Introduction
Colorectal cancer (CRC) is the third most common
cancer and the fourth leading cause of cancer mortality (1). Therefore, finding strategies to combat
CRC is still an emergent health problem. Phytochemicals can inhibit
CRC by disrupting multiple mechanisms that are central to cancer
progression (2,3). Flavonoids are one of the most numerous
and widely distributed family of phytochemicals in different types
of vegetables and fruits. Compared with conventional anticancer
drugs, plant-derived flavonoids have an extra margin of safety
since they show only marginal toxicity even at relatively high
concentrations (2,3).
Leaves of Toona sinensis (T. sinensis)
M.Roem., a popular vegetable in China, were reported to have
various biologically activated effects including antioxidative
(4–6), anticancer (7–9),
anti-inflammatory (10–12) and anti-hyperglycemic (13,14).
Crude water extracts of T. sinensis leaves have been shown
to inhibit cell proliferation and induce apoptosis of a number of
human cancers, including lung adenocarcinoma (A549) cells (15), human SKOV3 ovarian cancer cell lines
(16), HL-60 leukemia cells
(8,17,18),
H661 lung (19) and cervical
carcinoma HeLa cells (20), HepG
liver cancer (21) and MCF-7 breast
cancer cell (21), and prostate
cancer cells (22).
Antiproliferative effects on Caco-2 human colon cancer cells have
also been reported (21). Although
T. sinensis leaves have been used medicinally for a long
time, its effects are still not fully understood.
A previous study demonstrated that T.
sinensis leaf extracts are rich in active ingredients such as
flavonoids, volatile oils and alkaloids (17). The major identified flavonoids in
QTL are quercetrin, kaempferol-3-O-α-L-rhamopyranoside,
astragalin, quercetin, kaempferol, methyl and ethyl gallate, and
1,2,3,4,6-penta-O-galloyl-β-D-glucopyranose (4,17). The
present study was undertaken to evaluate the growth-inhibitory and
apoptosis-inducing effects of QTL and the probable molecular
mechanisms in the SW620 human CRC cell line.
Materials and methods
Materials
Dulbecco's modified Eagle's medium (DMEM), dimethyl
sulfoxide (DMSO),
3-(4,5-dimethylthiazol-2-yl)2,5-diphenyltetrazolium bromide (MTT)
2′,7′-dichlorofluorescin diacetate (DCFH-DA), N-acetyl
cysteine (NAC) and glutathione (GSH) were obtained from Sigma (St.
Louis, MO, USA), and fetal bovine serum (FBS) was purchased from
Gibco (Paisley, UK).
5,5,6,6-Tetrachloro-1,1,3,3-tetraethylbenzimidazolyl carbocyanine
iodide (JC-1) were purchased from Beyotime Institute of
Biotechnology (Shanghai, China). The commercially available assay
kits used detect the content of malondialdehyde (MDA), and the
activity of SOD, GPx and CAT were purchased from the Institute of
Jiancheng Biological Engineering (Nanjing, China). The primers for
superoxide dismutase (SOD), glutathione peroxidase (GPx) and
catalase (CAT) were designed and synthesized by Takara
Biotechnology Co., Ltd. (Dalian, China). Reagents, such as enzymes,
co-factors and nucleotides for internal standard construction and
reverse transcriptase-polymerase chain reaction (RT-PCR) were
obtained from Takara Biotechnology Co., Ltd. Rabbit anti-active
p53, p21, Bax, cytochrome c, caspase-9, Apaf-1 and caspase-3
polyclonal antibodies were purchased from Cell Signaling
Technologies (Beverly, MA, USA) and β-actin antibodies were
purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA).
Other chemicals were analytical grade.
Preparation of QTL
Leaves of T. sinensis were collected in
Shaanxi Province (China), in August 2015 and authenticated by
experts in the College of Forestry, Northwest A&F University
(Shaanxi, China). T. sinensis leaves (50 g) were cut into
pieces of ~2 cm in width and were dried. The leaves were then
soaked in a 70% ethanol solvent (1:10, w/v) for 2.5 h and were
sonicated in an ultrasonic bath at 200 kHz at 55°C for 45 min. The
samples were then filtered through a 0.45 µm microporous membrane
(Shanghai Wanzi Shiye Co., Ltd., Shanghai, China). The filtrate was
collected, and the solid was extracted two additional times using
the same volume of fresh solvent. The combined solutions were
concentrated and dried using a rotary evaporator. The dried crude
extract was added to distilled water and defatted with petroleum
ether and ethyl acetate. The ethyl acetate fractions of T.
sinensis leaves, eluted with EtOAc MeOH (8:1), were further
separated and purified by capillary electrophoresis using silica
gel column chromatography to yield quercetrin. QTL was concentrated
and dried using a rotary evaporator.
QTL was dissolved in DMSO immediately before use,
and the final concentration of DMSO did not exceed 0.1% (v/v) in
any of the experiments. Concentrations of QTL ranged from 12.5–400
µg/ml. DMSO at 0.1% was used as a control. All determinations were
performed in triplicate.
Cell culture
SW620 cells were maintained in DMEM supplemented
with 10% heat-inactivated FBS, 100 U/ml penicillin and 100 µg/ml
streptomycin (Thermo Fisher Scientific, Franklin Lakes, NJ, USA) in
a humidified 5% CO2 incubator at 37°C. The medium was
changed every 48 h. SW620 cells were cultured in 24- or 96-well
plates.
Cell viability
The cell survival rate was quantified using a
colorimetric MTT assay. Briefly, aliquots (20 µl) of the 2.5 mg/ml
MTT stock solution were pipetted into each well, and the plate was
incubated at 37°C in a humidified 5% CO2 incubator.
After 4 h, the medium was removed, and DMSO (200 µl) was added to
each well to dissolve the formazan. The optical density of each
well was assessed 10 min later at 570 nm by a spectrophotometer
(Tecan Infinite M200 PRO; Tecan, Männedorf, Switzerland).
Flow cytometric analysis of the cell
cycle
Cell suspensions (0.5–1×105/ml) were
prepared by trypsinization and washed twice with phosphate-buffered
saline (PBS). Τhe cells were then fixed with 70% ethanol at 4°C and
resuspended in PBS containing 0.25 mg/ml of RNase A (Thermo Fisher
Scientific). The suspension was incubated for 30 min at 37°C, and
then the cells were labeled with propidium iodide (PI) (50 µg/ml).
The total DNA content was quantified by fluorescence using a
Becton-Dickinson (BD Biosciences, San Jose, CA, USA) FACS flow
cytometer.
