Introduction
Hepatocellular carcinoma (HCC) is one of the most
common malignant tumors in China, and an increasing incidence has
been reported worldwide (1).
Although multimodal treatments such as surgical resection, liver
transplantation, chemotherapy and radiotherapy or a combination of
these options have been performed in recent years, the long-term
survival of HCC patients remains extremely poor due to a high
incidence of recurrence and metastasis (2,3).
Therefore, understanding of the underlying molecular mechanisms
leading to the initiation and metastasis of HCC are required for
the development of more effective therapeutic strategies.
MicroRNAs (miRNAs) are a class of short, endogenous,
non-coding RNAs known to negatively regulate the expression of
protein-coding genes by inhibiting translation or inducing
messenger RNA (mRNA) degradation by binding to the 3′ untranslated
regions (3′UTRs) of target genes (4,5). Since
miRNAs are able to regulate multiple targets (6), miRNAs are involved in the regulation
of various biological processes, including proliferation,
apoptosis, metastasis and angiogenesis (7). Alterations in miRNA expression have
been reported to play essential roles in tumorigenesis and tumor
progression processes, and function as oncogenes or tumor
suppressors (8). Accumulating
evidence has suggested that alterations in miRNA expression
contribute to HCC development and progression, and can serve as
diagnostic and prognostic markers, and therapeutic agents for HCC
(9,10).
The aberrant expression of microRNA-363 (miR-363)
and its role in tumor pathogenesis and progression have already
been reported in gallbladder (11),
colorectal (12), gastric (13), and head and neck (14), as well as in breast cancer (15). In HCC, miR-363 expression was found
to be downregulated and inhibited cell proliferation by
downregulating the expression of S1PR (16), and decreased the cisplatin
resistance of HCC cells, partly by targeting Mcl-1 (17). However, the detailed biological
role, particularly with respect to metastasis in HCC, has not been
fully elucidated. In the present study, we found that miR-363 was
decreased in HCC tissues and cell lines. In addition, a low
expression level of miR-363 was associated with tumor
differentiation, tumor-node-metastasis (TNM) stage and lymph node
metastasis. We found that miR-363 overexpression significantly
inhibited HCC cell proliferation, migration and invasion in
vitro and suppressed tumor growth in vivo by targeting
E2F transcription factor 3 (E2F3). These results suggest that
miR-363 may be a novel target for HCC.
Materials and methods
Patients and clinical tissue
specimens
Forty pairs of primary HCC and adjacent non-tumor
tissues were obtained from patients undergoing surgery at the
Department of Hepatopancreatobiliary Surgery, The First Hospital,
Jilin University (Changchun, China). The collection and use of
patient samples was approved by the Ethics Committee of the First
Hospital of Jilin University, and written informed consent was
obtained from all patients whose biological samples were used in
the study. All samples were immediately snap-frozen in liquid
nitrogen and stored at −80°C until use.
Cell culture and transfection
The 4 HCC cell lines (SMMC-7721, Hep3B, HepG2 and
Huh-7) as well as the normal human hepatocyte cell line HL-7702,
were purchased from the Institute of Biochemistry and Cell Biology,
Chinese Academy of Sciences (Shanghai, China). All cell lines were
maintained in high glucose Dulbeccos modified Eagles medium (DMEM)
supplemented with 10% fetal bovine serum (FBS) (both from Gibco,
Gaithersburg, MD, USA) at 37°C in a humidified chamber supplemented
with 5% CO2.
The miR-363 mimic and the appropriate negative
control oligonucleotide (miR-NC) were purchased from RiboBio
(Guangzhou, China). The E2F3 overexpression vector was amplified
and inserted into the pcDNA3.0 vector (Invitrogen Life
Technologies, Grand Island, NY, USA) at the
BamHI/EcoRI restriction sites. Transfection was
performed using Lipofectamine 2000 (Invitrogen) in accordance with
the manufacturer's protocol.
RNA isolation and qRT-PCR
Total RNA containing miRNA and mRNA was extracted
from tissue samples and cells using TRIzol® reagent
(Invitrogen) according to the manufacturer's instructions. RNA was
reverse transcribed to cDNA using Prime Script First Strand cDNA
Synthesis kit (Takara, Dalian, China) following the manufacturer's
protocol. The cDNAs were subjected to qRT-PCR using SYBR Premix Ex
Taq (Takara) under ABI 7900 Fast System (Applied Biosystems, Foster
City, CA, USA) to detect miR-363 and E2F3 mRNA. miR-363 and U6
primers were purchased from RiboBio. The primers for E2F3 and GAPDH
used in the present study were as previously described (18). U6 and GAPDH were used as internal
controls to detect miR-363 and E2F3 mRNA, respectively. Relative
expression of miR-363 and E2F5 was determined using the
2−ΔΔCt method.
Cell proliferation and colony
formation assays
Cell proliferation was assessed using Cell Counting
Kit-8 (CCK-8) assay (Dojindo Laboratories, Kumamoto, Japan)
according to the manufacturer's instructions. In brief,
5×103 transfected cells/well were seeded in 96-well
plates and cultured for 24–72 h. At the indicated time, an amount
of 20 µl of CCK-8 solution was added to each well and incubation
was carried out for 4 h. Absorbance at 450 nm was measured using an
enzyme-linked immunosorbent assay reader (Thermo Labsystems,
Helsinki, Finland).
Colony formation ability was determined using
plating at 500 transfected cells/well into 6-well plates and
culturing for 14 days. Cells were fixed with methanol and stained
with 0.1% crystal violet for 15 min. Stained colonies were imaged
and counted by a light microscope (Olympus, Tokyo, Japan).
Cell migration and invasion
assays
Cell migration was measured using a wound healing
assay. In brief, transfected cells were cultured in 6-well plates
(5×104 cells/well) and grown to confluency.
Subsequently, an artificial homogenous wound was scratched into the
monolayer using a sterile plastic micropipette tip, and then the
cells were cultured under standard conditions for 24 h. Following
several washes, the cultured cells were fixed and observed using an
inverted microscope (Olympus).
The invasion assay was performed using Transwell
insert chambers (Corning Inc., Corning, NY, USA). For the Transwell
invasion assay, 1×105 transfected cells were seeded into
each well of the upper chamber of Matrigel-coated inserts in
serum-free medium. In the lower chamber, DMEM with 20% FBS was
added to serve as a chemoattractant. After incubating for 24 h at
37°C with 5% CO2, the non-invading cells were gently
removed with a cotton swab, whereas cells that had invaded to the
lower surface of the filter were fixed in 70% ethanol for 30 min
and stained with 2% crystal violet for 10 min. Invaded cells were
photographed and quantified by counting them randomly in 5 fields
by a light microscope (Olympus).
