was synthesized from total RNA with High-Capacity cDNA
Reverse Transcription kit (Applied Biosystems, Foster City, CA,
USA). Quantitative RT-PCR analysis was performed by an ABI 7300
Real-Time PCR system (Applied Biosystems) with the cDNA and TaqMan
probes (Applied Biosystems) to XIAP (Hs00745222_s1) and GAPDH
(Hs02758991_g1).
RNAi
The MCF-7 and MDA-MB-231 cells were transfected with
each siRNA using Lipofectamine RNAiMAX (Life Technologies,
Carlsbad, CA, USA). The following siRNAs (Life Technologies) were
used: siRPS3 #1 (HSS184418), 5′-GAGCUGGCUGAAGAUGGCUACUCUG-3′;
siRPS3 #2 (HSS184419), 5′-GCGGAUUCGGGAACUGACUGCUGUA-3′; siRPS3 #3
(HSS184420), 5′-UGAUCCACAGCGGAGACCCUGUUAA-3′; and negative control
siRNAs (cat. no. 12935-112 and no. 12935-115). Only RNA sequences
of sense strands are shown.
Quantification of the NF-κB
activity
The nuclear extracts were prepared from the MCF-7
and MDA-MB-231 cells transfected with each siRNA using Nuclear
Extract kit (Active Motif, Carlsbad, CA, USA). Subsequently, the
activity of the p65 subunit of NF-κB in the nuclear extracts was
determined by the NF-κB p65 Transcription Factor Assay kit (TransAM
NFκB p65; Active Motif) and Multiskan FC (Thermo Fisher
Scientific).
Statistical analysis
The data from triplicate samples are represented as
the mean ± standard deviation (SD). Statistical analysis was
performed using non-repeated measures ANOVA followed by Bonferroni
post hoc test due to comparisons of more than two groups. P<0.05
was considered to indicate a statistically significant
difference.
Results
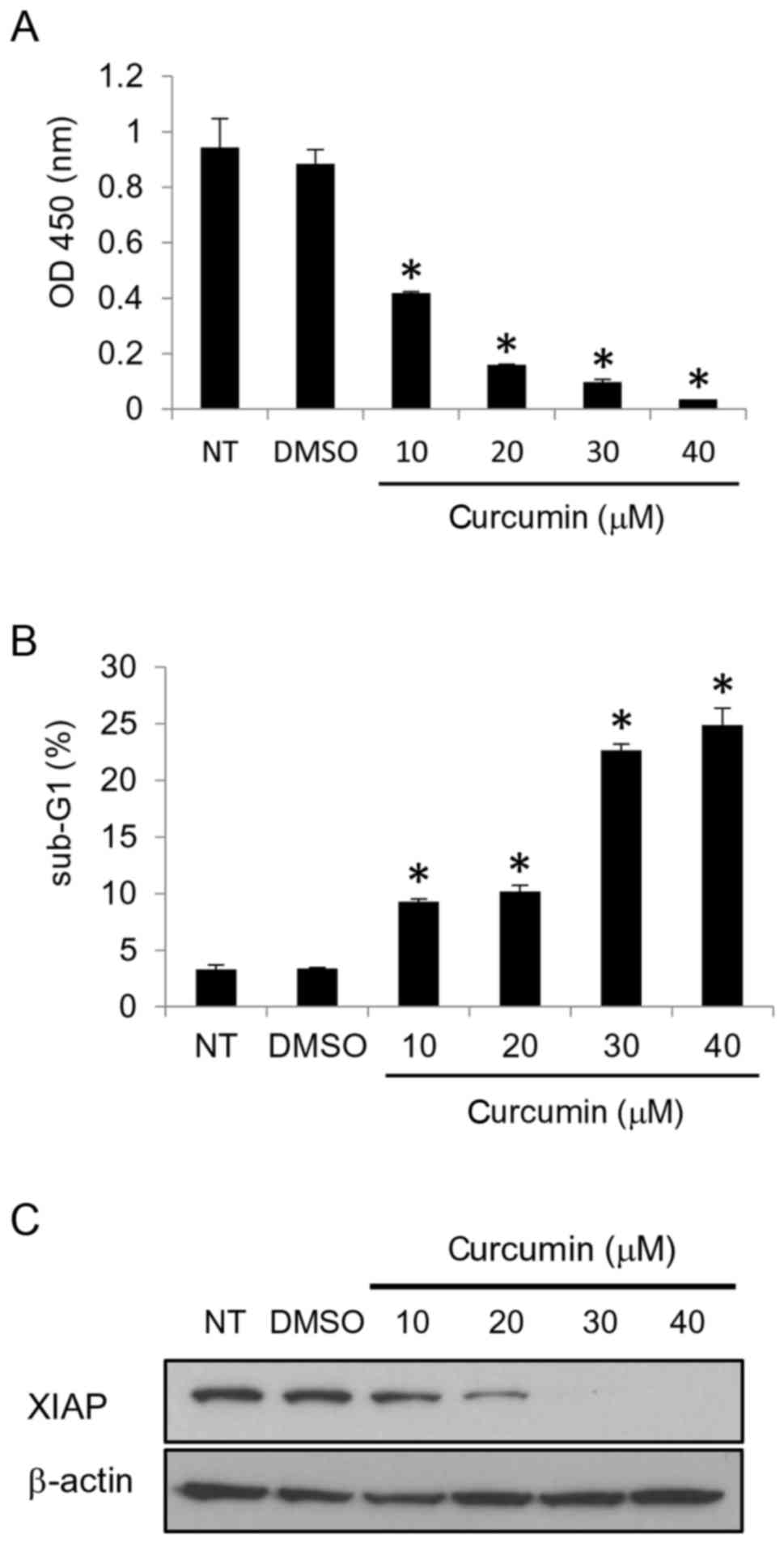
Curcumin downregulates the XIAP
protein in breast cancer cells
Previous studies have identified that XIAP is
strongly expressed in breast cancer tissues (16,17).