Flow cytometric analysis for
apoptosis
Prepared SW620 cells (1×106/ml) were
washed twice with cold PBS, and then re-suspended gently in 500 µl
binding buffer. Thereafter, the cells were stained in 5 µl Annexin
V-FITC and well shaken. Finally, 5 µl PI was added to these cells
and incubation for 20 min in a dark place followed. Subsequently,
the cells were analyzed by FACS flow cytometer, Becton-Dickinson
(BD Biosciences).
Western blotting
Cell lysates (30 µg of total protein) were analyzed
on 8–12% SDS-PAGE under a gradient concentration. The proteins were
transferred to an immunoblot polyvinylidene fluoride (PVDF)
membrane (Millipore, Billerica, MA, USA), which was blocked with 5%
skim milk in TBS containing 0.1% Tween-20 (TBS-T) for 1 h.
Subsequently, the primary monoclonal antibodies were added to the
TBS-T at a 1:1,000 dilution and were incubated for 1 h. Primary
antibody bindings were detected using a secondary antibody
conjugated to horseradish peroxidase and enhanced using an enhanced
chemiluminescence (ECL) assay kit (Amersham Pharmacia Biosciences,
Buckinghamshire, UK) according to the manufacturer's instructions.
Chemiluminescence was imaged on a FujiFilm LAS-3000 system
(FujiFilm, Tokyo, Japan). The basal levels of proteins were
normalized to the level of β-actin protein.
Assessment of the mitochondrial
membrane potential (ΔΨm)
The mitochondrial membrane potential (∆Ψm) was
analyzed by fluorescence spectrophotometry using a JC-1 dye. In
brief, SW620 cells were stained with 5 mM JC-1 at 37°C for 20 min
in 5% CO2. The cells were pelleted by centrifugation
(5,000 rpm for 5 min at 4°C), and then resuspended in PBS. The JC-1
fluorescence of the cell suspensions and PBS controls were assessed
in triplicate in Costar 96-well plates using a microplate reader
[Ex/Em (green)/(red): 485/538/590 nm] (Tecan Infinite M200 PRO).
FL2/FL1 ratios were then calculated. Each well was scanned by
measuring the intensity of each 25-square grid (of a 1
mm2 area), which was arranged in a 5×5 rectangular array
(bottom scanning). A higher red-to-green ratio indicated a more
polarized or more negative and hyperpolarized mitochondrial inner
membrane.
Assessment of intracellular ROS
level
Cellular ROS level was assessed using the
dichlorofluorescein assay. Cells were collected and incubated with
100 µM DCF-DA (dissolved in DMSO) for 30 min at 37°C. Then, the
cells were washed three times with PBS (pH 7.4) and the relative
levels of fluorescence were quantified on a spectrophotofluorimeter
(Tecan Infinite M200 PRO). The measured fluorescence values were
expressed as a percentage of the fluorescence in control cells.
Assays on the content of MDA and the
activity of GPx, SOD and CAT
The content of MDA and the activity of SOD, GPx and
CAT were determined by commercially available assay kits following
the manufacturer's protocols. The level of lipid peroxidation was
indicated by the amount of MDA in the cells as assayed using the
thiobarbituric acid reactions (TBARS). The absorbance was read at
548 nm. The CAT activity was determined by measuring the decrease
in absorbance at 405 nm due to the degradation of
H2O2. The SOD activity was
spectrophotometrically determined at 550 nm, indicating the
inhibition of the oxidation of oxymine by the xanthine/xanthine
oxidase system. The GPx activity was assayed by quantifying the
oxidation rate of glutathione to oxidized glutathione by
H2O2 at 412 nm. The protein concentration was
assessed using the BCA protein assay kit (Pierce, Rockford, IL,
USA) with BSA as a standard.
Quantitative RT-PCR analysis
Cells were washed in ice-cold PBS, and RNA was
extracted using TRIzol reagent (BioTeke Corporation, Beijing,
China) as described by the manufacturer. cDNA was synthesized from
mRNA using a PrimeScript™ RT reagent kit, followed by RT-PCR using
a SYBR Premix Ex Taq™ reagent kit (both from Takara, Dalian, China)
and the ABI PRISM 7500 sequence detection system (Applied
Biosystems, Foster City, CA, USA). The reactions were performed
with the following cycling conditions: 95°C for 30 sec, followed by
40 cycles of 95°C for 5 sec and 56°C for 30 sec. β-actin was used
as a housekeeping gene for normalization. All experiments were
repeated at least three times. The relative change in gene
expression was analyzed using the 2−ΔΔCt method. The
sequences for the RT-PCR primers were as follows: primers for SOD
were: 5′-CTTTGTTGCCGTGCGATG-3′ (forward) and
5′-CTAACCCCGTGAATGGACAGA-3′ (reverse); GPx,
5′-CCTCTAAACCTACGAGGGAGGAA-3′ (forward) and
5′-GGGAAACTCGCCTTGGTCT-3′ (reverse); primers for CAT,
5′-TCCAAGGCAAAGGTATTTGAGCA-3′ (forward) and
5′-CAACGAGATCCCAGTTACCATCTT C-3′ (reverse); primers for β-actin,
5′-CATCCGTAAAGACCTCTATGCCAA C-3′ (forward) and
5′-ATGGAGCCACCGATCCACA-3′ (reverse).
Statistical analysis
All data are expressed as the means ± SD of at least
three independent determinations for each experiment. Statistical
analyses were performed using SPSS version 13.0 (SPSS, Inc.,
Chicago, IL, USA). A one-way ANOVA with Duncan's multiple range
test was used to examine differences between groups. P<0.05 was
considered significant.