Dual-luciferase reporter assay
The 3′UTR of E2F3 containing the potential binding
sites of miR-363 (position 272–278) was amplified by PCR using
human liver cDNA, and inserting into the psiCHECK2 vector (Promega,
Wisconsin, WI, USA) within the XhoI/NotI restriction
sites. A mutant 3′UTR of E2F3 was constructed using QuikChange XL
Site-Directed Mutagenesis kit (Agilent Technologies, Santa Clara,
CA, USA), and cloned into the psiCHECK2 vector. For the luciferase
reporter assay, HepG2 cells were seeded in a 96-well plate. When
reaching 70% confluency, the cells were cotransfected with miR-363
mimic/miR-NC and wild/mutant-type 3′UTR of E2F3. Luciferase
activities were determined 48 h after transfection using the
Dual-Luciferase Reporter Assay System (Promega).
Western blot analysis and
antibodies
Total protein extraction, SDS-PAGE and western blot
analyses were performed as previously described (19). The primary antibodies against E2F3,
GAPDH, E-cadherin, N-cadherin and vimentin were purchased from Cell
Signaling Technology Inc. (Danvers, MA, USA). GAPDH was used as an
internal control for all experiments.
Xenograft mouse model
For in vivo tumorigenesis assays,
2×106 HepG2 cells carrying either miR-363 or miR-NC were
subcutaneously injected into the flanks of female BALB/c nude mice
(6–7 weeks of age) (Laboratory Animal Center of Jilin University).
Tumor volume was determined by measuring tumor length (L) and width
(W) every 7 days using the formula: Tumor volume = 0.5 × (L ×
W2). At 35 days after injection, animals were sacrificed
and subcutaneous tumors were stripped and weighed. All animal
handling and research protocols were approved by the Animal Care
and Use Ethics Committee of Jilin University.
Statistical analysis
In all experiments, the data are expressed as the
mean ± standard error of the mean (SEM) from at least 3 independent
experiments. The differences between groups were analyzed using
Student's t-test when only 2 groups were compared, or using a
one-way ANOVA when >2 groups were compared. Correlations between
miR-363 expression with E2F3 were evaluated by the Pearsons
correlation analysis. A value of P<0.05 was considered as an
indication of statistical significance. All analyses were performed
using SPSS 19.0 software (SPSS, Inc., Chicago, IL, USA).
Results
miR-363 is decreased in HCC tissues
and cell lines
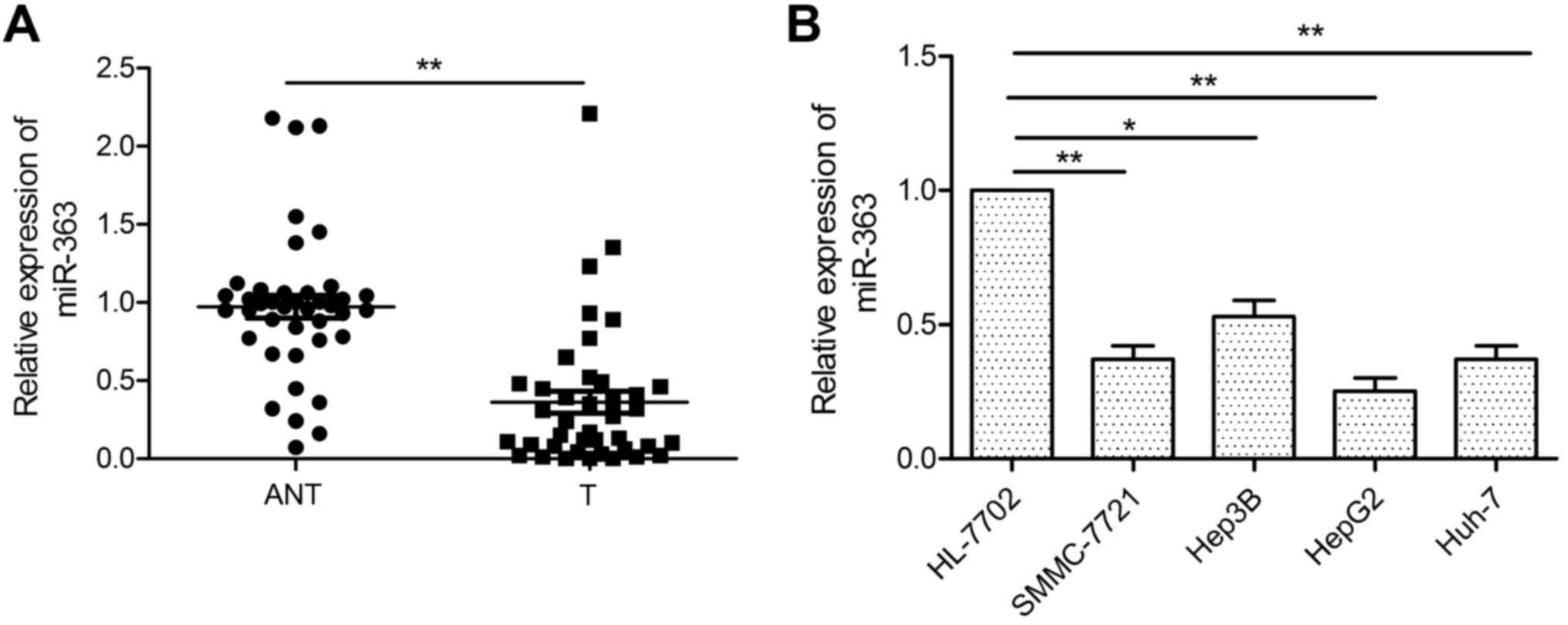
We examined the expression of miR-363 in 40 paired
HCC and corresponding adjacent non-tumor tissues by qRT-PCR. We
found that miR-363 was significantly decreased in the HCC tissues
(Fig. 1A). To assess the
association of miR-363 expression with HCC prognosis, patients were
divided into 2 groups using the mean miR-363 expression (0.33) as
cut-off: one with relative high miR-363 expressions (>0.33), and
another with relative low miR-363 expressions (<0.33). We found
that miR-363 expression was significantly associated with TNM
stage, tumor differentiation and lymph node metastasis (Table I). We next examined the expression
of miR-363 in a series of HCC cell lines. We found that the
expression of miR-363 in 4 human HCC cell lines (SMMC-7721, Hep3B,
HepG2 and Huh-7) was significantly downregulated compared with that
noted in the normal human hepatocyte cell line, HL-7702 (Fig. 1B). These data suggest that miR-363
is involved in the carcinogenesis of HCC.
 | Table I.Association of miR-363 expression
with clinicopathological factors of the 40 HCC patients. |
Table I.
