Introduction
Esophageal carcinoma is among the common types of
malignant tumors to develop in humans, and is classified into two
pathological types, namely esophageal squamous cell carcinoma
(ESCC) and esophageal adenocarcinoma. Approximately 70% of ESCC
cases in the world occur in China (1). The identification of novel low-toxic
therapeutic agents is urgently required for the prevention and
treatment of ESCC.
Natural plant ingredients or small synthetic
molecules are currently a topic of focus in the field of cancer
prevention and treatment. Flavonoids are polyphenols widely found
in a variety of edible plants, and have been shown to have
antioxidative, anti-inflammatory and antitumor activities in both
in vivo and in vitro systems (2–4).
Specifically, their antitumor effects include inhibition of cell
proliferation and promotion of apoptosis in vitro (5,6).
Various flavonoids have been reported to serve as cancer
chemopreventive agents that exert protective effects against cancer
development (5). Eupatilin is a
type of antitumorigenic flavonoid component, with the molecular
structure 5,7-dihydroxy-3′,4′,6-trimethoxyflavone, as shown in
Fig. 1A. It has been found that
eupatilin has anti-inflammatory and antitumor effects (6–10), but
the underlying mechanisms remain unclear.
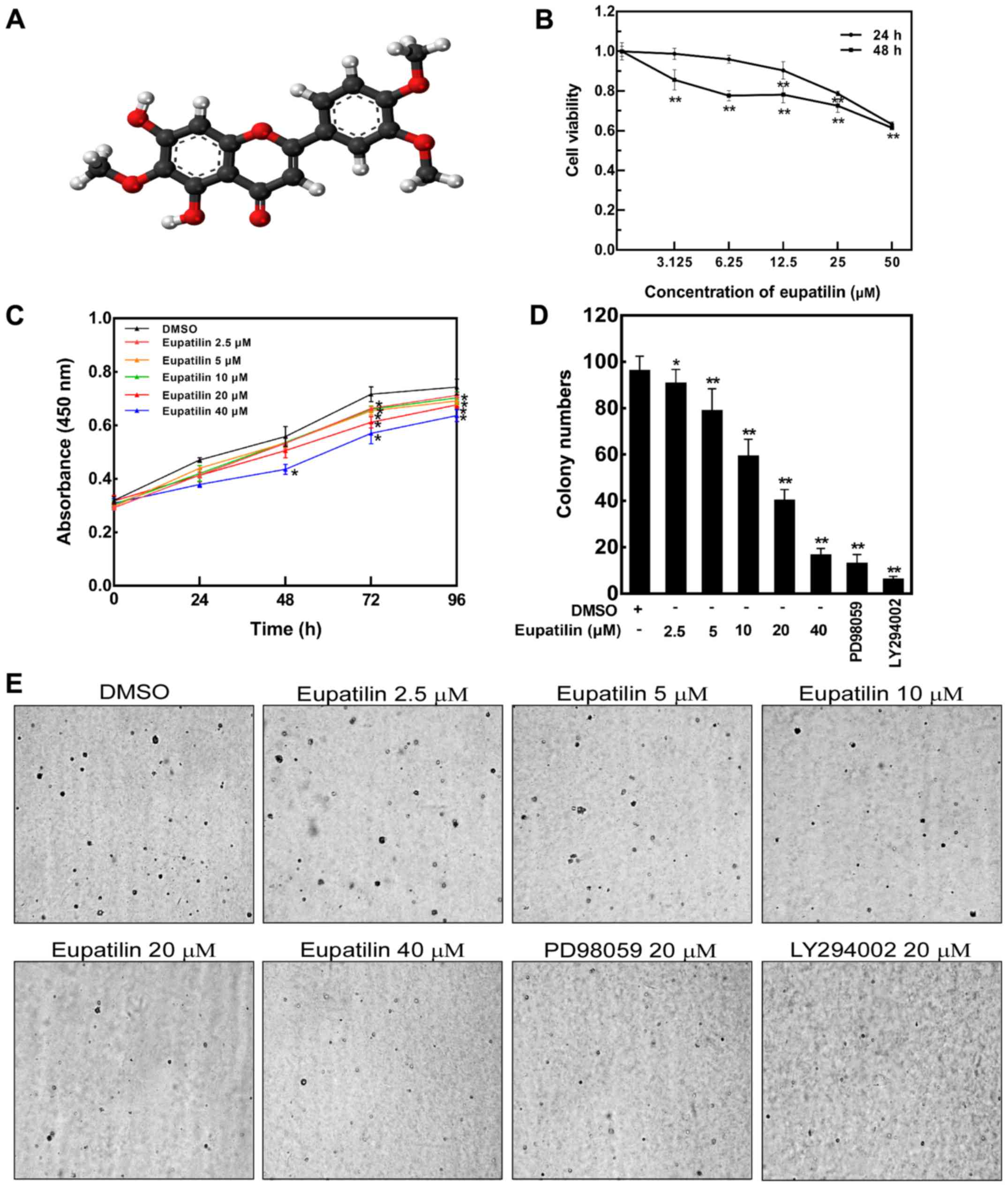 | Figure 1.Eupatilin inhibits the proliferation
and anchorage-independent growth of esophageal cancer TE1 cells.
(A) Chemical structure of eupatilin
(5,7-dihydroxy-3′,4′,6-trimethoxyflavone). Color code: carbon,
black; hydrogen, white; oxygen, red. (B) The viability of TE1 cells
was detected by Cell Counting Kit-8 (CCK-8) assay. TE1 cells were
treated With the indicated doses of eupatilin for 24 and 48 h. Data
are presented as mean values ± SD from triplicate experiments.
Statistical significance was determined by Student's t-test
(**P<0.01 vs. DMSO control). (C) Proliferation inhibition effect
of eupatilin on TE1 cells. TE1 cells were treated with different
concentrations of eupatilin for 0, 24, 48, 72 or 96 h, and the
absorbance of cells was assessed using a CCK-8 assay. Data are
presented as mean values ± SD from triplicate experiments
(*P<0.05 vs. DMSO control). (D and E) Inhibitory effect of
eupatilin on the colony formation capacity of TE1 cells. Cells were
treated with the indicated concentrations of eupatilin with or
without 20 µM PD98059 or 20 µM LY 294002. Images were captured
using a microscope (magnification, ×4). Representative images are
shown and data are presented as mean values ± SD from triplicate
experiments. As shown by the statistical results of the colony
assay, eupatilin obviously reduced the number and size of the TE1
colonies compared with the DMSO control (*P<0.05 and
**P<0.01). |
Several signaling pathways that regulate cell
proliferation and survival have been implicated as therapeutic
targets in cancer in preclinical and clinical trials (11). The Ras signaling pathway,
principally consisting of Ras, Raf, MEK and MAPK
kinase/extracellular signal-regulated kinase (ERK), plays a
significant role in the regulation of cell proliferation through
its influence on growth factors (12–14).
Uncontrolled Ras signaling has been related to the development of
many human cancers (15–17). Furthermore, MAPK activation often
leads to the growth or metastasis of tumors.
Additionally, abnormal activation of the PI3K/Akt
signaling pathway has been reported in several types of human
cancers, and its activation results in cancer development, namely
by influencing tumor cell proliferation, growth, angiogenesis and
survival (18). Moreover, the Akt
signal pathway may play an important role in cell cycle control by
targeting cyclin D1, which is also regulated by Ras-dependent
signaling (19–22). Thus, the MEK/ERK and PI3K/Akt
pathways may co-regulate cyclin D1 to affect cell cycling.
