Introduction
Human pancreatic adenocarcinoma (HPAC) is recognized
as one of the most fatal malignant neoplasms. Although therapeutic
management has been improved using a variety of treatment
approaches, the 5-year survival rate of patients with HPAC remains
7.7%, making HPAC a cancer with one of the worst prognoses of any
major malignancy (1,2). Thus, it is quite urgent to understand
the molecular mechanisms by which HPAC initiates, progresses,
invades, and recurs to develop novel and effective therapeutic
strategies for HPAC.
MicroRNAs (miRNAs) are 18–25 nucleotide-long,
single-stranded, non-coding RNA molecules that negatively regulate
gene expression by binding to the regions of sequence
complementarity to the 3′ untranslated region (3′UTR) of mRNAs,
leading to either the degradation or translational inhibition of
mRNAs (3). Previous research has
found that miRNAs are involved in various cellular processes,
including cell proliferation, apoptosis, migration and invasion,
the cell cycle and stem cell renewal (4,5). In
addition, evidence suggests that miRNAs are aberrantly expressed in
various cancers, and they play oncogenic or tumor-suppressive roles
in these cancers (6–8). Differentially expressed miRNAs may
also offer novel therapeutic approaches for the more successful
treatment of HPAC (9,10).
As a member of the miRNA family, microRNA-217
(miR-217) has been shown to be involved in the initiation and
development of many human types of cancers, and functions as a
tumor suppressor in the majority of cancers, such as ovarian
cancer, osteosarcoma, esophageal squamous cell carcinoma,
hepatocellular carcinoma, pancreatic ductal adenocarcinoma and lung
and colorectal cancer (11–17). However, the clinical significance of
miR-217 in pancreatic carcinoma and its underlying molecular
pathways involved in pancreatic carcinoma have not been
investigated.
Tumor protein D52-like 2, known as hD54 in previous
studies (Tpd52l2), is a member of the TPD52 family which has been
implicated in multiple human cancers (18–20).
Further research has demonstrated that the TPD52 gene encodes
regulators of cancer cell proliferation, indicating that TPD52 may
be important for maintaining tumorigenesis and metastasis of cancer
cells (21). However, its direct
effect and molecular mechanism involved in HPAC development have
not yet been elucidated.
In the present study, we investigated the expression
and role of miR-217 in human HPAC. The results indicated that
miR-217 functions as a tumor suppressor in HPAC. In addition,
overexpression of Tpd52l2 reversed the effects of miR-217
restoration in AsPC-1 cells. All these results suggest that miR-217
exerts a tumor-suppressor role in HPAC, at least in part, by
repressing Tpd52l2 expression. The miR-217/Tpd52l2 axis may be a
new therapeutic strategy with which to treat patients with HPAC in
the future.
Materials and methods
Tissues and cell lines
Thirty samples of pancreatic carcinoma and
para-carcinoma tissues were collected from The First Affiliated
Hospital of Nanjing Medical University. The pancreatic carcinoma
and normal tissues were snap-frozen in liquid nitrogen immediately
after resection and stored at −80°C until use. The present study
was approved by the Research Ethics Committee of The First
Affiliated Hospital of Nanjing Medical University.
The AsPC-1 cell line used in the present study was
purchased from the American Type Culture Collection (ATCC;
Manassas, VA, USA). Cells were cultured according to ATCC
recommendations: RPMI-1640 medium with 10% fetal bovine serum (FBS)
(Gibco, Grand Island, NY, USA) and penicillin/streptomycin (Sigma,
St. Louis, MO, USA) supplement in a 37°C humidified incubator
supplied with 95% air and 5% CO2.
Antibodies
Commercially available antibodies were used for all
immunoblotting and immunofluorescence studies. Anti-Tpd52l2,
anti-PIK3CA, anti-p-PIK3CA, anti-AKT1/2 and anti-p-AKT1/2 were
obtained from Abcam Co. (Cambridge, UK). Anti-GAPDH was obtained
from Kangchen KangChen Bio-Tech (Shanghai, China). All secondary
antibodies used were obtained from Boster (Beijing, China).
Cell transfection
miR-217 mimic (miR-217) and the corresponding
negative control, the siRNAs targeting human Tpd52l2c (si-Tpd52l2)
and the corresponding negative control were obtained form
GenePharma Co., Ltd. (Shanghai, China). Tpd52l2 overexpressing
plasmids were construct using pCDNA3.1 (+) basic vectors in our
laboratory. These molecular productions were transfected into
AsPC-1 cells when cells were grown to 80–90% confluence, using
Lipofectamine 2000 (Invitrogen, Carlsbad, CA, USA) according to the
manufacturers instructions.
Quantitative real-time PCR
Tumor specimens and cells were subjected to total
RNA extraction using TRIzol reagent (Takara, Tokyo, Japan). To
detect RNA expression, complementary DNA (cDNA) was synthesized
using a Bestar qPCR RT kit (DBI Bioscience, Ludwigshafen, Germany)
according to the manufacturer's instructions. Amplification and
detection of RNA were performed using a Bestar qPCR RT kit under
the ABI 9700 PCR amplifier system (Applied Biosystems, Foster City,
CA, USA). In detail, the primer sequences are as follows: for
miR-150 detection, forward primer, ACA CTC CAG CTG GGT CTC CCA ACC
CTT GTA CC and reverse primer, CTC AAC TGG TGT CGT GGA GTC GGC AAT
TCA GTT GAG CAC TGG T; for miR-138 detection, forward primer, CTC
AAC TGG TGT CGT GGA GTC GGC AAT TCA GTT GAG TCC AAT C and reverse
primer, CTC AAC TGG TGT CGT GGA GTC GGC AAT TCA GTT GAG CGG CCT G;
for miR-217 detection, forward primer, ACA CTC CAG CTG GGT ACT GCA
TCA GGA ACT GA and reverse primer, CTC AAC TGG TGT CGT GGA GTC GGC
AAT TCA GTT GAG TCC AAT C; for miR-205 detection, forward primer,
ACA CTC CAG CTG GGT CCT TCA TTC CAC CGG AG and reverse primer, CTC
AAC TGG TGT CGT GGA GTC GGC AAT TCA GTT GAG CAG ACT C; for miR-31
detection, forward primer, ACA CTC CAG CTG GGA GGC AAG ATG CTG GCA
TA and reverse primer, CTC AAC TGG TGT CGT GGA GTC GGC AAT TCA GTT
GAG AGC TAT G; for miR-34 detection, forward primer, ACA CTC CAG
CTG GGT GGC AGT GTC TTA GCT GG and reverse primer, CTC AAC TGG TGT
CGT GGA GTC GGC AAT TCA GTT GAG ACA ACC A; for Tpd52l2 detection,
forward primer, CATGACGTGCAGGTCTCTAGC and reverse primer,
CATGACGTGCAGGTCTCTAGC. The miRNA and mRNA expression levels were
normalized to those of GAPDH and U6, respectively, using the
2−ΔΔCt method.