Association of miR-363 expression
with clinicopathological factors of the 40 HCC patients.
|
|
| miR-363
expression |
|
|---|
|
|
|
|
|
|---|
| Variables | No. of cases | Low n (%) | High n (%) | P-value |
|---|
| Age (years) |
|
|
| >0.05 |
|
<55 | 19 | 10 (52.6) | 9 (47.4) |
|
|
≥50 | 21 | 13 (61.9) | 8 (38.1) |
|
| Sex |
|
|
| >0.05 |
|
Male | 23 | 14 (60.8) | 9 (39.2) |
|
|
Female | 17 | 9 (52.9) | 8 (47.1) |
|
| TNM stage |
|
|
| <0.01 |
|
I–II | 29 | 12 (41.4) | 17 (58.6) |
|
|
III–IV | 11 | 11 (100) | 0 (0) |
|
|
Differentiation |
|
|
| <0.01 |
|
Well/moderate | 30 | 14 (46.7) | 16 (53.3) |
|
|
Poor | 10 | 9 (90.0) | 1 (10.0) |
|
| Lymph node
metastasis |
|
|
| <0.01 |
| No | 31 | 14 (45.2) | 17 (54.8) |
|
|
Yes | 9 | 9 (100) | 0 (0) |
|
miR-363 inhibits HCC growth in vitro
and in vivo
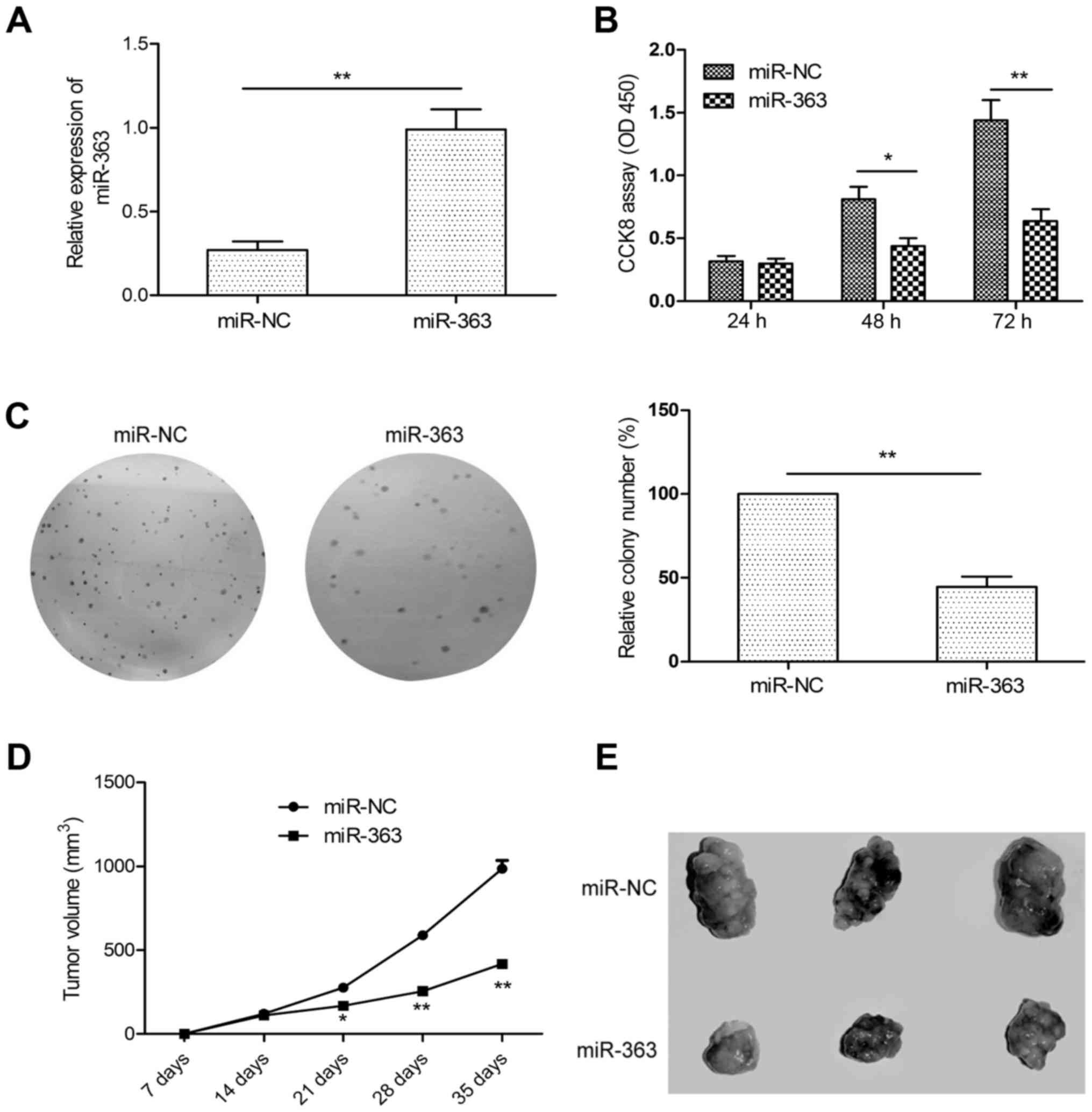
To assess the functional significance of miR-363 in
HCC growth, HepG2 cells with low expression of miR-363 were
transfected with the miR-363 mimic (miR-363) or miR-NC, and then
cell proliferation and colony formation ability were determined.
qRT-PCR results confirmed that miR-363 was upregulated in the HepG2
cells transfected with the miR-363 mimic as compared to cells
transfected with miR-NC (Fig. 2A).
CCK-8 assay revealed that overexpession of miR-363 significantly
suppressed proliferation of the HepG2 cells (Fig. 2B). Consistent with this result,
overexpression of miR-363 also significantly suppressed colony
formation of the HepG2 cells (Fig.
2C). To further confirm whether miR-363 suppresses tumor growth
in vivo, we created tumor xenograft mouse models.
