Introduction
Breast cancer is the most common malignant tumour
and the leading cause of cancer mortality in women, resulting in
14% of cancer-related deaths (1,2).
Although great improvements have been made in the diagnosis and
treatment of breast cancer, the prognosis of patients with advanced
breast cancer is still not satisfactory. In recent decades, many
oncogenes and tumour suppressors have been found to play key roles
in the malignant progression of breast cancer, and an understanding
of the molecular mechanisms underlying breast cancer progression
seems to be beneficial for the development of novel therapeutic
strategies for this disease (3,4).
MicroRNAs (miRs), a type of non-coding RNA that are
18–25 nucleotides in length, have been demonstrated to function as
gene expression regulators by directly binding to the
3-untranslated regions (UTR) of their target mRNAs, which
ultimately leads to mRNA degradation or translation inhibition
(5,6). In the most recent decade, various miRs
have been implicated in a variety of physiological and pathological
processes, such as cell development and differentiation,
angiogenesis and cell motility, as well as tumourigenesis (7,8).
Moreover, many miRs, including miR-133a, have been observed to be
significantly dysregulated in breast cancer and play suppressive or
oncogenic roles (9–11).
Recently, miR-133a has been suggested to play an
important role in some common human cancers (12,13).
For instance, Zheng et al (13) showed that miR-133a could inhibit
colorectal cancer cell invasion through inhibition of the protein
expression of Fascin1. Moreover, the suppressive role of miR-133a
in breast cancer has been suggested in several recent studies
(11,14,15).
For instance, Wu et al (14)
examined miR-133a expression in formalin-fixed paraffin-embedded
breast specimens from 26 benign, 34 pericancerous normal and 90
cancerous tissues using in situ hybridization and found that
miR-133a was gradually downregulated from normal to benign to
cancerous breast tissues. They further showed that downregulation
of miR-133a was associated with disease progression and poor
prognosis in breast cancer patients (14). Moreover, Ji et al (11) showed an in vivo antitumour
effect with ultrasound-mediated miRNA-133a microbubble delivery in
breast cancer. In addition, the oncogenic gene FSCN1 was identified
as a direct target gene of miR-133a in breast cancer (15). However, whether other target genes
of miR-133a exist in breast cancer still needs to be studied.
Growing evidence shows that LIM and SH3 domain
protein 1 (LASP1) is a multi-functional protein that plays
important roles in cytoskeletal formation and is a prognostic
marker in different cancers (16).
LASP1 expression is correlated with the grade, size and occurrence
of metastasis in clinical tumour samples (17). LASP1 is a target of miRNAs, such as
miR-203, miR-29a and miR-133b, and has been found in lung, breast
cancer and hepatocarcinoma (18–20).
However, whether LASP1 is targeted by miR-133a in breast cancer
remains unknown. Thus, the present study aimed to investigate the
molecular mechanisms by which miR-133a mediates the malignant
progression of this disease.
Materials and methods
Clinical samples
This study was approved by the Ethics Committee of
Dongying People's Hospital (Dongying, China). We collected a total
of 78 primary breast cancer tissues and their matched adjacent
non-tumour tissues when patients underwent surgical resection at
Dongying Peoples Hospital from April 2010 to October 2011. All the
patients were female and aged 28–74 years. Of the total patients,
35.9% (28/78) presented at advanced stage (III/IV) and 74.4%
(58/78) presented with nodal metastasis (Table I). No patient had received
radiotherapy or chemotherapy prior to surgery, and informed consent
was obtained. The clinical information for these patients is
summarized in Table I.
 | Table I.Association between the miR-133a
expression and clinicopathological characteristics in breast
cancer. |
Table I.
Association between the miR-133a
expression and clinicopathological characteristics in breast
cancer.
| Variables | N (n=78) | Low expression
(n=47) | High expression
(n=31) | P-value |
|---|
| Age (years) |
|
|
| 1.00 |
|
≤50 | 35 | 21 | 14 |
|
|
>50 | 43 | 26 | 17 |
|
| Tumor size |
|
|
| 0.496 |
|
T1-T2 | 44 | 25 | 19 |
|
|
T3-T4 | 34 | 22 | 12 |
|
| Grade |
|
|
| 0.485 |
| Well
and moderately | 49 | 28 | 21 |
|
|
Poor | 29 | 19 | 10 |
|
| Lymph node
metastasis |
|
|
| 0.038a |
|
Present | 58 | 39 | 19 |
|
|
Absent | 20 | 8 | 12 |
|
| Distant
metastasis |
|
|
| 0.0096b |
|
Present | 9 | 9 | 0 |
|
|
Absent | 69 | 38 | 31 |
|
| TNM stage |
|
|
| 0.001b |
|
I–II | 50 | 23 | 27 |
|
|
III–IV | 28 | 24 | 4 |
|
Cell culture and transfection
Human normal breast cell line Hs 578Bst and human
breast cancer cell lines MCF-7, MDA-MB-231, SK-BR-3, BT-20 and
MDA-MB-361 were purchased from the American Type Culture Collection
(ATCC; Manassas, VA, USA). All cell lines were cultured in
Dulbeccos modified Eagles medium (DMEM; Thermo Fisher Scientific,
Waltham, MA, USA) supplemented with 10% fetal bovine serum (FBS;
Thermo Fisher Scientific) in a 37°C humidified atmosphere of 5%
CO2. For the functional study of miR-133a and LASP1 in
breast cancer in vitro, SK-BR-3 and MDA-MB-231 cells were
transfected with scramble miR (namely, miR-NC), miR-133a mimic,
negative control (NC) inhibitor, miR-133a inhibitor, NC siRNA, or
LASP1 siRNA, or co-transfected with miR-133a mimic and blank
pc-DNA3.1 vector (namely, miR-133a+NC) or miR-133a mimic and
pc-DNA3.1-LASP1 plasmid (namely, miR-133a+LASP1) using
Lipofectamine 2000 (Thermo Fisher Scientific) according to the
manufacturers instructions. For in vivo study, SK-BR-3 and
MDA-MB-231 cells were stably transfected with the blank pLVTH
vector or pLVTH-miR-133a lentiviral plasmid, respectively. The
miR-133a inhibitor, miR-133a mimic and the negative control were
purchased from Shanghai GenePharma Co., Ltd. (Shanghai, China) and
the miR-133a lentiviral plasmid and blank plasmid were purchased
from GeneCopoeia (Guangzhou, China). At 48 h after cell
transfection, the mRNA and protein expression levels were
examined.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted using TRIzol reagent (Thermo
Fisher Scientific), according to the manufacturers instructions.
After that, the total RNA was converted into cDNA using a Reverse
Transcription kit (Thermo Fisher Scientific) according to the
manufacturers instructions. For miR or mRNA expression, real-time
PCR was performed using an All-in-One miRNA qPCR kit (GeneCopoeia,
Rockville, MD, USA) or a Thermo One-Step RT-PCR kit (Thermo Fisher
Scientific) according to the manufacturers instructions. U6 and
GAPDH were used as internal references. The specific primer pairs
used are as follows: miR-133a, forward,
5-TGCTTTGCTAGAGCTGGTAAAATG-3 and reverse, 5-AGCTACAGCTGGTTGAAGGG-3;
U6, forward, 5-CTCGCTTCGGCAGCACA-3 and reverse, 5-AACGCT
TCACGAATTTGCGT-3; LASP1, forward, 5-TGCGGCAA GATCGTGTATCC-3 and
reverse, 5-GCAGTAGGGCTTC TTCTCGTAG-3; GAPDH, forward, 5-GGAGCGAGATC
CCTCCAAAAT-3 and reverse, 5-GGCTGTTGTCATACT TCTCATGG-3. The PCR
reaction conditions were 95°C for 3 min, followed by 40 cycles of
denaturation at 95°C for 15 sec and an annealing/elongation step at
60°C for 30 sec. The relative expression was analysed by the
2−ΔΔCt method.