Western blotting
Monolayer cells were grown to 80% confluence and
then washed in ice-cold phosphate-buffered saline (PBS). Cells were
lysed in Universal protein extraction buffer (Beyotime, Shanghai,
China) applied with protease inhibitor phenylmethanesulfonyl
fluoride (PMSF) (Genebase Gen-Tech Co., Ltd., Shanghai, China) 48 h
after transfection. For western blotting, proteins were mixed with
5X SDS loading buffer [250 mM Tris-HCl (pH 6.8), 10% SDS, 0.5%
bromophenol blue, 50% glycerol, 5% β-mercaptoethanol] and heated to
95°C for 8 min before separating with 4–12% SDS-PAGE. Proteins were
then transferred to polyvinylidene difluoride membranes (Millipore,
Billerica, MA, USA), and detected with appropriate primary
antibodies and horseradish peroxidase-conjugated secondary
antibodies. Finally, the membranes were visualized using
chemiluminescent HRP substrate (Millipore). For Tpd52l2 detection,
anti-Tpd52l2 antibody was at 1:2,000 dilution, and the Tpd52l2
transfer membrane at a constant current 300 mA for 100 min. For
PIK3CA detection, the anti-PIK3CA antibody was at 1:2,500 dilution,
and the PIK3CA transfer membrane at a constant current 300 mA for
100 min. For p-PIK3CA detection, the anti-P-PIK3CA antibody was at
1:800 dilution, and the p-PIK3CA transfer membrane at a constant
current 300 mA for 100 min. For AKT1/2 detection, the anti-AKT1/2
antibody was at 1:1,000 dilution, and the AKT1/2 transfer membrane
at a constant current 300 mA for 50 min. For p-AKT1/2 detection,
the anti-P-AKT1/2 antibody was at 1:1,500 dilution, and the
p-AKT1/2 transfer membrane at a constant current 300 mA for 45 min.
For GAPDH detection, the anti-GAPDH antibody was at 1:10,000
dilution, and the GAPDH transfer membrane at a constant current 300
mA for 40 min.
Luciferase reporter assay
The target genes of miR-217 were predicted using
miRTarBase (http://mirtarbase.mbc.nctu.edu.tw/) and TargetScan
(http://www.Targetscan.org/). The
wild-type 3′UTR segment of the Tpd52l2 mRNA (not the full length of
Tpd52l2 3′UTR) containing miR-217 binding sites was amplified and
cloned into the Dual-luciferase reporter vector pGL3 (Promega,
Madison, WI, USA) termed as: Wt-Tpd52l2-3′UTR. A mutant construct
in the miR-217 binding sites of the Tpd52l2 3′UTR region also was
generated by synthesis, and subcloned into the pGL3-control vector
(Ambion, Foster City, CA, USA), and termed as Mut-Tpd52l2-3′UTR.
For the Dual-luciferase reporter assay, AsPC-1 cells were
transfected with miR-217 or normal control for 24 h, and then the
cells were transfected with the Wt/Mut-Tpd52l2-3′UTR reporter
plasmid using Lipofectamine 2000. Forty-eight hours later,
luciferase activity was measured using the Dual-Luciferase Reporter
Assay kit (Promega) according to the manufacturers instructions.
Renilla luciferase was used for normalization.
Transwell invasion assay
Serum-starved AsPC-1 cells (2×104) were
transferred to 8-µm pore size cell culture inserts coated with 0.1%
(w/v) collagen. The cells were incubated in serum-free medium with
or without thrombin/APC (both 10 nM), and the inserts were
incubated at 37°C for 10 h in serum-free medium with MCP-1 (50
ng/ml) as a chemoattractant. For microscopic analysis, cells on the
upper side of the Transwell membrane were removed with a cotton
swab after which the inserts were fixed and stained in a crystal
violet solution as previously described (22). The membranes were subsequently
mounted on a glass slide, and migrated cells were counted by light
microscopy. Cells were counted in 5 different fields using a
magnification of ×20.
Cell viability assay
Cell viability was determined using a Cell Counting
Kit-8 (CCK-8; Beyotime) based on WST-8. WST-8 existing in CCK-8
solution reacts with mitochondrial dehydrogenase, and results in
orange formazan deposition. A linear relationship between cell
number and shade was used to evaluate cell proliferation. Briefly,
AsPC-1 cells (2×103) were seeded into 96-well plates in
RPMI-1640 medium (100 µl) containing 10% FBS, and cultured
overnight. After transfection for 4 h, medium was renewed with
fresh medium, and continuously cultured for 72 h. CCK-8 solution
(10 µl) was added to each of the 96-well plates, and cultures were
incubated for 90 min at 37°C. Absorbance at 450 nm was measured
using an automatic microplate reader (BioTeke, Beijing, China). A
standard curve was constructed to deduce cell number. Experiments
were performed in sextuplicate and repeated 3 times. Results were
further analyzed using SPSS 20.0 software (SPSS, Inc., Chicago, IL,
USA).