Consistently, miR-363 significantly inhibited tumor growth in
vivo (Fig. 2D). After 35 days
of injection, the mice were sacrificed, and tumor tissues were
stripped. We found that the size of the subcutaneous tumors derived
from the miR-363-expressing HepG2 cells were markedly smaller than
the size of the tumors derived from the miR-NC-transfected cells
(Fig. 2E). Taken together, our data
support a growth inhibitory activity of miR-363 in HCC in
vitro and in vivo.
miR-363 inhibits HCC migration and
invasion, and ETM
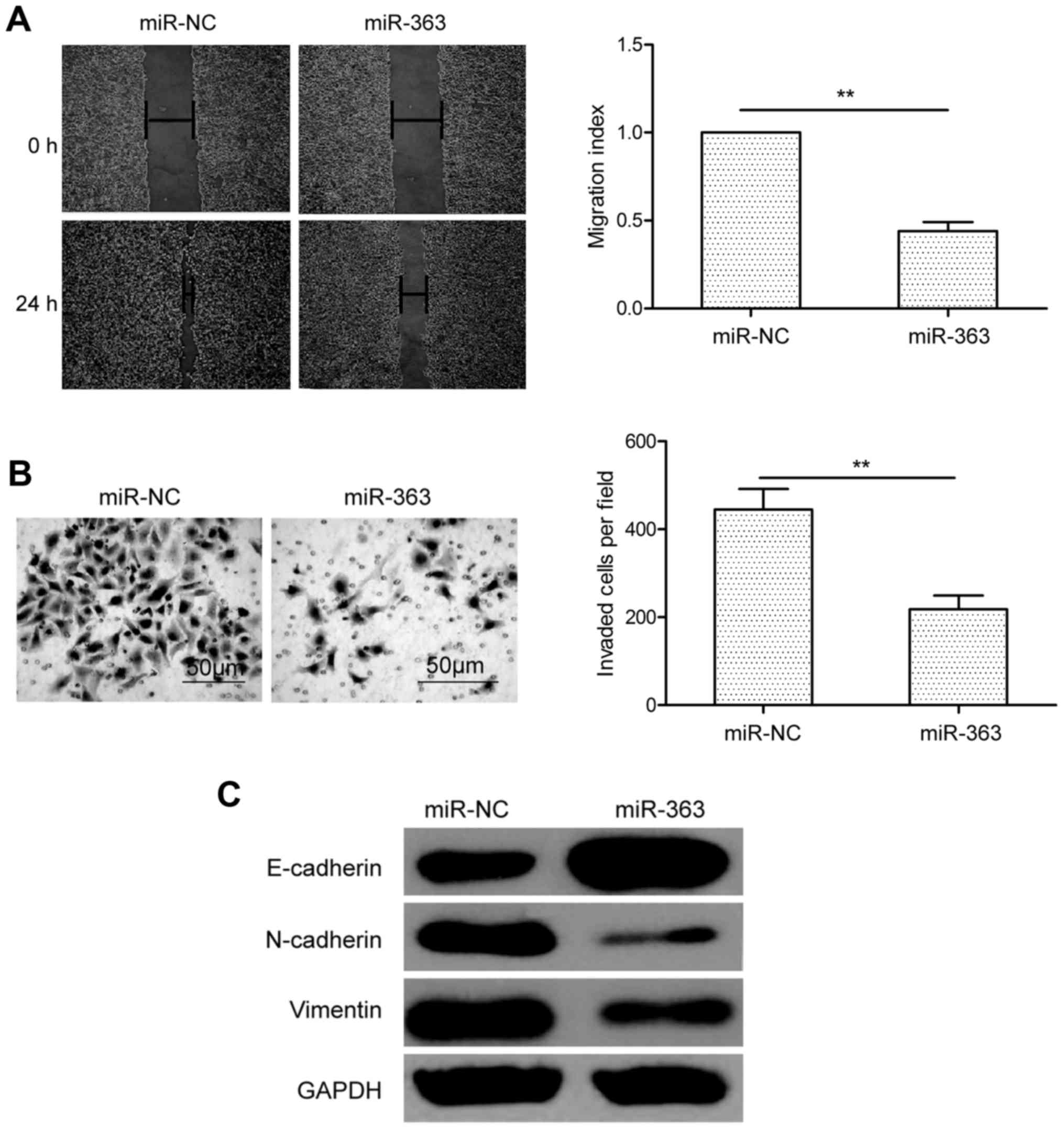
We next studied the effects of miR-363 on cell
migration and invasion in HCC cells using wound healing and
Transwell invasion assays, respectively. Notably, we found that
miR-363 overexpression significantly inhibited the migration and
invasion of HepG2 cells compared to these capacities in the miR-NC
group (Fig. 3A and B). Furthermore,
epithelial-mesenchymal transition (EMT) has been reported to play a
key role in the metastasis of HCC cells (18). To determine whether miR-363 affects
molecular changes typical of EMT in HCC cells, the expression of
mesenchymal markers, including N-cadherin and vimentin and the
epithelial marker, E-cadherin, was determined in the HepG2 cells.
As shown in Fig. 3C, overexpression
of miR-363 resulted in increased E-cadherin expression, and
decreased N-cadherin and vimentin expression. These finding
revealed that miR-363 inhibits HCC metastasis by regulating
EMT.
