Introduction
Cancer of the oral cavity (oral cancer) is the 11th
most frequent malignancy in the world, especially in developing
countries, ranking ninth in the total number of incidences
(1). Ninety-percent of oral cancers
histologically originate in squamous cells (2). The incidence and mortality of this
cancer exhibits variability resulting from: geographic location,
gender, as well as occurrence site (lips, tongue, gingiva, mouth
floor, parotid and saliva glands) (http://globocan.iarc.fr/Pages/summary_table_pop_sel.aspx).
Although with the development of iatrotechniques, the mortality of
oral squamous cell carcinoma (OSCC) has remained relatively
unchanged. Worse still, the age of oral cancer patients is trending
younger and younger. The overall prognosis of this cancer is poor;
the survival rate is roughly 55–65% (3). This is probably due to late diagnosis.
An early diagnosis is key in decreasing the mortality of oral
cancer, which subsequently improves the overall survival and
prognosis. Therefore, seeking a novel biomarker and therapeutic
target is of the utmost importance.
Circular RNAs (circRNAs) are special types of
non-coding RNAs characterized by their circular shape resulting
from covalently closed continuous loops, widely expressed in cells
that regulate the expression of relative genes in multiple pathways
(4,5). CircRNAs were initially thought to
originate due to a failure in alternative splicing owing to their
low expression levels (6). Since
next-generation sequencing technologies and bioinformatics analysis
have developped, the regulatory function of circRNAs in eukaryotic
cells is beginning to increasingly attract emphasis. As particular
non-coding RNAs, circRNAs are widely expressed, highly conserved
and stable in the cytoplasm (RNase R resistant) (6). With the deepening of research, two
circRNAs (CDR1as and Sry) have been revealed to be miRNA sponges
regulating the expression of relevant genes (7,8).
Moreover, they were also found to be regulators of alternative
splicing (9), and transcription
factors (10) and to encode for
proteins (11,12). CircRNAs participate in several
pathological states, including neurological diseases (13,14),
hematopathy (15), cartilage
degradation (16), Hirschsprung's
disease (17) and multiple cancers,
which may serve as promising diagnostic biomarkers and therapeutic
targets. It is known that circRNAs could compete for binding to
cancer-related miRNAs. Multiple circRNA/miRNA/mRNA axes in
cancer-associated pathways have been investigated in colorectal
cancer (18,19), pancreatic cancer (20), bladder carcinoma (21), esophageal squamous cell carcinoma
(ESCC) (22), glioblastoma
(23), cervical cancer (24), chronic lymphocytic leukemia (CLL)
(25), non-small cell lung cancer
(NSCLC) (26), cholangiocarcinoma
(27), ovarian cancer (28) and hepatocellular carcinoma (HCC)
(29,30), with both agonist and antagonist
effects on carcinogenesis. However, whether circRNAs harbor miRNAs
with regulatory roles in OSCC is still unknown.
MicroRNAs (miRNAs) are one of the large families of
non-coding RNAs with small, endogenous, and evolutionarily
conserved features, (approximately 18–24 nucleotides) which are
involved in regulating several essential, cellular and functional
processes such as cell differentiation, migration, growth,
proliferation, apoptosis, metabolism and tumorigenesis (31–33).
It is well known that miRNAs are known to play key roles in most
types of cancer and function as oncogenes or tumor suppressors in
upregulation or downregulation processes respectively (31). miRNAs have the potential to be used
as prognostic and diagnostic biomarkers. Generally, to be a
negative regulator, miRNAs bind to the 3′-untranslated region
(3′-UTR) of their target mRNA based on the principle of
complementary base pairing and inhibit translation (34). miRNAs play crucial roles in OSCC
initiation and development (35).
In this study, we investigated the molecular
mechanisms underlying the circRNAs in OSCC. We built an apoptosis
model by stimulating CAL-27 cells with TNF-α and acquired a
differentially expressed circRNA profile from the apoptosis model
and normal CAL-27 cells using high-throughput microarray. Several
significant differentially expressed circRNAs were further
confirmed with qRT-RCR and we found that the circRNA
has_circ_100721 (circDOCK1) was markedly overexpressed and the
knockdown of the expression of circDOCK1 led to the increase of
apoptosis. According to a set of bioinformatics approaches, we
constructed a circRNA/miRNA/gene network and determined that
circDOCK1 interacted with miR-196a-5p by competing with BIRC3,
eventually regulating cell apoptosis. Finally, we explored the
functions of circDOCK1 in OSCC in vivo and in vitro.
Hence, we concluded that circDOCK1 suppressed cell apoptosis via
inhibition of miR-196a-5p by targeting BIRC3 in OSCC.
Materials and methods
Ethics statement and tissue
samples
This study was approved and supervised by The Ethics
Committee of Guangzhou Medical University. All the experiments were
carried out in accordance with relevant guidelines and regulations.
All oral carcinoma tissues and adjacent normal tissues were
collected from patients who had undergone surgery at the Cancer
Center of Guangzhou Medical University and The First Affiliated
Hospital of Guangzhou Medical University from 2015 to 2016. All
carcinoma and paired normal tissues were ten groups each, which
were confirmed by pathological detection. The clinical and
pathological characteristics for each patient were noted before and
after surgery. The patients were made aware of the research
purposes and informed consent was given.
Cell culture
The human OSCC cell lines CAL-27, SCC-9 and SCC-25
were purchased from ATCC (American Type Culture Collection,
Manassas, VA, USA). Human normal oral epithelial keratinocytes
(HOK) were purchased from ScienCell Research Laboratories (San
Diego, CA, USA). HOK was cultured in Dulbecco's modified Eagle's
medium (DMEM) (Gibco, Carlsbad, CA, USA) containing with 10% fetal
bovine serum (FBS) (Gibco). The other cell lines were cultured in
Dulbecco's modified Eagle's medium: nutrient Mixture F-12
(DMEM/F12) (Gibco) with 10% FBS. All cell lines were incubated at
37°C in a humidified chamber containing 5% CO2.
Apoptosis model and flow cytometry
(FCM)
OSCC cell line CAL-27 cells were stimulated with 100
ng/ml recombinant human TNF-α (PeproTech, Rocky Hill, NJ, USA) for
48 h. The FITC Annexin V Apoptosis Detection Kit I (BD Biosciences,
Franklin Lakes, NJ, USA) was used according to the FITC Annexin V
Staining Protocol to detect apoptotic cells 48 h after being
treating with TNF-α. First, the cells were collected and washed
twice with cold PBS and then resuspended in 1X binding buffer at a
concentration of 1×106 cells/ml. One hundred microliters
of the solution (1×105 cells) was transferred to a 5 ml
culture tube. FITC Annexin V (5 µl) and propidium iodide (PI; 5 µl)
staining solutions were added to the cells. The cells were gently
cycled in a vortex and incubated for 15 min at RT (25°C) in the
dark. Then 400 µl of 1X binding buffer were added to each tube and
the cells were analyzed by flow cytometry (BD Biosciences) within 1
h.
CircRNA Microarray
An apoptosis model and a negative control group,
oral carcinoma tissues and adjacent normal tissues were used to
detect the expression of circRNAs by CircRNA Microarray. Total RNA
from each sample was quantified using the NanoDrop ND-1000. The
sample preparation and microarray hybridization were performed
based on the standard protocols of Arraystar Inc. (Rockville, MD,
USA). Briefly, total RNA from each sample was amplified and
transcribed into fluorescent cRNA utilizing a random primer
according to Arraystar's Super RNA Labeling protocol (Arraystar
Inc.). The labeled cRNAs were hybridized onto the Arraystar Human
circRNA Microarray (8×15K, Arraystar Inc.). After having washed the
slides, the arrays were scanned using the Agilent DNA Microarray
Scanner G2505C. Agilent Feature Extraction software (version
11.0.1.1) was used to analyze acquired array images while quantile
normalization and subsequent data processing were performed using
the R software package. Differentially expressed circRNAs with
statistical significance between two groups were identified through
volcano plot filtering and these two groups were also identified
through fold change filtering. Hierarchical clustering was
performed to reveal the distinguishable circRNA expression patterns
among samples. The microarray hybridization and the collection of
data were performed by KangChen Bio-tech, Shanghai, China.
Quantitative real-time PCR and circRNA
sequencing
Total RNA was extracted from OSCC cell lines and
oral cancer samples using TRIzol Reagent (Invitrogen, Carlsbad, CA,
USA) and quantified using NanoDrop 2000 (Thermo Scientific,
Carlsbad, CA, USA). After RNA extraction, SuperScript®
III Reverse Transcriptase (Invitrogen) was used for
reverse-transcription to cDNA following the manufacturer's
protocol. The RT products were used as templates for amplification
using the SYBR® Premix Ex Taq™ II PCR amplification
reagent (Takara, Shiga, Japan). Specific divergent primers were
designed to amplify the circular transcripts. The reaction system
of qRT-PCR was performed in 25 µl of reaction volume, including 2
µl of cDNA, 12.5 µl 2X SYBR Premix Ex Taq II, 0.5 µl of forward
primer (10 µM), 0.5 µl of reverse primer (10 µM) and 9.5 µl of
double distilled water. The reaction was set at 95°C for 30 sec for
pre-denaturation, then at 95°C for 5 sec, at 60°C for 30 sec and at
72°C for 30 sec for 39 cycles. Reactions for each sample were
performed in triplicate and the relative levels of target genes
were calculated using the 2−∆∆Ct method and normalized
to GAPDH. The products of qRT-PCR were purified using Agencourt
AMPure XP PCR purification kit from Beckman Coulter Inc. (Brea, CA,
USA). Direct PCR product Sanger sequencing was performed by LGC
Genomics (Berlin, Germany) Ready2 Run services. The forward primer
for circDOCK1 was provided for sequencing the product of circDOCK1.