In order to elucidate the mechanisms responsible for the
overexpression of XIAP, we searched for compounds that downregulate
XIAP in breast cancer cells. Curcumin, one of the most common
polyphenols, inhibited the growth of the MCF-7 breast cancer cells
(Fig. 1A) and induced apoptosis
(Fig. 1B). We then found that
curcumin downregulated the XIAP protein (Fig. 1C). These results indicate that
curcumin reduces the protein levels of XIAP in the MCF-7 breast
cancer cells.
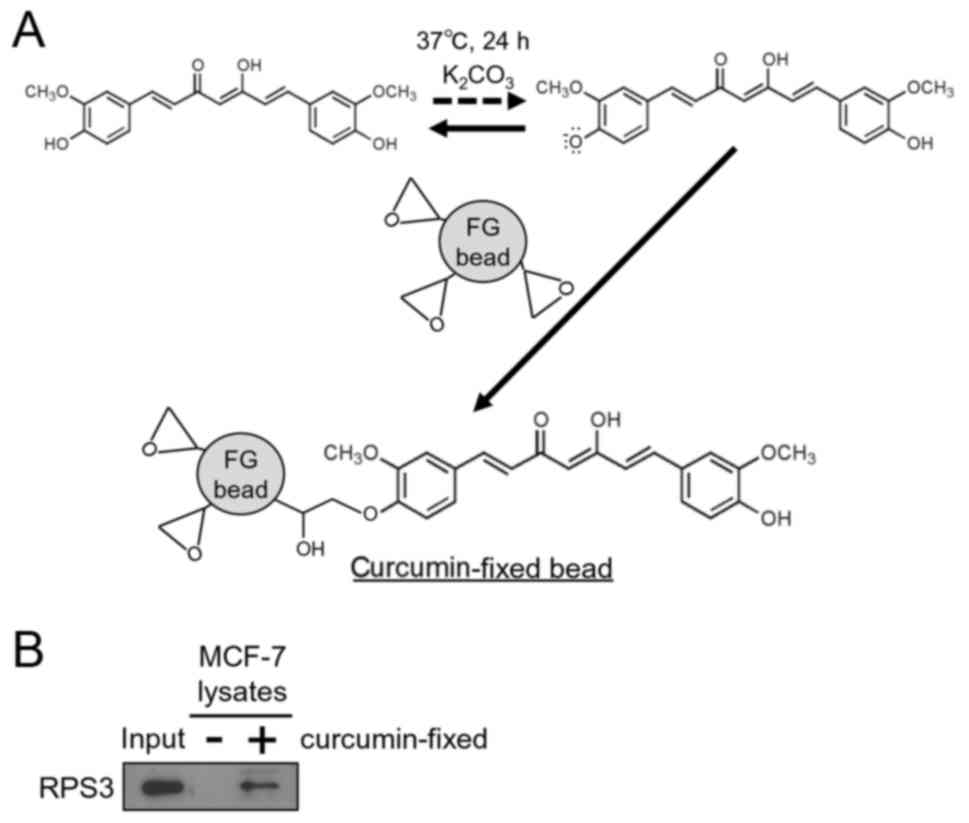
Identification of RPS3 as a
curcumin-binding protein
In an attempt to clarify the mechanisms by which
curcumin downregulates XIAP in MCF-7 cells, we examined the
curcumin-binding proteins using magnetic FG beads. Curcumin was
conjugated onto the beads with K2CO3
(Fig. 2A). We initially used PC-3
prostate cancer cells to identify the curcumin-binding proteins and
identified RPS3 as one of the curcumin-binding proteins (data not
shown). RPS3 was also purified from the whole cell extracts of the
MCF-7 cells using curcumin-fixed beads (Fig. 2B). These results reveal that RPS3 is
a target of curcumin.