E2F3 is a candidate target of miR-363
in HCC cells
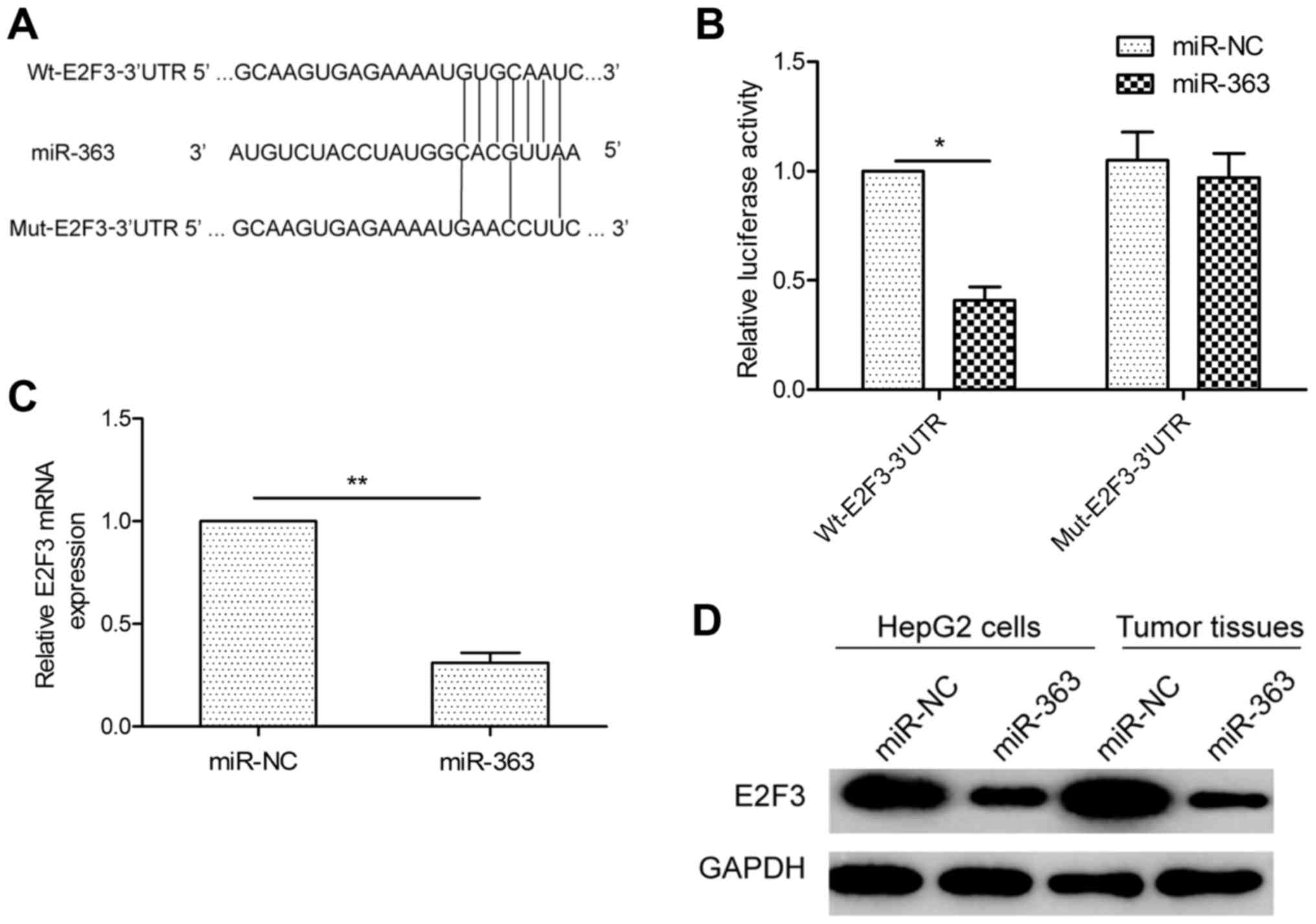
To understand the mechanisms by which miR-363
suppresses HCC growth and invasion, we searched for candidate
targets of miR-363 using 3 bioinformatic databases (TargetScan,
miRanda and PicTar). We identified the 3′UTR of E2F3 that was able
to bind to the ‘seed region’ of miR-363 (Fig. 4A). To confirm that miR-363 may bind
to the 3′UTR of E2F3, a human E2F3 3′UTR fragment containing the
binding sites of miR-363 (Fig. 4A),
or the mutant sites were amplified and inserted into the psiCHECK2
vector, and miR-363 mimic or miR-NC were co-transfected into HepG2
cells and cultured for 48 h, and then luciferase activities were
measured. As predicted, miR-363 was bound to E2F3 3′UTR, resulting
in markedly reduced luciferase activities (Fig. 4B). This effect was specific as
miR-363 failed to suppress the luciferase activity when the binding
site within E2F3 3′UTR was mutated (Fig. 4B). The results indicated that E2F3
is a downstream target of miR-363. Moreover, qRT-PCR and western
blot analysis further demonstrated that overexpression of miR-363
markedly suppressed E2F3 expression at the mRNA and protein levels
(Fig. 4C and D) in HepG2 cells. In
addition, the E2F3 protein level in tumor tissues isolated from
nude mice was determined by western blotting. The expression level
of E2F3 protein in the tumor tissues with miR-363 overexpression
was downregulated compared with the miR-NC group (Fig. 4D). Taken together, these data
indicate that E2F3 is a direct target of miR-363 in HCC cells.
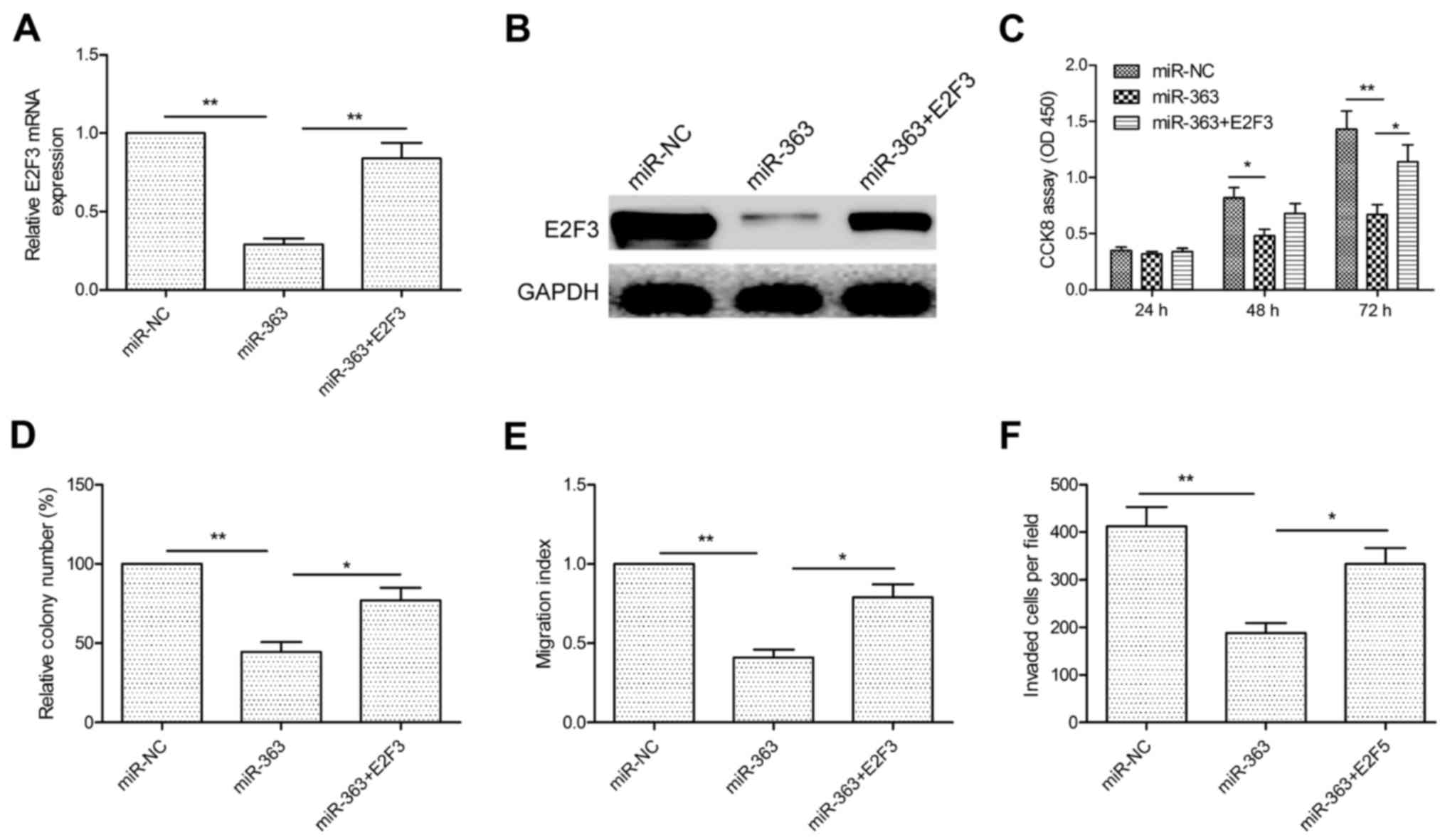
