Introduction
Breast cancer is one of the most common malignancies
affecting women worldwide, and is the second leading cause of
cancer-related death among women worldwide (1). Although early detection and extensive
use of adjuvant therapies have greatly reduced patient mortality,
tumor invasion and metastasis remain the most life-threatening
aspects of breast cancer. Metastasis is a multistep process
involving numerous interactions between a tumor and its
microenvironment. Of the multiple biological cascades of
metastasis, epithelial-mesenchymal transition (EMT) is a critical
process associated with increased aggressiveness, and is required
for the initiation of the metastatic spread of tumor cells to
distal tissues (2).
Since purinergic signaling was first proposed,
extracellular adenosine 5-triphosphate (ATP) and the receptors are
emerging as novel and important modulators of inflammation,
immunity and as such potential players in host-tumor interaction.
In addition to its function as the main form of intracellular
energy for all types of cells, ATP has been widely accepted as an
extracellular signaling molecule, which can extracellularly be
released under both physiologic and pathologic conditions (3,4). It
has been demonstrated that a high concentration of extracellular
ATP, in the hundreds micromolar range, is present in tumors,
particularly in the vicinity of necrotic areas (5,6),
whereas it is basically undetectable in normal tissues (4), indicating that ATP acts as a stimulus
in cancer progression. As a ubiquitous extracellular messenger, ATP
functions through purinergic receptors, which are divided into two
distinct families: ligand-gated cation permeable channel P2X
receptors and G protein-coupled P2Y receptors. Currently, 7
subtypes of the P2X family (P2X1-7) and 8 subtypes of the P2Y
family (P2Y1, P2Y2, P2Y4, P2Y6, P2Y11, P2Y12, P2Y13 and P2Y14) have
been cloned and functionally characterized (7).
ATP-induced purinergic signaling has been implicated
in many biological processes such as neurotransmission, muscle
contraction, cell proliferation, differentiation, migration and
apoptosis (7). Different P2Y
receptor subtypes have been identified to be expressed in several
types of cancer in both primary samples of human cancer tissues and
cell lines (8). Our previous
studies revealed that ATP enhanced the in vitro invasion of
prostate cancer cells via P2Y receptors (9,10).
Subsequently, we demonstrated that the P2Y2 receptor is one of the
critical receptors which mediate ATP-promoted invasion and
metastasis of prostate cancer cells both in vitro and in
vivo (11). As one of the G
protein-coupled receptors, the P2Y2 receptor was found to
participate in transactivation of EGFR as well as increased
activity of MAPK and PI3K signaling pathways in various cancer
cells (12–14). Additionally, various studies have
shown that the P2Y2 receptor is overexpressed in colon cancer and
was involved in the metastasis of colorectal cancer cells in a
mouse model (15,16). All of these studies strongly suggest
a critical role of the P2Y2 receptor in cancer progression. In the
present study, we aimed to investigate the function of the P2Y2
receptor in extracellular ATP-regulated migration and invasion, and
the underlying mechanisms in breast cancer progression.
Materials and methods
Chemicals and antibodies
ATP and UTP were purchased from Sigma (St. Louis,
MO, USA) and dissolved in ddH2O to a concentration of
100 mM. The antibodies of P2Y1 (H-120), P2Y2 (H-70), P2Y4 (H-60),
P2Y6 (H-70), Snail (H-130), E-cadherin (G-10) and β-actin were
purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA).
Absolute ethyl alcohol, dimethyl benzene, formaldehyde and hydrogen
peroxide were purchased from Beijing Chemical Works (Beijing,
China).
Breast tissues and clinical
information
Tumor samples were collected from 198 patients who
were diagnosed with breast carcinoma and who underwent modified
radical mastectomy between 2006 and 2010 at Peking University Third
Hospital. Among the 198 patients (median age, 51 years; range,
21–79 years), 148 patients (74.7%) had invasive ductal carcinomas.
Tumor characteristics and lymph node status were retrieved from the
pathology reports and various clinical data were gathered. The
tumor staging was defined according to the American Joint Committee
on Cancer (AJCC) Tumor-Node-Metastasis (TNM) Staging System for
Breast Cancer. Histological grading of tumors was performed
according to the Nottingham (Elston-Ellis) modification of the
Scarff-Bloom-Richardson grading system (17). The Nottingham Prognostic Index (NPI)
was calculated as follows: Lymph node (LN) stage (negative nodes, 1
point; 1–3 positive nodes, 2 points; ≥4 positive nodes, 3 points) +
Grade (1–3) + Maximum diameter (cm) × 0.2. A pevious
study divided the patients into 3 NPI groups: a good prognostic
group with an observed NPI range of 2–3.4, a moderate prognostic
group with an index range of 3.4–5.4, and a poor prognostic group
with an NPI ≥5.4 (18). Detailed
descriptions of the histological assessment, including nodal
status, histological grade and type, NPI grouping, estrogen
receptor (ER) and HER2 amplification status are presented in
Table I.
 | Table I.Statistical analysis of the
expression of the P2Y2 receptor (mean ± SD) and clinicopathological
characteristics of the breast cancer cases. |
Table I.
Statistical analysis of the
expression of the P2Y2 receptor (mean ± SD) and clinicopathological
characteristics of the breast cancer cases.
|
Characteristics | N | P2Y2 | P-value |
|---|
| Grade |
|
| 0.023a |
| 1 | 36 | 6.49±2.58 |
|
| 2 | 122 | 6.26±2.05 |
| 3 | 38 | 4.77±1.70 |
|
| TNM stage |
|
| 0.511b |
|
I+II | 134 | 6.11±2.24 |
|
|
III+IV | 63 | 5.80±2.92 |
|
| Lymph node
metastasis |
|
| 0.407b |
|
Yes | 116 | 6.17±2.03 |
|
| No | 81 | 5.79±2.29 |
|
| Distant
metastasis |
|
| 0.360b |
|
Yes | 28 | 6.53±2.70 |
|
| No | 170 | 5.93±2.20 |
|
| Location in the
tumor |
|
| 0.020b |
| Tumor
core | 92 | 1.99±1.18 |
|
|
Invasive edge or cancer
embolus | 92 | 2.48±1.63 |
|
|
NPI |
|
| 0.141a |
|
Good | 28 | 6.30±2.42 |
|
|
Moderate | 115 | 6.28±2.18 |
|
|
Poor | 52 | 5.28±2.82 |
|
| ER expression |
|
| 0.755b |
|
Yes | 83 | 5.94±1.27 |
|
| No | 104 | 6.09±2.08 |
|
| HER2
amplification |
|
| 0.008a |
|
Yes | 76 | 5.27±2.03 |
|
| No | 114 | 6.47±3.08 |
|
Cell lines and culture conditions
The MCF7 and Hs578T human breast cancer cell lines
were purchased from the American Type Culture Collection (ATCC;
Manassas, VA, USA). The MCF7 breast cancer cells were cultured in
RPMI-1640 medium (Gibco, Grand Island, NY, USA) supplemented with
10% fetal bovine serum (FBS). The Hs578T cells were cultured in
Dulbecco's modified Eagle's medium (DMEM) (Gibco) containing 10%
FBS. The cells were grown in a humidified atmosphere containing 5%
CO2 at 37°C.
Reverse transcription and real-time
PCR
Total RNA was extracted using TRIzol reagent
(Invitrogen, Carlsbad, CA, USA), and 2 µg of total RNA was
reversely transcribed into cDNA using Moloney Murine Leukemia Virus
(MMLV) reverse transcriptase (Promega, Madison, WI, USA), following
the manufacturer's instructions. Real-time PCR was carried out
using ABI StepOne Real-Time PCR System (Applied Biosystems, Foster
City, CA, USA). Due to no introns in the subtypes of P2Y receptors,
DNase (Promega) was used to totally eliminate DNA contamination in
the total RNA before reverse transcription. The expression of the
examined genes was normalized by β-actin. The specific primers were
synthesized and purified by Invitrogen and are listed in Table II. The thermal cycle conditions
were as follows: 10 min at 95°C, 30 cycles of 15 s at 95°C and 1
min at 60°C. The 2−ΔΔCt method was used for relative
quantification as previously described (19).
| Table II.Sequences of the primers used in the
real-time qPCR experiments. |
Table II.
Sequences of the primers used in the
real-time qPCR experiments.
| Target | Forward primer
sequence (5′-3′) | Reverse primer
sequence (5′-3′) | Length (bp) |
|---|
| E-cadherin |
CTGGGCTGGACCGAGAGA |
GAAGGTCAGCAGCTTGAACCA | 60 |
| Claudin-1 |
TGAAGTGCTTGGAAGACGATG |
GGCAACTAAAATAGCCAGACCT | 95 |
| IL-8 |
ACTGAGAGTGATTGAGAGTGGAC |
AACCCTCTGCACCCAGTTTTC | 112 |
| Snail |
AATCGGAAGCCTAACTACAGCG |
GTCCCAGATGAGCATTGGCA | 147 |
| P2Y1 |
GTGCTGGTGTGGCTCATTGT |
TGGTGTCGTAACAGGTGATGGT | 100 |
| P2Y2 |
GCTACAGGTGCCGCTTCAAC |
AGACACAGCCAGGTGGAACAT | 170 |
| P2Y4 |
GTGAGGTGGAGCTGGACTGTTG |
GGTCGGAGGCGGAAGATGA | 136 |
| P2Y6 |
CCACAGGCATCCAGCGTAAC |
AGGAAGCCGATGACAGTGAGAG | 106 |
| P2Y11 |
CCTTCTGTGTCCACCCTCTACTCT |
AGTGCTCTTGGCGTCCTCTG | 113 |
| P2Y12 |
CACTGCTCTACACTGTCCTGT |
AGTGGTCCTGTTCCCAGTTTG | 190 |
| P2Y13 |
CACTGCTCTACACTGTCCTGT |
AGTGGTCCTGTTCCCAGTTTG | 134 |
| P2Y14 |
TGGAGTGTCAGGATGGATATTCT |
GTCACCAAGGATCTTGAAAGGAA | 124 |
| β-actin |
GGATGCAGAAGGAGATCACTG |
CGATCCACACGGAGTACTTG | 90 |
Cell lysis and western blot
analysis
Cell extracts were generated using RIPA buffer
(Applygen Technologies Inc., Beijing, China) supplemented with
protease inhibitors (Roche, Mannheim, Germany). The concentration
of protein was determined using a BCA reagent (Applygen
Technologies Inc.). Equal amounts of protein were separated by
sodium dodecyl sulfate-polyacrylamide gel electrophoresis
(SDS-PAGE) gel and transferred onto nitrocellulose membranes
(Bio-Rad, Hercules, CA, USA), which were incubated separately with
primary antibodies against the P2Y2 receptor (1:500), E-cadherin
(1:500), claudin-1 (1:1,000), Snail (1:1,000) and β-actin (1:1,000)
at 4°C overnight. Immunoreactive proteins were visualized by
chemiluminescence (Applygen Technologies Inc.) and quantified by
densitometric analysis using Quantity One software (Bio-Rad).
siRNA and cell transfection
To investigate the role of the P2Y2 receptor in ATP-
and UTP-driven migration and invasion, two different small
interfering RNAs (siRNAs) (siRNA1 and siRNA2) were used to
downregulate the expression of the P2Y2 receptor in the breast
cancer cells. MCF7 and Hs578T breast cancer cells were transfected
with siRNAs directed against P2Y2 receptor (siRNA1 and siRNA2) gene
expression or a scramble sequence not targeting any known gene as a
control (NC). Two distinct siRNA oligonucleotides targeting the
P2Y2 receptor were used with sequences as follows: P2Y2 siRNA1,
5′-GUGCUAACAGUUGCCUUGA-3′ and P2Y2 siRNA2,
5′-GCCCAAGAGAUGAACAUCU-3′. A scramble siRNA was used as the
negative control (NC). Cells were transfected with siRNA using
Lipofectamine 2000 (Invitrogen) according to the manufacturer's
instructions.