Cell cycle assay
Cell cycle distribution was examined using a Cell
Cycle Analysis kit (MultiSciences, Susteren, The Netherlands).
Cells were fixed with 70% ethanol at 4°C overnight, and treated
with RNase A (0.02 mg/ml) in the dark at RT for 30 min, and then
stained with propidium iodide (PI) and analyzed using a FACSCalibur
flow cytometer (BD Biosciences, Franklin Lakes, NJ, USA) according
to the manufacturer's instructions.
Wound scratch assay
Scratch assays were essentially performed as
previously described (23). In
detail, cells were seeded into 6-well plates in RPMI-1640 medium
supplemented with 10% FBS. After the cells formed a confluent
monolayer, a scratch was created in the center of the monolayer
with a sterile p200 pipette tip. Next, the medium was removed and
cells were washed with serum-free medium to remove floating debris.
The cells were subsequently incubated for 18 h with serum-free
medium with plasmids or control transfection. The ability of cells
to close the wound was assessed by comparing the 0 and 24 h
phase-contrast micrographs of 6 marked points along the wounded
area. The percentage of non-recovered wound area was calculated by
dividing the non-recovered area after 24 h by the initial area at 0
h.
Statistical analysis
Data were analyzed using SPSS 20.0 software package
(SPSS, Inc.) with independent samples t-test between two groups.
All values were represented as mean ± standard deviation (SD).
Statistical significance was defined as P<0.05.
Results
miR-217 expression is downregulated in
HPAC tissues
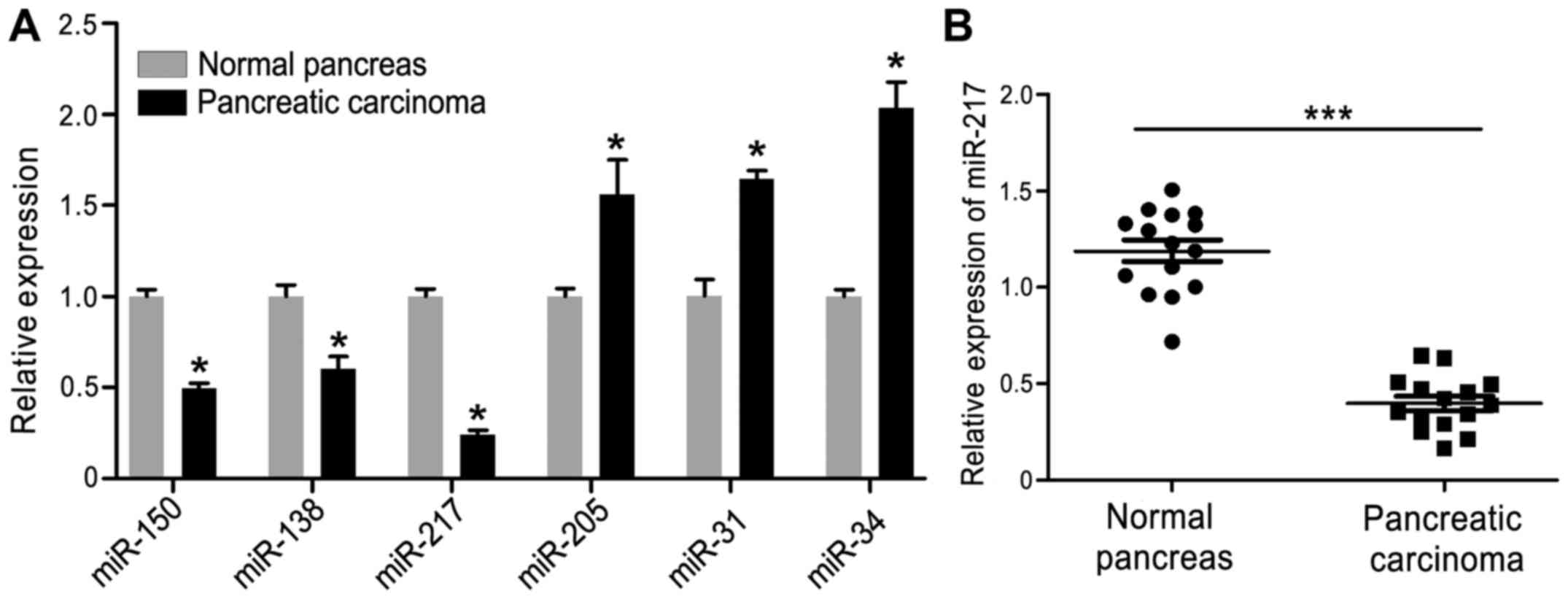
First, we examined various miRNAs, including the
difference in expression of miR-150, miR-138, miR-217, miR-205,
miR-31 and miR-34, in HPAC and normal pancreas tissues. As shown in
Fig. 1A, the expression of miR-150,
miR-138 and miR-217 were downregulated in HPAC tissues and miR-217
showed the largest decrease when compared with the other miRNAs,
while the expression of miR-205, miR-31 and miR-34 were increased.
Thus, miR-217 was selected for subsequent study. Furthermore, to
investigate the potential biological role of miR-217 expression in
human HPAC progression, we evaluated miR-217 expression in 15 HPAC
tissues and 15 normal pancreatic tissues by quantitative RT-PCR
(qRT-PCR). As shown in Fig. 1B, the
expression of miR-217 was significantly decreased in the HPAC
tissues compared with that noted in the normal tissues.