All the primer sequences of the genes used in this study are shown
in Table I.
 | Table I.All the primer sequences of the genes
used for this study. |
Table I.
All the primer sequences of the genes
used for this study.
| Genes | Primer sequence
(5′-3′) |
|---|
| GAPDH | F:
GCACCGTCAAGGCTGAGAAC |
|
| R:
TGGTGAAGACGCCAGTGGA |
| circDOCK1 | F:
GAAATCGTCCACAGTGACCT |
|
| R:
CACAGTGTCTCCGATCTGTAAA |
| circRNA_101531 | F:
TGCAGGGCGTCATCAAAACC |
|
| R:
CACACACCTGTTCTCCTGTGG |
| circRNA_101315 | F:
AGATGCGCAGGCTCAACATA |
|
| R:
CAATGGATTGTTCCTTGGCTGT |
| circRNA_104313 | F:
GCTACCATTTTGGCTGTTTGGA |
|
| R:
TCTTCAGCCTTGGGCTTCTTTT |
| circRNA_101226 | F:
GCCTGTGTATATCCCAGTTCCT |
|
| R:
TTTGGTCTGCATGTGAGGTTT |
| circRNA_104838 | F:
TTCCATAAATGGGGTCGAG |
|
| R:
TTGAGTGGTCCAAAATCTGC |
| circRNA_102765 | F:
AAGCCTTCACCTGCATCCAA |
|
| R:
AGACGTCTCTCAGTGACAAAG |
| circRNA_100790 | F:
CTGCGTCTCAGCCTCAAGTAT |
|
| R:
TGGTTCCTTCTGTGGTCGTT |
| circRNA_102367 | F:
TGGAAGAATTGCTGTGCTGTT |
|
| R:
TGCACCAAGATTTCCTGTTCG |
| circRNA_102956 | F:
GTTTTGGGCGTGCTAGAGGT |
|
| R:
GCTCGAATTCCTCCATGTCCA |
| BIRC3 | F:
CAACAGATCTGGCAAAAGCA |
|
| R:
TTGCTCAATTTTCCACCACA |
| FOXO1 | F:
AAGAGCGTGCCCTACTTCAA |
|
| R:
CTGTTGTTGTCCATGGATGC |
| MDM2 | F:
CATTGTCCATGGCAAAACAG |
|
| R:
GGCAGGGCTTATTCCTTTT |
| DOCK1 | F:
TCGGACGCCGTACTCTTTCTC |
|
| R:
CTGATTGCTTGGCAGATTCCT |
| miR-196a-5p | F:
TAGGTAGTTTCATGTTGTTGG |
| miR-138-2-3p | F:
CTATTTCACGACACCAGGGTT |
RNA interference, miRNA mimic and
transfection
The small interfering RNAs (siRNAs) (RiboBio,
Guangzhou, China) were designed for the junction of circDOCK1 and
the mimic was designed for miR-196a-5p. The concentration of the
mimic for miR-196a-5p, the siRNA for circDOCK1 and the siRNA for
the negative control was 50 nM. Transfection of the siRNA and mimic
was conducted with GenMute™ siRNA Transfection Reagent (SignaGen
Laboratories, Rockville, MD, USA) according to the standard
protocol. Before transfection, the complex which included 100 µl 1X
GenMute™ Transfection Buffer, 5 µl siRNA or mimic and 4 µl GenMute™
Reagent was incubated for 30 min at room temperature, and then the
above mixture was added to OSCC cell lines with 1 ml DMEM/F12
growth medium in 6-well plates. The cells were cultured in a
humidified atmosphere with 5% CO2 at 37°C. After
transfection for 48 h, the levels of circDOCK1 and miR-196a-5p were
assessed by qRT-PCR. All the sequence of siRNAs and mimics are
shown in Tables II and III.
| Table II.The sequence of siRNAs for
circDOCK1. |
Table II.
The sequence of siRNAs for
circDOCK1.
| siRNA | Target sequence
(5′-3′) |
|---|
| siRNA 1 |
CTTCACACAGCATGCTTTT |
| siRNA 2 |
TCACACAGCATGCTTTTTA |
| siRNA 3 |
CACACAGCATGCTTTTTAT |
| Table III.The sequence of miR-196a-5p
mimics. |
Table III.
The sequence of miR-196a-5p
mimics.
| miRNA mimics | Primer sequence
(5′-3′) |
|---|
| miR-196a-5p | F:
UAGGUAGUUUCAUGUUGUUGGG |
|
| R:
CCCAACAACAUGAAACUACCUA |
Prediction for circRNA/miRNA/mRNA
pathways
The relationship between circRNAs and miRNAs was
predicted by miRNA target prediction software based on miRanda and
TargetScan. Also, the circRNA/microRNA interaction was speculated
using Circinteractome according to the name of circRNAs and miRDB
according to the sequence of circRNAs. Through combination of the
three methods the capability of seed match sequences was ranked.
Using the Cytoscape 3.4.0, the graph of circRNA/miRNA network was
drawn. The top 15 miRNA pathways were predicted based on DIANA
TOOLS-miRPath v.3 and the cancer-related pathways and
apoptosis-related pathways of the top 15 miRNAs were selected. The
target genes were acquired based on Database for Annotation,
Visualization and Integrated Discovery (DAVID). Finally, several
cancer-related pathways and apoptosis-related pathways associating
with circRNAs, miRNAs and target genes were constructed.
Western blotting
The total protein of samples was extracted using
RIPA Lysis Buffer (Beyotime, China, which contained PMSF (100:1)
according to the standard procedure. The protein samples were
loaded onto 10% SDS PAGE and separated. Then, they were transferred
to a PVDF membrane. The membrane was blocked for 2 h with 0.5% skim
milk powder. The target proteins on the membranes were incubated
with the relative primary antibodies β-actin (1:4,000; cat. no.
ab8227) and BIRC3 (1:1,000; cat no. ab32059) (both from Abcam,
Cambridge, UK) for 2 h on table concentrator at 4°C. After being
washed three times, the membranes were sequentially incubated with
the HRP conjugated secondary antibody anti-Rabbit IgG H&L (HRP)
(1:4,000; cat. no. ab205718) (Abcam) for 1 h at RT and ECL
detection followed.
Statistical analysis
All of the experiments were repeated three times
independently. Data are expressed as the mean ± standard deviation
(SD). Student's t-test (two-tailed) was applied for data analysis.
The values P<0.05 indicated a statistically significant
difference.