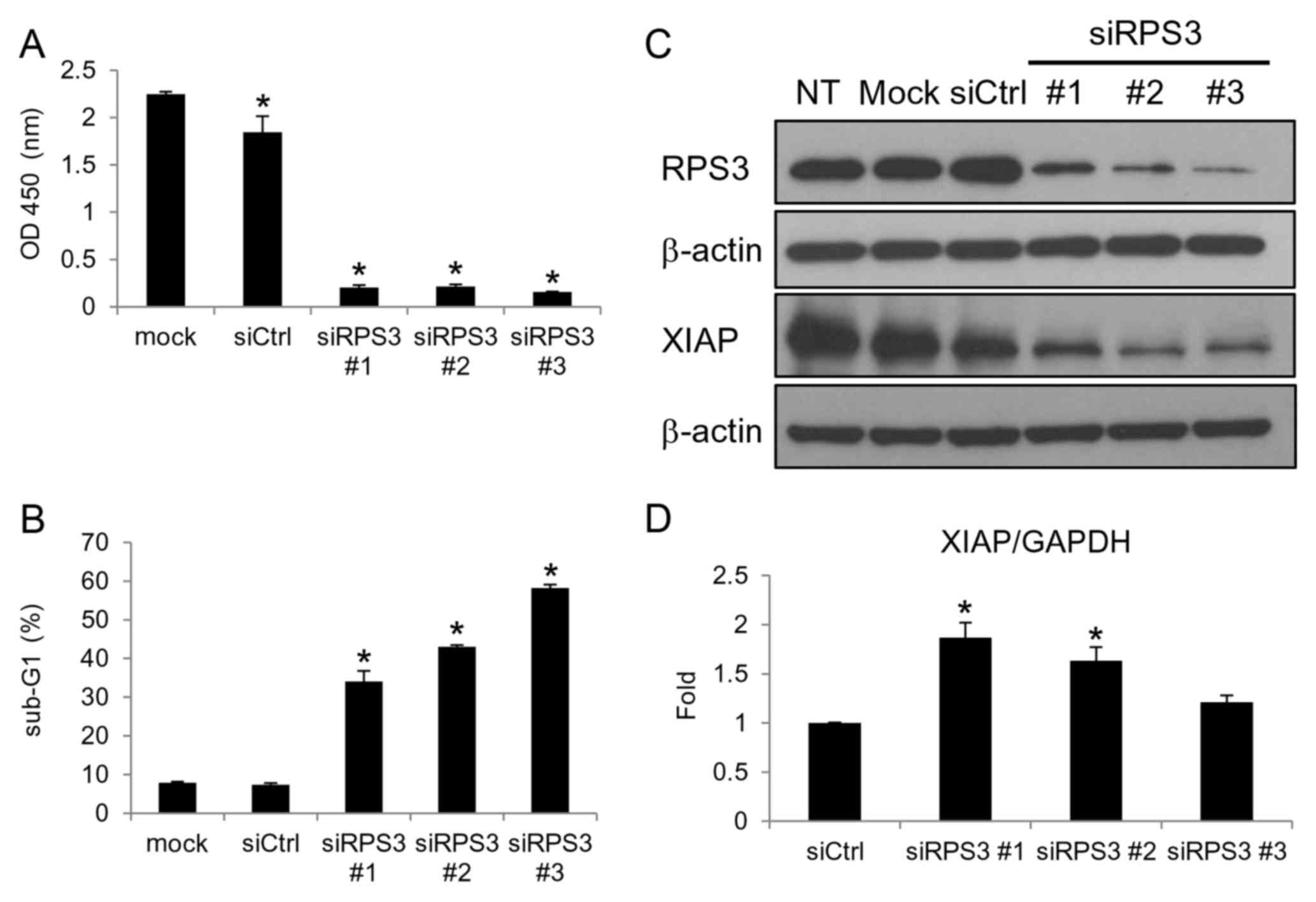
Knockdown of RPS3 downregulates the
XIAP protein independently of the NF-κB pathway
Several extraribosomal functions of the ribosomal
proteins have recently been identified (21,27–29).
RPS3 is known to be a subunit in NF-κB complexes and was found to
promote the transcription of anti-apoptotic molecules, including
XIAP in Jurkat leukemia cells (21). Therefore, we examined whether RPS3
also regulates XIAP at the mRNA level in breast cancer cells. The
knockdown of RPS3 inhibited cell growth (Fig. 3A) and induced apoptosis (Fig. 3B). Although the knockdown of RPS3
downregulated the XIAP protein (Fig.
3C), unexpectedly the mRNA level of XIAP was not reduced
(Fig. 3D). These findings reveal
that RPS3 regulates the expression of XIAP independently of the
NF-κB pathway in the MCF-7 breast cancer cells.