In vitro cell migration and invasion
assays
The migration and invasion abilities of the breast
cancer cells were evaluated using Boyden Chamber assay according to
the method described by Albini et al (20), with some modifications. Cell
migration capacity was analyzed using 24-well Transwell chambers
which contained 8-µm pore size polyethylene terephtalate membrane
cell culture inserts. The upper compartment was seeded with
0.5×105 viable cells and the lower compartment was
filled with 600 µl NIH3T3 conditioned medium as a chemoattractant.
After incubation with or without 100 µM ATP for 12 h at 37°C in a
humidified atmosphere containing 5% CO2, the chambers
were removed. Cells on the upper side of the chamber were removed
with cotton-tipped swabs, and the cells on the lower surface of the
membrane were fixed and stained with crystal violet. The number of
migrated cells was counted under a light microscope at a
magnification of ×200. The average numbers of migrated cells were
determined from 7 representative fields. Cell invasive ability was
assessed using the same inserts as mentioned above, but with the
membrane covered with a film of Matrigel (BD Biosciences, Franklin
Lakes, NJ, USA). In this case, 1×105 viable cells were
seeded in the upper compartment. Cells on the lower surface of the
membranes were stained with crystal violet and observed under a
microscope at a magnification of ×200. The numbers of invaded cells
in 7 fields were counted and the mean for each chamber was
determined. Each experiment was repeated at least 3 times, and the
results for migration and invasion were normalized to the
controls.
Immunohistochemical staining
To further verify the role of the P2Y2 receptor in
the invasion of breast cancer, immunohistochemical staining was
carried out to detect the expression of P2Y receptors and EMT
protein markers in breast cancer tissue specimens with the
avidin-biotin complex technique. Briefly, the slides were washed in
xylene to remove the paraffin, rehydrated through serial dilutions
of alcohol, followed by washings with a solution of
phosphate-buffered saline (PBS) (pH 7.2). Treated slides were then
placed in a citrate buffer (pH 6.0) and heated in a pressure cooker
for 3 min. The slides were then incubated overnight at 4°C with a
series of anti-P2YR rabbit polyclonal antibodies including
anti-P2Y1 (1:200 dilution), anti-P2Y2 (1:300 dilution), anti-P2Y4
(1:200 dilution) and anti-P2Y6 (1:400 dilution) antibodies, as well
as anti-Snail rabbit polyclonal (1:200 dilution) and
anti-E-cadherin mouse monoclonal antibodies (1:200 dilution),
respectively. The Histostain-Plus Streptavidin-Peroxidase method
(ZYMED kit 50–420Z; Life Technologies, Frederick, MD, USA) was
performed for signal development and the cells were counterstained
with hematoxylin. NCs were obtained by excluding the primary
antibody. Slides were mounted with gum for examination and images
were captured using an Olympus BX51 microscope (Olympus, Tokyo,
Japan) and Nikon Digital Camera System for study comparison (Nikon,
Tokyo, Japan).
Scoring of immunohistochemical
stainings
We interpreted the cytoplasmic and/or nuclear
staining as positive staining results. The German semi-quantitative
scoring system, which has been widely accepted and used in previous
studies (21), was also adopted to
consider the staining intensity and area extent. Every tumor was
scored according to the intensity of the nuclear or cytoplasmic
staining (no staining, 0; weak staining, 1; moderate staining, 2;
strong staining, 3) and the extent of stained cells (0%, 0; 1–10%,
1; 11–50%, 2; 51–80%, 3; 81–100%, 4). The final immunoreactive
score was determined by multiplying the intensity and extent
scores, with a minimum score of 0 and a maximum score of 12. For
each tumor, we also scored the staining intensity of invasive edge
areas and the tumor core areas.
Data analysis and statistics
All experiments, except immunohistochemistry, were
repeated at least 3 times. Statistical analyses were performed
using SPSS software (version 18.0; SPSS, Inc., Chicago, IL, USA).
Student's t-test was performed to determine whether there was a
significant difference between 2 groups, and one-way ANOVA analysis
was performed for comparison of multiple means. In addition, the
non-parametric Spearman's rho correlation coefficient was used to
analyze associations of the P2Y2 receptor expression with different
clinicopathological characteristics. Statistical significance is
indicated at P<0.05.
Ethics statement
The tissue samples used in the present study were
archived paraffin tissue blocks from the Department of Pathology,
Peking University Third Hospital. Many of the breast cancer
patients operated on before 2011 were lost to follow-up in our
hospital. Written or verbal consent could not be obtained from
these patients when they were integrated into the present study as
an essential part. We described this circumstance to the Peking
University Ethics Committee. The Ethics Committees/IRB approved
that this group could be exempt from informed consent. The
data/samples were used anonymously. The present study was approved
by the Peking University IRB (reference no. IRB00001052-11029).
Results
P2Y2 receptor is significantly
expressed in breast cancer cells
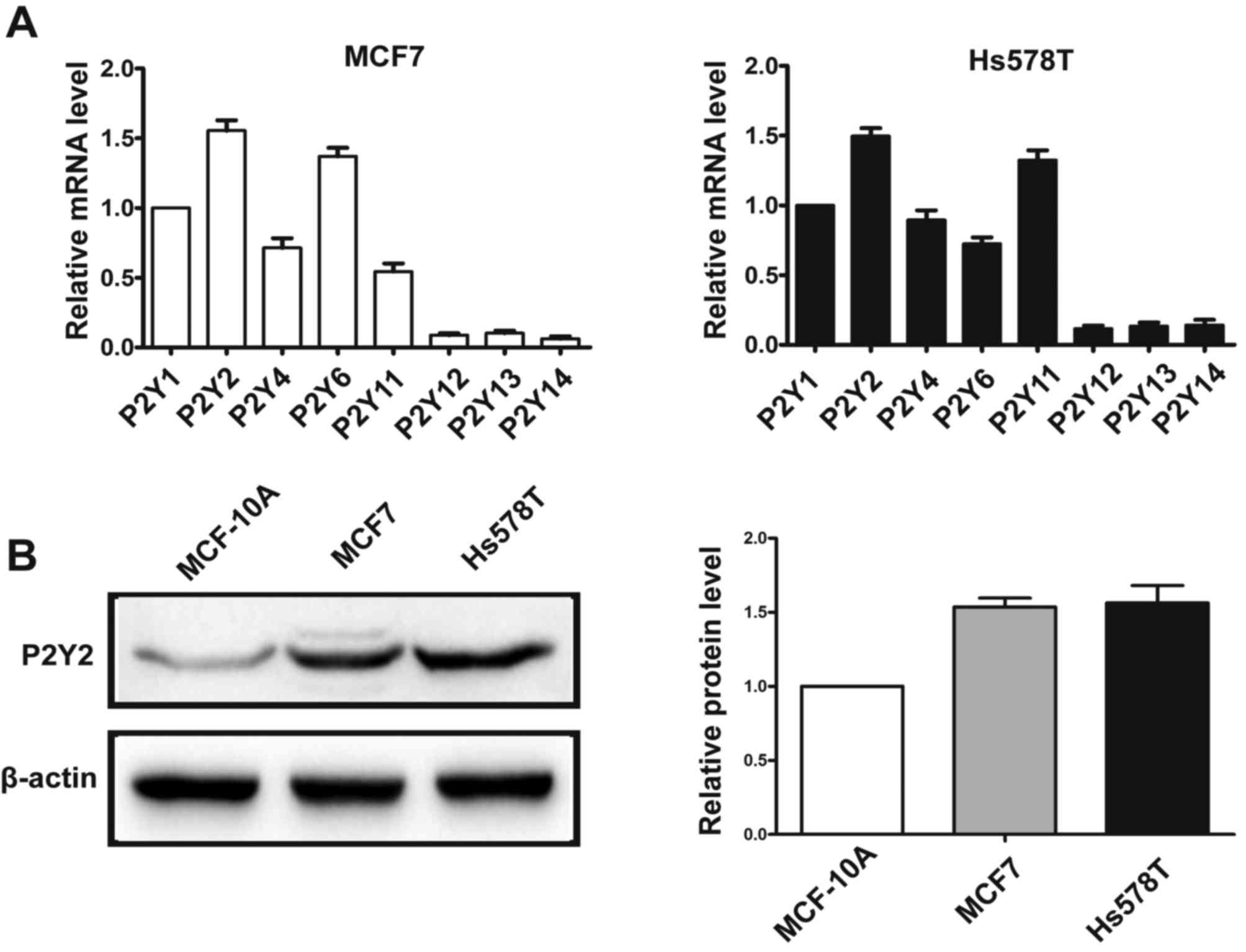
Real-time PCR analysis showed that MCF7 and Hs578T
breast cancer cells predominantly expressed mRNA for P2Y1, P2Y2,
P2Y4, P2Y6 and P2Y11 receptors (Fig.
1A). Of all the P2Y subtypes, the P2Y2 receptor exhibited the
highest expression in both breast cancer cell lines. In addition,
the protein expression of P2Y2 was examined by western blotting in
breast cancer cells and immortal normal mammary cells MCF-10A. We
found that P2Y2 protein was highly expressed in the MCF7 and Hs578T
breast cancer cell lines compared with the MCF-10A mammary
epithelial cells (Fig. 1B).