Results
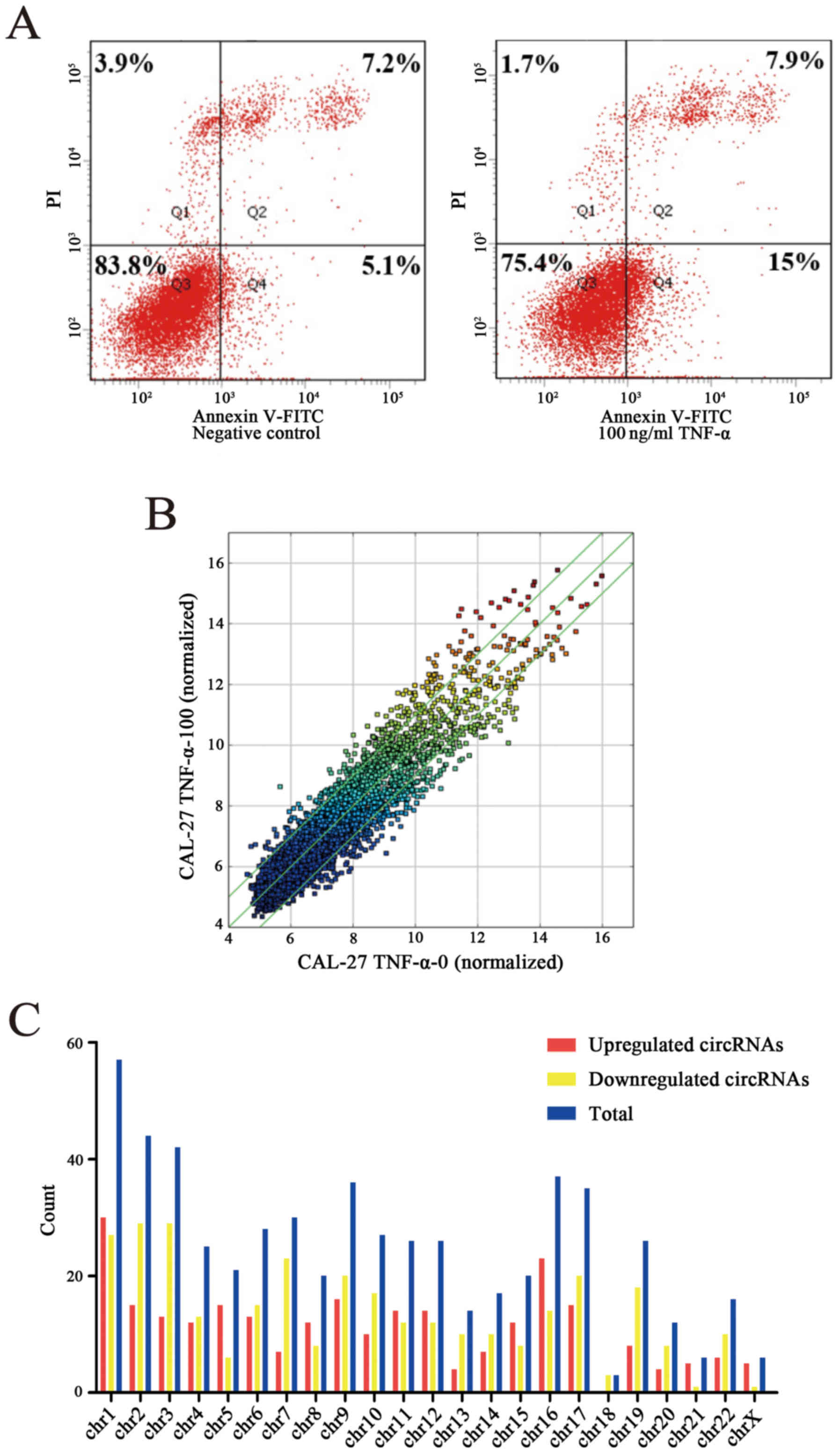
Construction of a cell apoptosis model
and its differentially expressed circRNA profiles
OSCC cell line CAL-27 was stimulated with 100 ng/ml
recombinant human TNF-α and cultured for 48 h. The apoptosis rate
of the apoptosis group and the negative group were detected through
FCM and the apoptosis rate of the apoptosis model was increased by
10.6% compared to the negative control (Fig. 1A). Following construction of the
apoptosis model the circRNA profile of the model was detected by
means of high throughput microarray assay. According to the fold
change (≥2.0), 628 circRNAs were differentially expressed, among
which 287 circRNAs were upregulated while 341 circRNAs were
downregulated (Fig. 1B). By
analyzing the data of differentially expressed circRNAs, we
classified their chromosomal origin (Fig. 1C) and determined that most of the
circRNAs came from the protein coding exons (Fig. 1D). Furthermore, we listed the top
twenty upregulated and downregulated circRNAs in line with the fold
changes of the expression of circRNAs (Table IV). Moreover, hierarchical
clustering was drawn to reveal the differentially expressed
circRNAs between the apoptosis model and the negative control
(Fig. 1E and F). All of the data
revealed that the expression of circRNAs in the apoptosis group was
different from that which was matched in the negative group.
| Table IV.Top 20 upregulated and downregulated
circRNAs were chosen in line with the fold changes of the
expression of circRNAs. |
Table IV.
Top 20 upregulated and downregulated
circRNAs were chosen in line with the fold changes of the
expression of circRNAs.
| Upregulated
circRNAs | Downregulated
circRNAs |
|---|
|
|
|---|
| CircRNA ID | CircRNA type | Chrom | Fold change | GeneSymbol | CircRNA ID | CircRNA type | Chrom | Fold change | GeneSymbol |
|---|
|
hsa_circRNA_100445 | Exonic | chr1 | 8.0656065 | TATDN3 |
hsa_circRNA_101531 | Exonic | chr15 | −7.5435655 | ALDH1A2 |
|
hsa_circRNA_103205 | Exonic | chr22 | 7.8829886 | FBXO7 |
hsa_circRNA_101315 | Exonic | chr14 | −6.1496856 | METTL3 |
|
hsa_circRNA_104650 | Exonic | chr8 | 7.2938337 | KIAA1429 |
hsa_circRNA_104313 | Exonic | chr7 | −5.1219124 | PHF14 |
|
hsa_circRNA_101906 | Exonic | chr16 | 5.4328383 | ANKRD11 |
hsa_circRNA_101226 | Exonic | chr13 | −4.9973297 | ZMYM2 |
|
hsa_circRNA_102895 | Exonic | chr2 | 5.3716472 | BMPR2 |
hsa_circRNA_102765 | Exonic | chr2 | −4.7427653 | SFXN5 |
|
hsa_circRNA_103108 | Exonic | chr21 | 5.0335912 | BC048201 |
hsa_circRNA_100790 | Exonic | chr11 | −4.7006624 | CAPRIN1 |
|
hsa_circRNA_104833 | Exonic | chr9 | 4.8887767 | C9orf3 |
hsa_circRNA_102367 | Exonic | chr18 | −4.6127982 | DYM |
|
hsa_circRNA_102991 | Exonic | chr20 | 4.875669 | TMEM230 |
hsa_circRNA_102956 | Exonic | chr2 | −4.2670317 | ILKAP |
|
hsa_circRNA_103951 | Exonic | chr5 | 4.7518169 | FAM13B |
hsa_circRNA_100721 | Exonic | chr10 | −4.1949433 | DOCK1 |
|
hsa_circRNA_104855 | Exonic | chr9 | 4.7120789 | TMEM245 |
hsa_circRNA_104838 | Exonic | chr9 | −4.1273329 | CDC14B |
|
hsa_circRNA_101141 | Exonic | chr12 | 4.5303581 | ANAPC7 |
hsa_circRNA_103563 | Exonic | chr3 | −4.1234249 | DLG1 |
|
hsa_circRNA_104040 | Exonic | chr6 | 4.5210355 | DUSP22 |
hsa_circRNA_102651 | Exonic | chr2 | −4.0871929 | ZNF512 |
|
hsa_circRNA_104839 | Exonic | chr9 | 4.4748769 | CDC14B |
hsa_circRNA_101695 | Exonic | chr16 | −4.0480481 | NAGPA |
|
hsa_circRNA_101287 | Exonic | chr13 | 4.3717281 | UGGT2 |
hsa_circRNA_104356 | Exonic | chr7 | −3.9404514 | POLR2J4 |
|
hsa_circRNA_100033 | Exonic | chr1 | 4.3075694 | RERE |
hsa_circRNA_100688 | Exonic | chr10 | −3.9125353 | ATRNL1 |
|
hsa_circRNA_102587 | Exonic | chr19 | 4.30598 | LIG1 |
hsa_circRNA_400068 | intronic | chr22 | −3.8772576 | RANBP1 |
|
hsa_circRNA_101850 | Exonic | chr16 | 4.1911027 | CIRH1A |
hsa_circRNA_102205 | Exonic | chr17 | −3.8669576 | TNRC6C |
|
hsa_circRNA_103360 | Exonic | chr3 | 4.0908172 | SMARCC1 |
hsa_circRNA_101055 | Exonic | chr12 | −3.8643985 | CERS5 |
|
hsa_circRNA_101115 | Exonic | chr12 | 3.9659427 | METAP2 |
hsa_circRNA_101747 | Exonic | chr16 | −3.8581905 | C16orf62 |
|
hsa_circRNA_101555 | Exonic | chr15 | 3.897013 | CSNK1G1 |
hsa_circRNA_100117 | Exonic | chr1 | −3.8552525 | EYA3 |
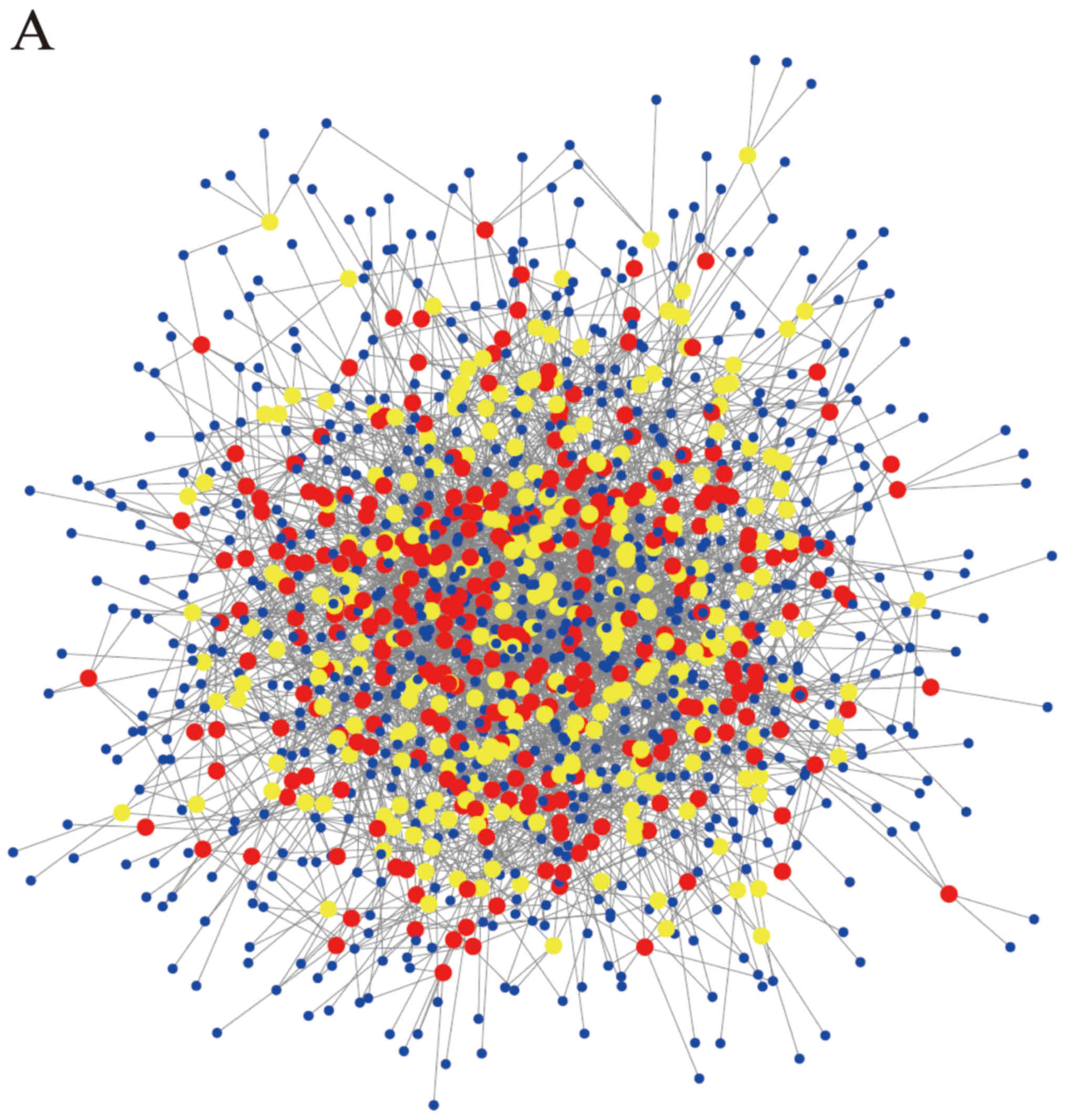
Construction of the circRNA/miRNA
connection network
According to the principle of complementary base
pairing and using the prediction software based on miRanda,
TargetScan and websites Circinteractome and miRDB, we found
hundreds of relative miRNAs targeting the differentially expressed
circRNAs. The top five candidate miRNAs out of a total of 628
circRNAs were selected in accordance with the strength of their
connection. The circRNA/miRNA network was depicted using Cytoscape
(Fig. 2A). In addition to the whole
data, the network of top 20 upregulated and downregulated circRNAs
and their candidate miRNAs was also depicted (Fig. 2B).
