Introduction
Gastric cancer is a common malignancy of the
gastrointestinal tract and has a high mortality rate worldwide.
According to the World Health Organization, gastric cancer causes
74,000 deaths (~9.7% of total cancer mortality) per year worldwide.
Although the incidence of gastric cancer has rapidly declined in
recent decades, it is currently the third leading cause of cancer
mortality in the world (GLOBOCAN 2012) (1). Surgical resection with radical lymph
node dissection still plays an important role in gastric cancer
therapy. Chemotherapy or targeted therapies are alternative
therapeutic strategies for incurable gastric cancer or tumor
recurrence after surgical resection. However, in the biological
molecular field, gastric carcinogenesis remains poorly
understood.
Hepatocyte growth factor (HGF), which is secreted by
mesenchymal cells, is a potent mitogenic factor for hepatocytes.
HGF binds to the c-Met receptor and activates tyrosine kinase
signaling pathways. HGF targets and acts on epithelial cells and
endothelial cells and is also associated with cell migration,
proliferation and angiogenesis (2–4).
Elevation of serum HGF levels and overexpression of the c-Met gene
are often found in patients with more advanced tumors or metastasis
(5,6). The correlation between COX-2 and c-Met
expression and human gastric cancer was found to be statistically
significant. In addition, the overexpression of COX-2 and elevation
of prostaglandin E2 were found to be involved in the
growth and metastasis of gastric cancer (2). These studies clearly show that HGF
targets c-Met and regulates its biological function in gastric
cancer.
Notch1 can act as an oncogene in breast, gastric,
pancreatic and colon cancers, or as a tumor-suppressor gene in
neuroendocrine tumors and skin cancer (7). Jagged1 is an intercellular ligand that
activates the Notch1 signaling pathway (8). The activity of Notch ligand Jagged1 is
associated with a poor prognosis in gastric cancer patients after
surgery (3). Notch1 intracellular
domain (N1IC), the intracellular activated form of the Notch1
receptor, translocates to the COX-2 promoter in the nucleus by
binding C promoter binding factor 1. Overexpression of N1IC was
demonstrated to increase cell proliferation and migration (9). COX-2 expression promotes the
production of prostaglandin E2. The interaction between
intercellular Jagged1 and the Notch1 receptor induces COX-2
expression, which causes growth and metastasis in gastric
cancer.
Notably, both the HGF/c-Met and Jagged1/Notch1
pathways are associated with COX-2 activity. COX-2 overexpression
was found to be associated with tumor invasion depth (10), lymphovascular invasion, lymph node
metastasis and more advanced gastric cancers (11). Elevated levels of COX-2 protein were
found in gastric cancer tumors. Moreover, COX-2-specific inhibitor
NS-398 and COX-2 knockdown suppressed the growth of human gastric
cancer SC-M1 cells (12). However,
the role of the interaction between the HGF/c-Met and Notch1
signaling pathway in gastric cancer cells remains obscure. The
present study aimed to elucidate the influence of HGF on
Jagged1/Notch signal transduction in SC-M1 gastric cancer
cells.
Materials and methods
Surgical specimens
From January 2003 to December 2005, human gastric
cancer samples were obtained from gastric cancer patients who
underwent gastric surgery at the Department of Surgery, Taipei
Veterans General Hospital. A total of 91 gastric cancer patients
(including 59 males and 32 females, age from 33–84 years) were
enrolled in this study and informed consent was obtained from all
patients. Gastric tumors and adjacent gastric tissues were fixed
overnight at 4°C with 4% neutral buffered paraformaldehyde,
dehydrated, cleared with Histo-Clear II (National Diagnostics,
Atlanta, GA, USA) and wax embedded. Five-micron sections were used
for hematoxylin and eosin (H&E) staining and
immunohistochemical staining. This study was approved by the
Institutional Review Board of Tapei Veterans General Hospital and
informed consent was provided by all subjects.