P2Y2 receptor is involved in ATP- or
UTP-promoted migration and invasion of breast cancer cells
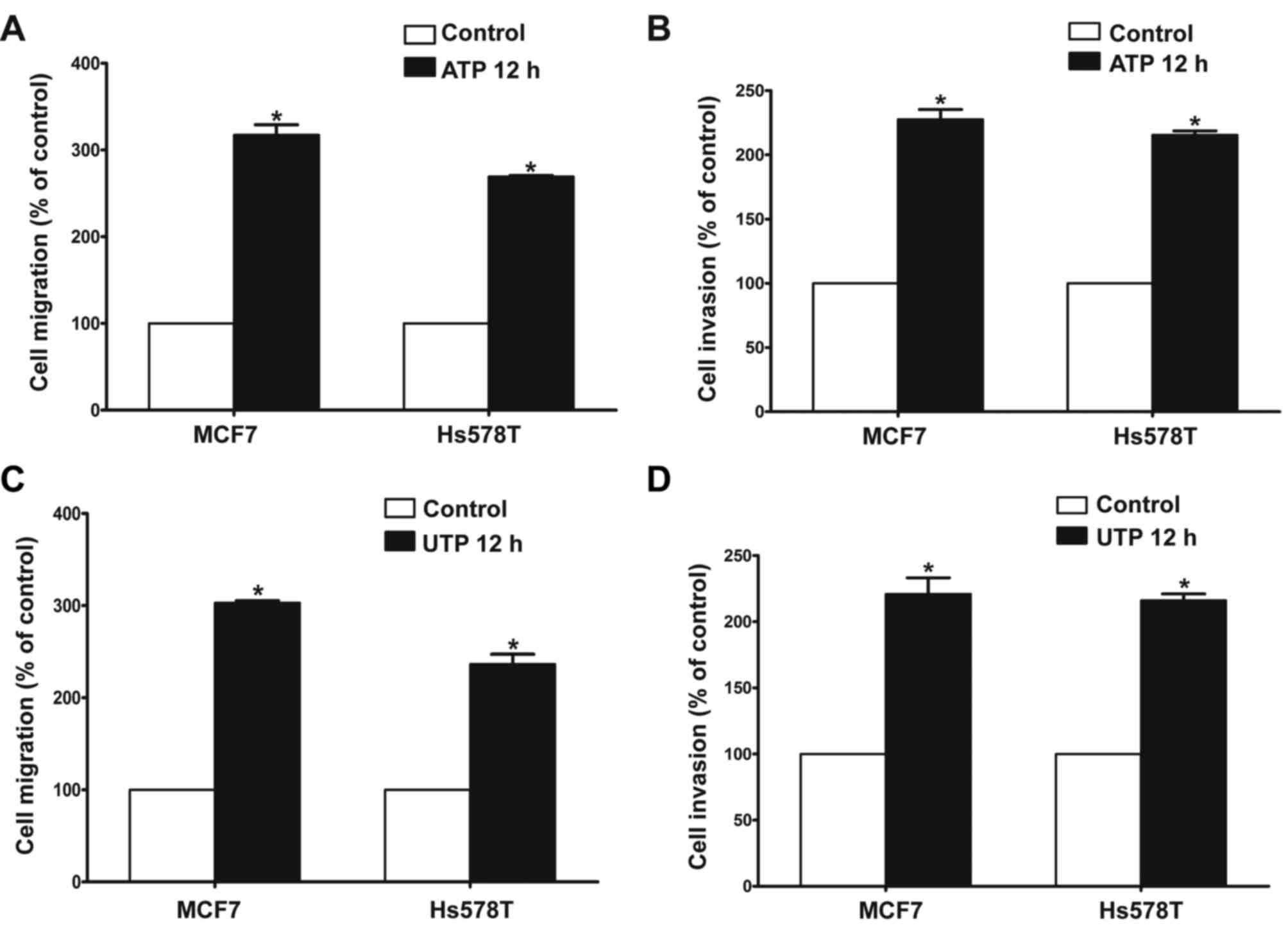
The Transwell assay revealed that ATP (100 µM)
promoted the migration and invasion of MCF7 and Hs578T breast
cancer cells. UTP (100 µM), as another preferred ligand of the P2Y2
receptor, also caused marked enhancement of migration and invasion
in the breast cancer cells, which was consistent with the effect of
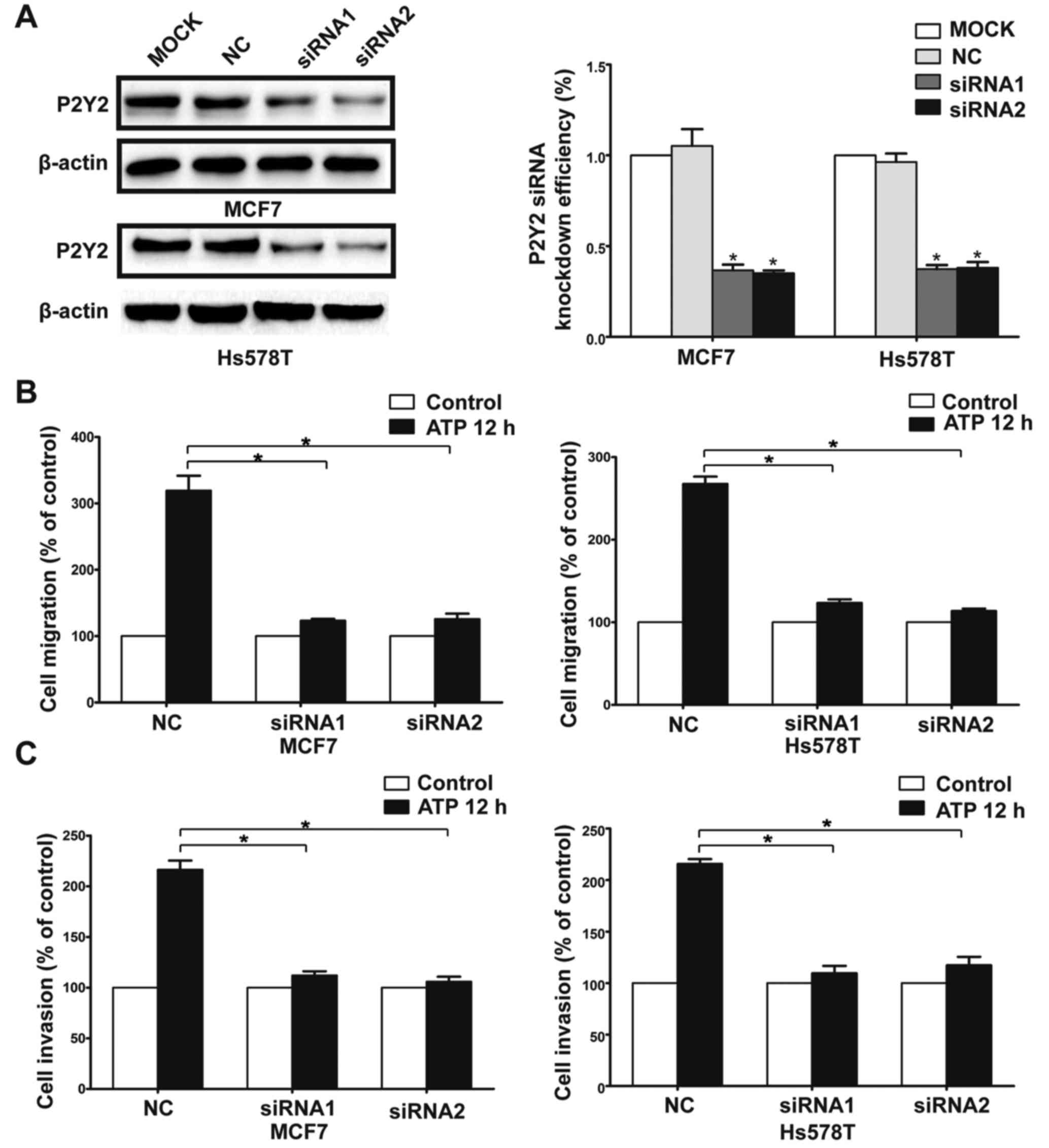
ATP stimulus (Fig. 2A-D). Then, we
used siRNAs (siRNA1 and siRNA2) to silence the expression of the
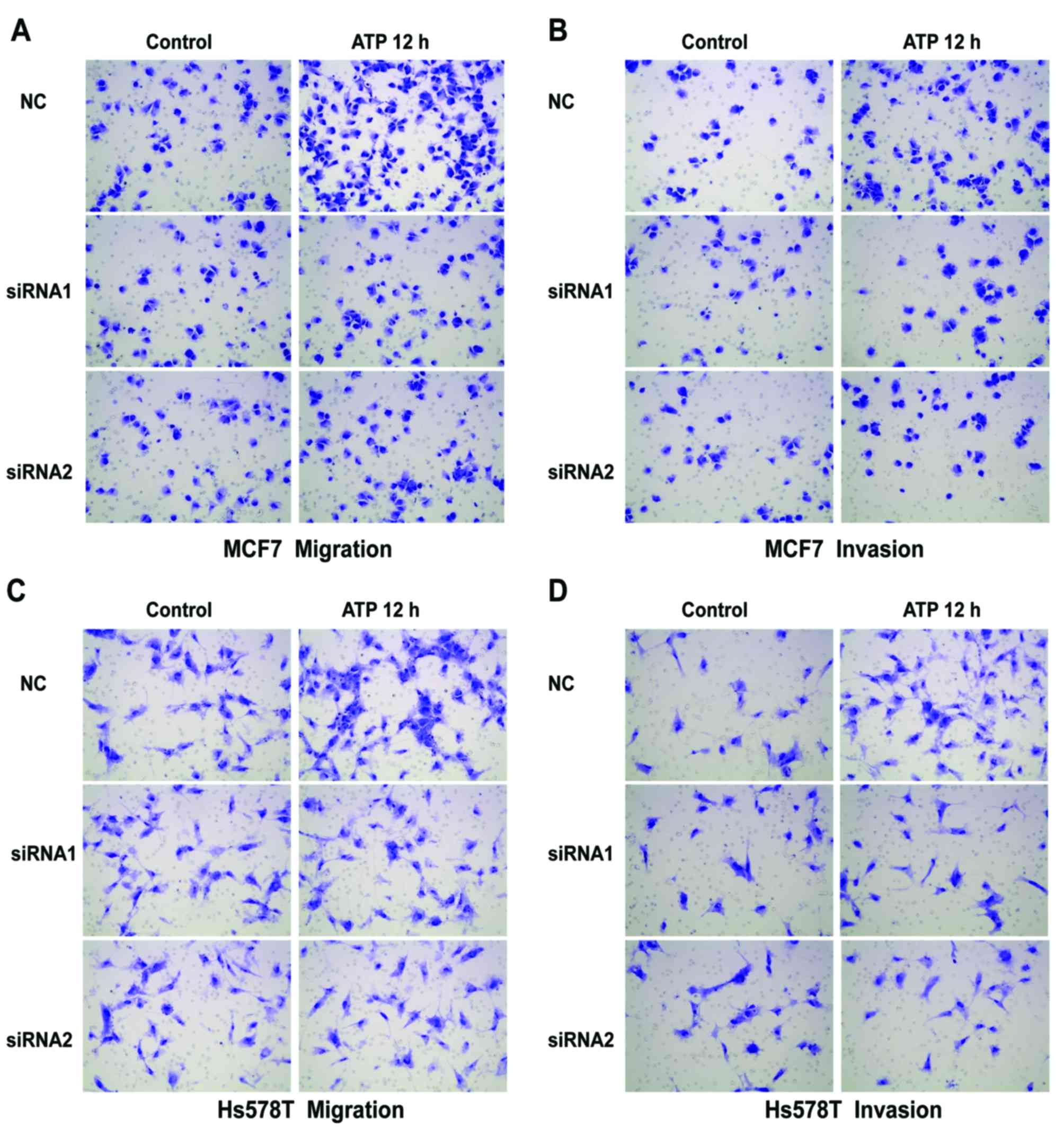
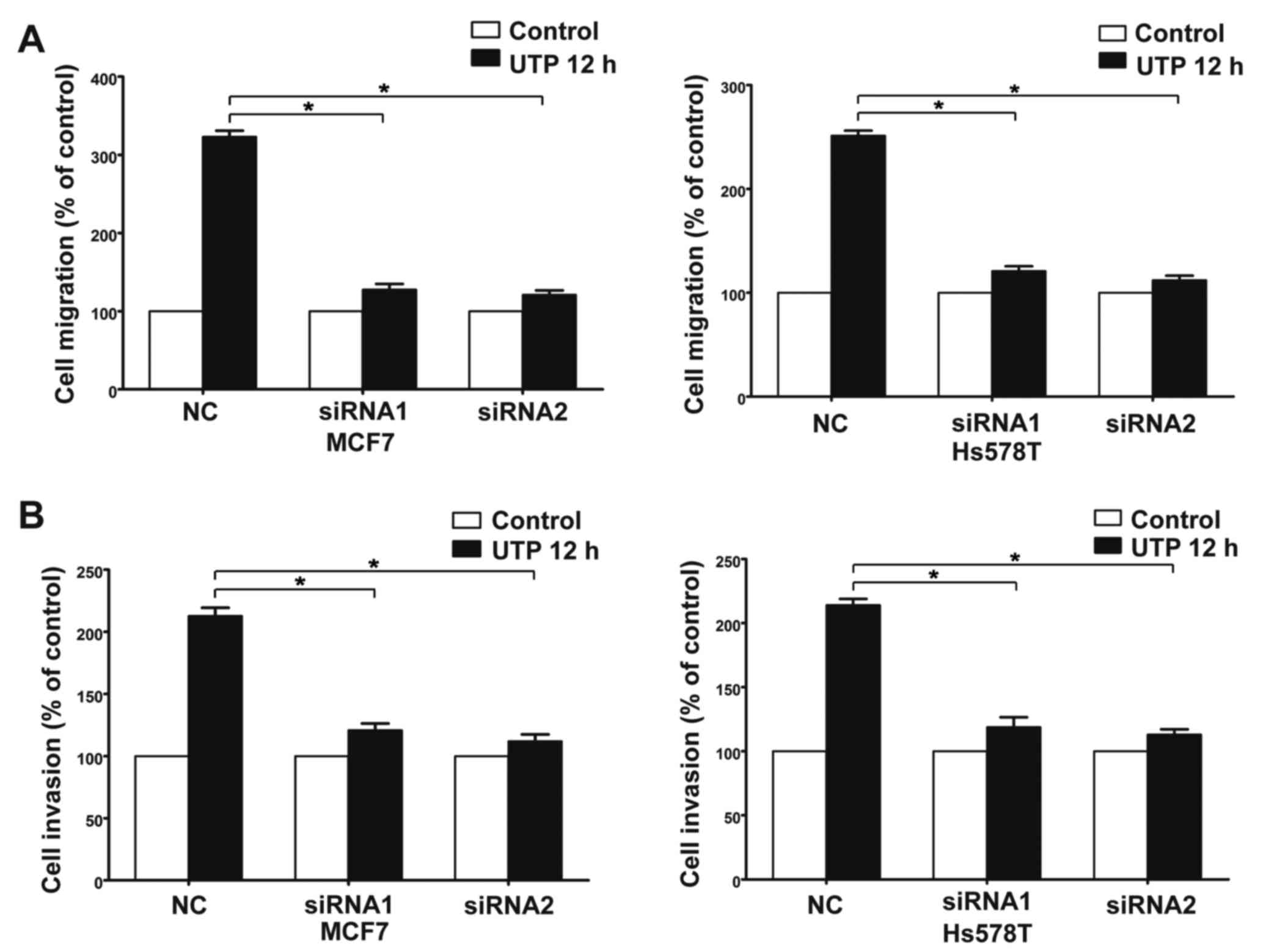
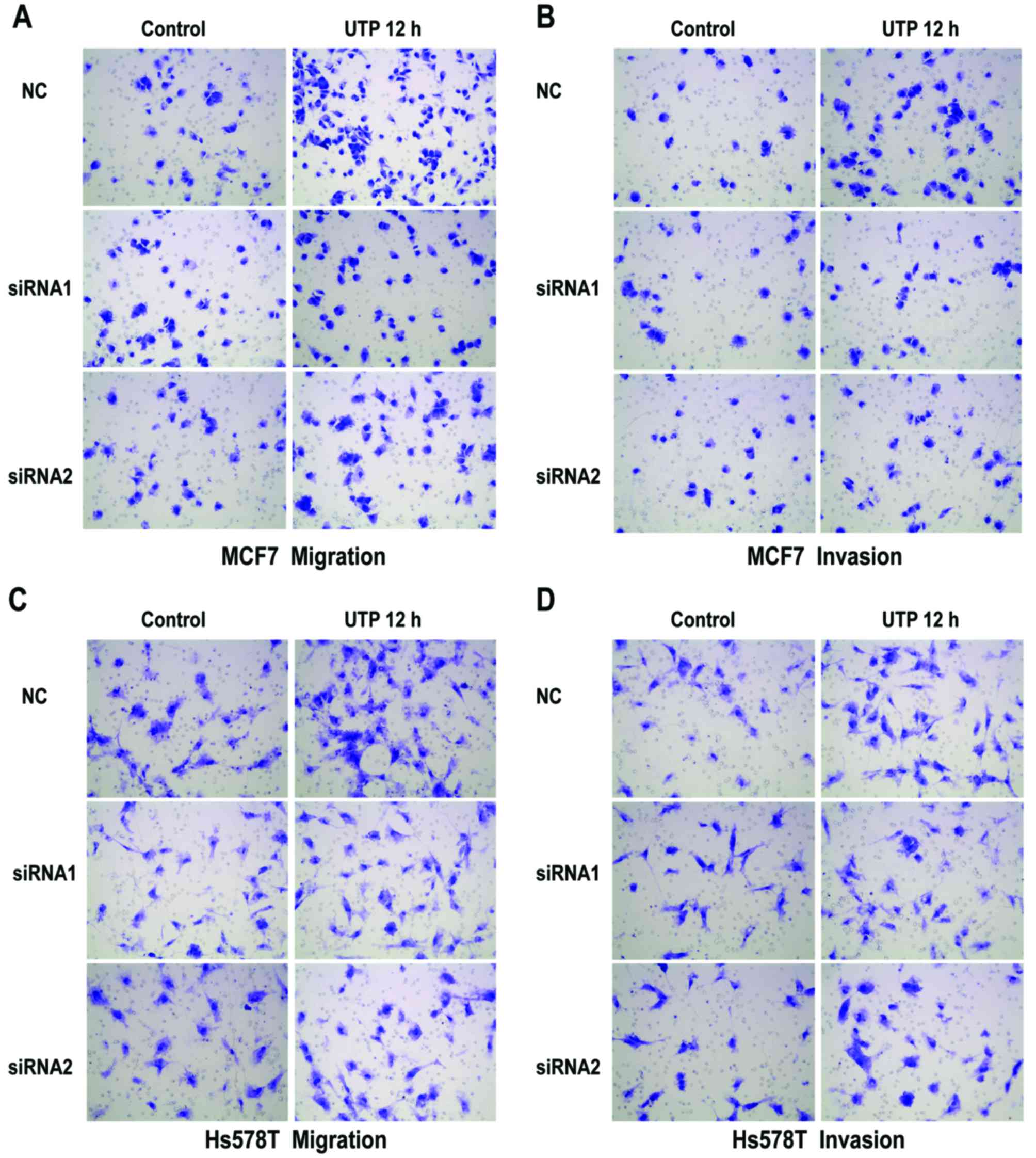
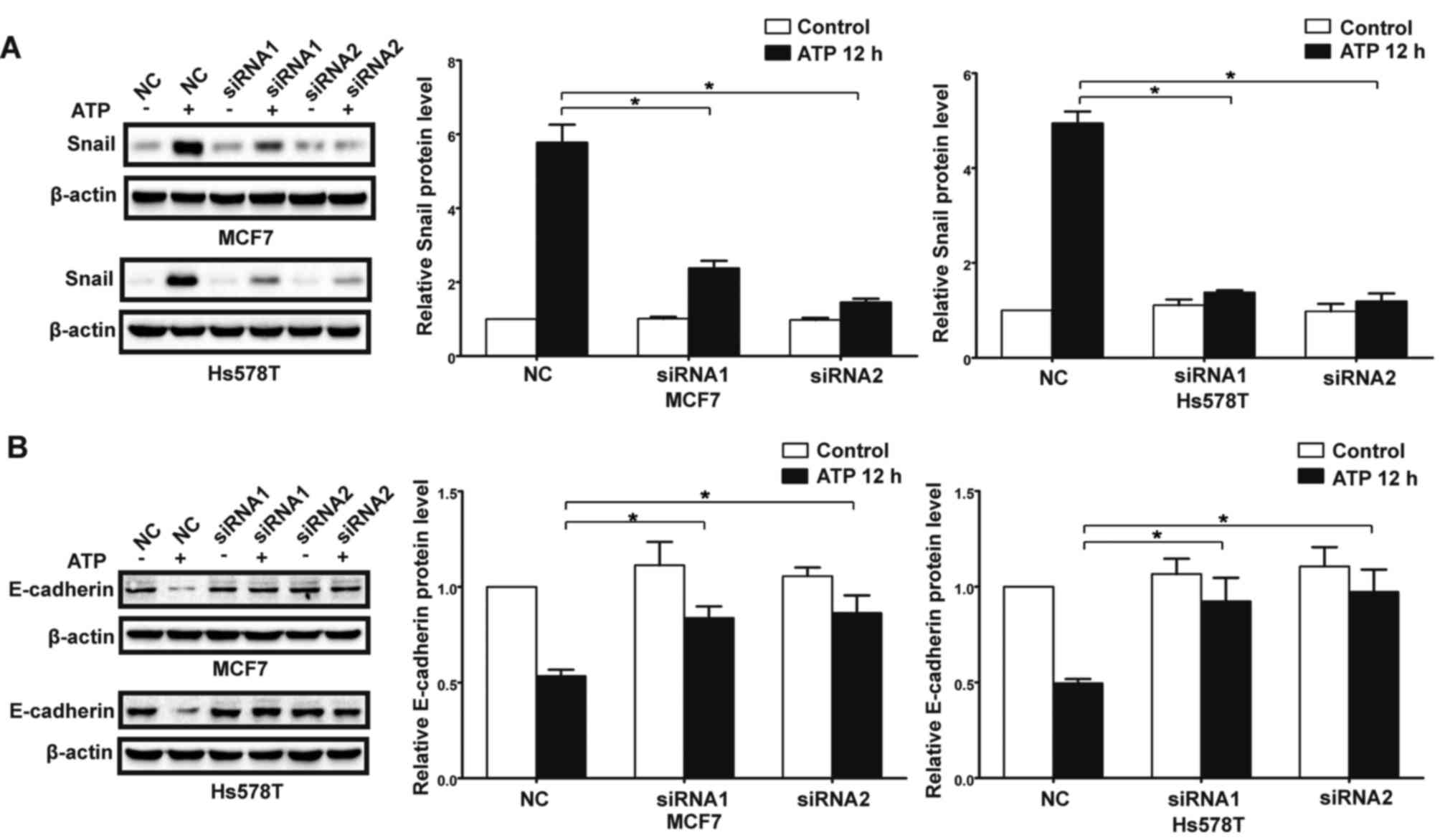
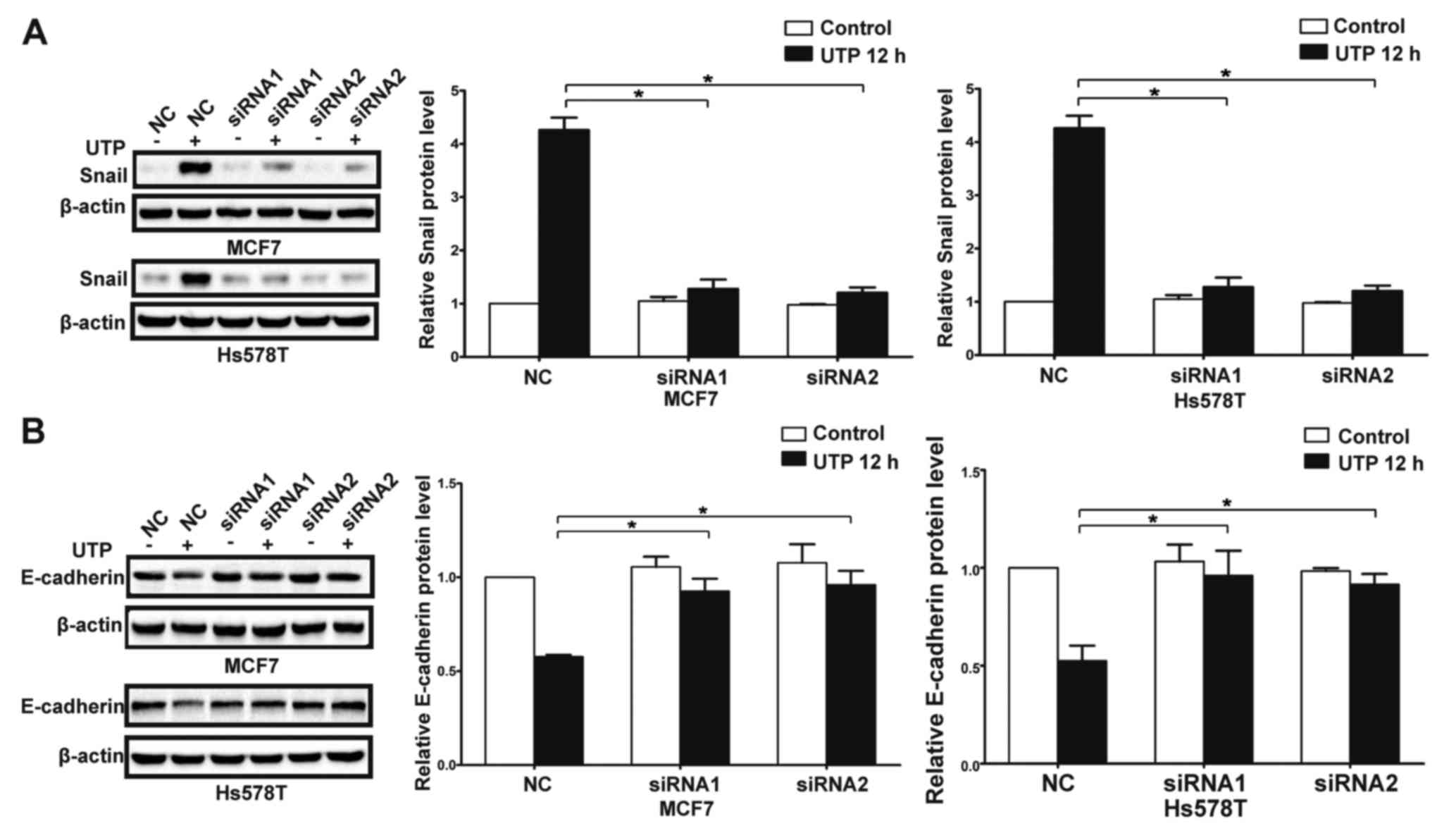