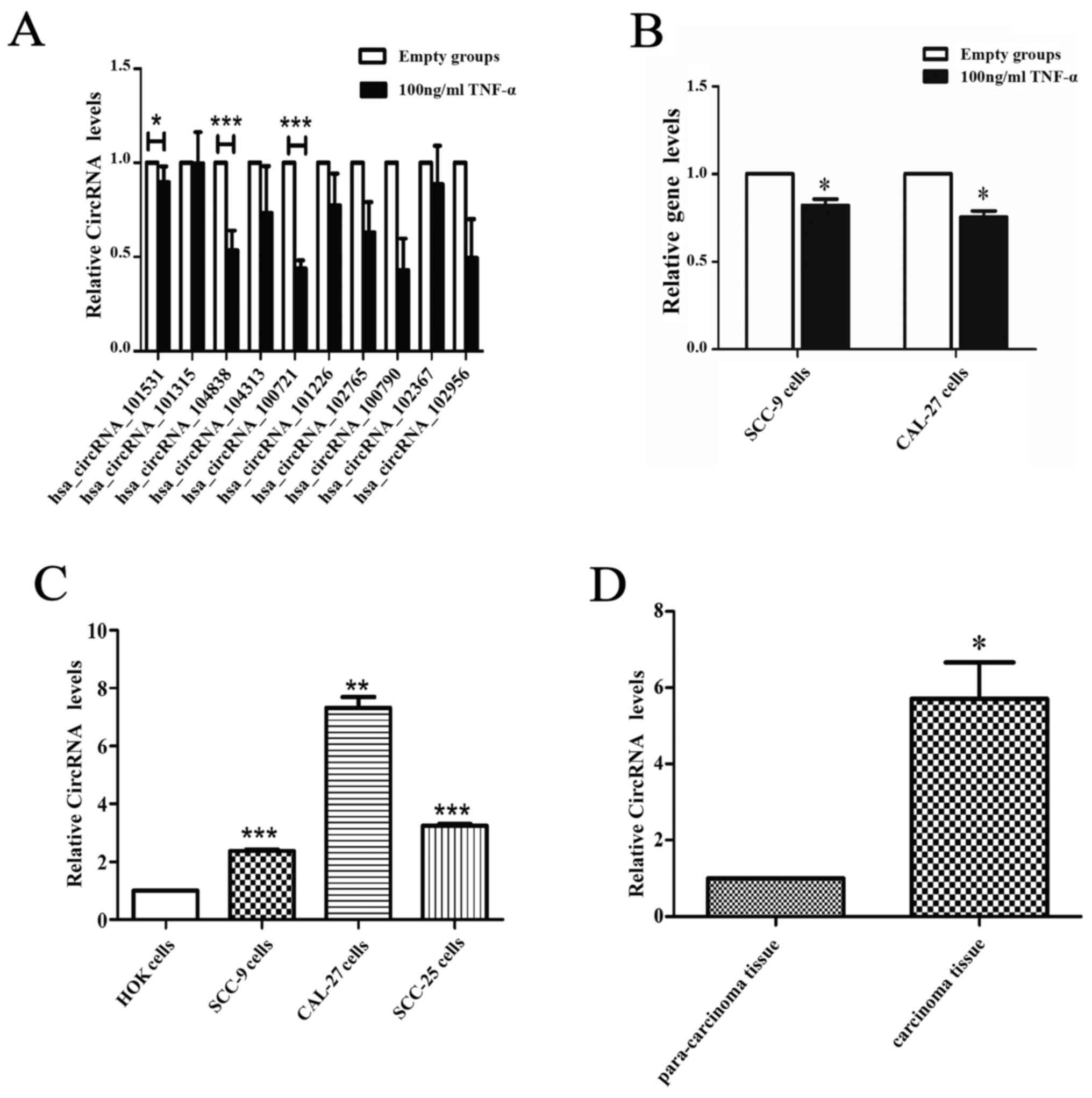
Validation of the differentially
expressed circRNAs in the apoptosis model and OSCC tissue by
qRT-PCR and sequencing
Ten circRNAs were selected to confirm the
differential expression between the apoptosis model and the
negative group via qRT-PCR. The results revealed, two circRNAs,
circCDC14B (hsa_circ_104838) and circDOCK1 (hsa_circ_100721) that
were significantly downregulated in the apoptosis group (Fig. 3A). However, the expression levels of
linear DOCK1 were slightly affected by TNF-α in SCC-9 and CAL-27
cells (Fig. 3B). To further confirm
the significance of these two circRNAs, their expression in OSCC
cell lines (CAL-27, SCC-9, SCC-25) and normal oral epithelial cells
(HOK), OSCC tissue and para-carcinoma tissue (ten groups) were
ascertained by qRT-PCR. Furthermore, the circRNA profiles of one of
the OSCC tissue groups compared with the para-carcinoma tissue was
detected via high throughput microarray assay. The results revealed
that circDOCK1 was distinctly upregulated in OSCC cell lines and
OSCC tissue (Fig. 3C and D). The
sequencing of circDOCK1 confirmed its presence in OSCC (Fig. 3E). The data indicated that
microarray analysis was consistent with the qRT-PCR results
regarding the expression levels of circDOCK1 (Fig. 3F). Consequently, circDOCK1 was
chosen to be studied due to its mechanism of action.