Results
Cell growth inhibition and
morphological changes
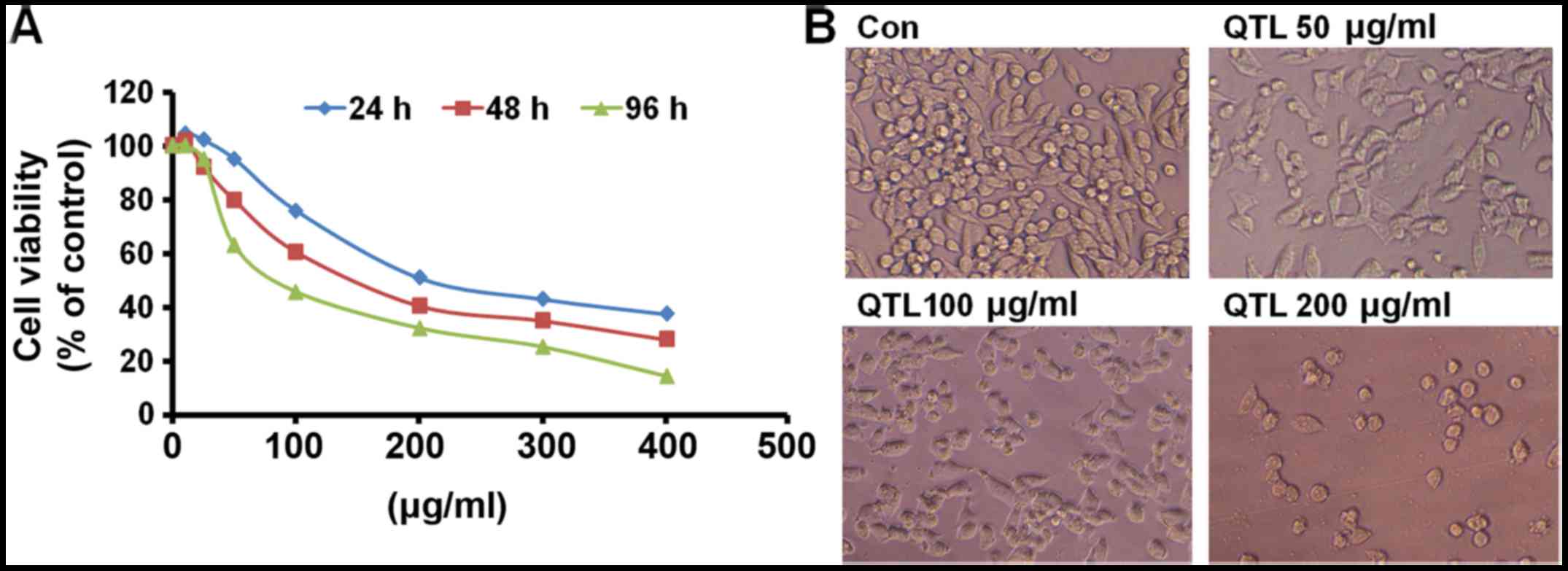
The growth inhibitory potential of QTL (12.5–400
µg/ml) in SW620 human CRC cells at different time-points after
addition (24, 48 and 96 h) is shown in Fig. 1A. Treatment with QTL exhibited
significant growth inhibition in a dose- and time-dependent manner.
The results clearly revealed that when SW620 cells were treated
with QTL for a prolonged period (96 h), the IC50 values
were the lowest (75.11 µg/ml). By contrast, when the cells were
treated with QTL for a short time (24 and 48 h), the
IC50 values were higher (198.21 and 112.55 µg/ml,
respectively).
When the cells were treated with different
concentrations of QTL for 48 h, morphological changes of SW620
cells were marked (Fig. 1B). The
cells treated with 50 µg/ml of QTL exhibited swelling, shrinkage
accompanied by rounding of cells, and membrane blebbing. Cells
incubated with 100 µg/ml of QTL shrunk and exhibited disrupted
intercellular contacts. In the group treated with 200 µg/ml of QTL,
the cells were binucleated and the majority of the cells were
detached from the wells, indicating cell death.
Analysis of cell cycle and apoptosis
status
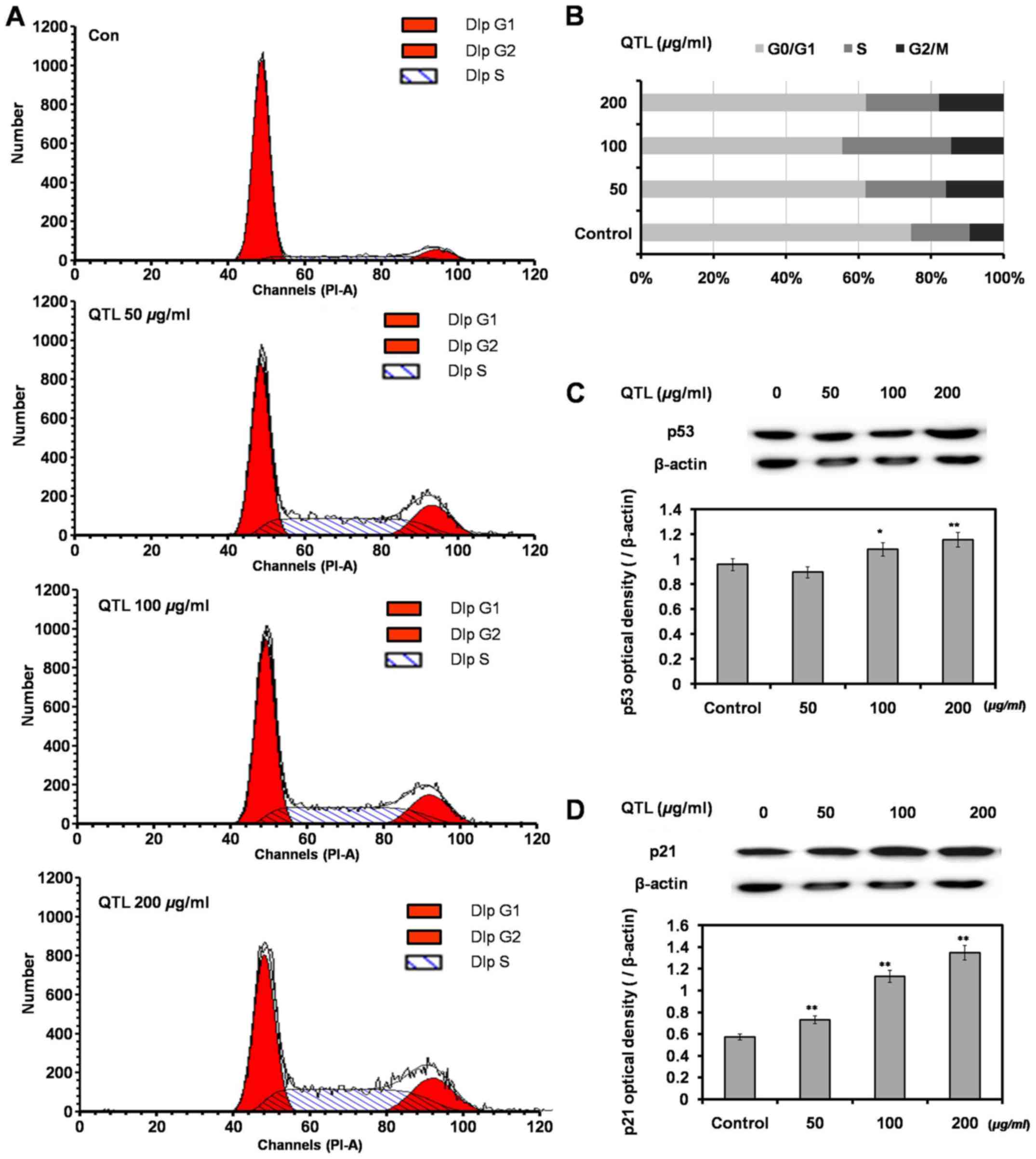
SW620 cells were exposed to QTL (50, 100 and 200
µg/ml) for 48 h, then washed, harvested and analyzed by flow
cytometry. A dose-dependent increase of cells in the G2/M phase
indicated the induction of a G2/M phase arrest (Fig. 2A and B). Western blot analysis
revealed that QTL increased the p53 and p21 protein expression
levels compared with the control groups (Fig. 2C and D), which was in accordance
with the results of the cell cycle analysis.