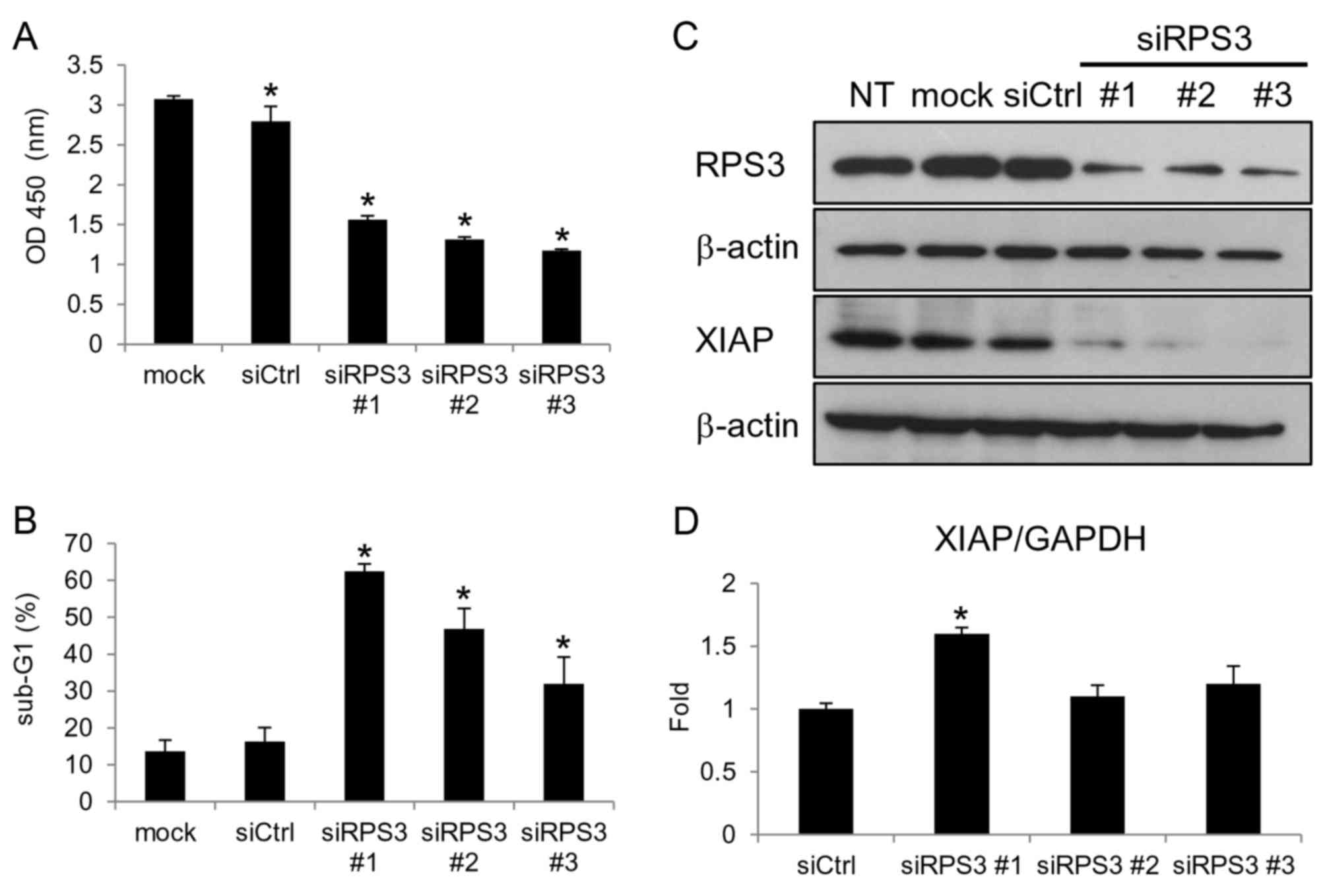 | Figure 3.Knockdown of RPS3 downregulates the
XIAP protein independently of the NF-κB pathway in MCF-7 breast
cancer cells. (A-D) The MCF-7 cells were transfected with siRPS3
#1, siRPS3 #2, siRPS3 #3, or a negative control siRNA (siCtrl).
After 120 h, cell growth (A) and apoptosis (B) were evaluated.
After 96 h, the protein (C) and mRNA (D) levels of XIAP were
analyzed. Columns, means of triplicate data; bars, SD. *P<0.05.