P2Y2 receptor in the MCF7 and Hs578T cells (Fig. 3A). Compared with the control cells,
ATP-driven migration and invasion were markedly inhibited after
downregulation of the P2Y2 receptor (Figs. 3B and C, and 4A-D). Similarly, knockdown of the P2Y2
receptor also significantly blocked UTP-mediated migration and
invasion of MCF7 and Hs578T cells (Figs. 5A and B, and 6A-D). All the above data indicate that the
P2Y2 receptor is involved in the regulation of migration and
invasion of breast cancer cells.
P2Y2 receptor mediates ATP or
UTP-driven expression of EMT-related genes Snail and
E-cadherin
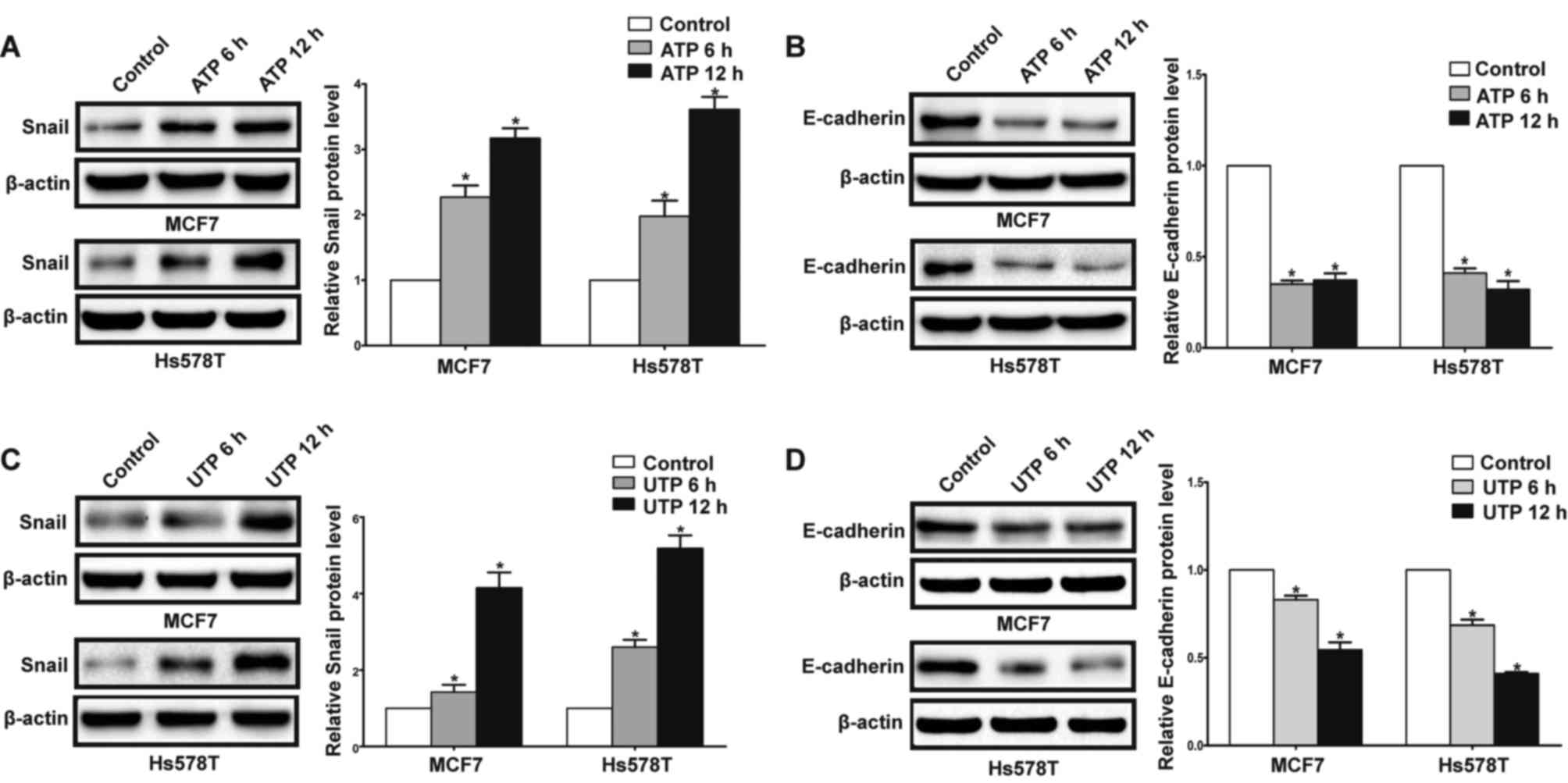
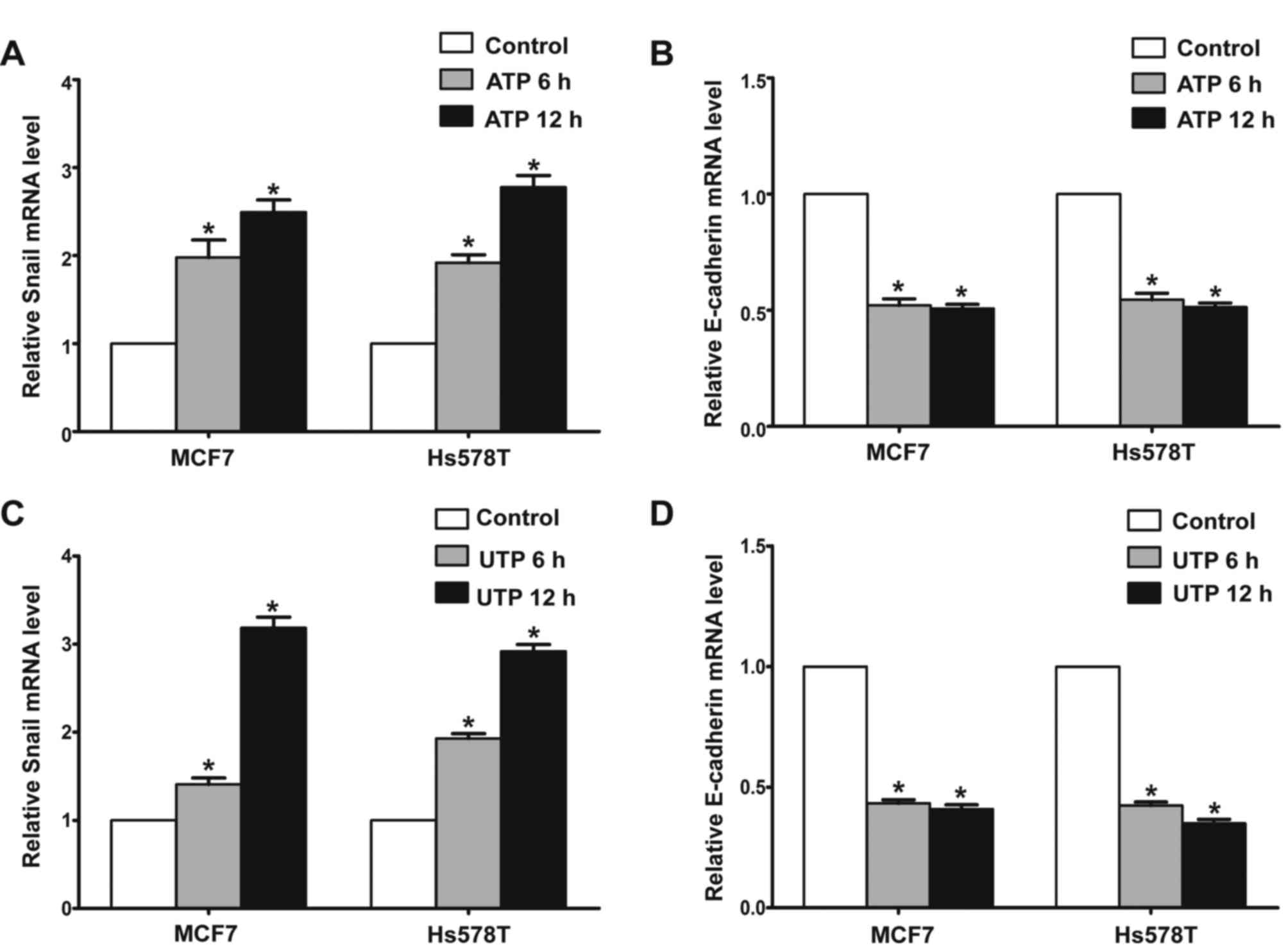
In the presence of ATP, expression of Snail was
significantly increased in the MCF7 and Hs578T cells, while
expression levels of E-cadherin and claudin-1 were markedly reduced