Researchers have found that the targets of many
flavonoid compounds are involved in multiple signaling pathways,
including the Ras/Raf/MEK/MAPK and PI3K/Akt/PTEN/mTOR pathways
(11,23). In the present study, we evaluated
the inhibitory effects of the flavonoid compound eupatilin on human
esophageal cancer cells. The results showed that eupatilin
inhibited the proliferation of esophageal cancer TE1 cells,
decreased anchorage-independent growth of the cells, and induced
G0/G1 phase cell cycle arrest. Furthermore,
our results indicated that eupatilin suppressed TE1 cell
proliferation by targeting the Akt/GSK3β and MAPK/ERK signaling
cascades. We also found that eupatilin decreased tumor volume in a
TE1 xenograft mouse model, while the body weights of all mice
remained stable after eupatilin treatment. Additionally, Ki-67
expression and the phosphorylation of Akt and ERK1/2 were
suppressed by eupatilin in the xenograft tumors. Overall, our
results indicate that the antiproliferative effect of eupatilin on
esophageal cancer TE1 cells is at least in part associated with
inhibition of the Akt and ERK pathways.
Materials and methods
Cell culture and chemicals
The human esophageal cancer cell line TE1 was
cultured in Dulbecco's modified Eagle's medium (DMEM), which
contained 10% fetal bovine serum (FBS), streptomycin (100 U/ml) and
penicillin (100 U/ml), at 37°C in a 5% CO2 saturated
humidity incubator. Eupatilin was supplied by Chunqiu (Nanjing,
China) and dissolved in dimethyl sulfoxide (DMSO). The primary
antibodies were purchased from the Cell Signaling Technology, Inc.
(Danvers, MA, USA) and Santa Cruz Biotechnology, Inc. (Dallas, TX,
USA). The secondary antibodies were purchased from Santa Cruz
Biotechnology, Inc. PD98059 (MEK inhibitor) and LY294002 (PI3K
inhibitor) were obtained from Beyotime Institute of Biotechnology
(Shanghai, China).
Determination of cell viability
We used the Cell Counting Kit-8 (CCK-8; KeyGen
Biotech Co., Ltd., Nanjing, China) method to detect the toxicity of
eupatilin in TE1 cells. TE1 cells (1×104) were plated
into the wells of 96-well plates. When the cells were adhered to
the bottom of the wells, the medium was replaced with medium
containing different concentrations of eupatilin (0, 2.5, 5, 10, 20
or 40 µM). After 24 and 48 h of incubation, 10 µl of CCK-8 solution
was added and the cells were incubated for 2 h in a CO2
incubator. Subsequently, cell absorbance was measured with a
microplate reader (Bio-Rad Laboratories, Inc., Hercules, CA, USA)
at 450 nm. The results were calculated as mean and standard
deviation, and are expressed as a percentage reduction for the
eupatilin-treated group, assuming that the absorbance of the
control cells was 100%.
Cell proliferation assay
The esophageal cancer TE1 cells (3×103)
were transferred into the wells of 96-well plates in 200 µl culture
medium and incubated for 6–8 h until the cells were adhered to the
wells. The medium was then replaced with medium containing
eupatilin at the indicated doses (0, 2.5, 5, 10, 20 or 40 µM), 20
µM PD98059 or 20 µM LY294002. The treated cells were cultured for
0, 24, 48, 72 or 96 h. Subsequently, 10 µl of CCK-8 solution was
added to each well, and after a 2-h incubation in a CO2
incubator, the optical densities of the wells at 450 nm were
measured with a microplate reader.
Anchorage-independent cell growth
assay
A volume of 3 ml basal medium eagle (BME)
(Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) including 10% FBS
and 0.5% agarose was spread in the wells of 6-well plates, to which
different concentrations of eupatilin, 20 µM PD98059 or 20 µM
LY294002 were added before the mixture became solidified. TE1 cells
were then suspended at a density of 8×103 cells in 1 ml
BME (10% FBS and 0.33% agarose) with the indicated concentrations
of eupatilin (0, 2.5, 5, 10, 20 or 40 µM), 20 µM PD98059 or 20 µM
LY294002, and the mixture was seeded onto the top of the agarose
layer in each well of the 6-well plates. The cells were cultured in
a CO2 incubator, and we observed and calculated the
colony number in each well on the 14th day of
culture.
Cell cycle assay by flow
cytometry
TE1 cells were treated with different concentrations
of eupatilin, 20 µM PD98059 or 20 µM LY294002 for 24 h. The cells
were then trypsinized and washed twice with cold phosphate-buffered
saline (PBS). Subsequently, the cells were fixed with 70% ethyl
alcohol, then stored at 4°C until use. To evaluate cell cycle
distribution, the stored cells were washed twice with PBS and
incubated with 50 µg/ml RNase A for 2 h at 37°C. The cells were
then stained with 50 µg/ml propidium iodide for 30 min in the dark
at room temperature. Following staining, the DNA contents of the
cells (1×104) was analyzed with a FACSCalibur flow
cytometer and CellQuest analysis software (BD Biosciences, Franklin
Lakes, NJ, USA).
Western blot analysis
TE1 cells were treated with different concentrations
of eupatilin, 20 µM PD98059 or 20 µM LY294002 for 24 h, after which
total cell protein was isolated using RIPA lysis buffer (50 mM
Tris-base, 1% NP-40, 0.25% SOD, 150 mM NaCl, 1 mM EDTA, 0.1% SDS, 1
mM Na3VO4, 1 mM NaF, 1 mM PMSF and 10 µl
protease inhibitor). The protein concentration of the supernatant
was measured using a protein assay kit (Beyotime Institute of
Biotechnology) with a microplate reader. Subsequently, 50 µg of
protein, dissolved in lysis buffer, was separated on a 10% SDS-PAGE
gel and transferred to polyvinylidene difluoride (PVDF) membranes.
The membranes were blocked with 5% non-fat milk-TBST buffer (TBS
containing 0.1% Tween-20) for 2 h at room temperature. After
blocking, the PVDF membranes were incubated with primary antibodies
against Akt (1:1,000 dilution; cat. no. 2938s; Cell Signaling
Technology, Inc.), GSK3β (1:1,000 dilution; cat. no. 9315; Cell
Signaling Technology, Inc.), CREB (1:1,000 dilution; cat. no. 9197;
Cell Signaling Technology, Inc.), ERK1/2 (1:1,000 dilution; cat.
no. 4695; Cell Signaling Technology, Inc.), phospho-Akt (Ser473)
(1:500 dilution; cat. no. 4060; Cell Signaling Technology, Inc.),
phospho-GSK3β (Ser9) (1:1,000 dilution; cat. no. 9336; Cell
Signaling Technology, Inc.), phospho-CREB (Ser133) (1:1,000
dilution; cat. no. 9191; Cell Signaling Technology, Inc.) and
phospho-ERK1/2 (Thr202/Tyr204) (1:1,000 dilution; cat. no. 9102;
Cell Signaling Technology, Inc.) overnight at 4°C. Equal protein
loading was compared using α-tubulin antibody (1:1,000 dilution;
Santa Cruz Biotechnology, Inc.). The membrane was washed 3 times
with TBST buffer for 10 min per wash, then incubated with HRP-IgG
secondary antibody (1:200 dilution; Santa Cruz Biotechnology, Inc.)
for 4 h at 4°C. Following washing of the membrane 3 times for 10
min each, the transferred proteins were detected as protein bands
using chemiluminescence reagent with an ImageQuant LAS 4000 imager
(GE Healthcare, Chicago, IL, USA).