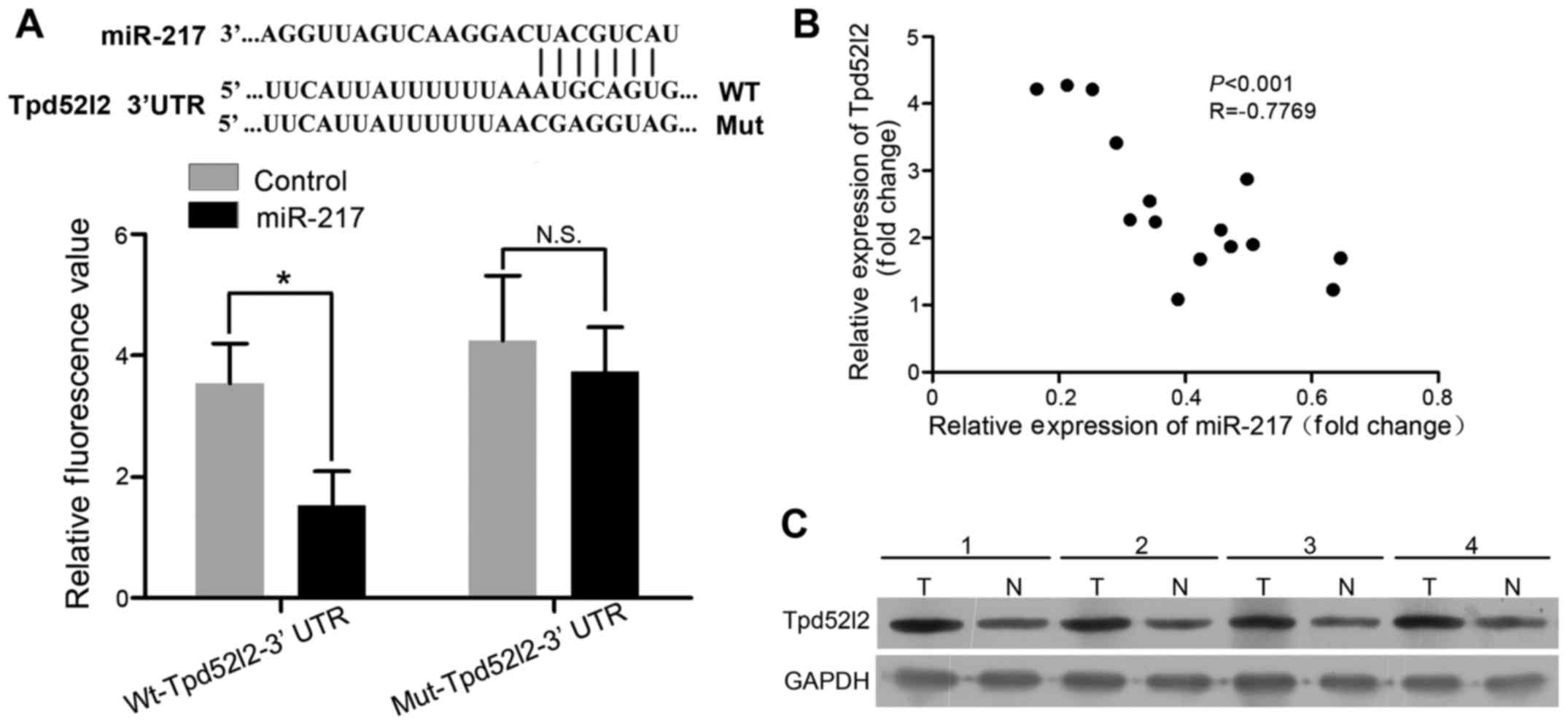
Tpd52l2 is a target of miR-217 in HPAC
cells
The target of miR-217 in HPAC cells was identified
used bioinformatics software (miRTarBase and TargetScan). It was
found that Tpd52l2 3′UTR has a binding sequences for miR-217 at
position 581–587 (Fig. 2A, upper
panel). To further verify Tpd52l2 as a direct target of miR-217,
luciferase activity assay was performed. We found that miR-217
significantly inhibited the luciferase activity of 3′UTR of Tpd52l2
in AsPC-1 cells (Fig. 2A, lower
panel). Further experiments were performed to investigate the
expression of Tpd52l2 in 4 paired HPAC tissues and normal pancreas
tissues. The results showed that Tpd52l2 expression was upregulated
in the HPAC tissues (Fig. 2C). The
relationship between miR-217 and Tpd52l2 expression in patients
with HPAC was also investigated. Spearman's correlation analysis
showed an inverse correlation between miR-217 expression and
Tpa52l2 mRNA level in the HPAC tissues (r=−0.7769; P<0.001)
(Fig. 2B). These data suggest that
Tpd52l2 is a direct target of miR-217.