Western blot analysis
Tissues and cells were lysed in cold
radioimmunoprecipitation assay buffer (Thermo Fisher Scientific),
and a Bicinchoninic Acid Protein Assay kit (Thermo Fisher
Scientific) was used to determine the protein concentration,
according to the manufacturers instructions. After that, 60 µg of
protein was separated with 10% SDS-PAGE and was then transferred to
a polyvinylidene difluoride membrane (Thermo Fisher Scientific).
The membrane was then blocked in 5% non-fat milk in
phosphate-buffered saline (PBS; Thermo Fisher Scientific)
containing 0.1% Tween-20 (Sigma-Aldrich, St. Louis, MO, USA) at
room temperature for 3 h. After being washed with PBS for 15 min,
the membrane was incubated with rabbit anti-LASP1 primary antibody
(1:500; Abcam, Cambridge, UK) or rabbit anti-GAPDH primary antibody
(1:500; Abcam) at room temperature for 3 h. After being washed with
PBS for 15 min, the membrane was incubated with goat anti-rabbit
secondary antibody (1:5,000; Abcam) at room temperature for 40 min.
After being washed with PBS for 15 min, the protein band was
detected using an Enhanced Chemiluminescence Western Blotting kit
(Thermo Fisher Scientific) and then quantified using Image Lab
analysis software 3.1 (Bio-Rad Laboratories, Inc., Hercules, CA,
USA), according to the manufacturers instructions.
MTT assay
An MTT assay was used to examine cell proliferation.
SK-BR-3 and MDA-MB-231 cells (5×103/well) were plated
into a 96-well plate and cultured at 37°C with 5% CO2
for 12, 24, 48 or 72 h. Subsequently, 20 µl MTT (5 mg/ml; Life
Technologies/Thermo Fisher Scientific) was added. Following
incubation at 37°C for 4 h, 150 µl dimethyl sulfoxide (DMSO; Thermo
Fisher Scientific) was added. Following incubation at room
temperature for 10 min, formazan production was detected by
determining the optical density at 570 nm using a Multiskan FC
enzyme immunoassay analyser (Thermo Fisher Scientific).
Wound healing assay
SK-BR-3 and MDA-MB-231 cells were cultured in DMEM
with 10% FBS to full confluence. After that, we created a wound
using a plastic scraper. After being washed with PBS, cells were
incubated in DMEM at 37°C for 24 h. Then, the medium was replaced
with DMEM with 10% FBS. After incubation at 37°C for 48 h, the
wound was observed under a microscope (Nikon, Tokyo, Japan).
Transwell assay
SK-BR-3 and MDA-MB-231 cell suspensions were added
to the Matrigel (Chemicon, Temecula, CA, USA) pre-coated upper
chamber (Chemicon), and 300 µl of DMEM with 10% FBS was added into
the lower chamber. Cells were then incubated at 37°C for 24 h.
After that, those cells not through the membrane of the insert were
removed using a cotton-tipped swab. Those cells on the lower
surface of the membrane were stained with gentian violet
(Sigma-Aldrich) and counted under a microscope (Olympus, Tokyo,
Japan).
Tumour growth in vivo
Male BALB/C-nu/nu nude mice (12 weeks, n=5 in each
group) were maintained under pathogen-free conditions at the Animal
Center of Central South University. MDA-MB-231 cells were stably
transfected with the blank pLVTH vector or pLVTH-miR-133a
lentiviral plasmid, which were injected subcutaneously in the
dorsal flank of the mice. All mice were sacrificed under
anaesthesia 60 days after cell injection. The tumour tissues were
photographed and the weight was examined. The volume of tumour
tissues was calculated according to: V (mm3) = 0.5 × a ×
b2 where a is the maximum length of diameter and b is
the maximum transverse diameter.
Bioinformatics prediction
The putative targets of miR-133a were predicted
using TargetScan software (http://www.targetscan.org), MiRanda (http://www.microrna.org) and PicTar (pictar.mdc-berlin.de) according to the manufacturers
instructions.
Luciferase reporter gene assay
The predicted miR-133a binding sites on the 3UTR of
LASP1 (500 bp) were cloned into the pGL3 vector (Promega, Madison,
WI, USA), named WT-LASP1-3UTR. The mutant miR-133a binding sites on
the 3UTR of LASP1 were constructed using a QuikChange Site-Directed
Mutagenesis kit (Stratagene; Agilent Technologies, Santa Clara, CA,
USA), which was then inserted into the pGL3 vector, named
Mut-LASP1-3UTR. SK-BR-3 and MDA-MB-231 cells were co-transfected
with WT-LASP1-3UTR or Mut-LASP1-3UTR plasmid and miR-NC or miR-133a
mimic, respectively, using Lipofectamine 2000. After transfection
for 48 h, the luciferase activity was examined using the
Dual-Luciferase reporter assay system (Promega) according to the
manufacturers instruction.
Statistical analysis
Data in the present study was expressed as the means
± standard error. The statistical analysis was conducted using
SPSS.20 software. The Students t-test was performed for two-group
comparisons, or one-way analysis of variance was performed for
multiple-group comparisons. The association of gene expression with
clinical characteristics was analysed using a chi-square test. A
P<0.05 was considered significantly different.
Results
Downregulation of miR-133a in breast
cancer
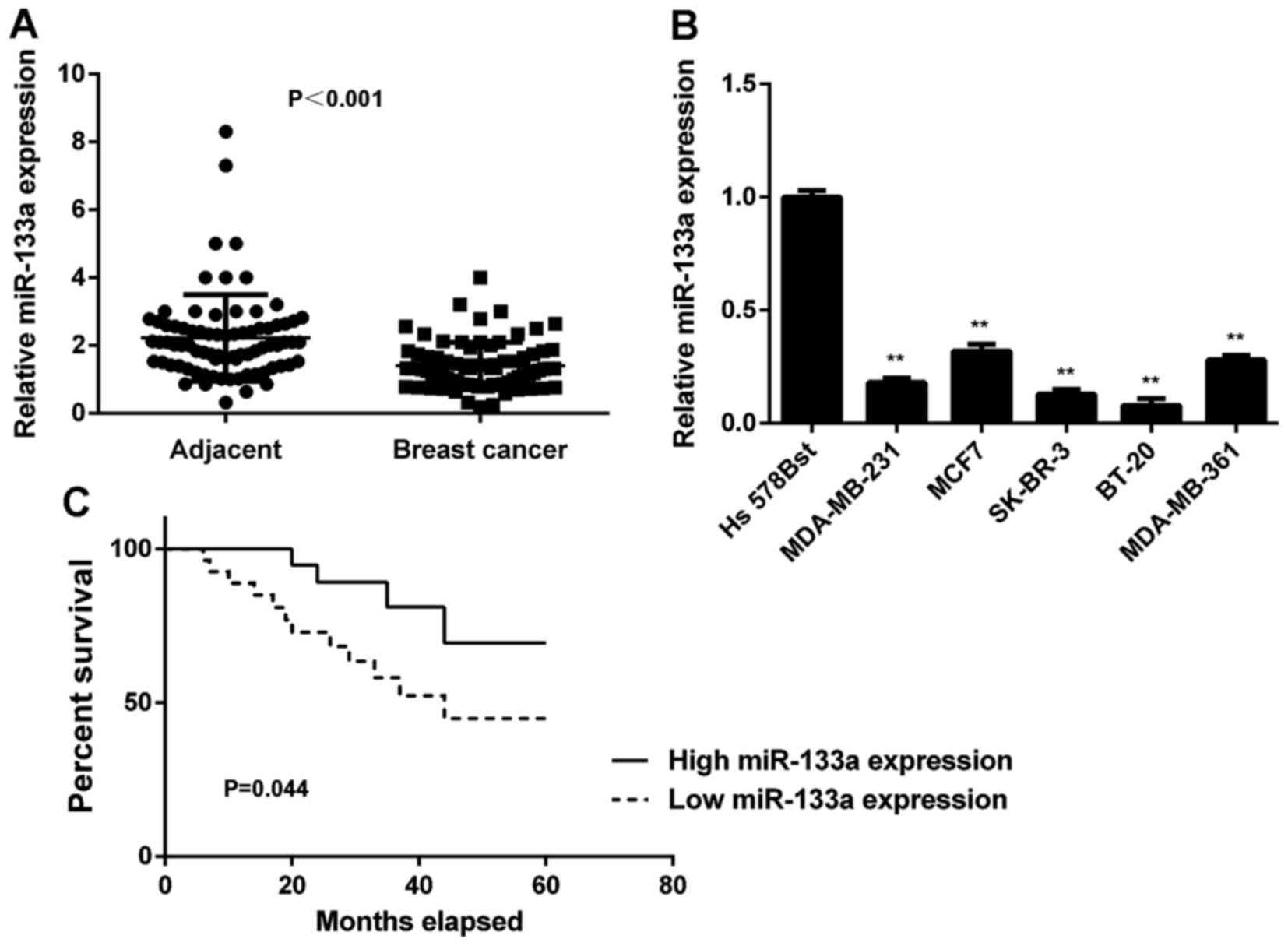
In this study, we first determined the miR-133a
levels in breast cancer tissues and matched adjacent non-tumour
tissues using real-time qPCR. We found that the expression levels
of miR-133a were significantly decreased in breast cancer tissues
compared with adjacent non-tumour tissues (Fig. 1A). To further confirm these
findings, we examined miR-133a expression in several common breast
cancer cell lines, including MCF-7, MDA-MB-231, MDA-MB-361, SK-BR-3
and BT-20. Normal human breast cell line Hs 578Bst was used as a
normal control. As indicated in Fig.