XIAP, X-linked inhibitor of apoptosis; RPS3, ribosomal protein S3;
sub-G1, hypodiploid population. |
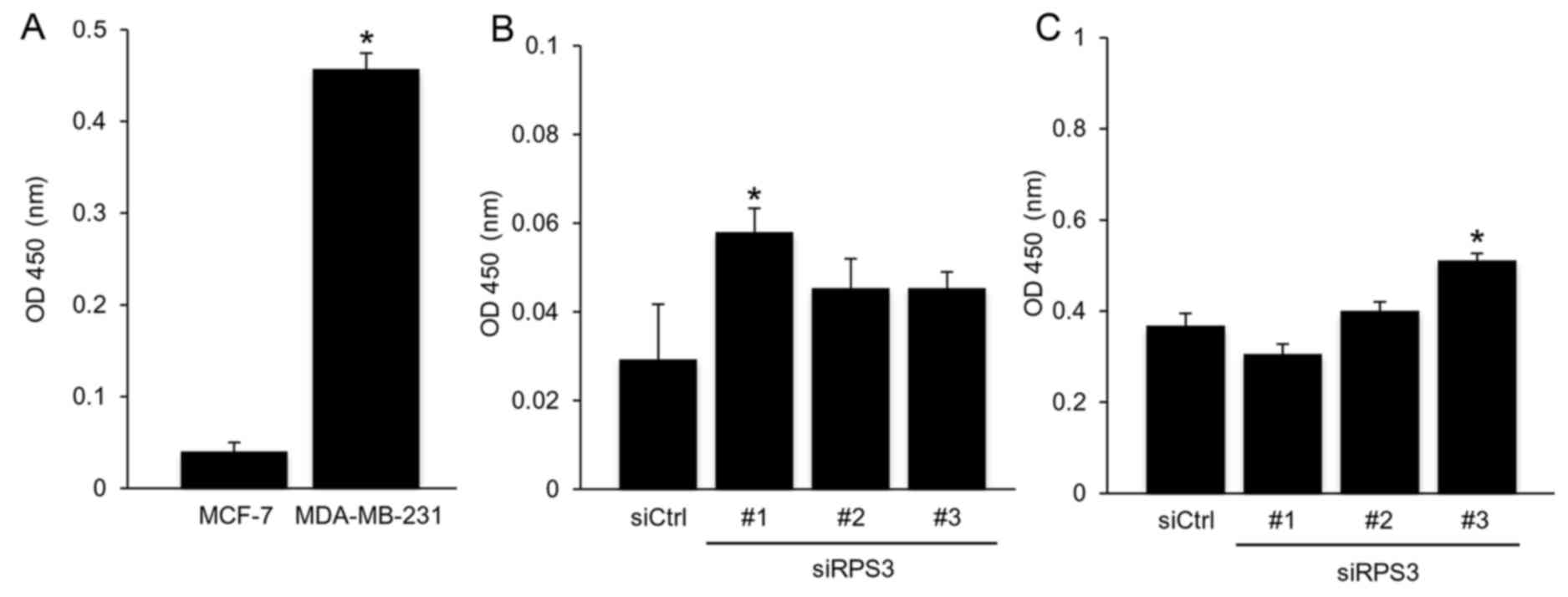
Additionally, in order to confirm that RPS3
regulates the expression of XIAP independently of the NF-κB
signaling, we used another breast cancer cell line, MDA-MB-231.
This cell line is classified as triple-negative breast cancer,
which is the most aggressive subtype (30,31)
and the NF-κB pathway is activated in this cell line (26). Similar to the MCF-7 breast cancer
cells, the knockdown of RPS3 inhibited the cell growth (Fig. 4A) and induced apoptosis (Fig. 4B). The RPS3 depletion downregulated
the XIAP protein (Fig. 4C), but not
the mRNA level of XIAP in the MDA-MB-231 cells (Fig. 4D). Collectively, these results
clearly reveal that RPS3 regulates the expression of XIAP
independently of the NF-κB pathway in breast cancer cells, which is
inconsistent with a previous study (21).
In addition, we investigated the activation of NF-κB
by quantifying the activity of the p65 subunit of the NF-κB. As
shown in Fig. 5A, the activity of
NF-κB in the MDA-MB-231 breast cancer cells was higher than that in
the MCF-7 breast cancer cells as previously reported (26). The knockdown of RPS3 did not reduce
the activity of NF-κB in the MCF-7 and MDA-MB-231 cells (Fig. 5B and C), whereas the RPS3 knockdown
downregulated the XIAP protein (Figs.
3C and 4C). These findings
further reveal that RPS3 regulates the expression of XIAP
independently of NF-κB.
Discussion
In the present study in order to elucidate the
mechanisms underlying the overexpression of XIAP in breast cancer
(16,17), we focused on the natural compound
curcumin, which downregulated the expression of XIAP in breast
cancer cells as shown in Fig. 1C.
We identified, for the first time, RPS3 as a curcumin-binding
protein and found that the knockdown of RPS3 downregulated the
expression of XIAP with the induction of apoptosis in breast cancer
cells, suggesting that RPS3 can increase the expression of XIAP.
According to previous findings obtained using Jurkat leukemia
cells, RPS3 directly binds to the p65 subunit of NF-κB complexes
and plays a role in gene transcription as part of these complexes
(21). In contrast to these
findings, our study revealed that RPS3 did not regulate the
expression of XIAP at the transcriptional level, suggesting that
RPS3 upregulates XIAP independently of the NF-κB pathway. Since
this discrepancy may be attributed to the low activity of NF-κB in
the MCF-7 breast cancer cells, we analyzed the expression of XIAP
regulated by RPS3 using the MDA-MB-231 breast cancer cells in which
the NF-κB pathway is more activated as reported previously
(26). However, the results
obtained using the MCF-7 and MDA-MB-231 breast cancer cells were
similar. Collectively, our results indicate that RPS3 regulates the
expression of XIAP independently of the NF-κB pathway in breast
cancer cells.