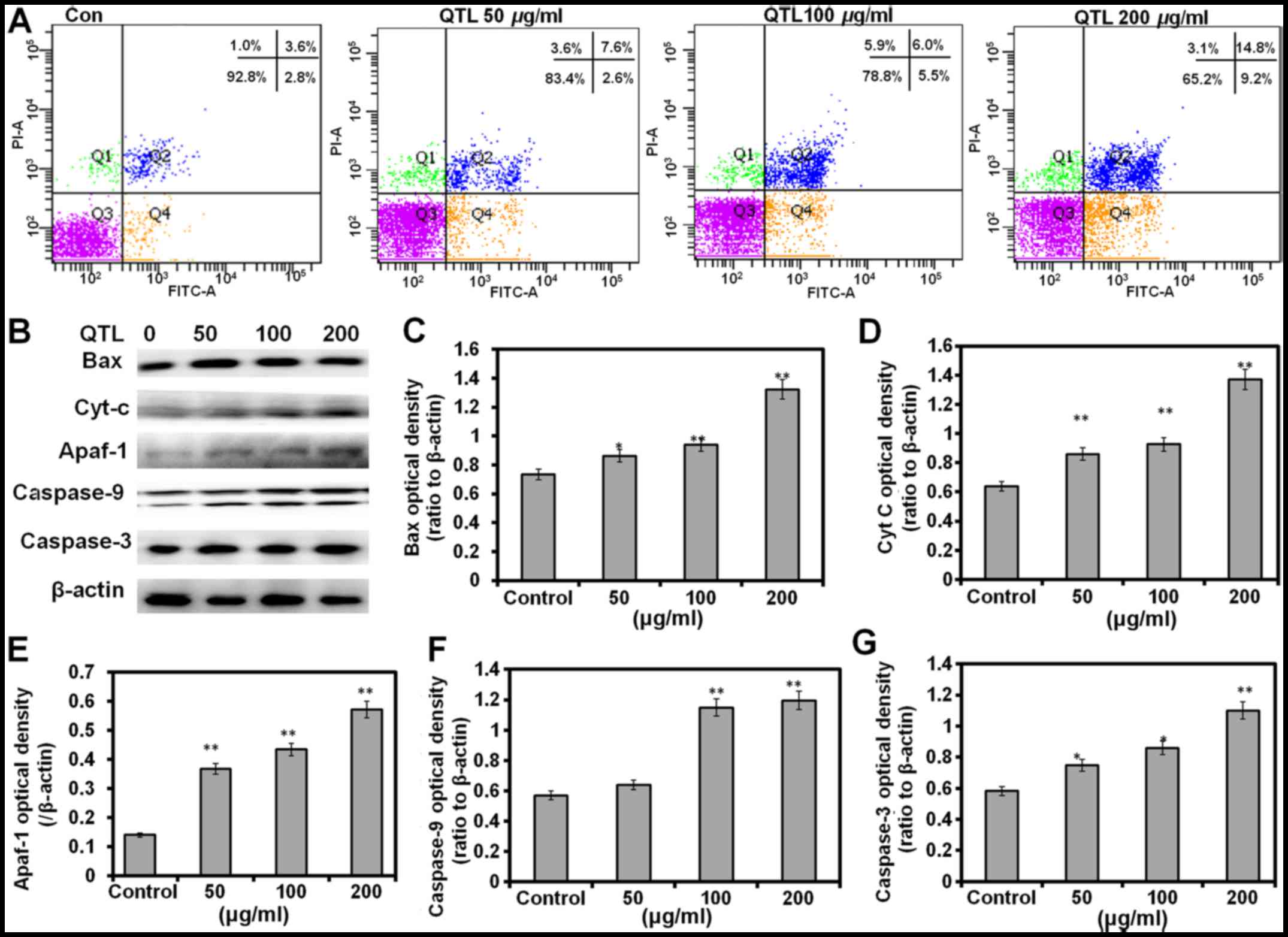
As shown in Fig. 3A,
untreated cells did not exhibit any significant signs of apoptosis,
whereas the cells became apoptotic after treatment with 50, 100 and
200 µg/ml QTL for 48 h, with apoptotic populations encompassing
~10.2, 11.5 and 24.0% of the total, respectively (Fig. 3A). The protein levels of apoptosis
markers Bax, cytochrome c caspase-9, Apaf-1 and caspase-3
exhibited a significant (P<0.01 in all cases) increase after
treatment with 200 µg/ml of QT compared with their respective
expression levels in the control groups (Fig. 3B-G). Our results therefore revealed
that the QTL-induced apoptotic response may be mediated by a
mitochondrial-dependent apoptotic pathway.