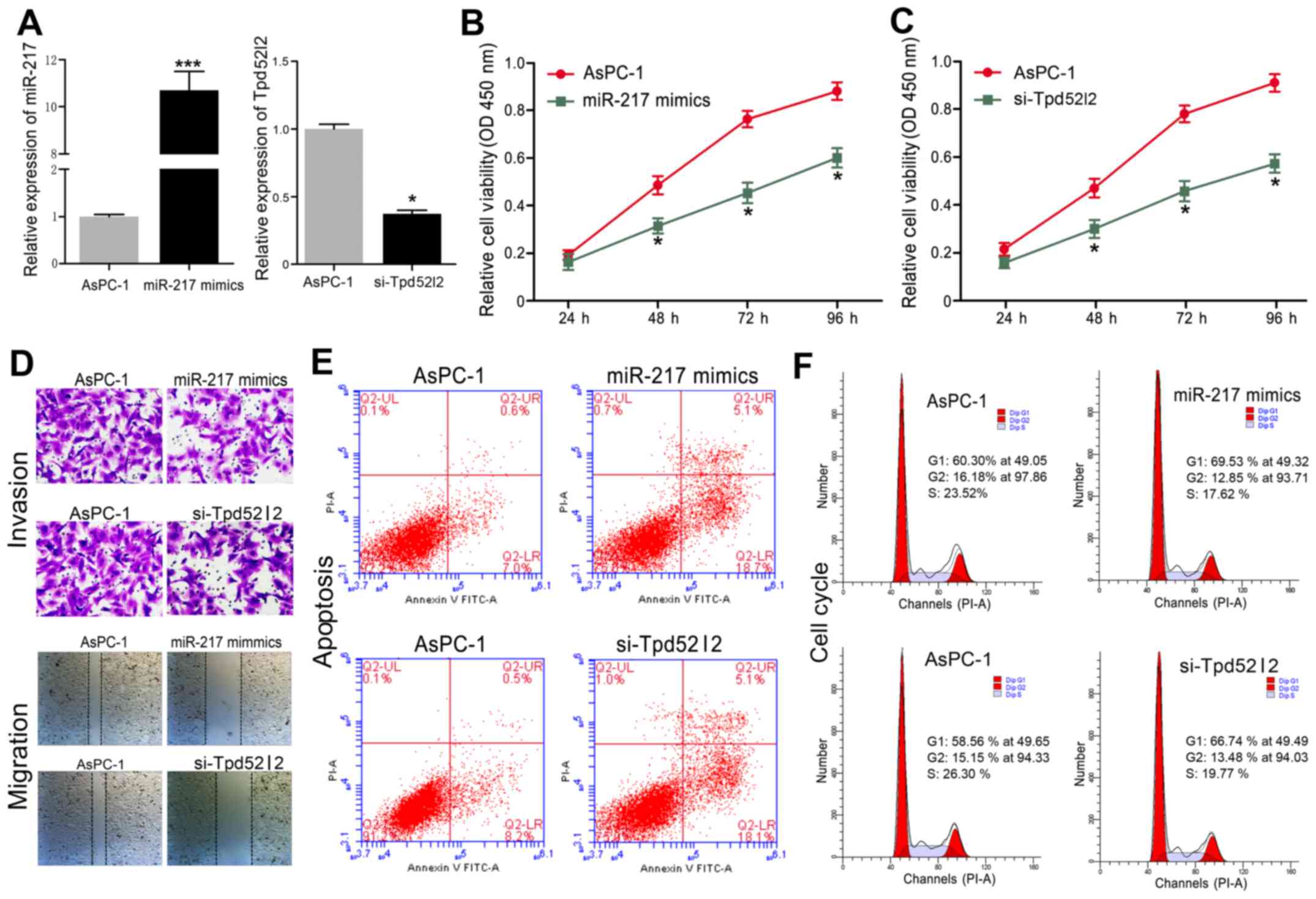
Downregulation of Tpd52l2 exhibits an
effect similar to that of miR-217 overexpression in HPAC cells
To investigate the biological functions of Tpd52l2
in HPAC cells, endogenous Tpd52l2 was knocked down in AsPC-1 cells
with specific siRNAs against Tpd52l2 (si-Tpd52l2). In addition,
miR-217 mimics were transfected into AsPC-1 cells. We found that
the mRNA level of miR-217 was increased by miR-217 mimic
transfection, and Tpd52l2 was significantly inhibited in AsPC-1
cells by si-Tpd52l2 (Fig. 3A).
Overexpression of miR-217 in AsPC-1 cells significantly inhibited
cell proliferation (Fig. 3B),
invasion and migration (Fig. 3D),
induced apoptosis (Fig. 3E) and
caused cell cycle arrest at the G0/G1 phase (Fig. 3F). Similarly, downregulation of
Tpd52l2 in AsPC-1 cells significantly inhibited cell proliferation
(Fig. 3C), invasion and migration
(Fig. 3D), induced apoptosis
(Fig. 3E) and cell cycle arrest
(Fig. 3F). These results suggest
that silencing of Tpd52l2 had a similar effect as miR-217
overexpression in the HPAC cells.
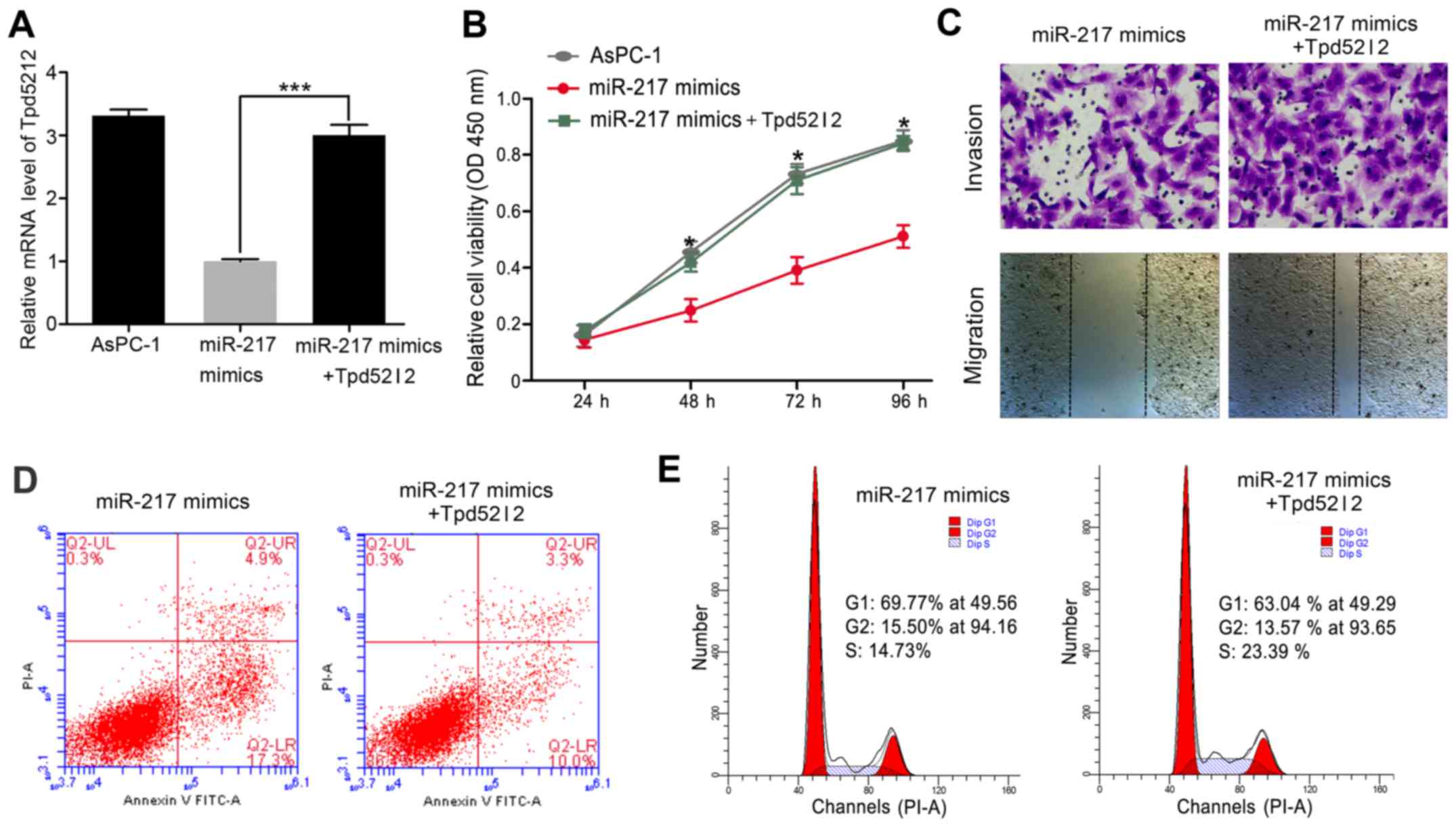
Overexpression of Tpd52l2 reverses the
effects of miR-217 in HPAC cells
To investigate the functional relevance of the
targeting of Tpd52l2 by miR-217, we assessed whether Tpd52l2