In vivo tumor growth assay
All animal studies conformed to guidelines approved
by the Research Ethics Committee of Zhengzhou University. Athymic
nude mice (BALB/c nude mice, female, 4–6 weeks old) were purchased
from Vital River Laboratory Animal Technology Co., Ltd. (Beijing,
China). The animals were kept at 23–25°C under a 12-h light/dark
cycle and administered sterile water and food ad libitum. After 1
week of adaptation, TE1 cells (1×107 in 0.1 ml) were
subcutaneously inoculated into the right flank of each mouse.
Animals were randomly divided into the following 3 groups (n=5 mice
per group): Vehicle group; 10 mg/kg eupatilin-treated group; and 50
mg/kg eupatilin-treated group. The mice were administered eupatilin
(10 or 50 mg/kg body weight in 1 µl of DMSO as vehicle) or vehicle
alone 3 times/week by intraperitoneal injection. Mice were weighed
and tumor size was measured with a caliper 3 times/week. Tumor
volume was calculated from cross-sectional measurements of the
individual tumor based on the following formula: Tumor volume
(mm3)=(length × width × width/2). Mice were monitored
until day 36, after which the mice were euthanized and the tumors
were extracted.
Hematoxylin and eosin staining
All acquired tumor tissues were subject to
hematoxylin and eosin (H&E) staining. Tumor tissues were fixed
with 10% formalin and embedded in paraffin, and 4-µm-thick specimen
sections were prepared on glass slides. The sections were
deparaffinized in xylene and rehydrated with graded alcohol, then
stained with H&E and scanned using TissueFAXS (TissueGnostics
GmbH, Vienna, Austria).
Immunohistochemical staining
Tumor tissues were fixed with 10% formalin and
embedded in paraffin, and 4-µm-thick specimen sections were
prepared on glass slides. The sections were deparaffinized in
xylene and rehydrated with graded alcohol, then soaked with 0.3%
hydrogen peroxide (H2O2) for 5 min. Antigen
retrieval was performed in sodium citrate buffer (pH 6.0) with a
microwave. Subsequently, the sections were incubated with
antibodies against Ki-67 (ZM-0165) (1:50 dilution; Zhongshan
Jinqiao Biotechnology Co., Ltd., Beijing, China), p-Akt (Ser473)
(1:50 dilution; cat. no. 4060; Cell Signaling Technology, Inc.),
p-ERK1/2 (Thr202/Tyr204) (1:100 dilution; cat. no. 9102; Cell
Signaling Technology, Inc.) at 4°C overnight, then with
HRP-conjugated secondary antibody (ZSGB-BIO, Beijing, China) at
37°C for 20 min. The peroxidase reaction was visualized by
incubating the sections with 2,4-diaminobenzidine substrate. After
rinsing in water, the sections were counterstained with
hematoxylin, then dehydrated with graded alcohol and cover slipped.
All of the sections were scanned using TissueFAXS analyzed using
HistoQuest 4.0 software (both from TissueGnostics GmbH) and
evaluated using the H-Score system.
Statistical analysis
All quantitative data are expressed as the mean ±
standard deviation. Multi-group comparisons of the means were
carried out by one-way analysis of variance (ANOVA) test with post
hoc contrasts by Student-Newman-Keuls test. ANOVA and Student's
t-test were used for statistical analysis using SPSS 22.0 software
(IBM Corp., Armonk, NY, USA). The sample sizes were chosen to allow
for statistical significance testing assuming a major effect and a
small variation. A value of P<0.05 was considered to indicate
statistical significance.
Results
Eupatilin inhibits the proliferation
of TE1 cells
Eupatilin is a type of flavonoid and exhibits
anti-inflammatory and antitumor effects. To evaluate the effect of
eupatilin on the proliferation of human esophageal cancer TE1
cells, the cytotoxic effect of eupatilin on TE1 cells was measured
by CCK-8 assay. TE1 cells were treated with different
concentrations of eupatilin (3.125, 6.25, 12.5, 25 or 50 µM) for 24
and 48 h. The results showed that the percentage of viable TE1
cells was reduced to 60% following treatment with 50 µM eupatilin
for 48 h (Fig. 1B). Following
treatment of TE1 cells with these concentrations of eupatilin (2.5,
5, 10, 20 or 40 µM) for different time periods (0, 24, 48, 72 or 96
h), the cell growth curve indicated that eupatilin inhibited TE1
cell proliferation in concentration-dependent and time-dependent
manners (Fig. 1C). The
concentration of the drug tested in the study was selected
according to the cytotoxicity of eupatilin shown in Fig. 1B. The results showed that the
percentage of viable TE1 cells was reduced to 80% following
treatment with 3.125 µM eupatilin for 48 h, which indicated that
eupatilin has a significant antiproliferation effect on human
esophageal cancer at low concentrations. When the concentration of
eupatilin was 50 µM, the cell viability was 60% for 48 h (Fig. 1B). Therefore, concentrations of
eupatilin from 2.5 to 40 µM were used for subsequent
experiments.
Eupatilin suppresses the
anchorage-independent growth of TE1 cells
To determine whether eupatilin inhibits the
anchorage-independent growth of TE1 cells, we observed cell colony
number in a soft agar assay after a 14-day culture. The results
indicated that eupatilin inhibited the colony formation capacity of
TE1 cells dose-dependently compared with the DMSO controls
(Fig. 1D and E). Additionally, MEK
and PI3K inhibitors, PD98059 and LY294002 respectively, were also
observed to inhibit TE1 colony formation in the assay.