1B, miR-133a was also significantly downregulated in these
breast cancer cell lines compared with Hs 578Bst cells. Therefore,
miR-133a is downregulated in breast cancer.
We then studied the association between miR-133a
expression and clinicopathological characteristics in breast
cancer. The median of miR-133a expression was used as the cut-off
value. If the expression of miR-133a was greater than the median,
it was defined as high expression; otherwise it was defined as low
expression. As indicated in Table
I, low expression of miR-133a was significantly associated with
lymph node metastasis and advanced clinical stage in breast cancer.
Therefore, downregulation of miR-133a is associated with the
malignant progress of breast cancer.
Next, we studied the association between miR-133a
expression and the prognosis of patients with breast cancer. As
shown in Fig. 1C, patients with low
miR-133a expression showed shorter survival time compared with
those with high miR-133a expression. These findings suggest that
the downregulation of miR-133a may predicate poor prognosis in
breast cancer patients.
Restoration of miR-133a expression
inhibits breast cancer cell growth, migration and invasion
As SK-BR-3 and MDA-MB-231 cells showed the most
significant reduction in miR-133a expression, we used these two
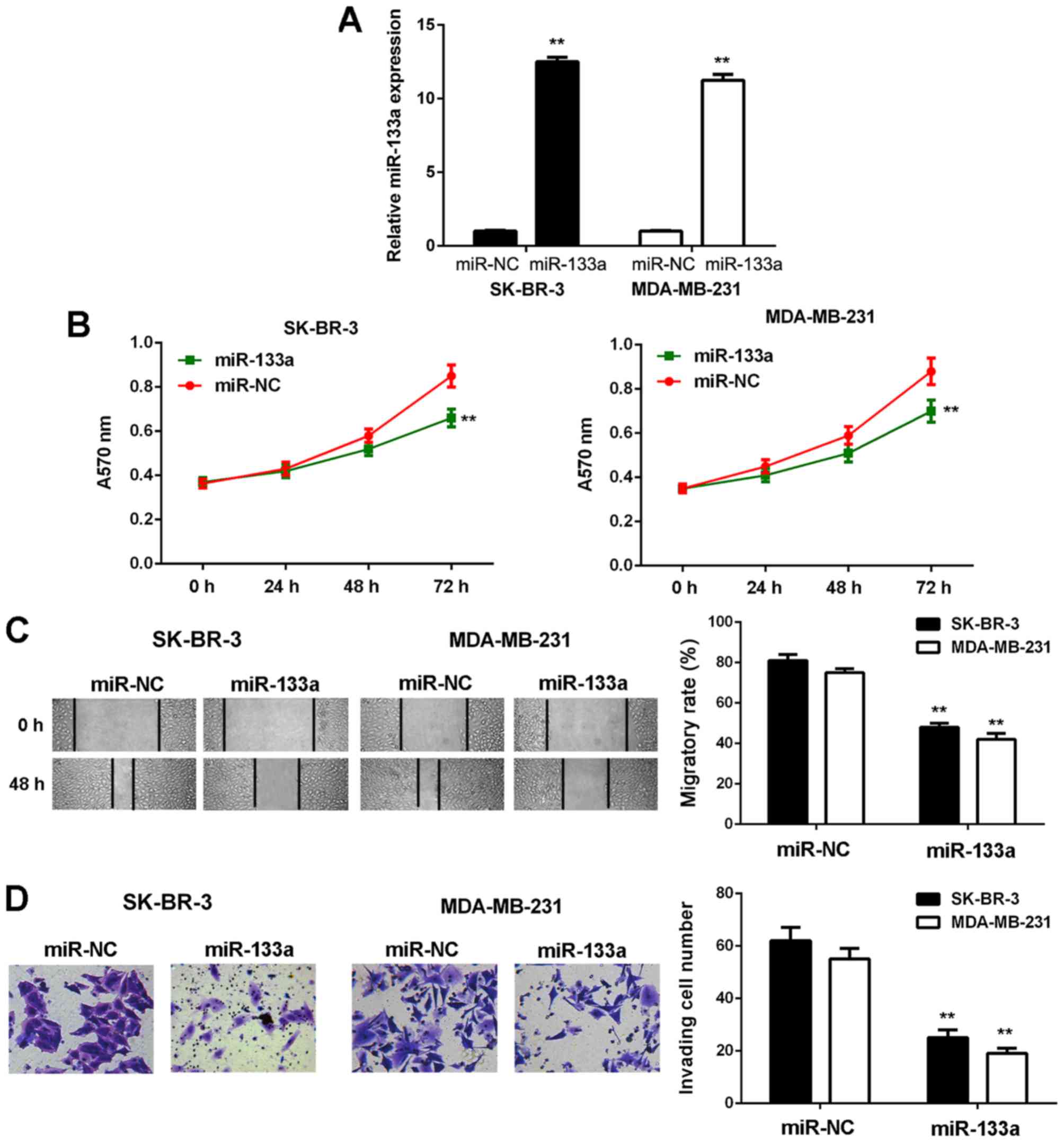
cell lines in the following experiments. To restore its expression,
SK-BR-3 and MDA-MB-231 cells were first transfected with miR-133a
mimic. Transfection with scramble miR mimic was used as the NC
group. As shown in Fig. 2A, the
miR-133a levels were significantly increased in the miR-133a group
compared with the NC group.
MTT, wound healing and Transwell assays were then
conducted to examine cell proliferation, migration and invasion
in vitro. As shown in Fig.
2, the proliferative, migratory and invasive capacities of
SK-BR-3 and MDA-MB-231 cells were significantly downregulated in
the miR-133a group compared with the NC group. These findings
demonstrate that miR-133a has suppressive effects on the malignant
phenotypes of breast cancer cells.