Overexpression of E2F3 reverses the
tumor-suppressive effect of miR-363 in HCC
We further studied whether E2F3 is involved in the
function of miR-363 in HCC cells. HepG2 cells with high expression
of miR-363 were transfected with the E2F3 overexpression plasmid,
and the expression of E2F3 was examined by qRT-PCR and western
blotting. As shown in Fig. 5A and
B, E2F3 expression at the mRNA and protein levels was restored
in the HepG2 cells. Then, we carried out CCK-8, colony formation,
would healing and Transwell assays to evaluate the effect of E2F3
overexpression on cell proliferation, colony formation, migration
and invasion in the HepG2 cells transfected with miR-363
with/without the E2F3 overexpression plasmid. As expected, E2F3
overexpression reversed the suppressive effect on cell
proliferation, colony formation, migration and invasion in HepG2
cells induced by miR-363 overexpression (Fig. 5C-F). Therefore, our data clearly
demonstrated that miR-363 inhibits HCC cell growth and invasion, at
least in part, by targeting E2F3.
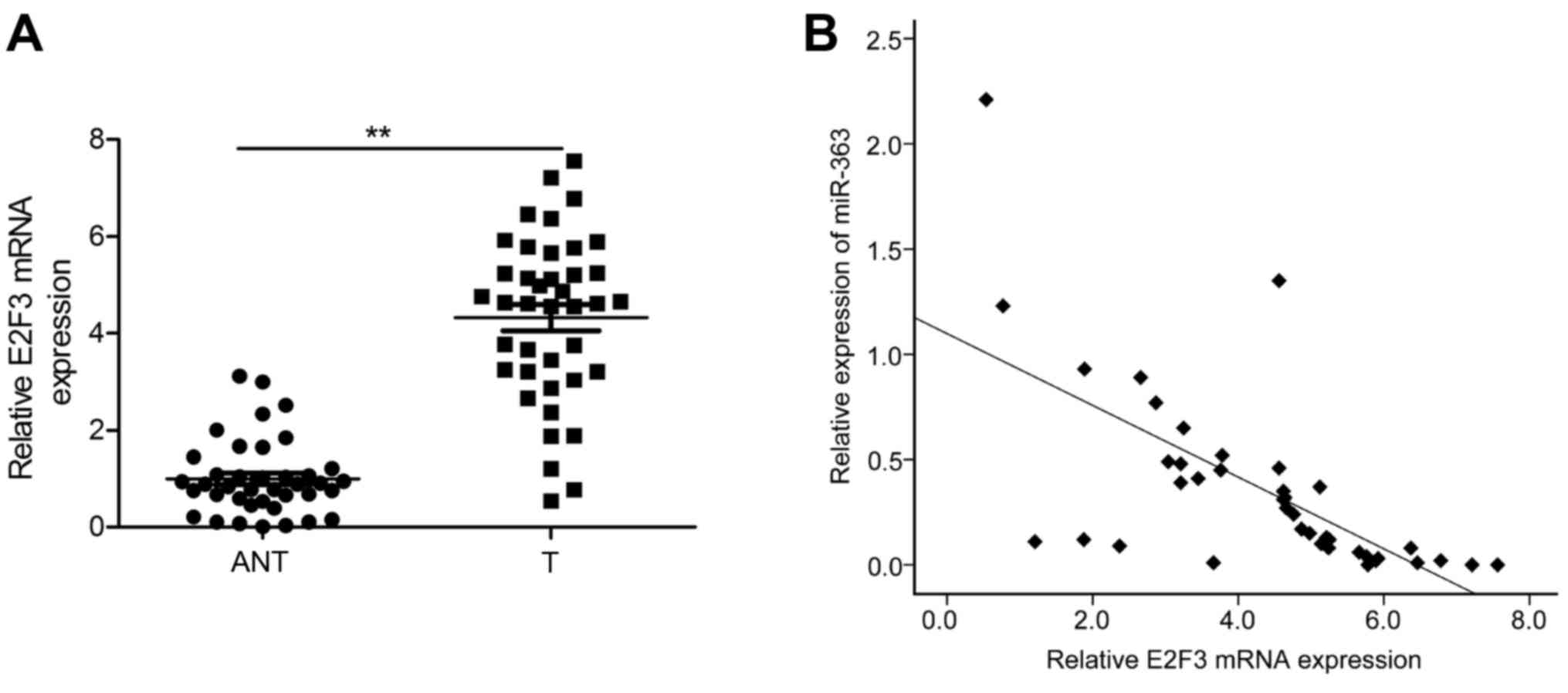
Inverse correlation between E2F3 and
miR-363 expression in HCC patients
We finally examined the E2F3 mRNA expression in 40
pairs of HCC tumors and the corresponding adjacent non-tumor
tissues by qRT-PCR. As shown in Fig.
6A, E2F5 mRNA expression was significantly increased in the HCC
tissues compared to that noted in the adjacent non-tumor tissues.
Pearsons correlation analysis revealed that the expression of
miR-363 was inversely correlated with the E2F3 mRNA level in the
HCC tissues (r=−0.648; P<0.001; Fig.
6B).