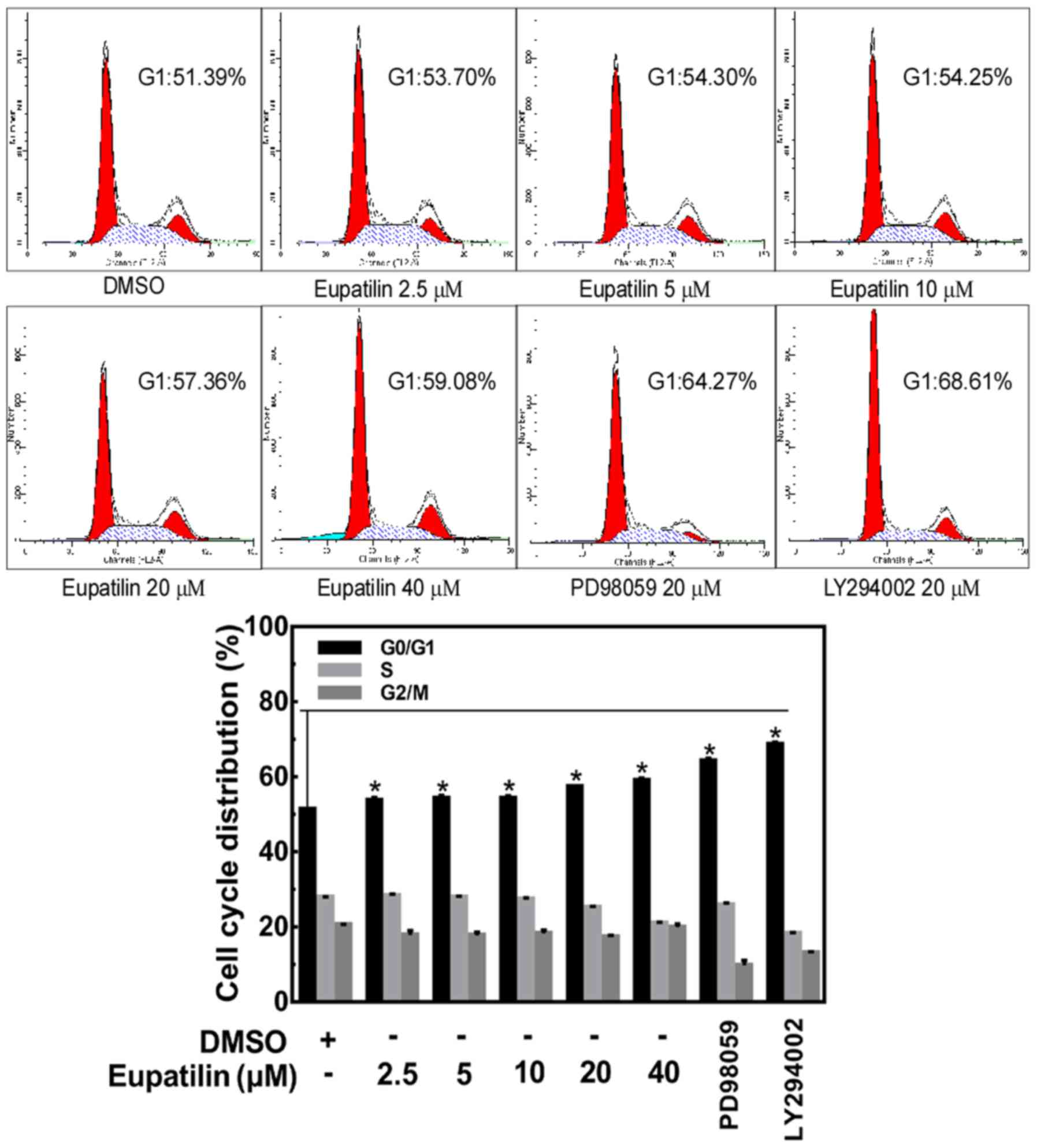
Eupatilin induces cell cycle arrest at
G0/G1 phase
To evaluate whether the inhibitory effect of
eupatilin on cell proliferation was related to induction of cell
cycle arrest, further experiments were performed in TE1 cells. TE1
cells were treated with different concentrations of eupatilin (0,
2.5, 5, 10, 20 or 40 µM), 20 µM PD98059 or 20 µM LY294002 for 24 h,
and the distribution of cells in the different phases of the cell
cycle was analyzed by flow cytometry. We found that there was an
accumulation of cells in the G0/G1 phase when
the cells were treated with eupatilin (Fig. 2). Similar changes also occurred in
the PD98059- and LY294002-treated groups. These results of flow
cytometry indicated that eupatilin induced
G0/G1 arrest in the TE1 cells.
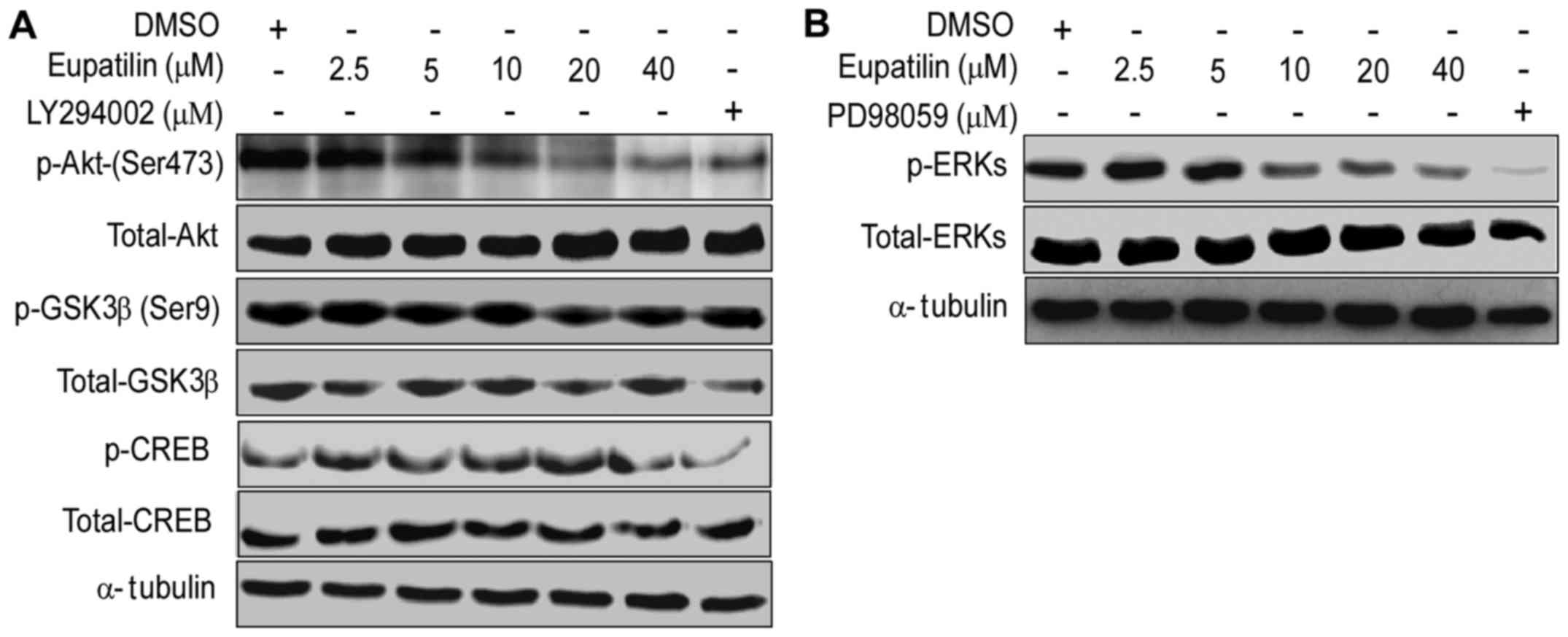
Eupatilin regulates the signaling
pathways of Akt/GSK3β/CREB and MAPK/ERK
To further explore the molecular mechanism
underlying the inhibitory effect of eupatilin on TE1 cell
proliferation, we detected the expression of key molecules in the
Akt/GSK3β and MAPK/ERK pathways. We observed that the treatments
with different concentrations of eupatilin in TE1 cells for 24 h
caused significant decreases in the levels of p-Akt, p-GSK3β,
p-CREB (Fig. 3A) and p-ERK
(Fig. 3B). Similar effects were
also observed following treatment with PD98059 or LY294002. These
data indicated that eupatilin inhibited TE1 cell proliferation by
regulating the signaling pathways of Akt/GSK3β and MAPK/ERK.