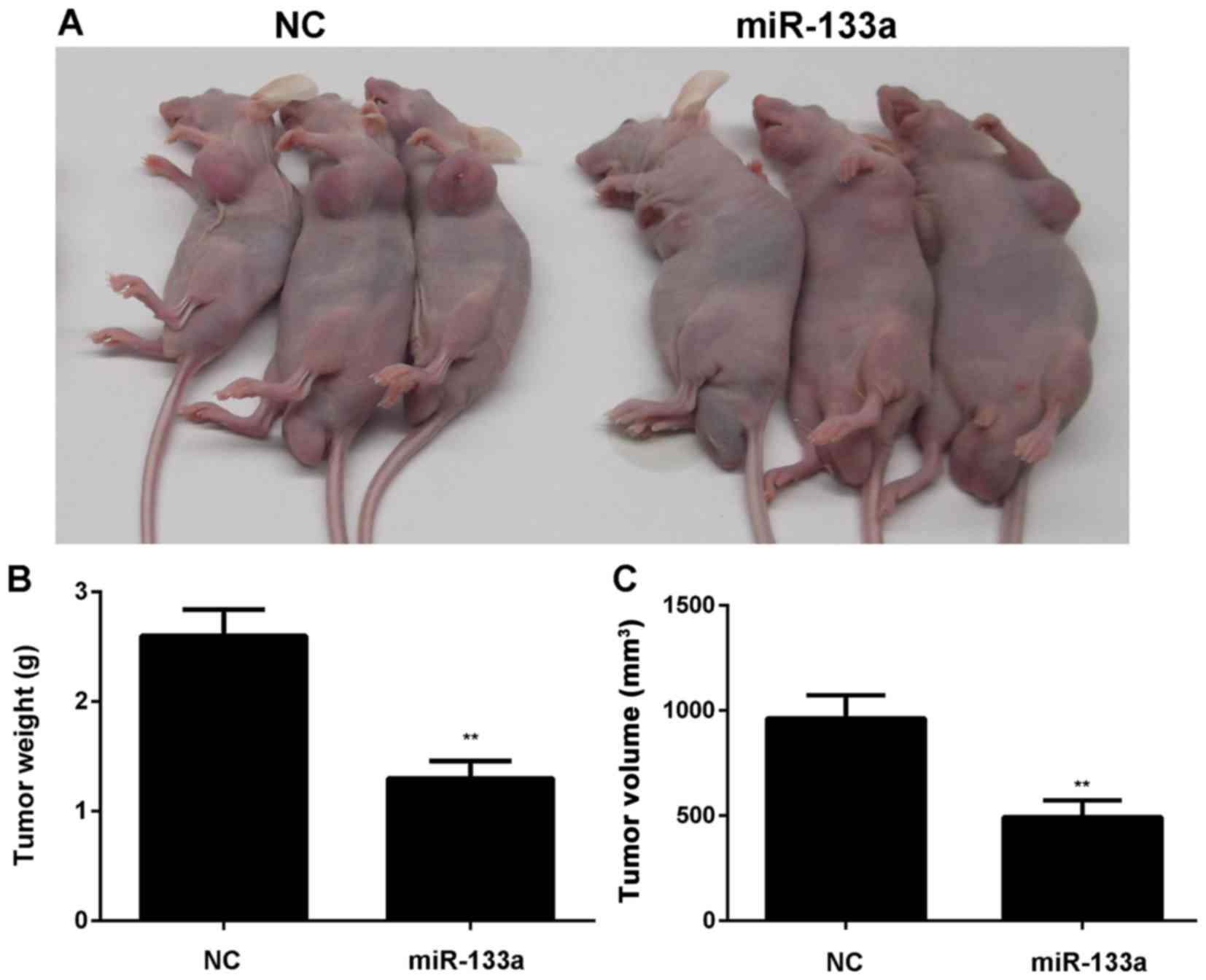
After that, MDA-MB-231 cells stably transfected with
miR-133a or miR-NC lentiviral plasmid were then subcutaneously
implanted into nude mice. The animals were sacrificed 60 days after
implantation. As indicated in Fig.
3, overexpression of miR-133a significantly inhibited the
growth of SK-BR-3 and MDA-MB-231 cells in vivo.
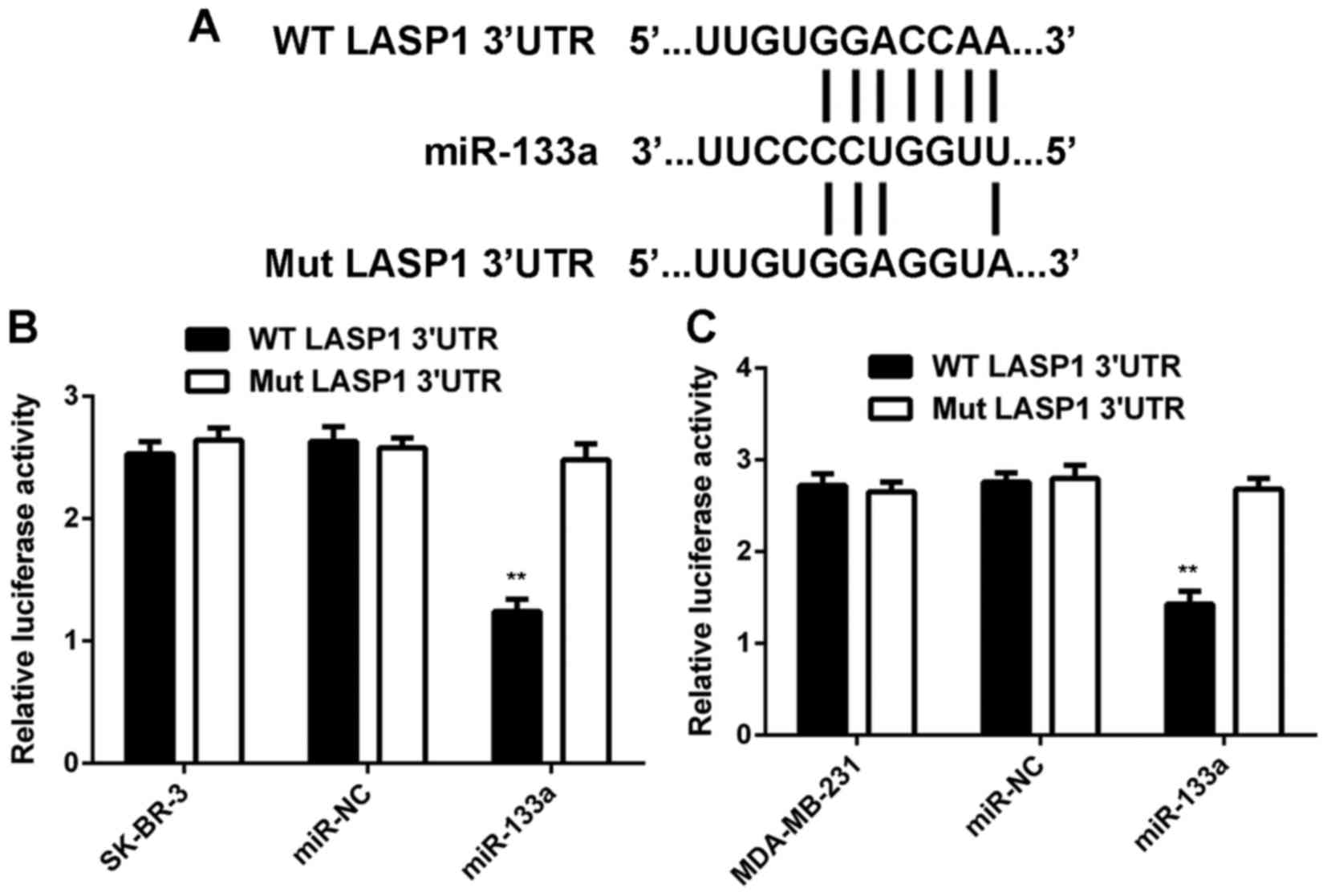
LASP1 is a target gene of miR-133a in
breast cancer cells
As miRs function through regulating the expression
of their target genes, we then studied the potential targets of
miR-133a in breast cancer cells. As shown in Fig. 4A, LASP1 was predicted to be a target
gene of miR-133a. To confirm this prediction, we generated
WT-LASP1-3UTR and Mut-LASP1-3UTR luciferase reporter plasmids
(Fig. 4A). A luciferase reporter
gene assay was conducted in SK-BR-3 and MDA-MB-231 cells. As shown
in Fig. 4B and C, luciferase
activity was significantly reduced in SK-BR-3 and MDA-MB-231 cells
co-transfected with miR-133a mimic and WT-LASP1-3UTR plasmid, but
unchanged in cells co-transfected with miR-133a mimic and
Mut-LASP1-3UTR plasmid. These data indicate that miR-133a can
directly bind to the 3UTR of LASP1 mRNA in breast cancer cells.