at both the mRNA and protein levels. Meanwhile, UTP exerted similar
effects on the regulation of Snail and E-cadherin in the MCF7 and
Hs578T cells (Figs. 7A-D, and
8A-D). Furthermore, after
downregulation of the P2Y2 receptor, the ATP-mediated increase in
Snail expression and decrease in E-cadherin expression (Fig. 9A and B) were significantly
attenuated. Similarly, knockdown of the P2Y2 receptor significantly
inhibited UTP-mediated expression changes in EMT-related genes in
the MCF7 and Hs578T cells (Fig. 10A
and B). These data indicate that the P2Y2 receptor mediates the
ATP- or UTP-induced expression changes of EMT-related genes in
breast cancer cells.
Expression of P2Y2, Snail and
E-cadherin in invasive breast cancer specimens
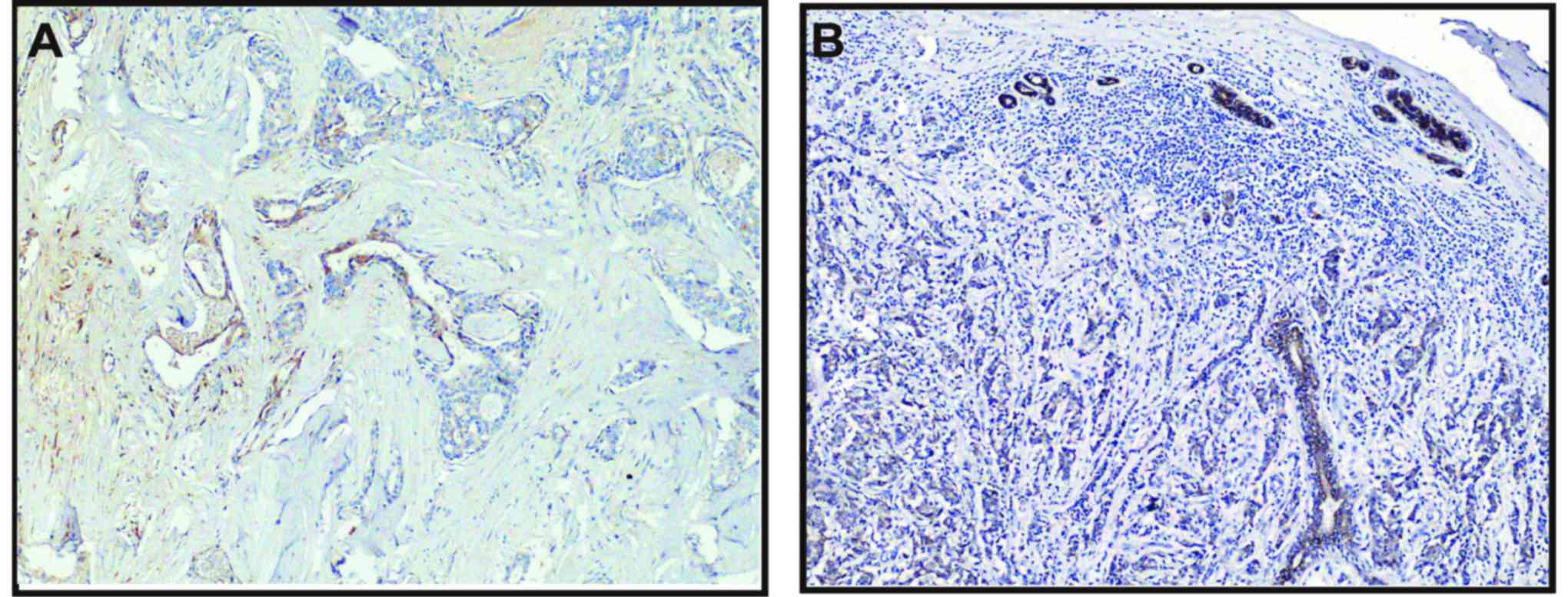
Positive immunoreactivity (brown) in cytoplasm was
observed for P2Y1, P2Y2, P2Y4 and P2Y6 receptors in most breast
cancer cells, but not detected in the tumor-adjacent normal
glandular epithelial cells. Positive P2Y2 staining could also be
noted in the nuclei of some ductal myoepithelial cells (Fig. 11A). The staining of P2Y2 and P2Y6
receptors was more intensive than that of P2Y1 and P2Y4 receptors.
Consistent with the results from breast cancer cell lines, the P2Y2
receptor was markedly expressed in breast tumor tissues.
Particularly, the expression of the P2Y2 receptor was higher at the
invasive edge of tumor, in infiltrating tumor cells in the breast
adipose tissue and the cancer embolus in the lymphatic sinuses,
than that at the tumor core areas (Fig. 11B-D).
 | Figure 11.Positive staining of the P2Y2
receptor in normal tumor-adjacent breast and breast cancer tissues.