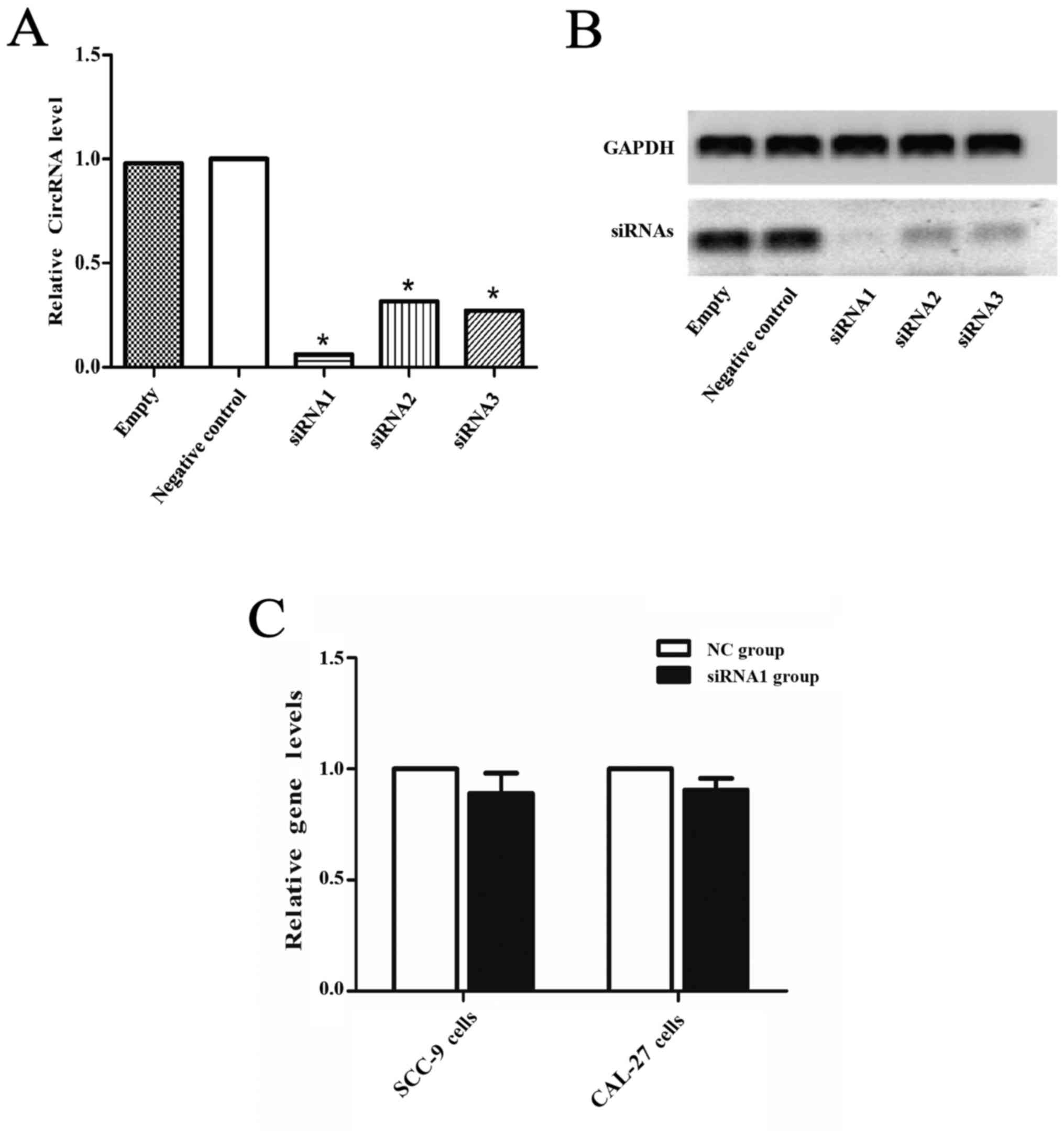
Confirmation of circDOCK1 apoptosis in
OSCC cell lines
CircDOCK1 was significantly downregulated in the
apoptosis model compared with the negative group and markedly
upregulated in the OSCC tissue compared with the para-carcinoma
tissue, however whether it could regulate the apoptosis rate in
OSCC cell lines was still unclear. Hence, a low expression model of
circDOCK1 was constructed with three siRNAs for circDOCK1 in CAL-27
cells. After transfection for 48 h, the effect of the three
different siRNAs against circDOCK1 (si-circDOCK1) was analyzed by
qRT-PCR and nucleic acid electrophoresis. The most marked
inhibitory result was achieved with si-circDOCK1-1 (siRNA1)
(Fig. 4A and B). However, the
expression levels of linear DOCK1 were not affected by siRNA1 in
SCC-9 and CAL-27 cells (Fig. 4C).
FCM detected the apoptosis rate of the low expression model in
CAL-27 and SCC-9 cells. Following transfection with si-circDOCK1-1
for 48 h, the apoptosis rate was increased by 13.7% and 9.6% in
CAL-27 and SCC-9 cells, respectively (Fig. 4D and E). Consequently, circDOCK1
regulated the apoptosis of OSCC cell lines.
Forecasting the circRNA/miRNA/mRNA
axis of circDOCK1
It is well known that circRNAs could serve as miRNA
sponges, indirectly regulating the expression of downstream mRNAs.
The combination of circRNAs and miRNAs as well as miRNAs and mRNAs
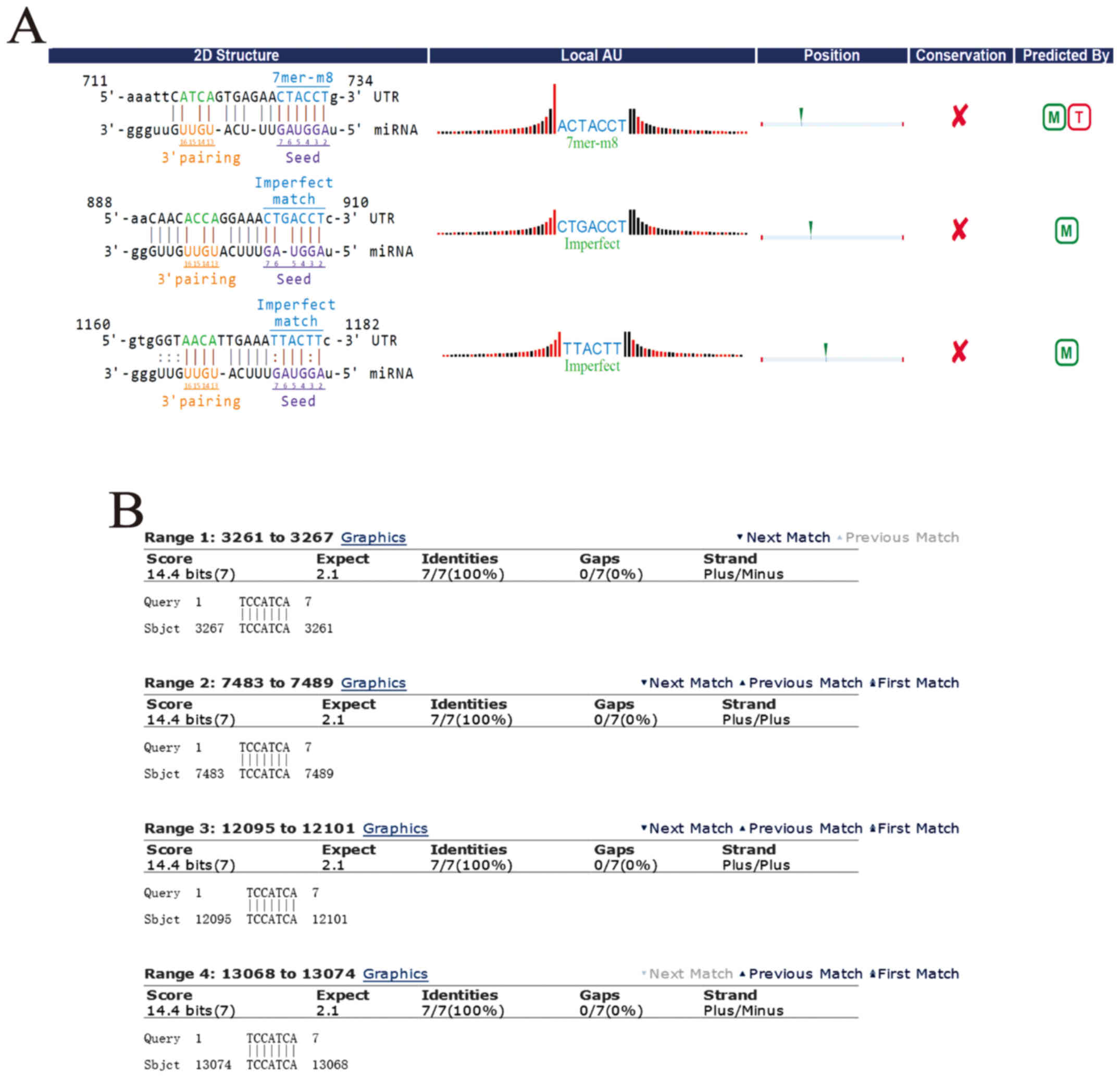
were based on complementary base pairing (Fig. 5A and B). To study the apoptosis
mechanism of circDOCK1, circDOCK1/miRNA/mRNA was predicted. DIANA
TOOLS-mirPath v.3 ascertained all the candidate miRNAs connecting
with circDOCK1, which participated in cancer-related pathways and
apoptosis-related pathways. According to the P-value (P<0.05),
the data revealed that the candidate miRNAs for circDOCK1
hsa-miR-138-2-3p and hsa-miR-196a-5p were involved in
cancer-related pathways. Further analysis of these two pathways
revealed that only hsa-miR-196a-5p was associated with
apoptosis-related pathways, including the p53 signaling pathway
(Fig. 5C); while 48 genes and 14
genes related to hsa-miR-196a-5p were involved in the
cancer-related pathways and the p53 signaling pathway, respectively
(Fig. 5D). In order to further
construct the circDOCK1/miR-196a-5p/mRNA axis which was associated
with the apoptosis pathway, DAVID functional annotation was used. A
total of 55 genes (7 genes were repetitive in cancer-related
pathways and the p53 signaling pathway) were analyzed (Fig. 5E and F). The analysis revealed that
some of these genes were significantly correlated with cell
apoptosis. Three genes were chosen: BIRC3, FOXO1, MDM2, which
participated in apoptosis-related pathways. Furthermore, it is well
known that Cyclin D1 and CDK4 could affect the cell cycle process
and cell proliferation. In addition E2F and c-Myc may block cell
differentiation. Ras, Rac1, ITGA, ITGB and HSP90 were involved in
the PI3K-Akt signaling pathway which regulated multiple
cancer-related phenotypes. In addition, FOXO could serve as a
tumor-inhibiting factor. This indicated that most of the genes
correlated to circDOCK1 were closely related to anti-apoptosis,
tumorigenesis and metastasis.