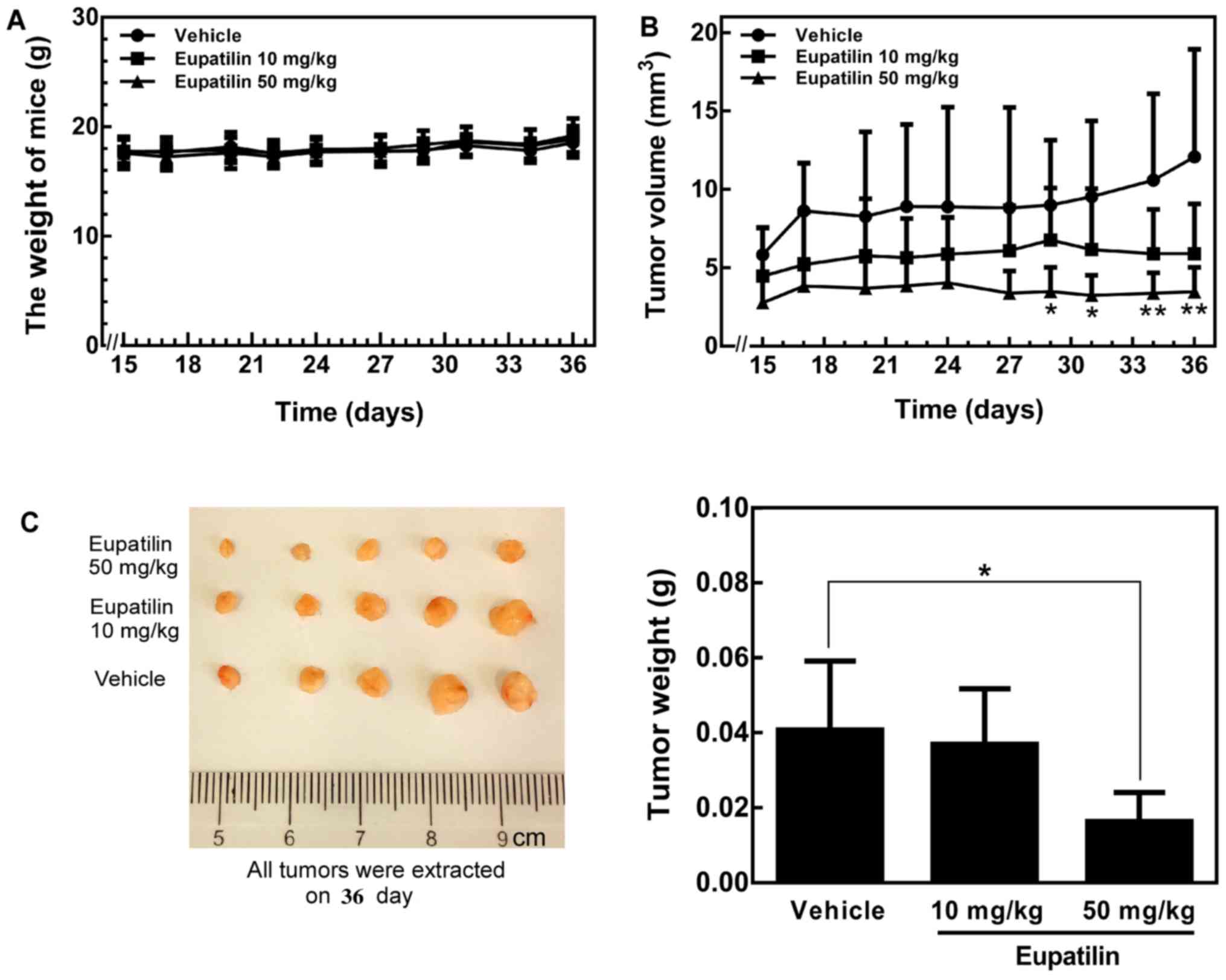
Eupatilin suppresses tumor growth in a
xenograft mouse model
To further investigate whether eupatilin could
suppress tumor growth in vivo, we evaluated the antitumor
activity of eupatilin in athymic nude mice implanted with TE1
cells. The results indicated a modest decrease in tumor volume in
the eupatilin-treated groups from the beginning to the end of the
experiment (Fig. 4B). Additionally,
the 50 mg/kg body weight eupatilin treatment caused significantly
greater inhibition compared with the lower dose treatment (10 mg/kg
body weight) (Fig. 4B). At the end
of the experiment, the mice were euthanized, the tumors were
extracted and weighed, and these results were confirmed (Fig. 4C). Notably, the average tumor size
in mice receiving 50 mg/kg eupatilin was significantly reduced
compared with tumors in the eupatilin-untreated mice (P<0.05 vs.
control). Meanwhile, in the 10 mg/kg eupatilin group, the average
tumor volume was smaller than that in the eupatilin-untreated group
without statistical significance (Fig.
4B). In addition, the body weights of all animals were
monitored and remained stable during the experiment (Fig. 4A), which suggested that the doses of
eupatilin used for the experiment were not severely toxic to the
mice. In the early stage, we carried out the acute toxicity test of
eupatilin in nude mice, and the maximum tolerated dose was 50
mg/kg. When the dosage of the drug was higher than 50 mg/kg, the
mice exhibited bradykinesia and weight loss and the mental state
was dispirited. When the dose reached 150 mg/kg, the mice died
overnight. In order to investigate whether eupatilin inhibits tumor
in a dose-dependent manner, we also set up a low dose of 10 mg/kg.
Overall, these data indicate that eupatilin can inhibit tumor
growth in vivo.
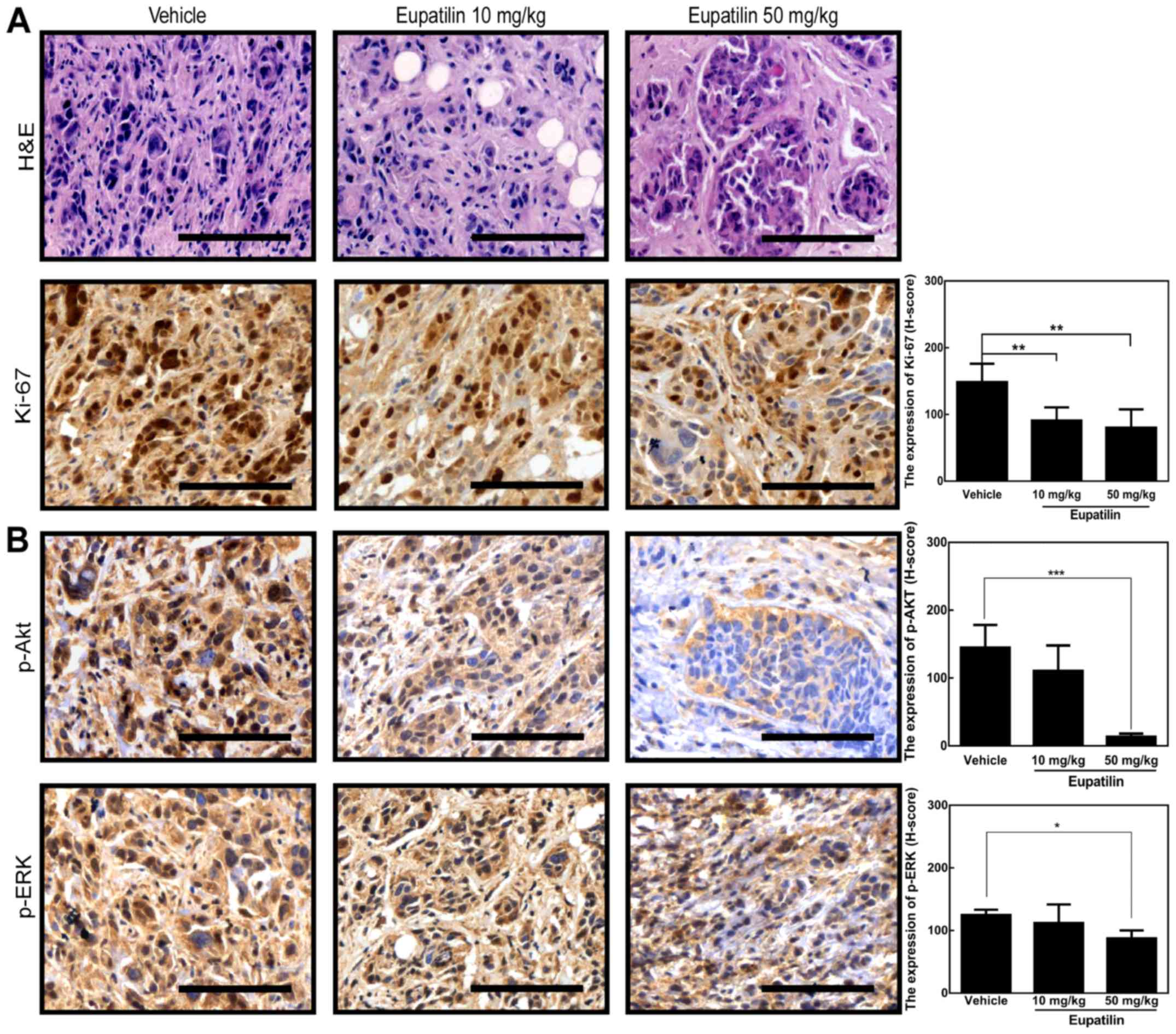
Eupatilin inhibits cell proliferation
and Akt and ERK signaling in tumors
To assess whether eupatilin suppressed tumor growth
in vivo by regulating the Akt and ERK signaling pathways, we
first detected the expression of Ki-67, a cellular biomarker of
proliferation, in the xenograft tumor tissues using
immunohistochemistry. We observed that 50 mg/kg eupatilin
significantly inhibited Ki-67 expression compared with vehicle and
10 mg/kg eupatilin (Fig. 5A). Next,
p-Akt and p-ERK were assessed by immunohistochemistry. We found
that the levels of Akt phosphorylation (Ser473) and ERK
phosphorylation in the 50 mg/kg eupatilin group were decreased
compared with the vehicle group, while phosphorylation changes in
the 10 mg/kg eupatilin group were observed to a lesser extent
(Fig. 5B). Taken together, these
data suggested that the inhibitory effect of eupatilin on tumor
growth in the mouse model may be associated with the Akt and ERK
signaling pathways.
Discussion
Both the incidence and mortality rates of esophageal
cancer in Eastern Asia are high, particularly in China. When the
disease is diagnosed, it is typically in the middle or late stage
of the cancer. The traditional treatment methods for esophageal
cancer are not reliable for improving prognosis. Thus, there is an
urgent need to identify more useful methods that can prevent or
treat the malignant cancer. Eupatilin, a flavone compound, has
potential anticancer activity (6,24). It
has been reported that eupatilin induces human renal cancer cell
apoptosis via ROS-mediated MAPK and PI3K/AKT signaling pathways
(25). Epatilin is a potential
chemopreventive agent in inhibition of skin cell transformation by
targeting PI3K, and is independent of the ERK-related signaling
pathway (ERK1/2, RSK2 and CREB) (24). However, to the best of our
knowledge, no study has explored the effect of eupatilin on human
esophageal cancer cells. Specifically, the antitumor effects
include inhibition of human esophageal cancer cell proliferation