Analysis of oxidative stress
markers
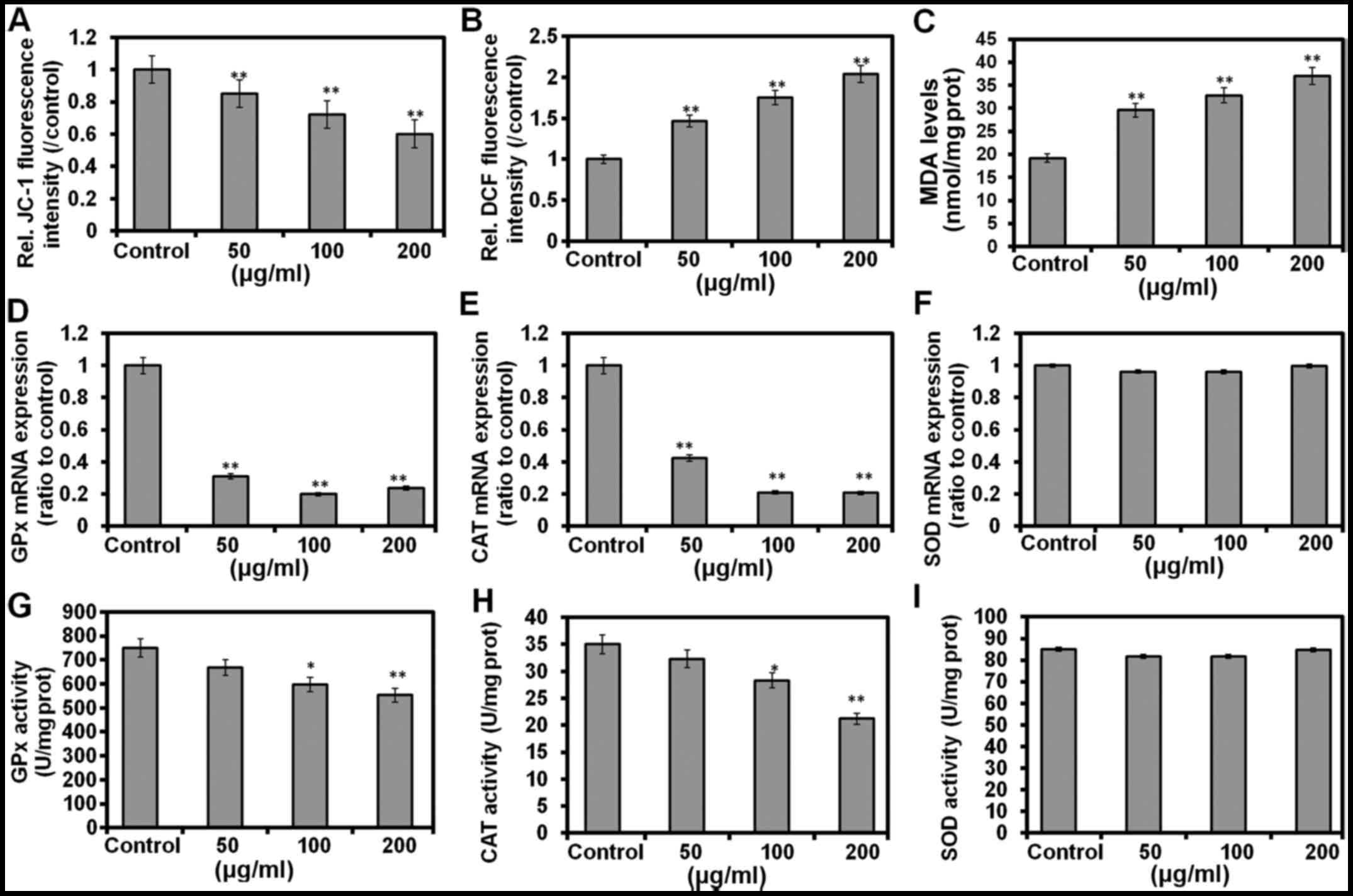
Alteration of the mitochondrial membrane potential
(ΔΨm) in the apoptotic cells was determined by JC-1 fluorescence,
and the results revealed a dose-dependent decrease in the membrane
potential (Fig. 4A). SW620 cells
treated with QTL (Fig. 4B) also
exhibited a significant (P<0.01) increase of intracellular ROS
production in a dose-dependent manner (48 h) as compared to the
control cells, indicating ROS-mediated apoptosis in these cells.
The MDA level was markedly increased by QTL (P<0.01 for all
doses; Fig. 4C). After incubation
with 50, 100 and 200 µg/ml of QTL for 48 h respectively, the
expression of CAT and GPx mRNAs was markedly decreased (P<0.01),
while the changes in the expression of SOD mRNA were
non-significantly (P>0.05) changed (Fig. 4D-F). Similarly, treatment with
25–100 µg/ml of QTL elicited no changes in the activity of SOD in
SW620 cells (Fig. 4I). Notably, a
significant decrease of GPx (P<0.01; Fig. 4G) and CAT (P<0.01; Fig. 4H) activities was observed after the
treatment of SW620 cells with 50 and 100 µg/ml of QTL for 48 h,
respectively.
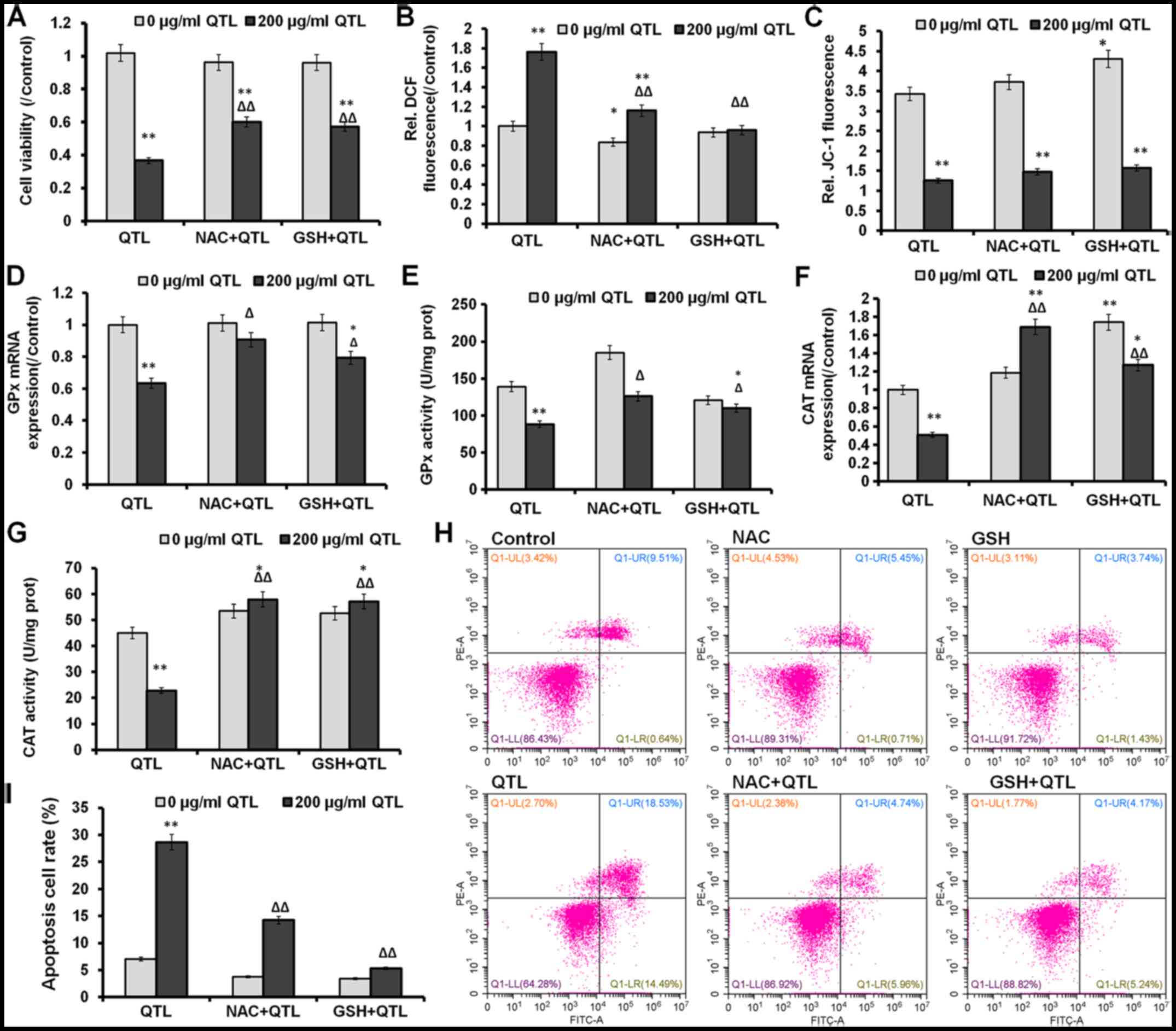
Effects of QTL in combination with NAC
or GSH on cell viability and oxidative stress markers in SW620
cells
SW620 cells were pretreated with 5 mM NAC (a ROS
scavenger) and 1 mM GSH (an antioxidant) and then exposed to 200
µg/ml of QTL for 24 h. The previously observed decrease of cell
viability in SW620 cells elicited by QTL was markedly (P<0.01)
reversed by NAC or GSH compared to cells treated with QTL only,
whereby NAC treatment only insignificantly (P>0.05) affected
cell viability (Fig. 5A).
Pretreatment with both NAC and GSH also partially abrogated the
QTL-mediated increase of ROS levels (P<0.01), while it had only
an insignificantly (P>0.05) effect on the QTL-mediated loss of
ΔΨm (Fig. 5C). In addition,
treatment with QTL partially (P<0.05 in both cases) reversed the
changes of GPx mRNA expression level and its activity (Fig. 5D and E). The pretreatment with both
NAC and GSH also significantly (P<0.01 in both cases) increased
the mRNA expression level and enzyme activity of CAT after
pretreatment with both NAC and GSH (Fig. 5F and G). As shown in Fig. 5H and I, QTL significantly increased
apoptosis (P<0.01) in SW620 cells, whereas QTL pretreatment with
both NAC and GSH markedly decreased (P<0.01 in both cases) the
apoptosis rate. Overall, these data demonstrated that QTL-induced
apoptosis in SW620 cells was mediated by the oxidative stress
pathway.
Discussion
Our data demonstrated that quercetrin from Toona
sinensis leaves (QTL) significantly decreased the viability of
the SW620 human colorectal cancer cell line, caused cell cycle
arrest in the G2/M phase and induced apoptosis. Cell cycle arrest
and apoptosis are common phenomena after DNA-damaging treatment. It
was reported that aqueous extracts from the leaves of T.
sinensis arrested SKOV3 ovarian cancer cells at the G2/M phase
and induced the apoptotic pathway (16). Cell cycle arrest or apoptosis in
response to DNA damage was mediated primarily by transcription
factor p53 (23). Moreover, p21 is
a very important checkpoint gene in the cell cycle, and it is also