As miRs generally regulate negatively the expression
of their target genes, we then investigated the effects of miR-133a
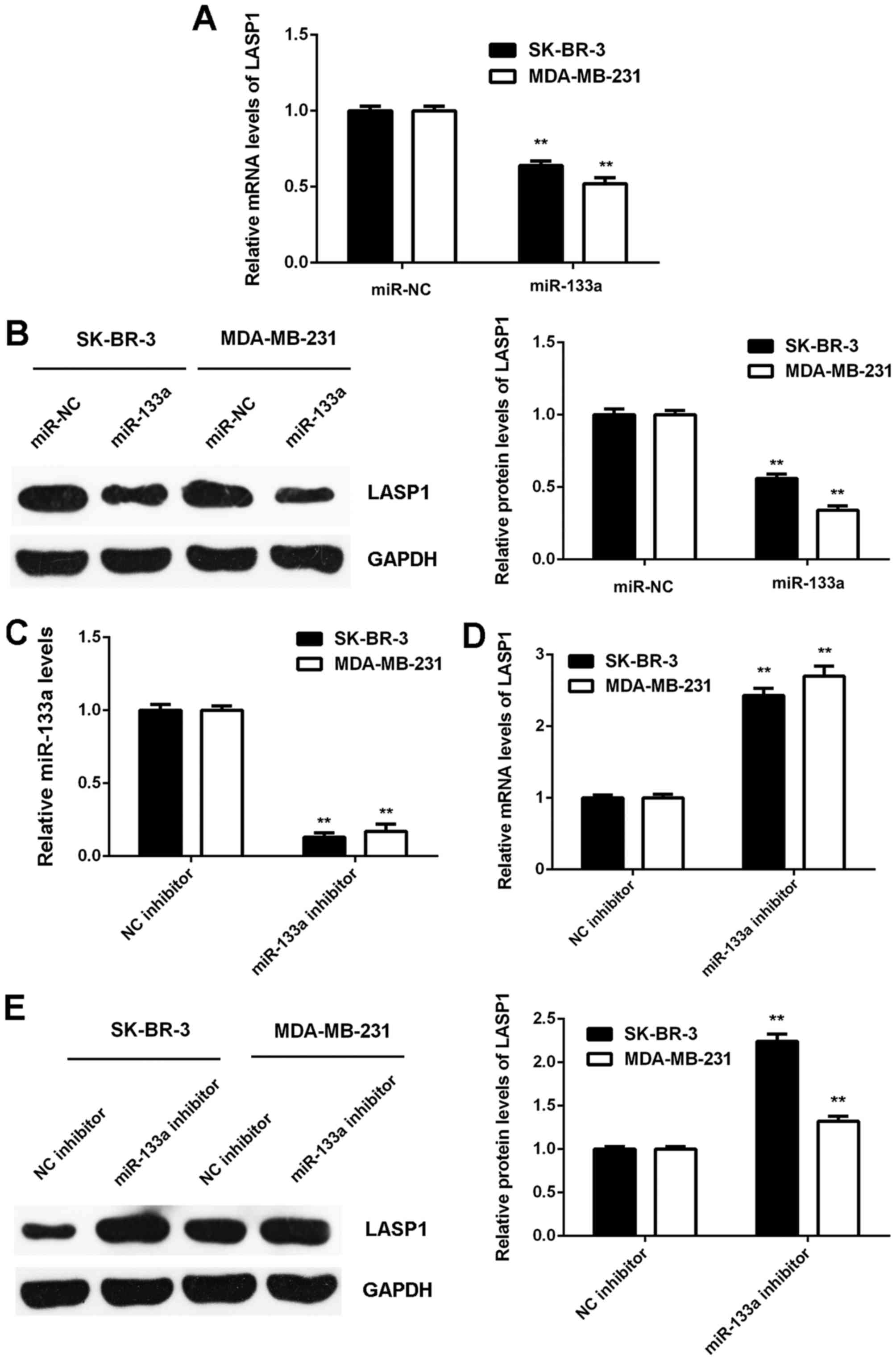
on LASP1 expression in breast cancer cells. We found that the mRNA
and protein expression of LASP1 was significantly downregulated in
SK-BR-3 and MDA-MB-231 cells transfected with miR-133a mimic when
compared to that in cells transfected with miR-NC (Fig. 5A and B). Next, SK-BR-3 and
MDA-MB-231 cells were transfected with NC inhibitor or miR-133a
inhibitor. After transfection, the miR-133a levels were
significantly downregulated in the miR-133a inhibitor group
compared to the NC inhibitor group (Fig. 5C). We then found that the mRNA and
protein expression of LASP1 were significantly increased in the
miR-133a inhibitor group compared with the NC inhibitor group
(Fig. 5D and E). Accordingly, the
expression of LASP1 is negatively mediated by miR-133a in breast
cancer cells.
LASP1 is upregulated in breast
cancer
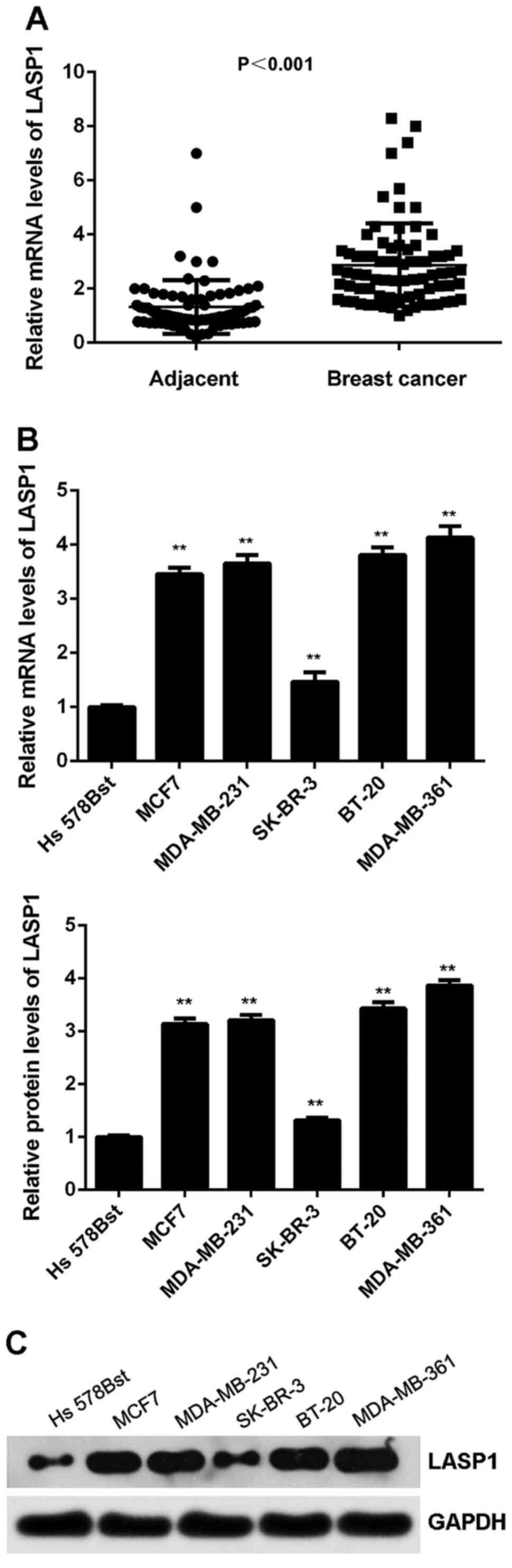
The expression of LASP1 was then examined in breast
cancer. As shown in Fig. 6A, the
mRNA expression of LASP1 was significantly increased in breast
cancer tissues compared with adjacent non-tumour tissues.