Immunohistochemical staining of c-Met
and Jagged1
The localization of c-Met receptor and Jagged1
ligand in gastric tissues was detected using an
avidin-biotin-peroxidase complex (ABC) technique according to a
protocol established and used for gastric cancer specimens
(2,3). Immunostaining of HGF/c-Met receptors
in gastric cancer tissue was performed. Briefly, tissue sections
were first microwaved in a sodium citrate buffer (10 mM, pH 6.0)
and, were then treated with peroxidase blocking solution (3%
H2O2) for 5–10 min. Serum blocking solution
(Histostain SP kit; Zymed Laboratories Inc., South San Francisco,
CA, USA) was applied to remove endogenous peroxidase activity and
to reduce nonspecific background staining. The antibodies used for
tissue sections were as follows: polyclonal rabbit anti-c-Met
(C-28) (1:200; cat. no. sc-161; Santa Cruz Biotechnology Inc.,
Dallas, TX, USA) at a 1:500 dilution and polyclonal rabbit
anti-human Jagged1 antibody (cat. no. sc8303; Santa Cruz
Biotechnology) at a 1:50 dilution. Slides were incubated overnight
at 4°C in a moist chamber. The tissue sections were subsequently
treated with post-primary block reagent: NovoLink polymer detection
system (cat. no. RE7140-K; Leica Biosystems, Wetzlar, Germany) for
10 min, streptavidin peroxidase-conjugated for 10 min, AEC
substrate chromogen for 5–10 min, and then counterstained with
hematoxylin for 5 min. Pre-immune rabbit IgG was used as a negative
control. The distribution pattern of chromogen in tumor cells was
examined by an experienced pathologist (A.F.-Y. Li).
The distribution of c-Met and Jagged1 proteins in
human gastric cancer specimens was evaluated by a semi-quantitative
system to calculate and estimate within the following arbitrary
ranges: (−) no positive cells; (+) 1–25%; (++) 26–75%; and (+++)
>75%.
Cell culture and transfection
Our previous study tested the Notch1 expression in
SC-M1, AGS, NUGC3 and KATO III gastric cancer cell lines. The
expression level of N1IC in SC-M1 gastric cancer cells was much
more abundant than that in the other gastric cancer cell lines
(3). Therefore, we chose SC-M1
cells for further experimentation. The overexpression of N1IC
increased Notch activity in SC-M1 cells. Human gastric cancer SC-M1
cells were cultured in RPMI-1640 medium (Gibco; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) with 10% fetal bovine serum
(FBS; HyClone Laboratory; GE Healthcare Life Sciences, Logan, UT,
USA) and 0.1% gentamicin (Gibco; Thermo Fisher Scientific, Inc.).
Our previous study showed that SC-M1 gastric cancer cell lines also
express other Notch family members (Notch 1, 2, 3 and 4). N1IC
expression was found to be significantly higher than the expression
of other Notch family members (13,14).
Therefore, we choose N1IC for further experimentation. To establish
stable expression of an HA-N1IC fusion protein in SC-M1 cells
(SC-M1/HA-N1IC), a pcDNA-HA-N1IC expression plasmid was transfected
into SC-M1 cells, and expression levels were screened by western
blot analysis according to a previous established protocol
(3). For the control, the pcDNA3
vector was also transfected into the SC-M1 cell line. The
SC-M1/HA-N1IC and SC-M1/pcDNA3 cell lines were selected with G418
(400 µg/ml; CalbioChem; EMD/Merck KGaA).