Discussion
Emerging evidence suggests that a wide range of
miRNAs play crucial roles in the development and progression of
HCC, and they act as oncogenes or tumor suppressors in HCC
(9,10). For example, miR-449a was found to
function as a tumor-suppressor miRNA by inhibiting cell
proliferation, colony formation, migration and invasion of HCC by
repressing ADAM10 expression (20).
miR-138 inhibited proliferation, colony formation, migration and
invasion in HCC cells by targeting SOX9 (19). miR-133a suppressed proliferation,
colony formation, migration and invasion of HCC cells and caused
cell cycle arrest at the G0/G1 stage and cell apoptosis in
vitro, and decreases tumor size and weight in a nude mouse
HepG2 xenograft model by targeting insulin-like growth factor-1
receptor (IGF-1R) (21). In the
present study, we investigated the miR-363 expression in 40 paired
cases of human HCC tissues and non-tumor tissues and in HCC cell
lines by quantitative RT-PCR. Significantly, we found that miR-363
was decreased in the HCC tissues and cell lines. We also revealed
that miR-363 suppressed HCC cell proliferation, colony formation,
migration and invasion, as well as tumor growth in nude mice by
targeting E2F3. These results imply that miR-363 may play a crucial
role in HCC progression.
Recently, the role of miR-363 in carcinogenesis and
cancer treatment has been a focus of research. Several studies have
demonstrated that miR-363 expression is downregulated in multiple
types of cancers, such as osteosarcoma (22), colorectal (12), head and neck (14), breast (15) and renal cancer (23), and neuroblastoma (24), suggesting that miR-363 functions as
a potential tumor suppressor in these types of cancer. However,
there are also contradictive studies. Chen et al
demonstrated that overexpression of miR-363 in prostate cancer
cells through transfection induced cell proliferation and
positively regulated cell transformation property as well as
promoted EMT in PC-3 cells by regulating c-myc (25). Zhang et al found that miR-363
overexpression promoted gastric cancer cell proliferation and
chemoresistance by directly targeting the tumor suppressor F-box
and WD repeat domain-containing 7 (FBW7) (13). In regards to HCC, the previous study
showed that miR-363 inhibited cell proliferation by downregulating
the expression of S1PR (16), and
decreased cisplatin resistance of HCC cell, partly by targeting
Mcl-1 (17). However, the detailed
roles concerning metastasis and the mechanism of its impact on HCC
cell growth and metastasis of miR-363 in HCC remain unclear. In the
present study, we showed that miR-363 was downregulated in HCC cell
lines and tissues, and was associated with lymph node metastasis
and TNM stage, which was consistent with previous results (16). We also first demonstrated that
miR-336 inhibited cell migration, invasion and EMT in HCC cells, as
well as suppressed HCC tumor growth in vivo by targeting
E2F3. These results further expand the function of miR-363 in HCC
and suggest that miR-363 functions as a tumor suppressor in
HCC.
E2F3, located at 6p22, is a member of the E2F
transcription factor family, which plays a key role in the
regulation of cell proliferation (26). E2F3 has been reported to be involved
in many biological processes, including apoptosis, DNA repair,
differentiation, development and tumorigenesis (27). Increasing evidence suggests that
dysregulation of E2F3 protein contributes to tumor formation by
regulating cell proliferation, cell cycle, apoptosis, migration and
invasion (28–30). Recently a study showed that E2F3
expression was upregulated in HCC tissues, and its expression was
associated with poor prognosis (31), suggesting that E2F3 is an oncogene
in HCC. In addition, E2F3 has been reported to be regulated in HCC
cells by several miRNAs, such as miR-141 (32), miR-144 (33), miR-503 (34), miR-214 (35) miR-424 (18) and miR-217 (36). In the present study, through
luciferase activity, qRT-PCR combined with western blotting, we
showed that E2F3 is a direct target of miR-363 in HCC cells.
Moreover, E2F3 overexpression partially attenuated the
tumor-suppressive effects of miR-363 overexpression in HCC cells,
and the E2F3 mRNA level was negatively correlated with the miR-363
level in HCC tissues. These results suggest that miR-363 exerts its
suppressive role in HCC, at least in part, by targeting E2F3.
In conclusion, the present study demonstrated that
miR-363 is downregulated in HCC cell lines and tissues, and is
associated with tumor differentiation, lymph node metastasis and
TNM stage. In addition, miR-363 suppressed HCC cell proliferation,
colony formation, migration and invasion in vitro, as well
as HCC tumor growth in vivo by directly targeting and
negatively regulating E2F3. This novel miR-363/E2F3 axis may
provide new insight into the mechanisms underlying HCC progression.
These results suggest that miR-363 may be a potential therapeutic
target for the treatment of HCC.
References
|
1
|
Raphael Waly S, Yangde Z and Yuxiang C:
Hepatocellular carcinoma: Focus on different aspects of management.
ISRN Oncol. 2012:4216732012.PubMed/NCBI
|
|
2
|
Mazzola A, Costantino A, Petta S,
Bartolotta TV, Raineri M, Sacco R, Brancatelli G, Cammà C and
Cabibbo G: Recurrence of hepatocellular carcinoma after liver
transplantation: An update. Future Oncol. 11:2923–2936. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Forner A, Llovet JM and Bruix J:
Hepatocellular carcinoma. Lancet. 379:1245–1255. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Orang Valinezhad A, Safaralizadeh R and
Kazemzadeh-Bavili M: Mechanisms of miRNA-mediated gene regulation
from common downregulation to mRNA-specific upregulation. Int J
Genomics. 2014:9706072014.PubMed/NCBI
|
|
5
|
Bushati N and Cohen SM: microRNA
functions. Annu Rev Cell Dev Biol. 23:175–205. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hwang HW and Mendell JT: MicroRNAs in cell
proliferation, cell death, and tumorigenesis. Br J Cancer.
94:776–780. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Calin GA and Croce CM: MicroRNA signatures
in human cancers. Nat Rev Cancer. 6:857–866. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Volinia S, Calin GA, Liu CG, Ambs S,
Cimmino A, Petrocca F, Visone R, Iorio M, Roldo C, Ferracin M, et
al: A microRNA expression signature of human solid tumors defines
cancer gene targets. Proc Natl Acad Sci USA. 103:2257–2261. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Mao B and Wang G: MicroRNAs involved with
hepatocellular carcinoma (Review). Oncol Rep. 34:2811–2820. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Giordano S and Columbano A: MicroRNAs: New
tools for diagnosis, prognosis, and therapy in hepatocellular
carcinoma? Hepatology. 57:840–847. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wang SH, Zhang WJ, Wu XC, Weng MZ, Zhang
MD, Cai Q, Zhou D, Wang JD and Quan ZW: The lncRNA MALAT1 functions
as a competing endogenous RNA to regulate MCL-1 expression by
sponging miR-363-3p in gallbladder cancer. J Cell Mol Med.