Similarly, its expression was also increased in breast cancer cell
lines compared with normal breast Hs 578Bst cells (Fig. 6B and C). We suggest that the
downregulation of miR-133a may contribute to the upregulation of
LASP1. After that, we studied the clinical significance of LASP1
expression in breast cancer. As shown in Table II, high expression of LASP1 was
significantly associated with lymph node metastasis and advanced
clinical stage in breast cancer, suggesting that the upregulation
of LASP1 may contribute to malignant progression in breast
cancer.
| Table II.Association between the LASP1
expression and clinicopathological characteristics in breast
cancer. |
Table II.
Association between the LASP1
expression and clinicopathological characteristics in breast
cancer.
| Variables | N (n=78) | Low expression
(n=37) | High expression
(n=41) | P-value |
|---|
| Age (years) |
|
|
| 1.000 |
|
≤50 | 35 | 17 | 18 |
|
|
>50 | 43 | 20 | 23 |
|
| Tumor size |
|
|
| 0.071 |
|
T1-T2 | 44 | 25 | 19 |
|
|
T3-T4 | 34 | 12 | 22 |
|
| Grade |
|
|
| 0.244 |
| Well
and moderately | 49 | 26 | 23 |
|
|
Poor | 29 | 11 | 18 |
|
| Lymph node
metastasis |
|
|
|
0.022a |
|
Present | 58 | 23 | 35 |
|
|
Absent | 20 | 14 | 6 |
|
| Distant
metastasis |
|
|
|
0.031a |
|
Present | 9 | 1 | 8 |
|
|
Absent | 69 | 36 | 33 |
|
| TNM stage |
|
|
|
0.004b |
|
I–II | 50 | 30 | 20 |
|
|
III–IV | 28 | 7 | 21 |
|
Knockdown of LASP1 inhibits the
proliferation, migration and invasion of breast cancer cells
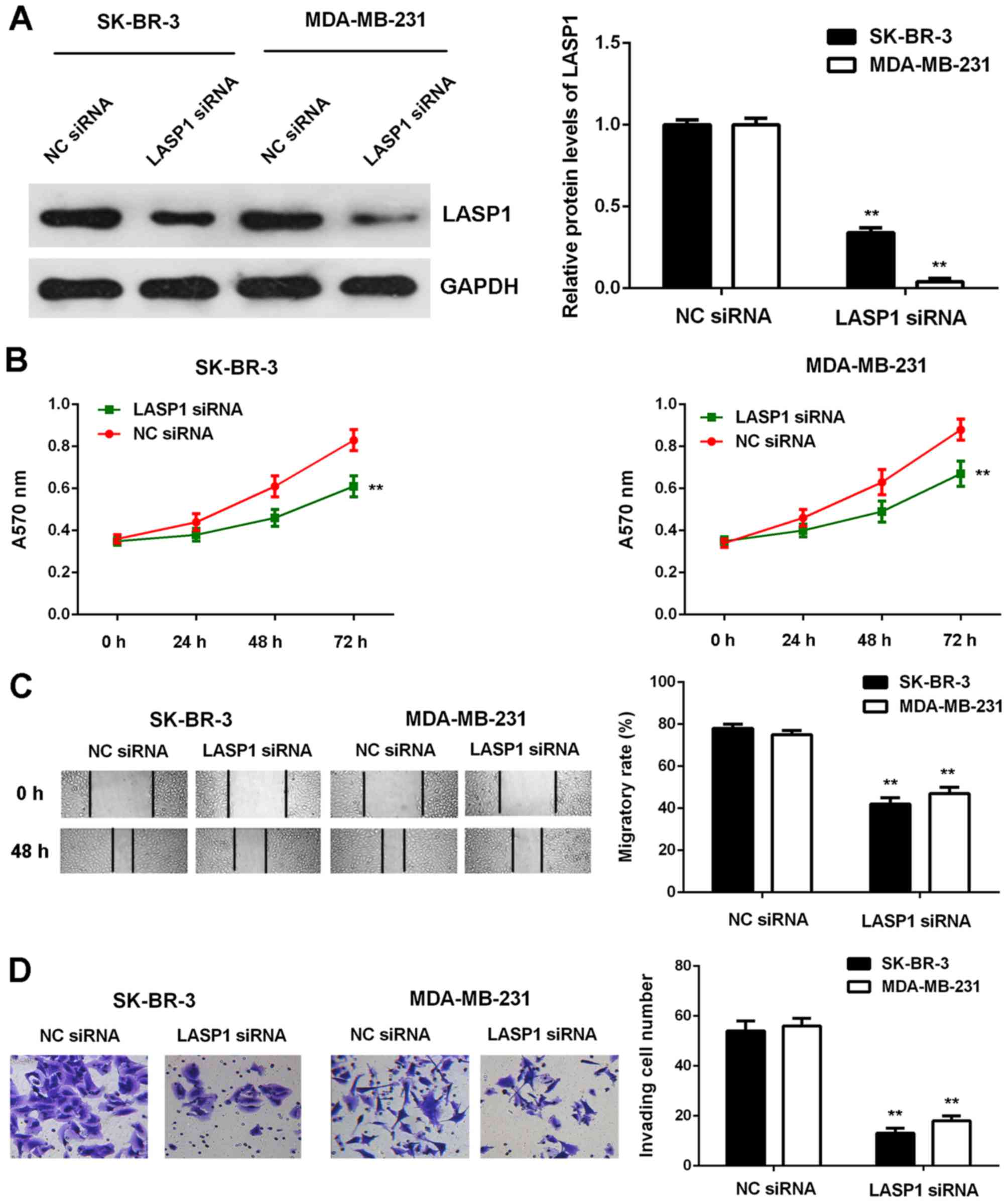
To further clarify the potential role of LASP1 in
breast cancer, SK-BR-3 and MDA-MB-231 cells were transfected with
NC siRNA or LASP1 siRNA. As shown in Fig. 7A, the protein expression of LASP1
was significantly reduced after transfection with LASP1 siRNA when
compared to transfection with NC siRNA. MTT, wound healing and
Transwell assay data further showed that the proliferative,
migratory and invasive capacities of SK-BR-3 and MDA-MB-231 cells
were significantly reduced in the LASP1 siRNA group, when compared
with those of the cells in the NC siRNA group (Fig. 7B-D). These findings suggest that
LASP1 plays a tumour-promoting role in breast cancer cells.