In normal tumor-adjacent breast tissues, nuclear staining of the
P2Y2 receptor was observed in various ductal myoepithelial cells
(A, magnification, ×200). In the breast tumor tissues, relative
overexpression of the P2Y2 receptor was noted at the invasive edge
of the tumor (B, magnification, ×200), in infiltrating tumor cells
in the breast adipose tissue (C, magnification, ×200) or the cancer
embolus in the lymphatic sinuses (D, magnification, ×200) compared
to the tumor core. Scale bars, 50 µm. |
Expression levels of EMT-related genes
Snail and E-cadherin were detected in breast cancer tissues
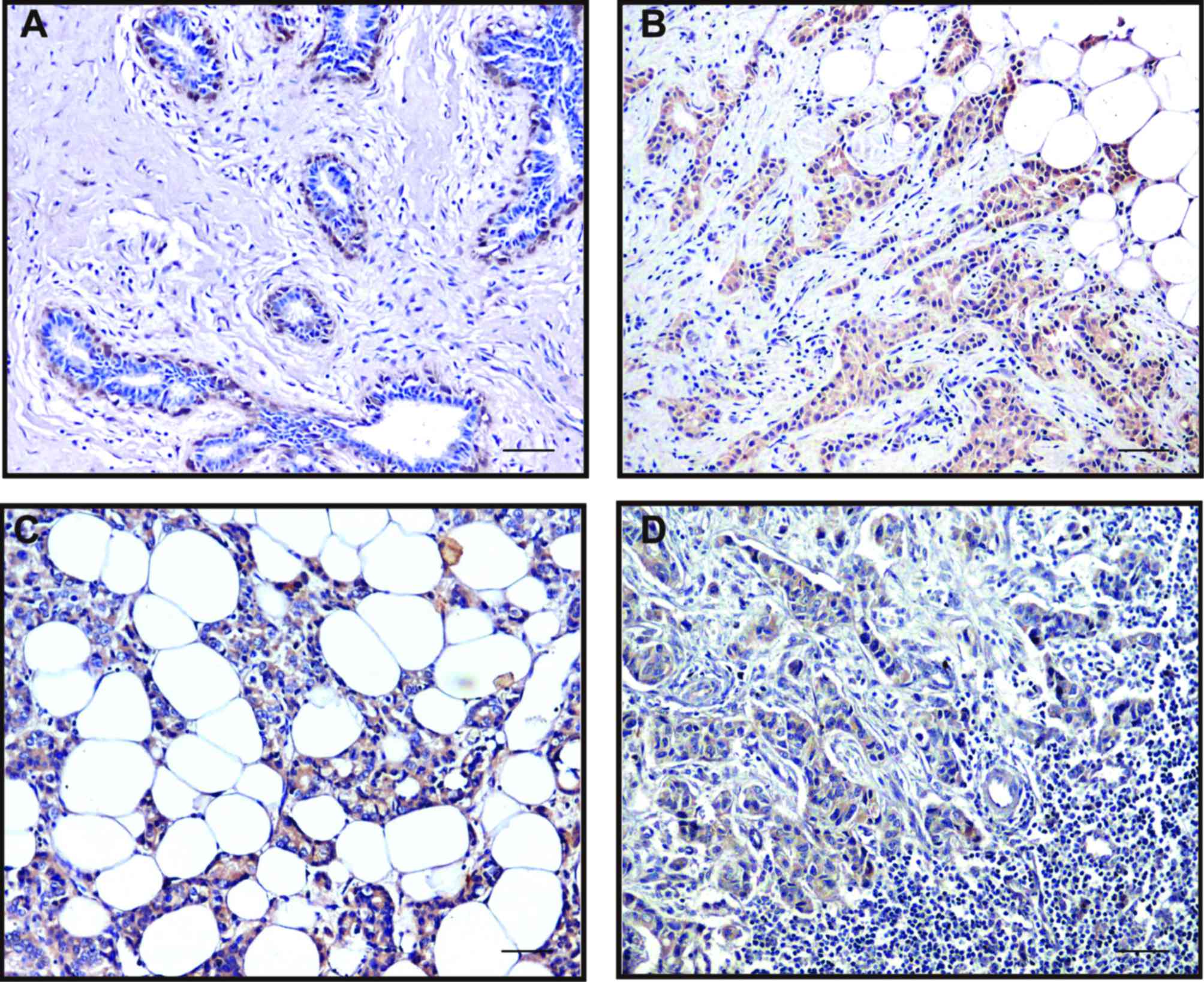
Snail expression in the tumor cells was either
nuclear, often in combination with weak-to-moderate cytoplasmic
positivity, or only cytoplasmic. High Snail expression was observed
at the invasive edge of tumor cells (Fig. 12A). In contrast, the intercellular
borders of the normal glandular epithelial cells were strongly
stained by E-cadherin. The invasive breast carcinoma cells and the
myoepithelial cells of remnant ducts and ductules showed much
weaker or negative staining of E-cadherin at cell-cell borders
(Fig. 12B). As a pair of adverse
markers of epithelial mesenchymal transition, the loss of
E-cadherin expression (negative or weak) was significantly
associated with the high expression of Snail (χ2=41.92,
P<0.01). Meanwhile, we observed the increased expression of the
P2Y2 receptor in the invasive edge of the tumor (Fig. 11B). This indicated a possibility
that upregulation of the P2Y2 receptor coexisted with an increase
in Snail or a reduction in E-cadherin, but a statistical
correlation was not found.
Association of P2Y2 receptor with
clinicopathological parameters of breast cancer
Finally, the associations between expression of P2Y2
receptor and some clinicopathological characteristics were analyzed
(Table I). We found that the
expression level of the P2Y2 receptor was negatively correlated
with histological grade (P=0.023). It was notable that the
immunostaining of the P2Y2 receptor was decreased in tumors with
higher grade. Furthermore, P2Y2 receptor expression was
significantly higher in the tumors with lower Ki-67 index
(r=−0.161, P=0.026) and no HER2 amplification (P=0.008). It is
possible that P2Y2 plays adverse roles in the malignant
proliferation and invasion of breast cancer. We found no
significant correlation between P2Y2 receptor expression and
various other clinical characteristics such as nodule status,
distant metastasis, clinical stage, as well as NPI grouping.
Discussion
In the present study, we demonstrated that
activation of the P2Y2 receptor enhanced breast cancer cell
invasiveness by induction of EMT. The P2Y2 receptor was found to be
highly expressed in MCF-7 and Hs578T cells. In addition, the
migratory and invasive capacities of the breast cancer cells were
markedly increased under stimulation of the P2Y2 receptor. We also
identified in breast cancer tissues that the P2Y2 receptor was
abundantly expressed at the invasive edge of the tumor, in
infiltrating tumor cells in the breast adipose tissue or the cancer
embolus in the lymphatic sinuses compared to the tumor core. In
addition, high expression of Snail and weak or negative expression
of E-cadherin were observed at the invasive edge of the tumor.
Extracellular ATP exerts diverse effects on a
variety of tissues and cell types via specific purinergic
receptors, consisting of P2X and P2Y families (7). Among these P2Y receptor subtypes, the
P2Y2 receptor displays distinct effects on progression of different
types of cancer. In the present study, we showed that the P2Y2
receptor was highly expressed in MCF-7 and Hs578T breast cancer
cell lines. Notably, in normal breast tissue, the P2Y2 receptor was
expressed in the ductal myoepithelial cells with nuclear staining
pattern, but not in the glandular epithelial cells of breast
lobules. However, in the breast cancer tissues, we found that
expression of the P2Y2 receptor in the tumor cells was
significantly higher than that in the tumor-adjacent normal
lobules. It has been reported that a high concentration of ATP, as
an agonist of the P2Y2 receptor, was observed at tumor sites
(4). ATP can be released from
infiltrating inflammatory cells as well as from the tumor cells via
different mechanisms, such as granule exocytosis, plasma membrane
channels, or cell lysis. Therefore, P2Y2 receptor expression may be
a sequential event due to the elevated extracellular ATP in the
microenvironment of breast cancer.
Some researchers have reported that activation of
the P2Y2 receptor promoted cell proliferation in melanoma and lung
cancer cells (22,23). On the contrary, the P2Y2 receptor
inhibited cell proliferation of esophageal cancer and some type of
colorectal cancer (24,25). Our previous studies demonstrated
that ATP promoted migration, invasion and metastasis of prostate
cancer cells, while it inhibited cell growth (10,11,26,27).
In the present study, we found that the P2Y2 receptor expression
was negatively correlated with histological grade, HER2
amplification and Ki67 index of breast cancer, which indicated that
the P2Y2 receptor may participate in the inhibition of tumor
proliferation.
Various studies strongly suggest that ATP is a
crucial microenvironmental element in driving metastatic spread
(28). Schumacher et al
pinpointed the nucleotide as an endothelia relaxant, facilitating
tumor cell escape (29).
Particularly, ATP release was inevitable in the borderline of the
tumor nest as a result of tissue destruction and disintegration of
inflammatory cells. Jelassi et al identified ATP as a
mediator of tumor infiltration (30). In the present study, the P2Y2
receptor mediated ATP-promoted migration and invasion of breast
cancer cells in vitro. Similarly, Chadet et al
demonstrated that activation of the P2Y2 receptor increased MCF-7
breast cancer cell migration through the MEK-ERK1/2 signaling
pathway (31). In breast cancer
tissues, we also observed a relative overexpression of the P2Y2
receptor at the invasive edge of the tumor, in infiltrating tumor
cells in the breast adipose tissue or the cancer embolus in the
lymphatic sinuses compared to the tumor core. These data strongly
suggest that the P2Y2 receptor is a key receptor mediating the
migration and invasion of breast cancer.
The tumor microenvironment plays a pivotal role in
tumor cell proliferation, survival and migration. Signals from the
tumor microenvironment could trigger cancer cells to adopt an
invasive phenotype through EMT, which is thought to promote cancer
cell progression and invasion into the surrounding
microenvironment. It is a multi-step process in which cells acquire
molecular alterations that facilitate dysfunctional cell-cell
adhesive interactions, loss of cell-cell junctions, and
reorganization of the cytoskeleton, all of which result in the loss
of epithelial characteristics and the acquisition of a more
spindle-shaped morphology defined as ‘transient mesenchymal
conversion’ (2,32,33).
EMT is typically characterized by the loss of intercellular
adhesion and polarity, downregulation of epithelial markers,
upregulation of mesenchymal markers and acquisition of increased
motility, invasiveness and metastatic capabilities (2). The full accomplishment of EMT requires
a complex genetic programme that is partially mediated by a number
of specific transcription factors, such as Snail, Slug and Twist.
These factors are expressed in a variety of cell types and modulate
directly or indirectly the expression of a great number of genes
involved in invasion and metastasis of cancer (34). It has been reported that ATP
participates in the EGF-induced EMT process in breast cancer cells
(35). Our previous studies
demonstrated that ATP induced EMT in prostate cancer cells
(11,26). In the present study, we found that
extracellular ATP led to significant upregulation of Snail and
marked downregulation of E-cadherin and claudin-1 in breast cancer
cells in vitro. After knockdown of the P2Y2 receptor, the
changes in the expression of ATP-induced EMT-related genes were
markedly inhibited, indicating that the P2Y2 receptor participates
in ATP-induced EMT in breast cancer cells. Davis et al
demonstrated that induction of EMT in breast cancer cells is
calcium signal dependent. However, it was reported that ATP
modulated intracellular Ca2+ by activation of P2Y2 and
P2Y4 receptors in MCF7 cells (36).
Therefore, it is possible that activation of the P2Y2 receptor
induces EMT via the calcium signaling pathway in breast cancer
cells. Several studies provide support for the presence of EMT
markers in invasive breast cancer, particularly in the
invasion-metastasis cascade (32,37).
Sarrió et al demonstrated that EMT-like changes occur
preferentially in the basal subtype of breast carcinomas with high
aggressiveness and metastatic capabilities (32). Similarly, Snail was found to be
expressed in all the infiltrating ductal carcinomas (IDC)
presenting lymph node metastasis and its expression was found to be
inversely correlated with the grade of differentiation of breast
tumors (38). As for the breast
cancer tissues in the present study, while the intercellular
borders of the tumor-adjacent normal glandular structures present
in breast carcinoma specimens were strongly stained by E-cadherin,
the invasive breast carcinoma cells showed much weaker or negative
staining at the cell-cell borders. Nevertheless, Snail was highly
expressed at the invasive edge of breast tumor tissue next to the
breast adipose tissue, where the elevated expression of the P2Y2
receptor was also observed. Collectively, these results indicate
that extracellular ATP may induce EMT at the invasive edge via
activation of the P2Y2 receptor. Additionally, it was reported that
P2Y receptors participate in TGFβ-induced EMT of MDCK cells
(39). Our previous study also
showed that the P2Y2 receptor mediated ATP-induced EMT in prostate
cancer cells (11). However, we did
not find any association between the expression of the P2Y2
receptor and E-cadherin or Snail protein in breast cancer tissues,
which could be due to the complex microenvironment of invasive
breast cancer. Nevertheless, all these data suggest the important
role of the P2Y2 receptor in mediating ATP-induced EMT in breast
cancer.