As shown in Figs. 3
and 4, the knockdown of RPS3
downregulated XIAP at the protein level, but not at the mRNA level.
The expression of XIAP is known to be regulated at the protein
level in various manners. For example, many microRNAs suppress the
translation of XIAP (32–36) and the E3 ligase TRIM32 degrades the
XIAP protein (37). In addition,
the TRIP-Br1 and TRIP-Br3 proteins inhibit the degradation of XIAP
(38) and a novel mechanism for
XIAP degradation through an autophagy pathway has been identified
(39). Therefore, multifaceted
analyses that consider these molecules are needed in order to
clarify the regulation of the expression of XIAP by RPS3.
It is considered important to search for biomarkers
that may be used to predict sensitivity to chemotherapy. Previous
studies reported that the anti-apoptotic protein XIAP is
overexpressed in breast cancer (16,17)
and furthermore, that the upregulation of XIAP is associated with
resistance to anthracycline or taxane, which are often used in
adjuvant therapies for breast cancer (12). Since our results revealed that RPS3
upregulates the expression of XIAP, these findings indicate that it
is possible to predict sensitivity to adjuvant chemotherapies based
on the RPS3 levels.
At present, we have analyzed the molecular
mechanisms by which various food factors and natural products exert
antitumor activities (40–44). In the course of our studies, we
identified several binding proteins of natural compounds using
magnetic FG beads and discovered novel functions through which
these binding proteins regulate tumor properties by referring to
the antitumor activities of these compounds (25,45–47).
In the present study, by using curcumin as a chemical probe, we
identified RPS3 as a curcumin-binding protein and found that RPS3
regulates the expression of XIAP independently of the NF-κB pathway
in breast cancer cells. As demonstrated it is useful for clarifying
novel molecular mechanisms in cells to apply natural compounds
exhibiting bioactivities as chemical probes.
Acknowledgements
This study was supported by a JSPS Grant-in-Aid for
Young Scientists (A) (no. 23689036 to Y.I.). We thank the Medical
English Service (http://www.med-english.com) for the English language
editing.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2015. CA Cancer J Clin. 65:5–29. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Baselga J, Campone M, Piccart M, Burris HA
III, Rugo HS, Sahmoud T, Noguchi S, Gnant M, Pritchard KI, Lebrun
F, et al: Everolimus in postmenopausal hormone-receptor-positive
advanced breast cancer. N Engl J Med. 366:520–529. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Verma S, Miles D, Gianni L, Krop IE,
Welslau M, Baselga J, Pegram M, Oh DY, Diéras V, Guardino E, et al
EMILIA Study Group, : Trastuzumab emtansine for HER2-positive
advanced breast cancer. N Engl J Med. 367:1783–1791. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Swain SM, Baselga J, Kim SB, Ro J,
Semiglazov V, Campone M, Ciruelos E, Ferrero JM, Schneeweiss A,
Heeson S, et al CLEOPATRA Study Group, : Pertuzumab, trastuzumab,
and docetaxel in HER2-positive metastatic breast cancer. N Engl J
Med. 372:724–734. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Rastogi P, Anderson SJ, Bear HD, Geyer CE,
Kahlenberg MS, Robidoux A, Margolese RG, Hoehn JL, Vogel VG, Dakhil
SR, et al: Preoperative chemotherapy: Updates of National Surgical
Adjuvant Breast and Bowel Project Protocols B-18 and B-27. J Clin
Oncol. 26:778–785. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Smith IC, Heys SD, Hutcheon AW, Miller ID,
Payne S, Gilbert FJ, Ah-See AK, Eremin O, Walker LG, Sarkar TK, et
al: Neoadjuvant chemotherapy in breast cancer: Significantly
enhanced response with docetaxel. J Clin Oncol. 20:1456–1466. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ekert PG, Silke J and Vaux DL: Caspase
inhibitors. Cell Death Differ. 6:1081–1086. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chai J, Shiozaki E, Srinivasula SM, Wu Q,
Datta P, Alnemri ES and Shi Y: Structural basis of caspase-7
inhibition by XIAP. Cell. 104:769–780. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Huang Y, Park YC, Rich RL, Segal D, Myszka
DG and Wu H: Structural basis of caspase inhibition by XIAP:
Differential roles of the linker versus the BIR domain. Cell.
104:781–790. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Riedl SJ, Renatus M, Schwarzenbacher R,
Zhou Q, Sun C, Fesik SW, Liddington RC and Salvesen GS: Structural
basis for the inhibition of caspase-3 by XIAP. Cell. 104:791–800.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Shiozaki EN, Chai J, Rigotti DJ, Riedl SJ,
Li P, Srinivasula SM, Alnemri ES, Fairman R and Shi Y: Mechanism of
XIAP-mediated inhibition of caspase-9. Mol Cell. 11:519–527. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
de Moraes G Nestal, Delbue D, Silva KL,
Robaina MC, Khongkow P, Gomes AR, Zona S, Crocamo S, Mencalha AL,
Magalhães LM, et al: FOXM1 targets XIAP and Survivin to modulate
breast cancer survival and chemoresistance. Cell Signal.