and promotion of apoptosis in vivo has rarely been
investigated. Accordingly, in the present study, we confirmed the
antiproliferative effect of eupatilin in human esophageal cancer
cells not only in vitro but also in vivo. To further
investigate whether eupatilin can suppress tumor growth in
vivo, we evaluated the antitumor activity of eupatilin in
athymic nude mice implanted with human esophageal cancer cells.
Obviously, we found that the high dose of eupatilin (50 mg/kg body
weight) strongly suppressed tumor growth. In addition, we confirmed
that eupatilin suppressed the anchorage-independent growth of
esophageal cancer cells, which can reflect the extent of malignancy
in part. In the present study, we partially identified the
mechanism by which eupatilin may inhibit the proliferation of
esophageal cancer TE1 cells. The results showed that the percentage
of viable TE1 cells was reduced to 80% following treatment with
3.125 µM eupatilin for 48 h (Fig.
1B). This indicated that eupatilin has a significant
antiproliferation effect on human esophageal cancer at very low
concentrations. With the increase in the concentration of
eupatilin, the antiproliferation effect had an increasing
trend.
Uncontrolled cell proliferation is a typical
characteristic of cancer cells. The MEK/MAPK pathway plays a
significant role in the growth and survival of cancer cells
(26,27). When MAPK/ERK signaling is activated
by certain stimuli, p-ERK enters the cell nucleus, which results in
excessive cell proliferation, aggressive transformation and tumor
progression, through the activation of various oncogenes. Studies
have demonstrated that some natural plant ingredients or small
synthetic molecules can inhibit cancer cell growth. For instance,
curcumin inhibits cell proliferation in a time- and dose-dependent
manner in human leukemia THP-1 cells by acting on the Akt/mTOR and
RAF/MEK/ERK signaling pathways (28). In the present study, eupatilin
inhibited the proliferation and anchorage-independent growth of TE1
cells (Fig. 1). Meanwhile,
activation of Akt and MAPK/ERK were decreased by eupatilin in the
TE1 cells, which had the same results as treatment with LY294002 or
PD98059, which are known to be inhibitors of the PI3K/Akt and ERK
pathways, respectively (Fig. 3). We
observed that the treatments with different concentrations of
eupatilin in TE1 cells for 24 h may cause decreases in the levels
of p-CREB (Fig. 3). As known, CREB
activity is regulated by phosphorylation at Ser133, which is a
target of various kinases depending on the specific signaling
stimulus and cell type (29,30).
CREB is a validated phosphorylated substrate of ERK, PKA, and
Ca2+/calmodulin-dependent protein kinases (31). Akt activation can sequentially
activate the downstream mTOR complex1 (mTORC1) and GSK3β, both of
which play important roles in cancer development (29). GSK-3 activity promotes conditional
association of CREB (32,33). CREB is a transcription factor
involved in various biological processes, including cancer, whose
activity is promoted by GSK-3 phosphorylation (34). In the present study, treatment with
eupatilin, as well as LY294002, caused significant decreases in the
levels of p-Akt, p-GSK3β and p-CREB. When the Akt-GSK3β signaling
pathway is blocked, it causes decreases in the levels of p-CREB. At
the same time, CREB is a validated phosphorylated substrate of ERK
(31). MAPK/ERK is involved in
indirect regulation of CREB via activation of MSK1 (30). The present study found that the
antiproliferative effect of eupatilin was facilitated by targeting
the Akt/GSK3β and MAPK/ERK signaling pathways. When the MAPK/ERK
signaling pathway is blocked, it probably causes decreases in the
levels of p-CREB. Taken together, the present study suggests that
both the Akt/GSK3β and MAPK/ERK signaling pathways may cause
decreases in the levels of p-CREB.