Overexpression of LASP1 attenuates the
inhibitory effects of miR-133a on breast cancer cells
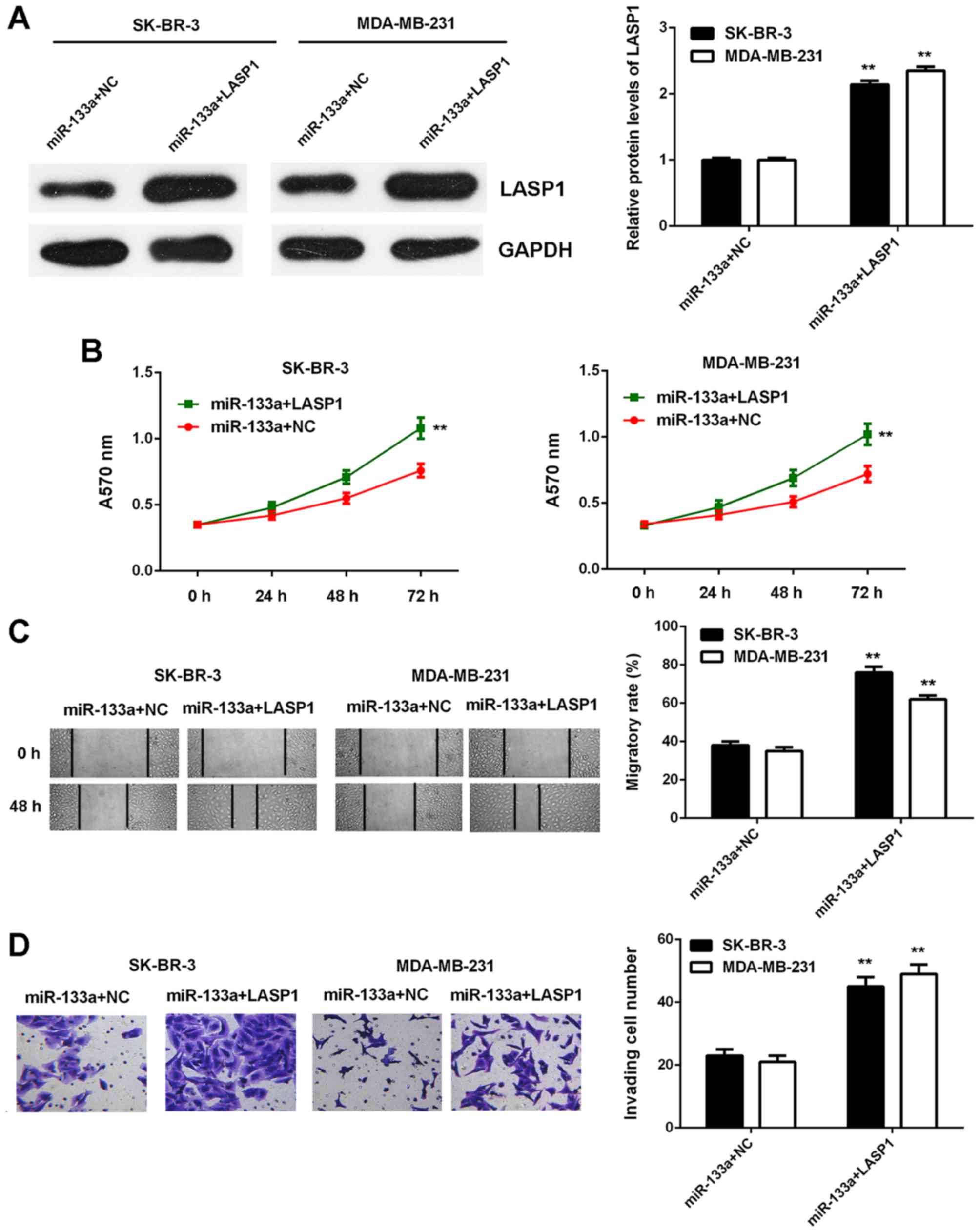
Based on the findings above, we speculated that
LASP1 might act as a downstream effector in the miR-133a-mediated
breast cancer cells. To determine whether this speculation was
correct, miR-133a-overexpressing breast cancer cells were
transfected with blank pcDNA3.1 vector or pcDNA3.1-LASP1 expression
plasmid. After transfection, LASP1 was significantly upregulated in
the miR-133a+LASP1 group compared with in the miR-133a+NC group
(Fig. 8A). We then found that the
proliferative, migratory and invasive capacities of SK-BR-3 and
MDA-MB-231 cells were significantly increased in the miR-133a+LASP1
group compared with the miR-133a+NC group (Fig. 8B-D). These findings indicate that
overexpression of LASP1 impaired the suppressive effects of
miR-133a on the malignant phenotypes of breast cancer cells.
Discussion
The regulatory mechanism of miR-133a underlying the
malignant progression of breast cancer remains largely unknown. In
the present study, we observed that miR-133a was significantly
downregulated in breast cancer, and low expression of miR-133a was
associated with disease progression and poor prognosis in breast
cancer. Overexpression of miR-133a reduced the proliferation,
migration and invasion of breast cancer cells in vitro and
tumour growth in vivo. LASP1 was identified as a target gene
of miR-133a in breast cancer cells, and upregulation of LASP1 was
significantly associated with disease progression in breast cancer.
Knockdown of LASP1 also inhibited the proliferation, migration and
invasion of breast cancers, and overexpression of LASP1 impaired
the suppressive effects of miR-133a upregulation on the malignant
phenotypes of breast cancer cells.
The suppressive role of miR-133a in breast cancer is
gradually revealed (11,14). Wu et al (14) reported that the downregulation of
miR-133a was associated with poor survival of breast cancer
patients and that restoration of miR-133a expression inhibited
breast cancer cell growth and invasion. Cui et al (15) found that miR-133a could inhibit
breast cancer cell proliferation by inducing cell cycle arrest via
the regulation of the epidermal growth factor receptor-mediated Akt
signalling pathway. In this study, we found that miR-133a was
significantly downregulated in breast cancer tissues and cell
lines. Moreover, low miR-133a expression was significantly
associated with lymph node metastasis, advanced clinical stage and
poor prognosis in breast cancer, consistent with a previous study.
Based on our findings and those of others, we suggest that miR-133a
downregulation contributes to the malignant progression of breast
cancer. Indeed, we found that restoration of miR-133a inhibited
breast cancer cell proliferation, migration and invasion in
vitro as well as tumour growth in vivo. Similarly, Ji
et al (11) also reported
that delivery of microbubbles containing miRNA-133a could
effectively inhibit breast cancer growth in vivo.
Accordingly, we suggest that miR-133a may become a promising
molecular candidate for breast cancer treatment. Notably, Chan
et al (21)studied
circulating miR signatures for breast cancer detection and found
that miR-133a was significantly upregulated in the serum of breast
cancer patients. Although miR-133a plays a suppressive role in
breast cancer intracellularly, it is still not known whether it has
antitumour effects in circulating form. Future studies could also
focus on this field, as circulating miRs have only been identified
over the last few years.
We then studied the regulatory mechanism of miR-133a
underlying breast cancer progression. Bioinformatics analysis and
luciferase reporter gene assay data confirmed that LASP1 was a
direct target gene of miR-133a. LASP1 is an important member of the
nebulin family of actin-binding proteins (22). It binds to the actin cytoskeleton at
extensions of the cell membrane depending on cAMP and cGMP
signalling (22). Recently, the
oncogenic role of LASP1 has also been reported (20,23,24).
For instance, LASP1 could induce TGF-beta mediated
epithelial-mesenchymal transition (EMT) in colorectal cancer by
regulating S100A4 expression (25).
Upregulation of LASP1 was strongly correlated with metastatic
dissemination and inferior overall and progression-free survival in
medulloblastoma (26). In this
study, we found that upregulation of LASP1 might contribute to
disease progression and that knockdown of LASP1 significantly
inhibited the proliferation, migration and invasion of breast
cancer cells, suggesting that it may be used as a potential
therapeutic target of this disease. Several other studies have also
reported that nuclear localization and cytosolic overexpression of
LASP1 correlated with advanced tumour size, lymph node metastasis,
and poor prognosis in breast cancer, and silencing of LASP1
inhibited breast cancer cell proliferation and migration (27). Moreover, Endres et al
(4) found that LASP1 could induce
the expression of MMP9 as well as its secretion into the
extracellular matrix, which was responsible for breast cancer cell
migration and invasion. As we found that miR-133a could negatively
regulate the expression of oncogenic LASP1 in breast cancer cells,
we speculated that LASP1 might be involved in the tumour
suppressive functions of miR-133a in breast cancer. Indeed,
overexpression of LASP1 impaired the suppressive effects of
miR-133a on the proliferation, migration and invasion of breast
cancer cells. These findings confirmed our speculation. In fact,
Wang et al (28) also
reported that miR-133a could inhibit tumour growth and metastasis
in colorectal cancer by targeting LASP1 and inhibiting the MAPK
pathway. Therefore, the miR-133a/LASP1 signalling axis may play an
essential role during the malignant progression of different
cancers.