OSCC cell apoptosis is regulated via
the circDOCK1/miR-196a-5p/BIRC3 pathway
The anti-apoptosis effect of circDOCK1 was
previously demonstrated. Moreover, a low expression model of
circDOCK1 was also constructed. Bioinformatic prediction indicated
that miR-196-5p may be one of the target miRNAs of circDOCK1.
Therefore, the low expression model of circDOCK1 was used to detect
the expression of miR-196a-5p. In addition, a negative group served
as a control. Following transfection for 48 h, the expression
levels of miR-196a-5p were upregulated distinctly in the
low-expression circDOCK1 group compared with the negative control
in CAL-27 and SCC-9 cells, while the expression levels of BIRC3 in
the low-expression circDOCK1 group were downregulated. However,
FOXO1 and MDM2 expressed no significant differences in expression
(Fig. 6A). Moreover, the expression
levels of miR-196a-5p in the OSCC tissue were lower than those in
the para-carcinoma tissue as determined by qRT-PCR, while higher
expression of BIRC3 was consistently detected in OSCC tissue (ten
groups) (Fig. 6B). Then, the
overexpression model of miR-196a-5p was constructed with mimics of
miR-196a-5p in CAL-27 cells. Following transfection with
miR-196a-5p mimics for 48 h, the expression levels increased nearly
seven 7,000-fold (Fig. 6C).
Accordingly, the expression levels of circDOCK1 and BIRC3 were
downregulated in CAL-27 and SCC-9 cells as determined by qRT-PCR
and western blotting (Fig. 6D and
E). Furthermore, FCM revealed that the cell apoptosis rate
after transfection with miR-196a-5p mimics increased by 11.4% and
9.1% in CAL-27 and SCC-9 cells, respectively (Fig. 6F). These results revealed that OSCC
cell apoptosis could be regulated through the
circDOCK1/miR-196a-5p/BIRC3 pathway.
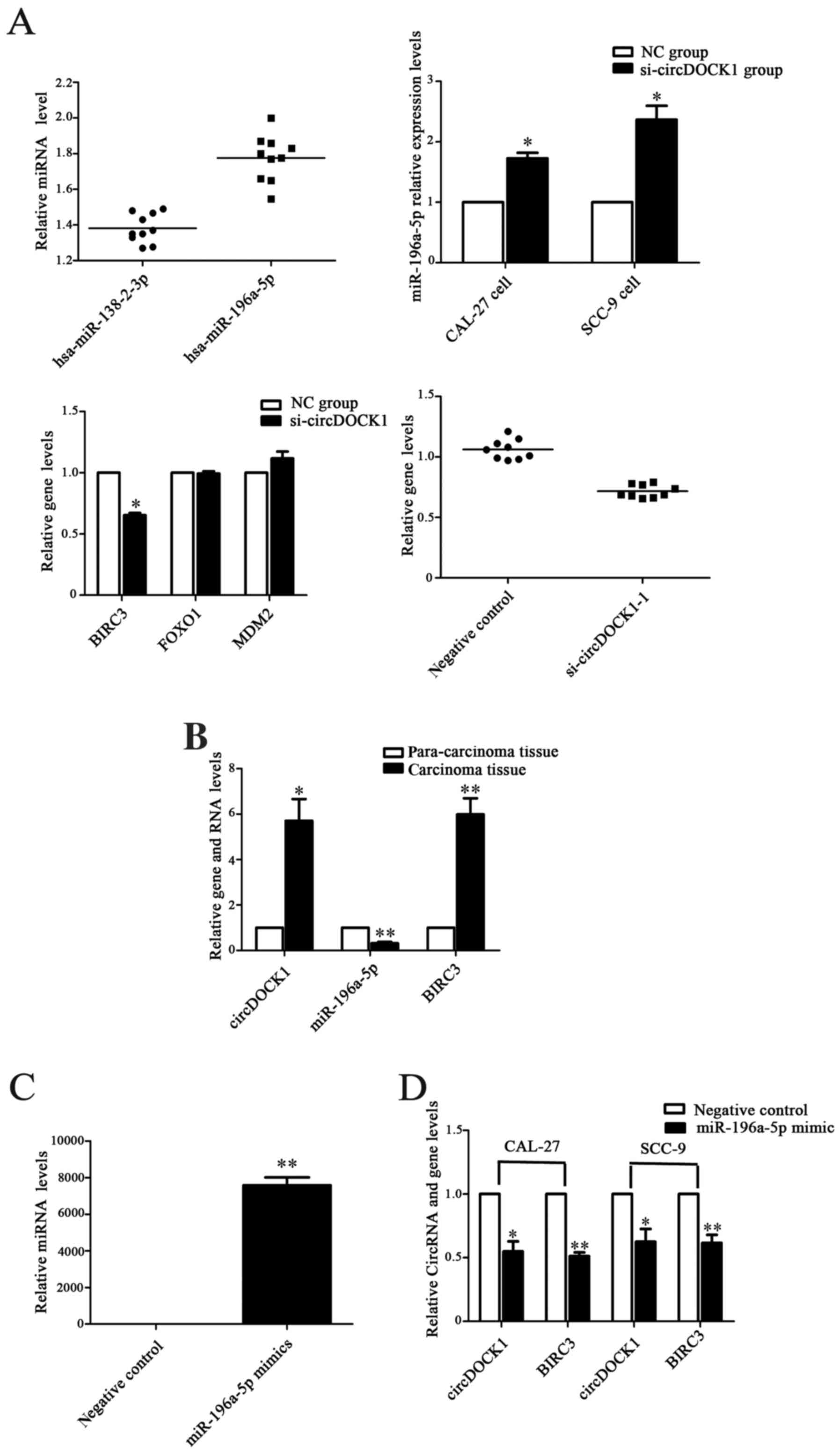 | Figure 6.The circDOCK1/miR-196a-5p/BIRC3
pathway regulated OSCC cell apoptosis. (A) Following transfection
with si-circDOCK1-1 for 48 h, the expression levels of miR-196-5p
were distinctly upregulated, while the expression level of BIRC3 in
the low-expression circDOCK1 group was downregulated. However,
FOXO1 and MDM2 exhibited no significant difference in expression.
(B) The expression level of miR-196a-5p was significantly
downregulated in OSCC tissue compared to para-carcinoma tissue,
while the expression levels of BIRC3 were upregulated. (C)
Following transfection with miR-196a-5p mimics, the expression
level was increased nearly 7,000-fold. (D) The expression levels of
circDOCK1 and BIRC3 were downregulated in CAL-27 and SCC-9 cells
after transfection with miR-196a-5p mimics as determined by
qRT-PCR. (E) The expression levels of circDOCK1 and BIRC3 were
downregulated in CAL-27 and SCC-9 cells after transfection with
miR-196a-5p mimics as determined by western blotting. (F) FCM
revealed that the cell apoptosis rate following transfection with
miR-196a-5p mimics was increased by 11.4% and 9.1% in CAL-27 and
SCC-9 cells, respectively. All data are expressed as the mean ± SE
of three independent experiments. *P<0.05, **P<0.01,
***P<0.001 vs. the control. OSCC, oral squamous cell carcinoma;
FCM, flow cytometry. |
Discussion
As early as 1976, circRNAs have been found in
viroids (36) however they were
often considered as splicing byproducts with low abundance
(6). With the emergence of new
generation sequencing technology and the progress of
bioinformatics, numerous circRNAs were found in eukaryotic
organisms and viruses (37–40). The regulatory effect of circRNAs in
eukaryotic cells was gradually recognized (4,10,11).
Sanger et al (36) found
that viroids were single-stranded covalently closed circRNA
molecules, which could infect certain higher plants. Unlike the
linear RNAs, the 3- and 5-ends of circRNAs were found to be joined
together to form covalently closed loop structures, which indicated
their potential important function (8). Salzman et al found that
numerous circRNAs existed in most normal cells and cancer cells.
The function of circRNAs were confirmed in succession (41). In addition to the potential
regulatory function, circRNAs may become new biomarkers of cancer
diagnosis and targeted therapy.
TNF-α could induce cancer cell apoptosis, which is
an accepted fact. The apoptosis effect in multiple types of cancer
have been demonstrated (42). The
construction of an apoptosis model in OSCC cell lines was
successful, which revealed that TNF-α could affect apoptosis in
OSCC.