overexpression reverses the inhibitory effects of miR-217
restoration on AsPC-1 cell proliferation, migration, invasion,
apoptosis and cell cycle distribution. AsPC-1 cells were
co-transfected with miR-217 or control mimics and Tpd52l2
overexpression plasmids. qRT-PCR analysis was used to validate the
Tpd52l2 mRNA level in the rescue experiment (Fig. 4A). In addition, our results also
showed that the exogenous expression of Tpd52l2 reversed the
effects of miR-217 overexpression on cell proliferation, migration,
invasion, apoptosis and cell cycle distribution (Fig. 4B-E).
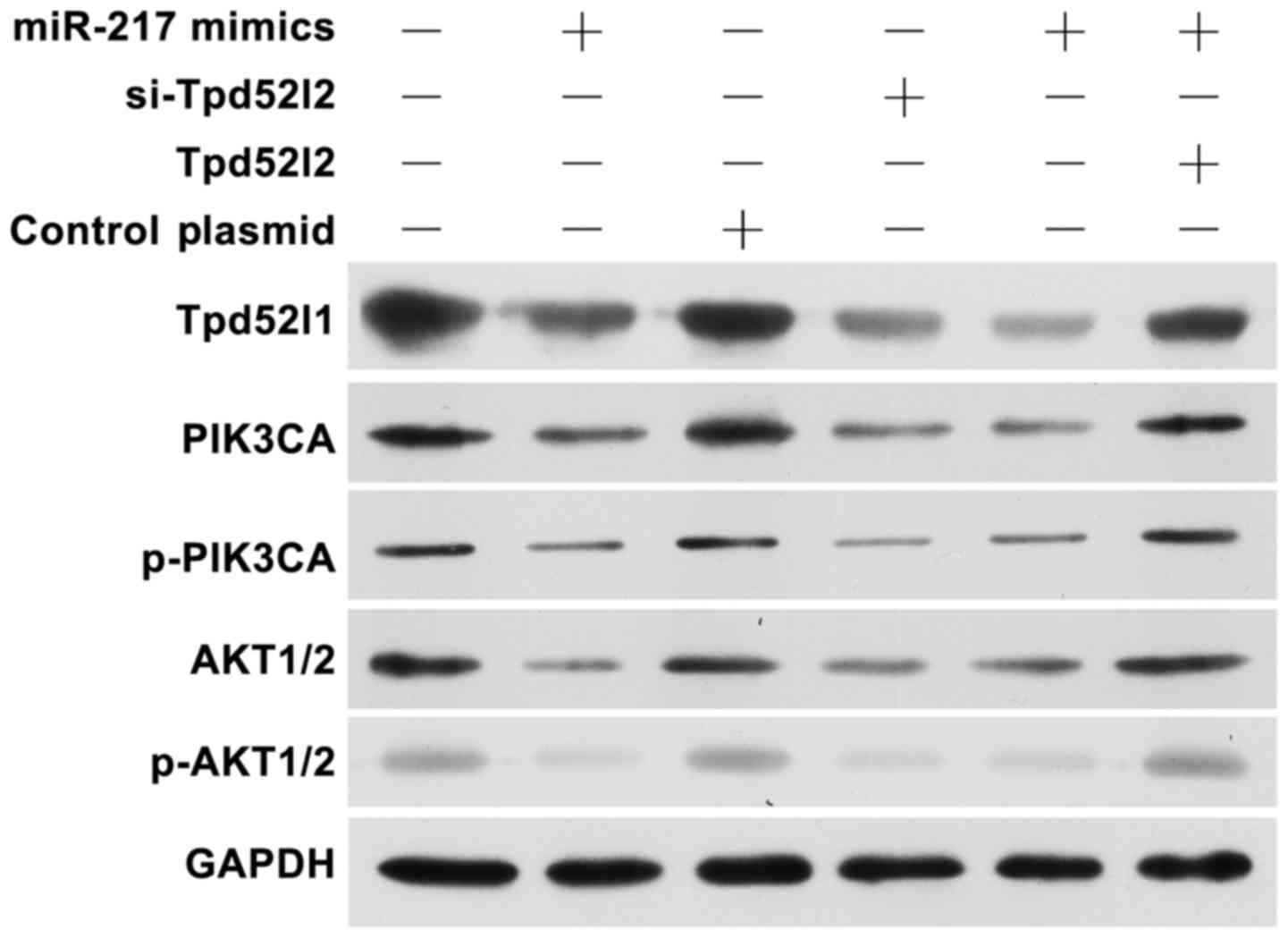
Overexpression of miR-217 or knockdown
of Tpd52l2 inhibits the PIK3CA/AKT signaling pathways
To further explain the effect of miR-217/Tpd52l2 on
HPAC cells, we detected the PIK3CA/AKT signaling pathways in
miR-217 mimic, si-Tpb52l2 or Tpd52l2 overexpression plasmid
transfected cells (Fig. 5). The
results showed that overexpression of miR-217 or knockdown of
Tpd52l2 suppressed the PIK3CA/p-PIK3CA and AKT1/2/p-AKT1/2 protein
levels. In addition, following co-transfection of miR-217 and the
Tpd52l2 plasmid, the inhibitory effect of miR-217 was reversed. All
these data indicated that upregulation of miR-217 exerts an
inhibitory effect on HPAC growth and metastasis partially by
suppressing Tpd52l2 expression.