Beside miR-133a, several other miRs have also been
demonstrated to directly target LASP1, and through this mechanism
they play tumour suppressive roles (19,20,23).
For instance, miR-203 inhibits cell proliferation and migration by
targeting LASP1 in triple-negative breast cancer cells (19). miR-218 inhibits gastric cancer cell
proliferation, migration, and invasion and promotes cell apoptosis
by targeting LASP1 (23). miR-133b
inhibits the proliferation, migration, and invasion of
hepatocarcinoma cells via targeting LASP1 (20). Therefore, the present study expands
the understanding of the function of miRs/LASP1 signalling in human
cancers. However, due to lack of molecular subtyping information
from breast cancer patients, we cannot clarify the relationship
between different subtypes of breast cancer and miR-133a-LASP1; we
therefore cannot investigate the molecular mechanisms that exist in
certain subtypes, which must be our next aim.
To the best of our knowledge, the present study is
the first to demonstrate that miR-133a acts as a tumour suppressor
in breast cancer, at least in part by directly targeting LASP1, and
thus suggests that miR-133a may become a promising therapeutic
candidate for the treatment of breast cancer.
References
|
1
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2015. CA Cancer J Clin. 65:5–29. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wu Y, Sarkissyan M and Vadgama JV:
Epithelial-mesenchymal transition and breast cancer. J Clin Med.
5:52016. View Article : Google Scholar :
|
|
4
|
Endres M, Kneitz S, Orth MF, Perera RK,
Zernecke A and Butt E: Regulation of matrix metalloproteinases
(MMPs) expression and secretion in MDA-MB-231 breast cancer cells
by LIM and SH3 protein 1 (LASP1). Oncotarget. 7:64244–64259. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ambros V: The functions of animal
microRNAs. Nature. 431:350–355. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
John B, Enright AJ, Aravin A, Tuschl T,
Sander C and Marks DS: Human MicroRNA targets. PLoS Biol.
2:e3632004. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Croce CM and Calin GA: miRNAs, cancer, and
stem cell division. Cell. 122:6–7. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zheng Y, Lv X, Wang X, Wang B, Shao X,
Huang Y, Shi L, Chen Z, Huang J and Huang P: MiR-181b promotes
chemoresistance in breast cancer by regulating Bim expression.
Oncol Rep. 35:683–690. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Xu X, Zhang Y, Jasper J, Lykken E,
Alexander PB, Markowitz GJ, McDonnell DP, Li QJ and Wang XF:
MiR-148a functions to suppress metastasis and serves as a
prognostic indicator in triple-negative breast cancer. Oncotarget.
7:20381–20394. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ji Y, Han Z, Shao L and Zhao Y: Evaluation
of in vivo antitumor effects of low-frequency ultrasound-mediated
miRNA-133a microbubble delivery in breast cancer. Cancer Med.
5:2534–2543. 2016. View
Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chen G, Fang T, Huang Z, Qi Y, Du S, Di T,
Lei Z, Zhang X and Yan W: MicroRNA-133a inhibits osteosarcoma cells
proliferation and invasion via targeting IGF-1R. Cell Physiol
Biochem. 38:598–608. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zheng K, Liu W, Liu Y, Jiang C and Qian Q:
MicroRNA-133a suppresses colorectal cancer cell invasion by
targeting Fascin1. Oncol Lett. 9:869–874. 2015.PubMed/NCBI
|
|
14
|
Wu ZS, Wang CQ, Xiang R, Liu X, Ye S, Yang
XQ, Zhang GH, Xu XC, Zhu T and Wu Q: Loss of miR-133a expression
associated with poor survival of breast cancer and restoration of
miR-133a expression inhibited breast cancer cell growth and
invasion. BMC Cancer. 12:512012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Cui W, Zhang S, Shan C, Zhou L and Zhou Z:
microRNA-133a regulates the cell cycle and proliferation of breast
cancer cells by targeting epidermal growth factor receptor through
the EGFR/Akt signaling pathway. FEBS J. 280:3962–3974. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lin X, Liu X, Fang Y and Weng X: LIM and
SH3 protein 1 promotes tumor proliferation and metastasis in lung
carcinoma. Oncol Lett. 12:4756–4760. 2016.PubMed/NCBI
|
|
17
|
Takeshita N, Mori M, Kano M, Hoshino I,
Akutsu Y, Hanari N, Yoneyama Y, Ikeda N, Isozaki Y, Maruyama T, et
al: miR-203 inhibits the migration and invasion of esophageal
squamous cell carcinoma by regulating LASP1. Int J Oncol.
41:1653–1661. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hu Z, Cui Y, Zhou Y, Zhou K, Qiao X, Li C
and Wang S: MicroRNA-29a plays a suppressive role in non-small cell
lung cancer cells via targeting LASP1. Onco Targets Ther.
9:6999–7009. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang C, Zheng X, Shen C and Shi Y:
MicroRNA-203 suppresses cell proliferation and migration by
targeting BIRC5 and LASP1 in human triple-negative breast cancer
cells. J Exp Clin Cancer Res. 31:582012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Li H, Xiang Z, Liu Y, Xu B and Tang J:
MicroRNA-133b inhibits proliferation, cellular migration, and
invasion via targeting LASP1 in hepatocarcinoma cells. Oncol Res.
25:1269–1282. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Chan M, Liaw CS, Ji SM, Tan HH, Wong CY,
Thike AA, Tan PH, Ho GH and Lee AS: Identification of circulating
microRNA signatures for breast cancer detection. Clin Cancer Res.
19:4477–4487. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Orth MF, Cazes A, Butt E and Grunewald TG:
An update on the LIM and SH3 domain protein 1 (LASP1): A versatile
structural, signaling, and biomarker protein. Oncotarget. 6:26–42.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang LL, Wang L, Wang XY, Shang D, Yin SJ,
Sun LL and Ji HB: MicroRNA-218 inhibits the proliferation,
migration, and invasion and promotes apoptosis of gastric cancer
cells by targeting LASP1. Tumour Biol. 37:15241–15252. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Du YY, Zhao LM, Chen L, Sang MX, Li J, Ma
M and Liu JF: The tumor-suppressive function of miR-1 by targeting
LASP1 and TAGLN2 in esophageal squamous cell carcinoma. J
Gastroenterol Hepatol. 31:384–393. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wang H, Shi J, Luo Y, Liao Q, Niu Y, Zhang
F, Shao Z, Ding Y and Zhao L: LIM and SH3 protein 1 induces
TGFβ-mediated epithelial-mesenchymal transition in human colorectal
cancer by regulating S100A4 expression. Clin Cancer Res.
20:5835–5847. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Traenka C, Remke M, Korshunov A, Bender S,
Hielscher T, Northcott PA, Witt H, Ryzhova M, Felsberg J, Benner A,
et al: Role of LIM and SH3 protein 1 (LASP1) in the metastatic
dissemination of medulloblastoma. Cancer Res. 70:8003–8014. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Frietsch JJ, Grunewald TG, Jasper S,
Kammerer U, Herterich S, Kapp M, Honig A and Butt E: Nuclear
localisation of LASP-1 correlates with poor long-term survival in
female breast cancer. Br J Cancer. 102:1645–1653. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wang H, An H, Wang B, Liao Q, Li W, Jin X,
Cui S, Zhang Y, Ding Y and Zhao L: miR-133a represses tumour growth
and metastasis in colorectal cancer by targeting LIM and SH3
protein 1 and inhibiting the MAPK pathway. Eur J Cancer.
49:3924–3935. 2013. View Article : Google Scholar : PubMed/NCBI
|