In the present study, we first constructed an OSCC
cell apoptosis model and corresponding differentially expressed
circRNA profiles. The circRNA profiles revealed that hundreds of
circRNAs were aberrantly expressed in the OSCC cell apoptosis model
compared with the negative control, suggesting that circRNAs were
involved in the regulation of cell phenotype. We also detected the
differentially expressed circRNA profiles in OSCC tissue and
para-carcinoma tissue. Upon comparison of these two sets of data,
we determined that dozens of circRNAs overlapped, such as
circDOCK1, circDLG1, circFBXL5, circPLCB1, circLOC401320,
circDHDDS, circZFYVE27, circPRDX3, circPPAPDC1A, circQSER1,
circCAPRIN1, circGAPDH, circRNFT2, circMETTL3, circEML1,
circHERC2P3, circCIB2, circABCC1, circCLN3, circABCC3, circPGPEP1,
circCACNG7, circLMF2, circRAD54L2, circABHD6, circLRIG1,
circYEATS2, circSENP5, circDEPDC1B, circG3BP1, circNUP153,
circLRRC16A, circBVES, circFAM120B, circSND1, circCREB3L2,
circTRIM24, circDOCK5, circKAT6A, circC9orf5, circSUSD1, circZMIZ1,
circRPL27A, circEXOC7, circMCM5, circQRICH1, circTJAP1,
circCOL26A1, which indicated that these overlapped differentially
expressed circRNAs may be significant in the regulation of OSCC
cell apoptosis. CircDOCK1 was confirmed to be markedly dysregulated
in the apoptosis model, OSCC cell lines and OSCC tissue compared
with the negative control, HOK and para-carcinoma tissue,
respectively. A previous study revealed that circRNAs from DOCK1
were strongly decreased by TGF-β treatment, while the DOCK1 mRNA
was increased 2-fold. Furthermore TGF-β treatment could induce
epithelial mesenchymal transformation (EMT). This indicated that
one of the functions of circRNAs from DOCK1 may induce the
downregulation of mRNAs in epithelial cells, keeping cellular
stability (43). However, numerous
circRNAs from DOCK1, may possess other functions in biological
regulation. Therefore, circDOCK1 may be a promising biomarker and
therapeutic target for OSCC, and its other functions in OSCC will
be detected continuously in our laboratory.
In addition, it is the first time that
differentially expressed circRNA profiles in OSCC were constructed.
We also constructed the circRNA/miRNA networks of OSCC, which
enriched competing endogenous RNAs (ceRNAs). As negative regulatory
factors, miRNAs could degrade or suppress their target mRNAs
(32,44,45).
However, in the past two years, circRNAs have been demonstrated to
serve as miRNA sponges, such as the circRNAs CDR1as (7), SRY(7) and circ-ITCH (46). They could interact with relevant
miRNAs based on conserved seed sequence matches, which suppress the
negative effect of miRNAs, thus upregulating the target mRNAs. For
instance, miR-7, which could be adsorbed by CDR1as, and could
inhibit the EGFR signaling pathway by targeting EGFR, Raf1, PSME3,
PLEC1, CKAP4, CNOT8, CNN3, CAPZA1, PFN2 and ARF4 (47). Furthermore, the EGFR signaling
pathway exhibited a significant connection with cancer development
(48). Therefore, circRNAs play a
crucial role in the ceRNA system. As an upstream regulator, the
change in the expression levels of circRNA may lead to the change
of miRNAs and mRNAs. Eventually the balance of cells would lead to
abnormity, including cancers. According to recent research, apart
from their function as miRNA-sponges, circRNAs could serve as
regulators of alternative splicing (9), transcription factors (10) and encode for proteins (11). The regulatory mechanism of circRNAs
is complex and it is not clear how many circRNAs could function as
miRNA-sponges. To date, there has been no report on circRNAs in
OSCC.
Improving the apoptosis rate is a great approach to
hinder the development of cancers. As a result, we found that
circDOCK1 was significantly downregulated in the apoptosis model
compared to the control. Moreover, our results revealed that
circDOCK1 was highly expressed in OSCC tissue and cell lines
compared to para-carcinoma and HOK cells, respectively. In order to
further confirm the biological function of circDOCK1 in OSCC, we
downregulated its expression using siRNAs and the apoptosis rate
increased, which was consistent with our previous results. With the
use of bioinformatics analysis, we surmised that circDOCK1 could
serve as miR-196a-5p sponge. DIANA-miRPath analysis revealed that
miR-196a-5p may be implicated in cancer-associated pathways and
regulate apoptosis-related genes. Based on diverse bioinformatics
data, we depicted a circRNA/miRNA/mRNA regulatory pathway in OSCC.
The data revealed that 61 genes, which were related to miR-196a-5p,
were involved in cancer-related pathways and the p53 signaling
pathway. Following the use of DAVID functional annotations, we
found that BIRC3, FOXO1 and MDM2 participated in apoptosis-related
pathways. In order to validate the circDOCK1/miR-196a-5p/mRNA axis,
our results revealed that when the expression level of circDOCK1
was downregulated, miR-196-5p was upregulated while BIRC3 was
downregulated accordingly. Moreover, when the expression level of
miR-196-5p was upregulated, circDOCK1 and BIRC3 were downregulated
accordingly, too. We also detected the expression level of
miR-196a-5p and BIRC3 in OSCC tissue and cell lines compared to
para-carcinoma and HOK cells, respectively and found that
miR-196a-5p was expressed at a low level while BIRC3 was highly
expressed in OSCC tissue and cell lines. Both the RNA level and
protein level exhibited the same outcome. A recent study
demonstrated that both in vivo and in vitro BIRC3
upregulation led to apoptosis evasion and therapeutic resistance in
genome-wide analysis of glioblastoma (GBM) (49). Inhibition of IAP protein expression
by small molecule inhibitors or molecular biological assays could
promote tumor cell apoptosis and improve sensitivity to
chemotherapy (50), which was
consistent with our results. However, we noted that the apoptotic
rate increased predominately in quadrant 2 rather than quadrant 4
[compared figs. 4D and E, 6F and 1A
(apoptosis model)] in the siRNA experiment, which may signify that
more late apoptosis/necrosis occurred.
In conclusion, our results demonstrated that
circDOCK1 was upregulated and revealed its apoptosis-regulating
function in OSCC. Using multiple bioinformatic methods, we
constructed the circRNA/miRNA/mRNA regulatory network in OSCC.
Finally, the apoptosis-regulating function of the
circDOCK1/miR-196a-5p/BIRC3 axis was further confirmed in OSCC. In
the future, other functions of circDOCK1 may be detected in our
laboratory however our present results revealed that circDOCK1 may
serve as a promising diagnostic biomarker and therapeutic target in
OSCC.
References
|
1
|
Fitzmaurice C, Dicker D, Pain A, Hamavid
H, Moradi-Lakeh M, MacIntyre MF, Allen C, Hansen G, Woodbrook R,
Wolfe C, et al Global Burden of Disease Cancer Collaboration, : The
Global Burden of Cancer 2013. JAMA Oncol. 1:505–527. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lingen MW, Kalmar JR, Karrison T and
Speight PM: Critical evaluation of diagnostic aids for the
detection of oral cancer. Oral Oncol. 44:10–22. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ghantous Y, Yaffi V and Abu-Elnaaj I; Oral
cavity cancer, : epidemiology and early diagnosis. Refuat Hapeh
Vehashinayim (1993). 3255–63. (71)2015.(In Hebrew). PubMed/NCBI
|
|
4
|
Jeck WR and Sharpless NE: Detecting and
characterizing circular RNAs. Nat Biotechnol. 32:453–461. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lasda E and Parker R: Circular RNAs:
Diversity of form and function. RNA. 20:1829–1842. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Cocquerelle C, Mascrez B, Hétuin D and
Bailleul B: Mis-splicing yields circular RNA molecules. FASEB J.
7:155–160. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Hansen TB, Jensen TI, Clausen BH, Bramsen
JB, Finsen B, Damgaard CK and Kjems J: Natural RNA circles function
as efficient microRNA sponges. Nature. 495:384–388. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Memczak S, Jens M, Elefsinioti A, Torti F,
Krueger J, Rybak A, Maier L, Mackowiak SD, Gregersen LH, Munschauer
M, et al: Circular RNAs are a large class of animal RNAs with
regulatory potency. Nature. 495:333–338. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ashwal-Fluss R, Meyer M, Pamudurti NR,
Ivanov A, Bartok O, Hanan M, Evantal N, Memczak S, Rajewsky N and
Kadener S: circRNA biogenesis competes with pre-mRNA splicing. Mol
Cell. 56:55–66. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Li Z, Huang C, Bao C, Chen L, Lin M, Wang
X, Zhong G, Yu B, Hu W, Dai L, et al: Exon-intron circular RNAs
regulate transcription in the nucleus. Nat Struct Mol Biol.
22:256–264. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wang Y and Wang Z: Efficient backsplicing
produces translatable circular mRNAs. RNA. 21:172–179. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Thomas LF and Sætrom P: Circular RNAs are
depleted of polymorphisms at microRNA binding sites.