In conclusion, the present study demonstrated that
the P2Y2 receptor was critical for mediating ATP-promoted migration
and invasion as well as ATP-driven expression changes in
EMT-related genes Snail and E-cadherin. The critical role of the
P2Y2 receptor indicates it could be a potential therapeutic target
for the treatment of breast cancer. However, there are multiple P2Y
receptor subtypes expressed in breast cancer cells, and the
contribution of other receptor subtypes in ATP-mediated progression
of breast cancer cannot be fully excluded. Investigation of the
detailed molecular mechanisms of purinergic signaling in the
progression of breast cancer may still be our focus in future
research.
Acknowledgements
We thank all the individuals who participated in the
present study. The present study was supported by grants to W.-G.F.
from the National Natural Science Foundation of China
(81621063).
References
|
1
|
Siegel R, Ma J, Zou Z and Jemal A: Cancer
statistics, 2014. CA Cancer J Clin. 64:9–29. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Thiery JP: Epithelial-mesenchymal
transitions in tumour progression. Nat Rev Cancer. 2:442–454. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Murata Y, Yasuo T, Yoshida R, Obata K,
Yanagawa Y, Margolskee RF and Ninomiya Y: Action potential-enhanced
ATP release from taste cells through hemichannels. J Neurophysiol.
104:896–901. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Pellegatti P, Raffaghello L, Bianchi G,
Piccardi F, Pistoia V and Di Virgilio F: Increased level of
extracellular ATP at tumor sites: In vivo imaging with plasma
membrane luciferase. PLoS One. 3:e25992008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wang X, Arcuino G, Takano T, Lin J, Peng
WG, Wan P, Li P, Xu Q, Liu QS, Goldman SA, et al: P2×7 receptor
inhibition improves recovery after spinal cord injury. Nat Med.
10:821–827. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Raffaghello L, Chiozzi P, Falzoni S, Di
Virgilio F and Pistoia V: The P2×7 receptor sustains the growth of
human neuroblastoma cells through a substance P-dependent
mechanism. Cancer Res. 66:907–914. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Di Virgilio F: Purines, purinergic
receptors, and cancer. Cancer Res. 72:5441–5447. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Stagg J and Smyth MJ: Extracellular
adenosine triphosphate and adenosine in cancer. Oncogene.
29:5346–5358. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chen L, He HY, Li HM, Zheng J, Heng WJ,
You JF and Fang WG: ERK1/2 and p38 pathways are required for P2Y
receptor-mediated prostate cancer invasion. Cancer Lett.
215:239–247. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhang Y, Gong LH, Zhang HQ, Du Q, You JF,
Tian XX and Fang WG: Extracellular ATP enhances in vitro invasion
of prostate cancer cells by activating Rho GTPase and upregulating
MMPs expression. Cancer Lett. 293:189–197. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Li WH, Qiu Y, Zhang HQ, Liu Y, You JF,
Tian XX and Fang WG: P2Y2 receptor promotes cell invasion and
metastasis in prostate cancer cells. Br J Cancer. 109:1666–1675.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Liu J, Liao Z, Camden J, Griffin KD,
Garrad RC, Santiago-Pérez LI, González FA, Seye CI, Weisman GA and
Erb L: Src homology 3 binding sites in the P2Y2 nucleotide receptor
interact with Src and regulate activities of Src, proline-rich
tyrosine kinase 2, and growth factor receptors. J Biol Chem.
279:8212–8218. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Tu MT, Luo SF, Wang CC, Chien CS, Chiu CT,
Lin CC and Yang CM: P2Y2 receptor-mediated proliferation of C6
glioma cells via activation of Ras/Raf/MEK/MAPK pathway. Br J
Pharmacol. 129:1481–1489. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Muscella A, Elia MG, Greco S, Storelli C
and Marsigliante S: Activation of P2Y2 receptor induces c-FOS
protein through a pathway involving mitogen-activated protein
kinases and phosphoinositide 3-kinases in HeLa cells. J Cell
Physiol. 195:234–240. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Nylund G, Hultman L, Nordgren S and Delbro
DS: P2Y2- and P2Y4 purinergic receptors are over-expressed in human
colon cancer. Auton Autacoid Pharmacol. 27:79–84. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Künzli BM, Bernlochner MI, Rath S, Käser
S, Csizmadia E, Enjyoji K, Cowan P, d'Apice A, Dwyer K, Rosenberg
R, et al: Impact of CD39 and purinergic signalling on the growth
and metastasis of colorectal cancer. Purinergic Signal. 7:231–241.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Early Breast Cancer Trialists'
Collaborative Group (EBCTCG), . Effects of chemotherapy and
hormonal therapy for early breast cancer on recurrence and 15-year
survival: An overview of the randomised trials. Lancet.
365:1687–1717. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Blamey RW, Ellis IO, Pinder SE, Lee AH,
Macmillan RD, Morgan DA, Robertson JF, Mitchell MJ, Ball GR,
Haybittle JL, et al: Survival of invasive breast cancer according
to the Nottingham Prognostic Index in cases diagnosed in 1990–1999.
Eur J Cancer. 43:1548–1555. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Schmittgen TD and Livak KJ: Analyzing
real-time PCR data by the comparative C(T) method. Nat Protoc.
3:1101–1108. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Albini A, Iwamoto Y, Kleinman HK, Martin
GR, Aaronson SA, Kozlowski JM and McEwan RN: A rapid in vitro assay
for quantitating the invasive potential of tumor cells. Cancer Res.
47:3239–3245. 1987.PubMed/NCBI
|
|
21
|
Matos LL, Stabenow E, Tavares MR, Ferraz
AR, Capelozzi VL and Pinhal MA: Immunohistochemistry quantification
by a digital computer-assisted method compared to semiquantitative
analysis. Clinics. 61:417–424. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Schafer R, Sedehizade F, Welte T and
Reiser G: ATP- and UTP-activated P2Y receptors differently regulate
proliferation of human lung epithelial tumor cells. Am J Physiol
Lung Cell Mol Physiol. 285:L376–L385. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
White N, Ryten M, Clayton E, Butler P and
Burnstock G: P2Y purinergic receptors regulate the growth of human
melanomas. Cancer Lett. 224:81–91. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Maaser K, Höpfner M, Kap H, Sutter AP,
Barthel B, von Lampe B, Zeitz M and Scherübl H: Extracellular
nucleotides inhibit growth of human oesophageal cancer cells via
P2Y2-receptors. Br J Cancer. 86:636–644. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Höpfner M, Maaser K, Barthel B, von Lampe
B, Hanski C, Riecken EO, Zeitz M and Scherübl H: Growth inhibition
and apoptosis induced by P2Y2 receptors in human colorectal
carcinoma cells: Involvement of intracellular calcium and cyclic
adenosine monophosphate. Int J Colorectal Dis. 16:154–166. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Qiu Y, Li WH, Zhang HQ, Liu Y, Tian XX and
Fang WG: P2×7 mediates ATP-driven invasiveness in prostate cancer
cells. PLoS One. 9:e1143712014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Fang WG, Pirnia F, Bang YJ, Myers CE and
Trepel JB: P2-purinergic receptor agonists inhibit the growth of
androgen-independent prostate carcinoma cells. J Clin Invest.
89:191–196. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Adinolfi E: New intriguing roles of ATP
and its receptors in promoting tumor metastasis: Presented by Maria
P. Abbracchio. Purinergic Signal. 9:487–490. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Schumacher D, Strilic B, Sivaraj KK,
Wettschureck N and Offermanns S: Platelet-derived nucleotides
promote tumor-cell transendothelial migration and metastasis via
P2Y2 receptor. Cancer Cell. 24:130–137. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Jelassi B, Anchelin M, Chamouton J,
Cayuela ML, Clarysse L, Li J, Goré J, Jiang LH and Roger S:
Anthraquinone emodin inhibits human cancer cell invasiveness by
antagonizing P2×7 receptors. Carcinogenesis. 34:1487–1496. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Chadet S, Jelassi B, Wannous R, Angoulvant
D, Chevalier S, Besson P and Roger S: The activation of P2Y2
receptors increases MCF-7 breast cancer cells migration through the
MEK-ERK1/2 signalling pathway. Carcinogenesis. 35:1238–1247. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Sarrió D, Rodriguez-Pinilla SM, Hardisson
D, Cano A, Moreno-Bueno G and Palacios J: Epithelial-mesenchymal
transition in breast cancer relates to the basal-like phenotype.
Cancer Res. 68:989–997. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Trimboli AJ, Fukino K, de Bruin A, Wei G,
Shen L, Tanner SM, Creasap N, Rosol TJ, Robinson ML, Eng C, et al:
Direct evidence for epithelial-mesenchymal transitions in breast
cancer. Cancer Res. 68:937–945. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Lee JM, Dedhar S, Kalluri R and Thompson
EW: The epithelial-mesenchymal transition: New insights in
signaling, development, and disease. J Cell Biol. 172:973–981.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Davis FM, Kenny PA, Soo ET, van Denderen
BJ, Thompson EW, Cabot PJ, Parat MO, Roberts-Thomson SJ and
Monteith GR: Remodeling of purinergic receptor-mediated
Ca2+ signaling as a consequence of EGF-induced
epithelial-mesenchymal transition in breast cancer cells. PLoS One.
6:e234642011. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Bilbao P Scodelaro, Boland R, de Boland A
Russo and Santillán G: ATP modulation of mitogen activated protein
kinases and intracellular Ca2+ in breast cancer (MCF-7)
cells. Arch Biochem Biophys. 466:15–23. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Ma L, Teruya-Feldstein J and Weinberg RA:
Tumour invasion and metastasis initiated by microRNA-10b in breast
cancer. Nature. 449:682–688. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Blanco MJ, Moreno-Bueno G, Sarrio D,
Locascio A, Cano A, Palacios J and Nieto MA: Correlation of Snail
expression with histological grade and lymph node status in breast
carcinomas. Oncogene. 21:3241–3246. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Wolff CI, Bundey RA and Insel PA:
Involvement of P2Y receptors in TGFβ-induced EMT of MDCK cells.
FASEB J. 22:942–945. 2008.
|