regulated by the transcription of p53. Our findings are consistent
with previous studies demonstrating that QTL increased the
expression level of p53 and p21 proteins (24). Those results revealed that the
QTL-induced inhibition of SW620 cell growth was partially due to
the induction of G2/M arrest. Apoptosis is an ordered and
orchestrated cellular event by which cells undergo inducible
non-necrotic cellular suicide and thus it plays a crucial role in
preventing carcinogenesis (25).
Various phytochemicals induce apoptosis and change the morphology
of cancer cells due to their pro-apoptotic activity, and
consequently they are used for cancer prevention and cancer
chemotherapy (26). In the present
study, QTL was able to induce apoptosis in the SW620 cells in a
dose-dependent manner, as was demonstrated by flow cytometric
analysis. The tumor-suppressor p53 has been revealed to
differentially regulate Bcl-2 and Bax levels both in vitro
and in vivo (27). Using
western blotting, increases in the protein levels of the apoptosis
markers Bax, cytochrome c caspase-9, Apaf-1 and caspase-3
were detected indicating that QTL-induced apoptosis of SW620 was
tightly associated with the mitochondrial apoptotic pathway.
Mitochondria are key organelles crucial for cell
survival, and are conversely a source of ROS during apoptosis. The
loss of ΔΨm is often, but not always observed to be associated with
the early stages of apoptosis (28). The collapse of this potential is
believed to coincide with the opening of the mitochondrial
permeability transition pores, leading to the release of cytochrome
c into the cytosol. In the cytoplasm, cytochrome c
combines with caspase-9, Apaf-1 and dATP to form the apoptosome
complex which in turn activates caspase-9, −3 and −7. ROS have been
implicated as a second messengers in multiple signaling pathways
and can play a significant role in the process of apoptosis by
regulating the activity of certain enzymes involved in the cell
death pathway (29). In cancer
cells, ROS initiates cell transformation by causing alterations
during DNA replication (30).
Moreover, enhancement of ROS production has long been associated
with the apoptotic response induced by anticancer agents (31–33).
Several natural compounds, such as phenolic phytochemicals used for
cancer treatment have been shown to decrease the ΔΨm leading to the
increased generation of intracellular ROS and apoptosis (31). Similar results were observed in the
present study when SW620 colorectal cancer cells were treated with
QTL. Our findings along with other studies, therefore revealed that
QTL acts as an antiproliferative agent via the overproduction of
ROS and the loss of ΔΨm.
Furthermore, previous in vitro anticancer
studies of anticancer agents revealed that the increased ROS levels
in primary cancer cells were associated with a decrease of
antioxidants, such as SOD, GPx and catalase (CAT) (34). It is also known that many types of
human cancer cells can exist in a highly oxidative state due to the
decreased levels of protective antioxidant enzymes as compared to
their normal tissue counterparts (35). Natural phytochemicals are known to
deplete intracellular GSH and increase intracellular ROS to a level
that can cause cell death (36,37).