Drug treatment
Human gastric cancer SC-M1, SC-M1/HA-N1IC and
SC-M1/pcDNA3 cells were treated with HGF alone (0–25 ng/ml; Gibco;
Thermo Fisher Scientific, Inc.), NS398 alone (50 µM; Cayman
Chemical Company, Ann Arbor, MI, USA), or HGF in combination with
NS398. HGF promotes gastric cancer cell growth, and NS398 is a
COX-2-specific inhibitor that has anti-inflammatory and analgesic
effects. To assess the effects on cell growth, the gastric cancer
cell lines were treated with HGF and NS398 alone, or combined HGF
and NS398 for 24, 48 or 72 h. To assess the effects on migration,
cells were treated with HGF (25 ng/ml) for 64 h, and cell migration
was observed at 8, 32 and 64 h. For western blot analysis, the
gastric cancer cell lines were incubated with HGF (25 ng/ml), and
protein samples were collected at 1, 2, 4 and 24 h to analyze
c-Met, Jagged1 and COX-2 protein expression.
MTT assay
SC-M1 cells were seeded into a 96-well plate at a
density of 1.2×104 cells/well and incubated for 24 h.
After treatment with HGF and/or NS398 for 24, 48, or 72 h, the
medium was replaced with 100 µl of medium with
3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyl-tetrazolium bromide
(MTT). Then, the plate was incubated in a 37°C incubator. After
discarding the medium, 100 µl dimethyl sulfoxide (DMSO) was added.
The optical density was measured at an absorption wavelength of 570
nm by an ELISA reader.
Wound healing migration study
The migration ability of the drug- or
vehicle-treated gastric cancer cells was evaluated by wound healing
migration analysis. Culture-inserts (Ibidi-cells in focus, Munich,
Germany) were placed in 6-well dishes, and 8×104 SC-M1
cells were seeded onto the inserts and incubated for 24 h. Inserts
were removed and gently washed to remove floating cells. After
addition of 0.5% FBS medium with or without HGF (25 ng/ml), images
of cell migration were captured at 8, 32 and 64 h. The percentage
of the migration area was compared to the initial area after
removing the insert. The migration area of gastric cancer cells was
measured by ImageJ version 1.46 software (National Institutes of
Health, Bethesda, MD, USA).
Transwell migration assay
Cell migration assays were analyzed using 24-well
Transwell inserts with 5-µm pores (Corning Costar, Cambridge, MA,
USA). Approximately 5×104 cells were resuspended in 100
µl of low serum (0.1% FBS)-containing medium and loaded into the
inserts with low serum-containing medium in the lower chamber.
After incubating for 12 h, either DMSO or HGF in low
serum-containing medium at indicated concentrations was added to
the inserts, which were then immediately shifted to new wells with
high serum (20% FBS)-containing medium to allow migration in a
humidified 37°C incubator with 5% CO2 for 24 h. SC-M1,
SC-M1/HA-N1IC and SC-M1/pcDNA3 cells were then treated with HGF
and/or NS398. After incubation for 24 h, the inserts were removed,
fixed in ice-cold methanol for 20 min, and then stained using Liu
staining solutions (Tonyar Biotech, Inc., Taoyuan, Taiwan). After
staining, the cells that remained on the upper sides of the inserts
were carefully wiped-off using rinsed cotton swabs. The migrated
cells found on the bottom of the inserts were then imaged (at ×100
magnification) and counted. Migrated cells from 10 random fields of
each inserts were counted by ImageJ version 1.46 software (National
Institutes of Health) and normalized.