Bioinformatics. 30:2243–2246. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kumar L, Shamsuzzama, Haque R, Baghel T
and Nazir A: Circular RNAs: the emerging class of non-coding RNAs
and their potential role in human neurodegenerative diseases. Mol
Neurobiol. Oct 29–2016.(Epub ahead of print).
|
|
14
|
Nan A, Chen L, Zhang N, Liu Z, Yang T,
Wang Z, Yang C and Jiang Y: A novel regulatory network among
LncRpa, CircRar1, miR-671 and apoptotic genes promotes lead-induced
neuronal cell apoptosis. Arch Toxicol. 91:1671–1684. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Bonizzato A, Gaffo E, Te Kronnie G and
Bortoluzzi S: CircRNAs in hematopoiesis and hematological
malignancies. Blood Cancer J. 6:e4832016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Liu Q, Zhang X, Hu X, Dai L, Fu X, Zhang J
and Ao Y: Circular RNA related to the chondrocyte ECM regulates
MMP13 expression by functioning as a miR-136 ‘sponge’ in human
cartilage degradation. Sci Rep. 6:225722016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Peng L, Chen G, Zhu Z, Shen Z, Du C, Zang
R, Su Y, Xie H, Li H, Xu X, et al: Circular RNA ZNF609 functions as
a competitive endogenous RNA to regulate AKT3 expression by
sponging miR-150-5p in Hirschsprung's disease. Oncotarget.
8:808–818. 2017.PubMed/NCBI
|
|
18
|
Wang X, Zhang Y, Huang L, Zhang J, Pan F,
Li B, Yan Y, Jia B, Liu H, Li S, et al: Decreased expression of
hsa_circ_001988 in colorectal cancer and its clinical
significances. Int J Clin Exp Pathol. 8:16020–16025.
2015.PubMed/NCBI
|
|
19
|
Xie H, Ren X, Xin S, Lan X, Lu G, Lin Y,
Yang S, Zeng Z, Liao W, Ding YQ, et al: Emerging roles of
circRNA_001569 targeting miR-145 in the proliferation and invasion
of colorectal cancer. Oncotarget. 7:26680–26691. 2016.PubMed/NCBI
|
|
20
|
Li H, Hao X, Wang H, Liu Z, He Y, Pu M,
Zhang H, Yu H, Duan J and Qu S: Circular RNA expression profile of
pancreatic ductal adenocarcinoma revealed by microarray. Cell
Physiol Biochem. 40:1334–1344. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhong Z, Lv M and Chen J: Screening
differential circular RNA expression profiles reveals the
regulatory role of circTCF25-miR-103a-3p/miR-107-CDK6 pathway in
bladder carcinoma. Sci Rep. 6:309192016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Xia W, Qiu M, Chen R, Wang S, Leng X, Wang
J, Xu Y, Hu J, Dong G, Xu PL, et al: Circular RNA has_circ_0067934
is upregulated in esophageal squamous cell carcinoma and promoted
proliferation. Sci Rep. 6:355762016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yang P, Qiu Z, Jiang Y, Dong L, Yang W, Gu
C, Li G and Zhu Y: Silencing of cZNF292 circular RNA suppresses
human glioma tube formation via the Wnt/β-catenin signaling
pathway. Oncotarget. 7:63449–63455. 2016.PubMed/NCBI
|
|
24
|
Hao Z, Yang J, Wang C, Li Y, Zhang Y, Dong
X, Zhou L, Liu J, Zhang Y and Qian J: MicroRNA-7 inhibits
metastasis and invasion through targeting focal adhesion kinase in
cervical cancer. Int J Clin Exp Med. 8:480–487. 2015.PubMed/NCBI
|
|
25
|
Berg V, Rusch M, Vartak N, Jüngst C,
Schauss A, Waldmann H, Hedberg C, Pallasch CP, Bastiaens PI, Hallek
M, et al: miRs-138 and −424 control palmitoylation-dependent
CD95-mediated cell death by targeting acyl protein thioesterases 1
and 2 in CLL. Blood. 125:2948–2957. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Liu R, Liu X, Zheng Y, Gu J, Xiong S,
Jiang P, Jiang X, Huang E, Yang Y, Ge D, et al: MicroRNA-7
sensitizes non-small cell lung cancer cells to paclitaxel. Oncol
Lett. 8:2193–2200. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wang Q, Tang H, Yin S and Dong C:
Downregulation of microRNA-138 enhances the proliferation,
migration and invasion of cholangiocarcinoma cells through the
upregulation of RhoC/p-ERK/MMP-2/MMP-9. Oncol Rep. 29:2046–2052.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ayaz L, Çayan F, Balci Ş, Görür A, Akbayir
S, Yıldırım Yaroğlu H, Doğruer Unal N and Tamer L: Circulating
microRNA expression profiles in ovarian cancer. J Obstet Gynaecol.
34:620–624. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Shang X, Li G, Liu H, Li T, Liu J, Zhao Q
and Wang C: Comprehensive circular RNA profiling reveals that
hsa_circ_0005075, a new circular RNA biomarker, is involved in
hepatocellular carcinoma development. Medicine (Baltimore).
95:e38112016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Qin M, Liu G, Huo X, Tao X, Sun X, Ge Z,
Yang J, Fan J, Liu L and Qin W: Hsa_circ_0001649: A circular RNA
and potential novel biomarker for hepatocellular carcinoma. Cancer
Biomark. 16:161–169. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Krol J, Loedige I and Filipowicz W: The
widespread regulation of microRNA biogenesis, function and decay.
Nat Rev Genet. 11:597–610. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
32
|
Bartel DP: MicroRNAs: Target recognition
and regulatory functions. Cell. 136:215–233. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Carthew RW and Sontheimer EJ: Origins and
mechanisms of miRNAs and siRNAs. Cell. 136:642–655. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Huntzinger E and Izaurralde E: Gene
silencing by microRNAs: Contributions of translational repression
and mRNA decay. Nat Rev Genet. 12:99–110. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Gomes CC, de Sousa SF and Gomez RS:
MicroRNAs: Small molecules with a potentially role in oral squamous
cell carcinoma. Curr Pharm Des. 19:1285–1291. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Sanger HL, Klotz G, Riesner D, Gross HJ
and Kleinschmidt AK: Viroids are single-stranded covalently closed
circular RNA molecules existing as highly base-paired rod-like
structures. Proc Natl Acad Sci USA. 73:pp. 3852–3856. 1976;
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Arnberg AC, Van Ommen GJ, Grivell LA, Van
Bruggen EF and Borst P: Some yeast mitochondrial RNAs are circular.
Cell. 19:313–319. 1980. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Kos A, Dijkema R, Arnberg AC, van der
Meide PH and Schellekens H: The hepatitis delta (delta) virus
possesses a circular RNA. Nature. 323:558–560. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Capel B, Swain A, Nicolis S, Hacker A,
Walter M, Koopman P, Goodfellow P and Lovell-Badge R: Circular
transcripts of the testis-determining gene Sry in adult mouse
testis. Cell. 73:1019–1030. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Zaphiropoulos PG: Circular RNAs from
transcripts of the rat cytochrome P450 2C24 gene: Correlation with
exon skipping. Proc Natl Acad Sci USA. 93:pp. 6536–6541. 1996;
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Salzman J, Gawad C, Wang PL, Lacayo N and
Brown PO: Circular RNAs are the predominant transcript isoform from
hundreds of human genes in diverse cell types. PLoS One.
7:e307332012. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Balkwill F: Tumour necrosis factor and
cancer. Nat Rev Cancer. 9:361–371. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Conn SJ, Pillman KA, Toubia J, Conn VM,
Salmanidis M, Phillips CA, Roslan S, Schreiber AW, Gregory PA and
Goodall GJ: The RNA binding protein quaking regulates formation of
circRNAs. Cell. 160:1125–1134. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Mallory AC and Vaucheret H: MicroRNAs:
Something important between the genes. Curr Opin Plant Biol.
7:120–125. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Li F, Zhang L, Li W, Deng J, Zheng J, An
M, Lu J and Zhou Y: Circular RNA ITCH has inhibitory effect on ESCC
by suppressing the Wnt/β-catenin pathway. Oncotarget. 6:6001–6013.
2015.PubMed/NCBI
|
|
47
|
Chen YJ, Chien PH, Chen WS, Chien YF, Hsu
YY, Wang LY, Chen JY, Lin CW, Huang TC, Yu YL, et al: Hepatitis B
virus-encoded X protein downregulates EGFR expression via inducing
microRNA-7 in hepatocellular carcinoma cells. Evid Based Complement
Alternat Med. 2013:6823802013.PubMed/NCBI
|
|
48
|
Xu W, Jing H and Zhang F: Epidermal growth
factor receptor-targeted therapy in colorectal cancer. Front Biosci
(Landmark Ed). 21:410–418. 2016. View
Article : Google Scholar : PubMed/NCBI
|
|
49
|
Wang D, Berglund A, Kenchappa RS, Forsyth
PA, Mulé JJ and Etame AB: BIRC3 is a novel driver of therapeutic
resistance in Glioblastoma. Sci Rep. 6:217102016. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Hird AW, Aquila BM, Hennessy EJ, Vasbinder
MM and Yang B: Small molecule inhibitor of apoptosis proteins
antagonists: A patent review. Expert Opin Ther Pat. 25:755–774.
2015. View Article : Google Scholar : PubMed/NCBI
|