20:2299–2308. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Hu F, Min J, Cao X, Liu L, Ge Z, Hu J and
Li X: MiR-363-3p inhibits the epithelial-to-mesenchymal transition
and suppresses metastasis in colorectal cancer by targeting Sox4.
Biochem Biophys Res Commun. 474:35–42. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhang PF, Sheng LL, Wang G, Tian M, Zhu
LY, Zhang R, Zhang J and Zhu JS: miR-363 promotes proliferation and
chemo-resistance of human gastric cancer via targeting of FBW7
ubiquitin ligase expression. Oncotarget. 7:35284–35292.
2016.PubMed/NCBI
|
|
14
|
Chapman BV, Wald AI, Akhtar P, Munko AC,
Xu J, Gibson SP, Grandis JR, Ferris RL and Khan SA: MicroRNA-363
targets myosin 1B to reduce cellular migration in head and neck
cancer. BMC Cancer. 15:8612015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhang R, Li Y, Dong X, Peng L and Nie X:
MiR-363 sensitizes cisplatin-induced apoptosis targeting in Mcl-1
in breast cancer. Med Oncol. 31:3472014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhou P, Huang G, Zhao Y, Zhong D, Xu Z,
Zeng Y, Zhang Y, Li S and He F: MicroRNA-363-mediated
downregulation of S1PR1 suppresses the proliferation of
hepatocellular carcinoma cells. Cell Signal. 26:1347–1354. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ou Y, Zhai D, Wu N and Li X:
Downregulation of miR-363 increases drug resistance in
cisplatin-treated HepG2 by dysregulating Mcl-1. Gene. 572:116–122.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yang H, Zheng W, Shuai X, Chang RM, Yu L,
Fang F and Yang LY: MicroRNA-424 inhibits Akt3/E2F3 axis and tumor
growth in hepatocellular carcinoma. Oncotarget. 6:27736–27750.
2015.PubMed/NCBI
|
|
19
|
Liu Y, Zhang W, Liu K, Liu S, Ji B and
Wang Y: miR-138 suppresses cell proliferation and invasion by
inhibiting SOX9 in hepatocellular carcinoma. Am J Transl Res.
8:2159–2168. 2016.PubMed/NCBI
|
|
20
|
Liu S, Liu K, Zhang W, Wang Y, Jin Z, Jia
B and Liu Y: miR-449a inhibits proliferation and invasion by
regulating ADAM10 in hepatocellular carcinoma. Am J Transl Res.
8:2609–2619. 2016.PubMed/NCBI
|
|
21
|
Zhang W, Liu K, Liu S, Ji B, Wang Y and
Liu Y: MicroRNA-133a functions as a tumor suppressor by targeting
IGF-1R in hepatocellular carcinoma. Tumour Biol. 36:9779–9788.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Li X, Liu X, Fang J, Li H and Chen J:
microRNA-363 plays a tumor suppressive role in osteosarcoma by
directly targeting MAP2K4. Int J Clin Exp Med. 8:20157–20167.
2015.PubMed/NCBI
|
|
23
|
Li Y, Chen D, Li Y, Jin L, Liu J, Su Z, Qi
Z, Shi M, Jiang Z, Ni L, et al: Oncogenic cAMP responsive
element binding protein 1 is overexpressed upon loss of tumor
suppressive miR-10b-5p and miR-363-3p in renal cancer. Oncol Rep.
35:1967–1978. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Qiao J, Lee S, Paul P, Theiss L, Tiao J,
Qiao L, Kong A and Chung DH: miR-335 and miR-363 regulation of
neuroblastoma tumorigenesis and metastasis. Surgery. 154:226–233.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chen Y, Lu X, Wu B, Su Y, Li J and Wang H:
MicroRNA 363 mediated positive regulation of c-myc translation
affect prostate cancer development and progress. Neoplasma.
62:191–198. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Rady B, Chen Y, Vaca P, Wang Q, Wang Y,
Salmon P and Oberholzer J: Overexpression of E2F3 promotes
proliferation of functional human β cells without induction of
apoptosis. Cell Cycle. 12:2691–2702. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ginsberg D: E2F3-a novel repressor of the
ARF/p53 pathway. Dev Cell. 6:742–743. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Gao Y, Li H, Ma X, Fan Y, Ni D, Zhang Y,
Huang Q, Liu K, Li X, Wang L, et al: E2F3 upregulation promotes
tumor malignancy through the transcriptional activation of HIF-2a
in clear cell renal cell carcinoma. Oncotarget. Jul 13–2016.(Epub
ahead of print). doi: 10.18632/oncotarget.10568.
|
|
29
|
Rennhack J and Andrechek E: Conserved E2F
mediated metastasis in mouse models of breast cancer and HER2
positive patients. Oncoscience. 2:867–871. 2015.PubMed/NCBI
|
|
30
|
Trikha P, Sharma N, Pena C, Reyes A, Pécot
T, Khurshid S, Rawahneh M, Moffitt J, Stephens JA, Fernandez SA, et
al: E2f3 in tumor macrophages promotes lung metastasis. Oncogene.
35:3636–3646. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zeng X, Yin F, Liu X, Xu J, Xu Y, Huang J,
Nan Y and Qiu X: Upregulation of E2F transcription factor 3 is
associated with poor prognosis in hepatocellular carcinoma. Oncol
Rep. 31:1139–1146. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Xue J, Niu YF, Huang J, Peng G, Wang LX,
Yang YH and Li YQ: miR-141 suppresses the growth and metastasis of
HCC cells by targeting E2F3. Tumour Biol. 35:12103–12107. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Cao T, Li H, Hu Y, Ma D and Cai X: miR-144
suppresses the proliferation and metastasis of hepatocellular
carcinoma by targeting E2F3. Tumour Biol. 35:10759–10764. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Xiao F, Zhang W, Chen L, Chen F, Xie H,
Xing C, Yu X, Ding S, Chen K, Guo H, et al: MicroRNA-503 inhibits
the G1/S transition by downregulating cyclin D3 and E2F3 in
hepatocellular carcinoma. J Transl Med. 11:1952013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Yang Y, Chang S, Zhao Z, Hou NI, He K,
Wang X, Gao L, Wang L, Cai D, Guo BO, et al: MicroRNA-214
suppresses the proliferation of human hepatocellular carcinoma
cells by targeting E2F3. Oncol Lett. 10:3779–3784. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Su J, Wang Q, Liu Y and Zhong M: miR-217
inhibits invasion of hepatocellular carcinoma cells through direct
suppression of E2F3. Mol Cell Biochem. 392:289–296. 2014.
View Article : Google Scholar : PubMed/NCBI
|