27:2496–2505. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Lima RT, Martins LM, Guimarães JE, Sambade
C and Vasconcelos MH: Specific downregulation of bcl-2 and xIAP by
RNAi enhances the effects of chemotherapeutic agents in MCF-7 human
breast cancer cells. Cancer Gene Ther. 11:309–316. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Bilim V, Kasahara T, Hara N, Takahashi K
and Tomita Y: Role of XIAP in the malignant phenotype of
transitional cell cancer (TCC) and therapeutic activity of XIAP
antisense oligonucleotides against multidrug-resistant TCC in
vitro. Int J Cancer. 103:29–37. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Sasaki H, Sheng Y, Kotsuji F and Tsang BK:
Down-regulation of X-linked inhibitor of apoptosis protein induces
apoptosis in chemoresistant human ovarian cancer cells. Cancer Res.
60:5659–5666. 2000.PubMed/NCBI
|
|
16
|
Jaffer S, Orta L, Sunkara S, Sabo E and
Burstein DE: Immunohistochemical detection of antiapoptotic protein
X-linked inhibitor of apoptosis in mammary carcinoma. Hum Pathol.
38:864–870. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Xu YC, Liu Q, Dai JQ, Yin ZQ, Tang L, Ma
Y, Lin XL and Wang HX: Tissue microarray analysis of X-linked
inhibitor of apoptosis (XIAP) expression in breast cancer patients.
Med Oncol. 31:7642014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Stehlik C, de Martin R, Kumabashiri I,
Schmid JA, Binder BR and Lipp J: Nuclear factor
(NF)-kappaB-regulated X-chromosome-linked iap gene expression
protects endothelial cells from tumor necrosis factor alpha-induced
apoptosis. J Exp Med. 188:211–216. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Warner JR and McIntosh KB: How common are
extraribosomal functions of ribosomal proteins? Mol Cell. 34:3–11.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang Y and Lu H: Signaling to p53:
Ribosomal proteins find their way. Cancer Cell. 16:369–377. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wan F, Anderson DE, Barnitz RA, Snow A,
Bidere N, Zheng L, Hegde V, Lam LT, Staudt LM, Levens D, et al:
Ribosomal protein S3: A KH domain subunit in NF-kappaB complexes
that mediates selective gene regulation. Cell. 131:927–939. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kim J, Chubatsu LS, Admon A, Stahl J,
Fellous R and Linn S: Implication of mammalian ribosomal protein S3
in the processing of DNA damage. J Biol Chem. 270:13620–13629.
1995. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Woo JH, Kim YH, Choi YJ, Kim DG, Lee KS,
Bae JH, Min DS, Chang JS, Jeong YJ, Lee YH, et al: Molecular
mechanisms of curcumin-induced cytotoxicity: Induction of apoptosis
through generation of reactive oxygen species, down-regulation of
Bcl-XL and IAP, the release of cytochrome c and inhibition of Akt.
Carcinogenesis. 24:1199–1208. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Park S, Cho DH, Andera L, Suh N and Kim I:
Curcumin enhances TRAIL-induced apoptosis of breast cancer cells by
regulating apoptosis-related proteins. Mol Cell Biochem. 383:39–48.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Iizumi Y, Oishi M, Taniguchi T, Goi W,
Sowa Y and Sakai T: The flavonoid apigenin downregulates CDK1 by
directly targeting ribosomal protein S9. PLoS One. 8:e732192013.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yamaguchi N, Ito T, Azuma S, Ito E, Honma
R, Yanagisawa Y, Nishikawa A, Kawamura M, Imai J, Watanabe S, et
al: Constitutive activation of nuclear factor-kappaB is
preferentially involved in the proliferation of basal-like subtype
breast cancer cell lines. Cancer Sci. 100:1668–1674. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lohrum MAE, Ludwig RL, Kubbutat MH, Hanlon
M and Vousden KH: Regulation of HDM2 activity by the ribosomal
protein L11. Cancer Cell. 3:577–587. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Dai MS and Lu H: Inhibition of
MDM2-mediated p53 ubiquitination and degradation by ribosomal
protein L5. J Biol Chem. 279:44475–44482. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Dai MS, Zeng SX, Jin Y, Sun XX, David L
and Lu H: Ribosomal protein L23 activates p53 by inhibiting MDM2
function in response to ribosomal perturbation but not to
translation inhibition. Mol Cell Biol. 24:7654–7668. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lin NU, Vanderplas A, Hughes ME, Theriault
RL, Edge SB, Wong YN, Blayney DW, Niland JC, Winer EP and Weeks JC:
Clinicopathologic features, patterns of recurrence, and survival
among women with triple-negative breast cancer in the National
Comprehensive Cancer Network. Cancer. 118:5463–5472. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Dent R, Trudeau M, Pritchard KI, Hanna WM,
Kahn HK, Sawka CA, Lickley LA, Rawlinson E, Sun P and Narod SA:
Triple-negative breast cancer: Clinical features and patterns of
recurrence. Clin Cancer Res. 13:4429–4434. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Liu S, Zhang P, Chen Z, Liu M, Li X and
Tang H: MicroRNA-7 downregulates XIAP expression to suppress cell
growth and promote apoptosis in cervical cancer cells. FEBS Lett.