In the present study, treatment with QTL significantly decreased
the mRNA expression and activity of CAT and GPx in the SW620
cells.
Previous studies demonstrated that leaf extracts of
Toona sinensis are cytotoxic to DU145 prostate cancer cells,
due to the generation of reactive oxygen species (ROS) and
mitochondria-mediated apoptosis, which was reversed by the
antioxidants CAT and N-acetyl cysteine (NAC) (22). The present study also investigated
the effects of NAC (a well-known antioxidant and a precursor of GSH
biosynthesis) or GSH on QTL-treated SW620 cells. Treatment with
only NAC did not significantly affect cell viability, ΔΨm and mRNA
expression of CAT and GSH in SW620 cells. However, NAC
significantly mitigated QTL-induced cytotoxicity and ROS
production, which was accompanied by the decrease of CAT and GPx
mRNA expression in these cells. Treatment with only GSH slightly
decreased the reduction of cell viability, and did not enhance ROS
levels in SW620 cells. However, it should be noted that GSH
significantly mitigated the reduction of cell viability, the
increase of ROS levels and the decrease of CAT and GPx activity, as
well as the apoptosis rate in QTL-treated SW620 cells. Therefore,
our results support the notion that QTL-induced apoptotic cell
death in the SW620 cancer cell line was mediated by the excessive
generation of ROS.
In conclusion, we present evidence for the first
time that QTL has a potential role in suppressing human colorectal
cancer via inhibition of tumor cell proliferation, induction of
cell cycle arrest and apoptosis and enhancement of oxidative
stress, providing a therapeutic strategy for human colorectal
cancer treatment.
Acknowledgements
The present study was financed by the Special Fund
for Forest Scientific Research in the Public Welfare (201304811),
by the National Natural Science Foundation of China (31101266), and
by the Scientific and Technical Foundation of Shaanxi Province
(2014JM41005).
Glossary
Abbreviations
Abbreviations:
|
CRC
|
colorectal cancer
|
|
QTL
|
quercetrin from Toona sinensis
leaves
|
|
ROS
|
reactive oxygen species
|
|
ΔΨm
|
mitochondrial membrane potential
|
|
MDA
|
malondialdehyde
|
|
SOD
|
superoxide dismutase
|
|
GPx
|
glutathione peroxidase
|
|
CAT
|
catalase
|
|
NAC
|
N-acetyl cysteine
|
|
DMEM
|
Dulbecco's modified Eagle's medium
|
|
DMSO
|
dimethyl sulfoxide
|
|
RT-PCR
|
reverse transcriptase-polymerase chain
reaction
|
|
MTT
|
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide
|
|
DCFH-DA
|
2′,7′-dichlorofluorescin diacetate
|
|
FBS
|
fetal bovine serum
|
|
JC-1
|
5,5′,6,6/−tetra'hloro-1,1/,3,3/−tetraethylbenzimidazolyl
carbocyanine iodide
|
|
GSH
|
glutathione
|
References
|
1
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Redondo-Blanco S, Fernández J,
Gutiérrez-Del-Río I, Villar CJ and Lombó F: New insights toward
colorectal cancer chemotherapy using natural bioactive compounds.
Front Pharmacol. 8:1092017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Aghajanpour M, Nazer MR, Obeidavi Z,
Akbari M, Ezati P and Kor NM: Functional foods and their role in
cancer prevention and health promotion: A comprehensive review. Am
J Cancer Res. 7:740–769. 2017.PubMed/NCBI
|
|
4
|
Yang H, Gu Q, Gao T, Wang X, Chue P, Wu Q
and Jia X: Flavonols and derivatives of gallic acid from young
leaves of Toona sinensis (A. Juss.) Roemer and evaluation of
their anti-oxidant capacity by chemical methods. Pharmacogn Mag.
10:185–190. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yu WJ, Chang CC, Kuo TF, Tsai TC and Chang
SJ: Toona sinensis Roem leaf extracts improve antioxidant
activity in the liver of rats under oxidative stress. Food Chem
Toxicol. 50:1860–1865. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Yang HL, Chen SC, Lin KY, Wang MT, Chen
YC, Huang HC, Cho HJ, Wang L, Kumar KJ and Hseu YC: Antioxidant
activities of aqueous leaf extracts of Toona sinensis on
free radical-induced endothelial cell damage. J Ethnopharmacol.
137:669–680. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wu JG, Peng W, Yi J, Wu YB, Chen TQ, Wong
KH and Wu JZ: Chemical composition, antimicrobial activity against
Staphylococcus aureus and a pro-apoptotic effect in SGC-7901
of the essential oil from Toona sinensis (A. Juss.) Roem.
leaves. J Ethnopharmacol. 154:198–205. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Huang PJ, Hseu YC, Lee MS, Kumar Senthil
KJ, Wu CR, Hsu LS, Liao JW, Cheng IS, Kuo YT, Huang SY, et al: In
vitro and in vivo activity of gallic acid and Toona sinensis
leaf extracts against HL-60 human premyelocytic leukemia. Food Chem
Toxicol. 50:3489–3497. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yang CJ, Huang YJ, Wang CY, Wang CS, Wang
PH, Hung JY, Wang TH, Hsu HK, Huang HW, Kumar SP, et al:
Antiproliferative and antitumorigenic activity of Toona
sinensis leaf extracts in lung adenocarcinoma. J Med Food.
13:54–61. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Su YF, Yang YC, Hsu HK, Hwang SL, Lee KS,
Lieu AS, Chan TF and Lin CL: Toona sinensis leaf extract has
antinociceptive effect comparable with non-steroidal
anti-inflammatory agents in mouse writhing test. BMC Complement
Altern Med. 15:702015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yang HL, Huang PJ, Liu YR, Kumar KJ, Hsu
LS, Lu TL, Chia YC, Takajo T, Kazunori A and Hseu YC: Toona
sinensis inhibits LPS-induced inflammation and migration in
vascular smooth muscle cells via suppression of reactive oxygen
species and NF-κB signaling pathway. Oxid Med Cell Longev.