Discussion
Evidence shows that miRNAs are involved in
tumorigenesis and metastasis of various types of human cancers,
including HPAC (15,24,25).
Aberrant expression of miR-217 has been found in various human
cancers (11–17). Frequently, miR-217 expression is
downregulated and acts as a tumor suppressor, while it is
overexpressed and functions as an oncogene in breast cancer
(17) and B cell lymphoma (26). However, the function and relevant
mechanisms of miR-217 in HPAC have not been comprehensively
identified.
In the present study, we found that the expression
of miR-217 was significantly downregulated in HPAC tissues and
cells. We also demonstrated that restoration of miR-217 expression
in HPAC cells inhibited proliferation, migration, and invasion,
induced apoptosis and caused cell cycle arrest. These results
suggested that miR-217 functions as a tumor suppressor in HPAC.
To further investigate the molecular mechanism of
the tumor-suppressor role of miR-217 in HPAC, we used
bioinformatics software to identify the target of miR-217 in HPAC
cells. Tpd52l2 3′UTR was found to have a binding sequence for
miR-217 at position 581–587. Luciferase activity assay, qRT-PCR and
western blot assay further confirmed that Tpd52l2 is a target gene
of miR-217. Tpd52l2 is a member of the TPD52 family which has been
implicated in multiple human cancers (18–20).
Studies have demonstrated that TPD52 may be important for
maintaining tumorigenesis and metastasis of cancer cells (21). In the present study, we found that
Tpd52l2 expression was upregulated and inversely correlated with
miR-217 expression in HPAC tissues. In addition, we also found that
downregulation of Tpd52l2 had a similar effect as the restoration
of miR-217 expression in HPAC cells, and overexpression of Tpd52l2
reversed the effects of miR-217 in AsPC-1 cells. These results
suggest that miR-217 exerts a tumor-suppressor role in HPAC, at
least in part, by suppressing Tpd52l2 expression.
In summary, the present study demonstrated that
miR-217 expression is downregulated in HPAC cells and tissues, and
that restoration of miR-217 expression inhibited proliferation,
migration and invasion, induced apoptosis and cell cycle arrest of
HPAC cells by suppressing Tpd52l2. PIK3CA/AKT signaling pathways
generally act to promote survival through inhibition of
pro-apoptotic factors and activation of anti-apoptotic factors
(27). Activation of the PIK3CA/AKT
pathway signals through mTOR to promote protein translation and
cell cycling, and other effectors participate in regulation of
transcription, apoptosis and cellular metabolism (28–30).
In addition, PIK3A, at the top of the pathway, is an upstream
catalytic enzyme that, when active, leads to cell growth and
proliferation and in particular inhibition of cell death (31). Finally, we found that overexpression
of miR-217 or knockdown of Tpd52l2 inhibited the PIK3CA/AKT
signaling pathways. In addition, this may explain the effect of
miR-217/Tpd52l2 on HPAC development. Taken together, targeting of
the miR-217/Tpd52l2 axis may be a new therapeutic strategy by which
to treat patients with HPAC.
Acknowledgements
The present study was supported by grants from the
National Nature Science Foundation of China (nos. 81672449 and
81272239). The present study was also supported by Talents Planning
of Six Summit Fields of Jiangsu Province (WSN-025).
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2015. CA Cancer J Clin. 65:5–29. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ryan DP, Hong TS and Bardeesy N:
Pancreatic adenocarcinoma. N Engl J Med. 371:1039–1049. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Orang Valinezhad A, Safaralizadeh R and
Kazemzadeh-Bavili M: Mechanisms of miRNA-mediated gene regulation
from common downregulation to mRNA-specific upregulation. Int J
Genomics. 2014:9706072014.PubMed/NCBI
|
|
4
|
Bushati N and Cohen SM: microRNA
functions. Annu Rev Cell Dev Biol. 23:175–205. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hwang HW and Mendell JT: MicroRNAs in cell
proliferation, cell death, and tumorigenesis. Br J Cancer. Suppl
96:R40–R44. 2007.PubMed/NCBI
|
|
6
|
Kwak PB, Iwasaki S and Tomari Y: The
microRNA pathway and cancer. Cancer Sci. 101:2309–2315. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zhang S, Liu X, Liu J, Guo H, Xu H and
Zhang G: PGC-1 alpha interacts with microRNA-217 to functionally
regulate breast cancer cell proliferation. Biomed Pharmacother.