587:2247–2253. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Xie Y, Tobin LA, Camps J, Wangsa D, Yang
J, Rao M, Witasp E, Awad KS, Yoo N, Ried T, et al: MicroRNA-24
regulates XIAP to reduce the apoptosis threshold in cancer cells.
Oncogene. 32:2442–2451. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Li X, Chen W, Zeng W, Wan C, Duan S and
Jiang S: microRNA-137 promotes apoptosis in ovarian cancer cells
via the regulation of XIAP. Br J Cancer. 116:66–76. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Ren Y, Han X, Yu K, Sun S, Zhen L, Li Z
and Wang S: microRNA-200c downregulates XIAP expression to suppress
proliferation and promote apoptosis of triple-negative breast
cancer cells. Mol Med Rep. 10:315–321. 2014.PubMed/NCBI
|
|
36
|
Wang C, Ju H, Shen C and Tong Z: miR-429
mediates δ-tocotrienol-induced apoptosis in triple-negative breast
cancer cells by targeting XIAP. Int J Clin Exp Med. 8:15648–15656.
2015.PubMed/NCBI
|
|
37
|
Ryu YS, Lee Y, Lee KW, Hwang CY, Maeng JS,
Kim JH, Seo YS, You KH, Song B and Kwon KS: TRIM32 protein
sensitizes cells to tumor necrosis factor (TNFα)-induced apoptosis
via its RING domain-dependent E3 ligase activity against X-linked
inhibitor of apoptosis (XIAP). J Biol Chem. 286:25729–25738. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Li C, Jung S, Lee S, Jeong D, Yang Y, Kim
KI, Lim JS, Cheon CI, Kim C, Kang YS, et al: Nutrient/serum
starvation derived TRIP-Br3 down-regulation accelerates apoptosis
by destabilizing XIAP. Oncotarget. 6:7522–7535. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Wang N, Feng Y, Zhu M, Siu FM, Ng KM and
Che CM: A novel mechanism of XIAP degradation induced by
timosaponin AIII in hepatocellular carcinoma. Biochim Biophys Acta.
1833:2890–2899. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Hosokawa N, Hosokawa Y, Sakai T, Yoshida
M, Marui N, Nishino H, Kawai K and Aoike A: Inhibitory effect of
quercetin on the synthesis of a possibly cell-cycle-related 17-kDa
protein, in human colon cancer cells. Int J Cancer. 45:1119–1124.
1990. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Horinaka M, Yoshida T, Shiraishi T, Nakata
S, Wakada M, Nakanishi R, Nishino H, Matsui H and Sakai T: Luteolin
induces apoptosis via death receptor 5 upregulation in human
malignant tumor cells. Oncogene. 24:7180–7189. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Matsui TA, Sowa Y, Yoshida T, Murata H,
Horinaka M, Wakada M, Nakanishi R, Sakabe T, Kubo T and Sakai T:
Sulforaphane enhances TRAIL-induced apoptosis through the induction
of DR5 expression in human osteosarcoma cells. Carcinogenesis.
27:1768–1777. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Taniguchi H, Yoshida T, Horinaka M, Yasuda
T, Goda AE, Konishi M, Wakada M, Kataoka K, Yoshikawa T and Sakai
T: Baicalein overcomes tumor necrosis factor-related
apoptosis-inducing ligand resistance via two different
cell-specific pathways in cancer cells but not in normal cells.
Cancer Res. 68:8918–8927. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Ichikawa M, Sowa Y, Iizumi Y, Aono Y and
Sakai T: Resibufogenin induces G1-phase arrest through the
proteasomal degradation of cyclin D1 in human malignant tumor
cells. PLoS One. 10:e01298512015. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Kume K, Iizumi Y, Shimada M, Ito Y, Kishi
T, Yamaguchi Y and Handa H: Role of N-end rule ubiquitin ligases
UBR1 and UBR2 in regulating the leucine-mTOR signaling pathway.
Genes Cells. 15:339–349. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Taniguchi T, Iizumi Y, Watanabe M, Masuda
M, Morita M, Aono Y, Toriyama S, Oishi M, Goi W and Sakai T:
Resveratrol directly targets DDX5 resulting in suppression of the
mTORC1 pathway in prostate cancer. Cell Death Dis. 7:e22112016.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Watanabe M, Iizumi Y, Sukeno M,
Iizuka-Ohashi M, Sowa Y and Sakai T: The pleiotropic regulation of
cyclin D1 by newly identified sesaminol-binding protein ANT2.
Oncogenesis. 6:e3112017. View Article : Google Scholar : PubMed/NCBI
|