2014:9013152014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Hsiang CY, Hseu YC, Chang YC, Kumar KJ, Ho
TY and Yang HL: Toona sinensis and its major bioactive
compound gallic acid inhibit LPS-induced inflammation in nuclear
factor-κB transgenic mice as evaluated by in vivo bioluminescence
imaging. Food Chem. 136:426–434. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Liu HW, Huang WC, Yu WJ and Chang SJ:
Toona sinensis ameliorates insulin resistance via AMPK and
PPARγ pathways. Food Funct. 6:1855–1864. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hsieh TJ, Tsai YH, Liao MC, Du YC, Lien
PJ, Sun CC, Chang FR and Wu YC: Anti-diabetic properties of
non-polar Toona sinensis Roem extract prepared by
supercritical-CO2 fluid. Food Chem Toxicol. 50:779–789.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chang HC, Hung WC, Huang MS and Hsu HK:
Extract from the leaves of Toona sinensis roemor exerts
potent antiproliferative effect on human lung cancer cells. Am J
Chin Med. 30:307–314. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Chang HL, Hsu HK, Su JH, Wang PH, Chung
YF, Chia YC, Tsai LY, Wu YC and Yuan SS: The fractionated Toona
sinensis leaf extract induces apoptosis of human ovarian cancer
cells and inhibits tumor growth in a murine xenograft model.
Gynecol Oncol. 102:309–314. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kakumu A, Ninomiya M, Efdi M, Adfa M,
Hayashi M, Tanaka K and Koketsu M: Phytochemical analysis and
antileukemic activity of polyphenolic constituents of Toona
sinensis. Bioorg Med Chem Lett. 24:4286–4290. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yang HL, Chang WH, Chia YC, Huang CJ, Lu
FJ, Hsu HK and Hseu YC: Toona sinensis extracts induces
apoptosis via reactive oxygen species in human premyelocytic
leukemia cells. Food Chem Toxicol. 44:1978–1988. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang CY, Lin KH, Yang CJ, Tsai JR, Hung
JY, Wang PH, Hsu HK and Huang MS: Toona sinensis extracts
induced cell cycle arrest and apoptosis in the human lung large
cell carcinoma. Kaohsiung J Med Sci. 26:68–75. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhen H, Zhang Y, Fang Z, Huang Z, You C
and Shi P: Toona sinensis and Moschus decoction
induced cell cycle arrest in human cervical carcinoma HeLa cells.
Evid Based Complement Alternat Med. 2014:1212762014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Liu J, You L, Wang C and Liu R:
Antioxidization and antiproliferation of extract from leaves of
Toona sinensis. Zhong Nan Da Xue Xue Bao Yi Xue Ban.
37:42–47. 2012.PubMed/NCBI
|
|
22
|
Chen HM, Wu YC, Chia YC, Chang FR, Hsu HK,
Hsieh YC, Chen CC and Yuan SS: Gallic acid, a major component of
Toona sinensis leaf extracts, contains a ROS-mediated
anti-cancer activity in human prostate cancer cells. Cancer Lett.
286:161–171. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Mirzayans R, Andrais B, Kumar P and Murray
D: Significance of wild-type p53 signaling in suppressing apoptosis
in response to chemical genotoxic agents: Impact on chemotherapy
outcome. Int J Mol Sci. 18:182017.
|
|
24
|
Fischer M, Quaas M, Steiner L and Engeland
K: The p53-p21-DREAM-CDE/CHR pathway regulates G2/M cell cycle
genes. Nucleic Acids Res. 44:164–174. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Mbaveng AT, Kuete V and Efferth T:
Potential of Central, Eastern and Western Africa medicinal plants
for cancer therapy: Spotlight on resistant cells and molecular
targets. Front Pharmacol. 8:3432017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Shukla S, Meeran SM and Katiyar SK:
Epigenetic regulation by selected dietary phytochemicals in cancer
chemoprevention. Cancer Lett. 355:9–17. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Abraha AM and Ketema EB: Apoptotic
pathways as a therapeutic target for colorectal cancer treatment.
World J Gastrointest Oncol. 8:583–591. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Suzuki Y, Hasegawa H, Tsuji T, Tsuruda K,
Sasaki D, Ishihara K, Nagai K, Yanagihara K, Yamada Y and Kamihira
S: Relationships of diverse apoptotic death process patterns to
mitochondrial membrane potential (Δψm) evaluated by three-parameter
flow cytometric analysis. Cytotechnology. 65:59–70. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Marchi S, Giorgi C, Suski JM, Agnoletto C,
Bononi A, Bonora M, De Marchi E, Missiroli S, Patergnani S, Poletti
F, et al: Mitochondria-ros crosstalk in the control of cell death
and aging. J Signal Transduct. 2012:3296352012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Panieri E and Santoro MM: ROS homeostasis
and metabolism: A dangerous liason in cancer cells. Cell Death Dis.
7:e22532016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Bishayee K, Ghosh S, Mukherjee A,
Sadhukhan R, Mondal J and Khuda-Bukhsh AR: Quercetin induces
cytochrome-c release and ROS accumulation to promote
apoptosis and arrest the cell cycle in G2/M, in cervical carcinoma:
Signal cascade and drug-DNA interaction. Cell Prolif. 46:153–163.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhou Y, Shu F, Liang X, Chang H, Shi L,
Peng X, Zhu J and Mi M: Ampelopsin induces cell growth inhibition
and apoptosis in breast cancer cells through ROS generation and
endoplasmic reticulum stress pathway. PLoS One. 9:e890212014.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kim J, Kim J and Bae JS: ROS homeostasis
and metabolism: A critical liaison for cancer therapy. Exp Mol Med.
48:e2692016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Abdal Dayem A, Hossain MK, Lee SB, Kim K,
Saha SK, Yang GM, Choi HY and Cho SG: The role of reactive oxygen
species (ROS) in the biological activities of metallic
nanoparticles. Int J Mol Sci. 18:182017.
|
|
35
|
Banerjee K and Mandal M: Oxidative stress
triggered by naturally occurring flavone apigenin results in
senescence and chemotherapeutic effect in human colorectal cancer
cells. Redox Biol. 5:153–162. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Ferraresi R, Troiano L, Roat E, Lugli E,
Nemes E, Nasi M, Pinti M, Fernandez MI, Cooper EL and Cossarizza A:
Essential requirement of reduced glutathione (GSH) for the
anti-oxidant effect of the flavonoid quercetin. Free Radic Res.
39:1249–1258. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Ramos AM and Aller P: Quercetin decreases
intracellular GSH content and potentiates the apoptotic action of
the antileukemic drug arsenic trioxide in human leukemia cell
lines. Biochem Pharmacol. 75:1912–1923. 2008. View Article : Google Scholar : PubMed/NCBI
|