Components of the MAPK and PI3K signaling pathways
are mediated by receptor tyrosine kinases in esophageal cancer
(35). Upon stimulation by growth
factors, cytokines and physical factors, PI3K is activated, and Akt
becomes phosphorylated. Researchers have found that Akt activation
promotes cell proliferation through multiple downstream targets
(GSK3β, for instance) that affect the cell cycle (36). Moreover, the MEK/ERK pathway also
participates in the regulation of cyclin D1 expression (21). In the present study, we found that
eupatilin induced cell cycle arrest at the
G0/G1 phase, which was associated with
eupatilin-mediated inhibition of the Akt/GSK3β and ERK pathways
(Figs. 2 and 3). Our previous study demonstrated that
eupatilin induced cell cycle arrest at the
G0/G1 phase by inhibiting the activity of
cyclin D1 in EGF-treated JB6 cells (24). However, G2/M arrest of
the cell cycle has been observed in eupatilin-treated human
endometrial carcinoma Hec1A cells (37). This difference between
G0/G1 and G2/M arrest may be
determined by differences in signaling responses in different cell
types, in cellular state, and in the environment at the time of
eupatilin treatment.
Molecular-targeted therapy for tumors has made
substantial progress, and new molecular-targeted drugs have
exhibited efficacy in clinical practice (38,39).
The epidermal growth factor receptor (EGFR) signaling pathway plays
important roles in tumor proliferation, survival and metastasis,
among other processes (19–22,40).
Activation of EGFR can promote multiple intracellular signaling
cascades involved in tumor growth, and thus these signaling
molecules have been focused on as targets for cancer therapy.
Raf/MEK/MAPK and PI3K/Akt/PTEN/mTOR are both downstream signaling
pathways of EGFR. ERK and Akt can regulate cell cycle-related
proteins, which generally results in cell growth and proliferation
(41,42). In our xenograft mouse model in the
present study, we found that the higher dose of eupatilin (50 mg/kg
body weight) strongly suppressed tumor growth (Fig. 4B and C). Meanwhile, phosphorylation
of Akt and ERK were suppressed in the high-dose eupatilin treatment
group compared with the vehicle or low-dose eupatilin (10 mg/kg
body weight) treatment groups (Fig.
5). These findings indicated that eupatilin regulated the Akt
and ERK pathways to inhibit tumor growth.
In conclusion, eupatilin suppressed the
proliferation of esophageal cancer TE1 cells, cause cell cycle
arrest at G0/G1 phase, and suppressed TE1
xenograft tumor growth by decreasing activation of the Akt and
MAPK/ERK signaling pathways. In future studies, we aim to explore
the underlying mechanisms in more detail to identify the potential
targets of eupatilin in the two pathways, in order to determine
possible therapeutic targets in esophageal cancer.
Acknowledgements
Not applicable.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant nos. 81372269, 81472324
and 81572812) and the Science Foundation of Henan Education
Department (grant no. 14A310011).
Availability of data and materials
The datasets used during the present study are
available from the corresponding author upon reasonable
request.
Authors' contributions
JiZ, KL, XW and YZhu conceived and designed the
study. XW, YZhu, LZ, YZhao, YS, XChen, YX, FL and YJ performed the
experiments. JL, YH, XChang, JuZ, XL, KL, MZ and ZD helped perform
the data analyses and interpreted the results. All authors read and
approved the manuscript and agree to be accountable for all aspects
of the research in ensuring that the accuracy or integrity of any
part of the work are appropriately investigated and resolved.
Ethics approval and consent to
participate
All animal studies conformed to guidelines approved
by the Research Ethics Committee of Zhengzhou University
(Zhengzhou, Henan, China).
Consent for publication
Not applicable.
Competing interests
The authors state that they have no competing
interests.
References
|
1
|
Pennathur A, Gibson MK, Jobe BA and
Luketich JD: Oesophageal carcinoma. Lancet. 381:400–412. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Xiao Z, Peng Z, Peng M, Yan W, Ouyang Y
and Zhu H: Flavonoids health benefits and their molecular
mechanism. Mini Rev Med Chem. 11:169–177. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Li ZG, Shimada Y, Sato F, Maeda M, Itami
A, Kaganoi J, Komoto I, Kawabe A and Imamura M: Inhibitory effects
of epigallocatechin-3-gallate on N-nitrosomethylbenzylamine-induced
esophageal tumorigenesis in F344 rats. Int J Oncol. 21:1275–1283.
2002.PubMed/NCBI
|
|
4
|
Kumar S and Pandey AK: Chemistry and
biological activities of flavonoids: An overview.
ScientificWorldJournal. 2013:1627502013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ye F, Zhang GH, Guan BX and Xu XC:
Suppression of esophageal cancer cell growth using curcumin,
(−)-epigallocatechin-3-gallate and lovastatin. World J
Gastroenterol. 18:126–135. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kim MJ, Kim DH, Na HK, Oh TY, Shin CY and
Surh Ph D Professor YJ: Eupatilin, a pharmacologically active
flavone derived from Artemisia plants, induces apoptosis in
human gastric cancer (AGS) cells. J Environ Pathol Toxicol Oncol.
24:261–269. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Seo HJ, Park KK, Han SS, Chung WY, Son MW,
Kim WB and Surh YJ: Inhibitory effects of the standardized extract
(DA-9601) of Artemisia asiatica Nakai on phorbol
ester-induced ornithine decarboxylase activity, papilloma
formation, cyclooxygenase-2 expression, inducible nitric oxide
synthase expression and nuclear transcription factor kappa B
activation in mouse skin. Int J Cancer. 100:456–462. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Min SW, Kim NJ, Baek NI and Kim DH:
Inhibitory effect of eupatilin and jaceosidin isolated from
Artemisia princeps on carrageenan-induced inflammation in
mice. J Ethnopharmacol. 125:497–500. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yun C, Jung Y, Chun W, Yang B, Ryu J, Lim
C, Kim JH, Kim H and Cho SI: Anti-inflammatory effects of
Artemisia leaf extract in mice with contact dermatitis in
vitro and in vivo. Mediators Inflamm. 2016:80275372016.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wang Y, Hou H, Li M, Yang Y and Sun L:
Anticancer effect of eupatilin on glioma cells through inhibition
of the Notch-1 signaling pathway. Mol Med Rep. 13:1141–1146. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Steelman LS, Chappell WH, Abrams SL, Kempf
RC, Long J, Laidler P, Mijatovic S, Maksimovic-Ivanic D, Stivala F,
Mazzarino MC, et al: Roles of the Raf/MEK/ERK and
PI3K/PTEN/Akt/mTOR pathways in controlling growth and sensitivity
to therapy-implications for cancer and aging. Aging (Albany NY).
3:192–222. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Sewing A, Wiseman B, Lloyd AC and Land H:
High-intensity Raf signal causes cell cycle arrest mediated by
p21Cip1. Mol Cell Biol. 17:5588–5597. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Woods D, Parry D, Cherwinski H, Bosch E,
Lees E and McMahon M: Raf-induced proliferation or cell cycle
arrest is determined by the level of Raf activity with arrest
mediated by p21Cip1. Mol Cell Biol. 17:5598–5611. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Knauf JA, Sartor MA, Medvedovic M,
Lundsmith E, Ryder M, Salzano M, Nikiforov YE, Giordano TJ,
Ghossein RA and Fagin JA: Progression of BRAF-induced thyroid
cancer is associated with epithelial-mesenchymal transition
requiring concomitant MAP kinase and TGFβ signaling. Oncogene.
30:3153–3162. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Clair T, Miller WR and Cho-Chung YS:
Prognostic significance of the expression of a ras protein with a
molecular weight of 21,000 by human breast cancer. Cancer Res.