Western blot analysis
SC-M1 cells were treated with or without HGF (25
ng/ml) for 1, 2 or 4 h. The cells were mixed with NP-40 lysis
buffer, and then added with mammalian protein extraction reagents
(M-PER) in an ice bath. Cell lysates were collected after
centrifugation at 23,220 × g for 10 min at 4°C, and cell lysates
were heated at 90°C for 5 min. Protein concentrations were
quantitatively determined using Bradford method. Protein (30 µg)
samples were electrophoresed on 6% (for c-Met and Jagged1) or 7.5%
(for COX-2) SDS-polyacrylamide gels. Each gel was then transferred
to a polyvinylidene fluoride (PVDF) membrane (Immobilon-P; EMD
Millipore, Bedford, MA, USA). The PVDF membrane was blocked with 5%
non-fat milk for 30 min and probed with rabbit anti-c-Met antibody
(C-28) (cat. no. sc-161; Santa Cruz Biotechnology) at a 1:250
dilution, rabbit anti-Jagged1 antibody (cat. no. sc8303; Santa Cruz
Biotechnology) at a 1:500 dilution, or mouse anti-COX-2 antibody
(cat. no. 160112; Cayman Chemical Company) overnight at 4°C. After
washing with Tris-buffered saline containing 0.1% Tween-20 (TBS-T),
the membranes were incubated with secondary anti-rabbit antibody
(cat. no. 7074; Cell Signaling Technology, Danvers, MA, USA) at a
1:2,000 dilution. Enhanced protein bands were detected by a
chemiluminescence kit (Amersham Life Science, Piscataway, NJ,
USA).
Statistical analyses
All statistical analyses were carried out using SPSS
software version 18.0 (SPSS, Inc., Chicago, IL, USA).
Clinicopathological differences were compared with Chi-square
tests, and the correlation between Jagged1 and c-Met expression in
each specimen was estimated with linear regression modeling. A
P-value <0.05 was considered to indicate a statistically
significant result.
Results
Expression of c-Met and Jagged1
expression in gastric cancer tissue in relation to survival
To evaluate the correlation between c-Met and
Jagged1 in gastric cancer specimens, we collected samples from 91
gastric cancer patients in our study. The expression of c-Met and
Jagged1 in gastric cancer tissue was analyzed by
immunohistochemical staining of the 91 gastric cancer samples.