85:541–548. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yates LA, Norbury CJ and Gilbert RJ: The
long and short of microRNA. Cell. 153:516–519. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Cioffi M, Trabulo SM, Sanchez-Ripoll Y,
Miranda-Lorenzo I, Lonardo E, Dorado J, Vieira Reis C, Ramirez JC,
Hidalgo M, Aicher A, et al: The miR-17-92 cluster counteracts
quiescence and chemoresistance in a distinct subpopulation of
pancreatic cancer stem cells. Gut. 64:1936–1948. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Neesse A and Gress TM: Emerging role of
microRNAs to tackle drug resistance in pancreatic cancer. Gut.
64:1842–1843. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Li J, Li D and Zhang W: Tumor suppressor
role of miR-217 in human epithelial ovarian cancer by targeting
IGF1R. Oncol Rep. 35:1671–1679. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wei R, Deng Z and Su J: miR-217 targeting
Wnt5a in osteosarcoma functions as a potential tumor suppressor.
Biomed Pharmacother. 72:158–164. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Su J, Wang Q, Liu Y and Zhong M: miR-217
inhibits invasion of hepatocellular carcinoma cells through direct
suppression of E2F3. Mol Cell Biochem. 392:289–296. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhao WG, Yu SN, Lu ZH, Ma YH, Gu YM and
Chen J: The miR-217 microRNA functions as a potential tumor
suppressor in pancreatic ductal adenocarcinoma by targeting KRAS.
Carcinogenesis. 31:1726–1733. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Guo J, Feng Z, Huang Z, Wang H and Lu W:
MicroRNA-217 functions as a tumour suppressor gene and correlates
with cell resistance to cisplatin in lung cancer. Mol Cells.
37:664–671. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wang B, Shen ZL, Jiang KW, Zhao G, Wang
CY, Yan YC, Yang Y, Zhang JZ, Shen C, Gao ZD, et al: MicroRNA-217
functions as a prognosis predictor and inhibits colorectal cancer
cell proliferation and invasion via an AEG-1 dependent mechanism.
BMC Cancer. 15:4372015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhang Q, Yuan Y, Cui J, Xiao T and Jiang
D: miR-217 promotes tumor proliferation in breast cancer via
targeting DACH1. J Cancer. 6:184–191. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Byrne JA, Balleine RL, Fejzo Schoenberg M,
Mercieca J, Chiew YE, Livnat Y, St Heaps L, Peters GB, Byth K,
Karlan BY, et al: Tumor protein D52 (TPD52) is overexpressed and a
gene amplification target in ovarian cancer. Int J Cancer.
117:1049–1054. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Rubin MA, Varambally S, Beroukhim R,
Tomlins SA, Rhodes DR, Paris PL, Hofer MD, Storz-Schweizer M,
Kuefer R, Fletcher JA, et al: Overexpression, amplification, and
androgen regulation of TPD52 in prostate cancer. Cancer Res.
64:3814–3822. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhu H, Lam DC, Han KC, Tin VP, Suen WS,
Wang E, Lam WK, Cai WW, Chung LP and Wong MP: High resolution
analysis of genomic aberrations by metaphase and array comparative
genomic hybridization identifies candidate tumour genes in lung
cancer cell lines. Cancer Lett. 245:303–314. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Byrne JA, Mattei MG and Basset P:
Definition of the tumor protein D52 (TPD52) gene family
through cloning of D52 homologues in human (hD53) and
mouse (mD52). Genomics. 35:523–532. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Queiroz KC, Shi K, Duitman J, Aberson HL,
Wilmink JW, van Noesel CJ, Richel DJ and Spek CA:
Protease-activated receptor-1 drives pancreatic cancer progression
and chemoresistance. Int J Cancer. 135:2294–2304. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lin C, von der Thüsen J, Daalhuisen J, ten
Brink M, Crestani B, van der Poll T, Borensztajn K and Spek CA:
Pharmacological targeting of protease-activated receptor 2 affords
protection from bleomycin-induced pulmonary fibrosis. Mol Med.
21:576–583. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chang W, Liu M, Xu J, Fu H, Zhou B, Yuan T
and Chen P: MiR-377 inhibits the proliferation of pancreatic cancer
by targeting Pim-3. Tumour Biol. 37:14813–14824. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yonemori K, Kurahara H, Maemura K and
Natsugoe S: MicroRNA in pancreatic cancer. J Hum Genet. 62:33–40.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Slattery ML, Lundgreen A, Herrick JS, Caan
BJ, Potter JD and Wolff RK: Associations between genetic variation
in RUNX1, RUNX2, RUNX3, MAPK1 and eIF4E and risk of
colon and rectal cancer: Additional support for a TGF-β-signaling
pathway. Carcinogenesis. 32:318–326. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Nathan N, Keppler-Noreuil KM, Biesecker
LG, Moss J and Darling TN: Mosaic disorders of the
PI3K/PTEN/AKT/TSC/mTORC1 signaling pathway. Dermatol Clin.
35:51–60. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Samuels Y and Velculescu VE: Oncogenic
mutations of PIK3CA in human cancers. Cell Cycle. 3:1221–1224.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Hennessy BT, Smith DL, Ram PT, Lu Y and
Mills GB: Exploiting the PI3K/AKT pathway for cancer drug
discovery. Nat Rev Drug Discov. 4:988–1004. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhao L and Vogt PK: Class I PI3K in
oncogenic cellular transformation. Oncogene. 27:5486–5496. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Fleisher B, Clarke C and Ait-Oudhia S:
Current advances in biomarkers for targeted therapy in
triple-negative breast cancer. Breast Cancer. 8:183–197.
2016.PubMed/NCBI
|