47:5290–5293. 1987.PubMed/NCBI
|
|
16
|
Janes PW, Daly RJ, deFazio A and
Sutherland RL: Activation of the Ras signalling pathway in human
breast cancer cells overexpressing erbB-2. Oncogene. 9:3601–3608.
1994.PubMed/NCBI
|
|
17
|
Bos JL: Ras oncogenes in human cancer: A
review. Cancer Res. 49:4682–4689. 1989.PubMed/NCBI
|
|
18
|
Wee P and Wang Z: Epidermal growth factor
receptor cell proliferation signaling pathways. Cancers (Basel).
9:2017.PubMed/NCBI
|
|
19
|
Kaul R, Saha P, Saradhi M, Prasad RL,
Chatterjee S, Ghosh I, Tyagi RK and Datta K: Overexpression of
hyaluronan-binding protein 1 (HABP1/p32/gC1qR) in HepG2 cells leads
to increased hyaluronan synthesis and cell proliferation by
up-regulation of cyclin D1 in AKT-dependent pathway. J Biol Chem.
287:19750–19764. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lv C, Qin W, Zhu T, Wei S, Hong K, Zhu W,
Chen R and Huang C: Ophiobolin O isolated from Aspergillus
ustus induces G1 arrest of MCF-7 cells through interaction with
AKT/GSK3β/cyclin D1 signaling. Mar Drugs. 13:431–443. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wang HY, Yang SL, Liang HF and Li CH: HBx
protein promotes oval cell proliferation by up-regulation of cyclin
D1 via activation of the MEK/ERK and PI3K/Akt pathways. Int J Mol
Sci. 15:3507–3518. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Thompson KN, Whipple RA, Yoon JR, Lipsky
M, Charpentier MS, Boggs AE, Chakrabarti KR, Bhandary L, Hessler
LK, Martin SS and Vitolo MI: The combinatorial activation of the
PI3K and Ras/MAPK pathways is sufficient for aggressive tumor
formation, while individual pathway activation supports cell
persistence. Oncotarget. 6:35231–35246. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Bode AM and Dong Z: Signal transduction
and molecular targets of selected flavonoids. Antioxid Redox
Signal. 19:163–180. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Li F, Tao Y, Qiao Y, Li K, Jiang Y, Cao C,
Ren S, Chang X, Wang X, Wang Y, et al: Eupatilin inhibits
EGF-induced JB6 cell transformation by targeting PI3K. Int J Oncol.
49:1148–1154. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhong WF, Wang XH, Pan B, Li F, Kuang L
and Su ZX: Eupatilin induces human renal cancer cell apoptosis via
ROS-mediated MAPK and PI3K/AKT signaling pathways. Oncol Lett.
12:2894–2899. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Blackhall FH, Pintilie M, Michael M,
Leighl N, Feld R, Tsao MS and Shepherd FA: Expression and
prognostic significance of kit, protein kinase B, and
mitogen-activated protein kinase in patients with small cell lung
cancer. Clin Cancer Res. 9:2241–2247. 2003.PubMed/NCBI
|
|
27
|
Huynh H, Nguyen TT, Chow KH, Tan PH, Soo
KC and Tran E: Over-expression of the mitogen-activated protein
kinase (MAPK) kinase (MEK)-MAPK in hepatocellular carcinoma: Its
role in tumor progression and apoptosis. BMC Gastroenterol.
3:192003. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Guo Y, Shan Q, Gong Y, Lin J, Shi F, Shi R
and Yang X: Curcumin induces apoptosis via simultaneously targeting
AKT/mTOR and RAF/MEK/ERK survival signaling pathways in human
leukemia THP-1 cells. Pharmazie. 69:229–233. 2014.PubMed/NCBI
|
|
29
|
de Groot RP, Ballou LM and Sassone-Corsi
P: Positive regulation of the cAMP-responsive activator CREM by the
p70 S6 kinase: An alternative route to mitogen-induced gene
expression. Cell. 79:81–91. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Xing J, Ginty DD and Greenberg ME:
Coupling of the RAS-MAPK pathway to gene activation by RSK2, a
growth factor-regulated CREB kinase. Science. 273:959–963. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Tan Y, Rouse J, Zhang A, Cariati S, Cohen
P and Comb MJ: FGF and stress regulate CREB and ATF-1 via a pathway
involving p38 MAP kinase and MAPKAP kinase-2. EMBO J. 15:4629–4642.
1996.PubMed/NCBI
|
|
32
|
Fiol CJ, Williams JS, Chou CH, Wang QM,
Roach PJ and Andrisani OM: A secondary phosphorylation of CREB341
at Ser129 is required for the cAMP-mediated control of gene
expression. A role for glycogen synthase kinase-3 in the control of
gene expression. J Biol Chem. 23:32187–32193. 1994.
|
|
33
|
Wang Z, Iwasaki M, Ficara F, Lin C,
Matheny C, Wong SH, Smith KS and Cleary ML: GSK-3 promotes
conditional association of CREB and its coactivators with MEIS1 to
facilitate HOX-mediated transcription and oncogenesis. Cancer Cell.
17:597–608. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Horike N, Sakoda H, Kushiyama A, Ono H,
Fujishiro M, Kamata H, Nishiyama K, Uchijima Y, Kurihara Y,
Kurihara H and Asano T: AMP-activated protein kinase activation
increases phosphorylation of glycogen synthase kinase 3beta and
thereby reduces cAMP-responsive element transcriptional activity
and phosphoenolpyruvate carboxykinase C gene expression in the
liver. J Biol Chem. 283:33902–33910. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Lin DC, Hao JJ, Nagata Y, Xu L, Shang L,
Meng X, Sato Y, Okuno Y, Varela AM, Ding LW, et al: Genomic and
molecular characterization of esophageal squamous cell carcinoma.
Nat Genet. 46:467–473. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Manning BD and Cantley LC: AKT/PKB
signaling: Navigating downstream. Cell. 129:1261–1274. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Cho JH, Lee JG, Yang YI, Kim JH, Ahn JH,
Baek NI, Lee KT and Choi JH: Eupatilin, a dietary flavonoid,
induces G2/M cell cycle arrest in human endometrial cancer cells.
Food Chem Toxicol. 49:1737–1744. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Li K and Li J: Current molecular targeted
therapy in advanced gastric cancer: A comprehensive review of
therapeutic mechanism, clinical trials, and practical application.
Gastroenterol Res Pract. 2016:41056152016. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Wang H, Xu T, Jiang Y, Xu H, Yan Y, Fu D
and Chen J: The challenges and the promise of molecular targeted
therapy in malignant gliomas. Neoplasia. 17:239–255. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Borlak J, Meier T, Halter R, Spanel R and
Spanel-Borowski K: Epidermal growth factor-induced hepatocellular
carcinoma: Gene expression profiles in precursor lesions, early
stage and solitary tumours. Oncogene. 24:1809–1819. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Adlung L, Kar S, Wagner MC, She B,
Chakraborty S, Bao J, Lattermann S, Boerries M, Busch H, Wuchter P,
et al: Protein abundance of AKT and ERK pathway components governs
cell type-specific regulation of proliferation. Mol Syst Biol.
13:9042017. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Saez-Rodriguez J, MacNamara A and Cook S:
Modeling signaling networks to advance new cancer therapies. Annu
Rev Biomed Eng. 17:143–163. 2015. View Article : Google Scholar : PubMed/NCBI
|