While the positive c-Met expression rate was 71.4% (65/91), the
positive Jagged1 expression rate was 76.9% (70/91). Patients with
positive c-Met expression had poorer overall survival than patients
with negative c-Met expression (P=0.043) (Fig. 1A). Our previous study showed that
patients with positive Jagged1 expression also had poorer prognosis
than patients with negative Jagged1 expression (3). Jagged1 expression was positively
correlated with c-Met expression in the human gastric cancer
tissues (P=0.004) (Fig. 1B). Taken
together, these results indicate a positive correlation between the
c-Met and Jagged1 signaling pathways.
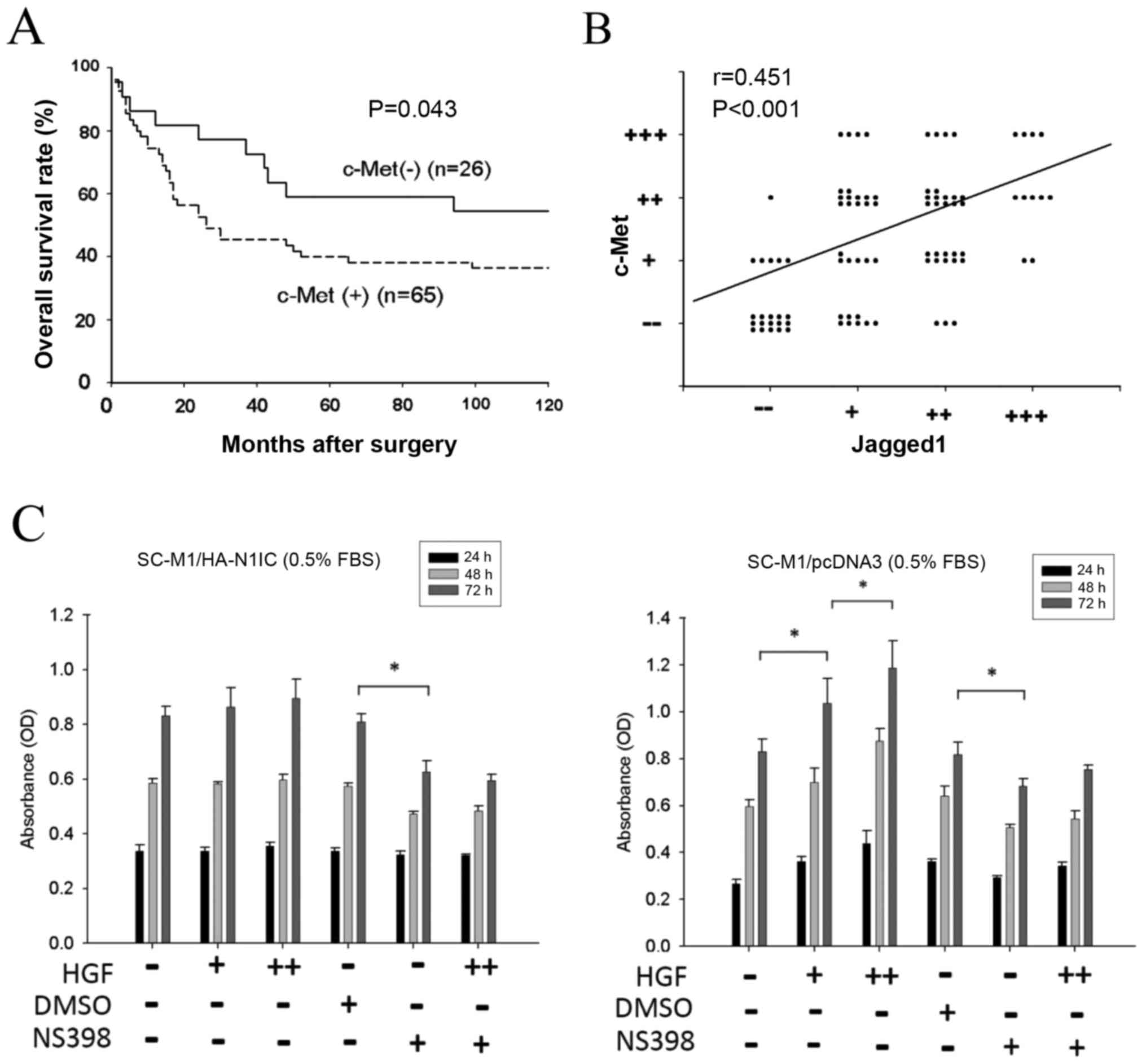 | Figure 1.Clinical relevance of c-Met and
Jagged1 expression in gastric cancer tissues. (A) Approximately 91
gastric cancer tissues were collected for immunohistochemical
staining of c-Met and Jagged1. Patients with positive c-Met
expression had poorer overall survival than patients without c-Met
expression (P=0.043). (B) Statistical correlation between c-Met and
Jagged1 expression was compared using the linear regression model
(P=0.004). (C) Effect of HGF and NS398 on the growth of
SC-M1/HA-N1IC and SC-M1/pcDNA3 cells. Transfected SC-M1 cells were
cultured in the presence or absence of HGF (0, 5 or 25 ng/dl) and
NS398 (0 or 50 µM) for 3 days, and the cell growth was measured by
MTT assay. Representative data are the average of 6 replicates.
(HGF dosage: -, +, and ++ denotes 0, 5, and 25 ng/dl, respectively;
NS398 dosage: - and + denotes 0 and 50 µM, respectively; *P<0.01
at 72 h). HGF, hepatocyte growth factor; N1IC, Notch1 intracellular
domain. |
Effect of HGF stimulation on the
growth of SC-M1/pcDNA3 and SC-M1/HA-N1IC cells
To evaluate the effect of HGF on the Notch1/Jagged1
pathway, N1IC-overexpressing (SC-M1/HA-N1IC) and control
(SC-M1/pcDNA3) human gastric cancer SC-M1 cells were used in our
experiment. To reduce the influence of the growth factors in FBS,
the concentration of FBS was tapered to 0.5%. Cells were treated
with various concentration of HGF (0, 5 or 25 ng/ml) (−, + and ++,
respectively in the figures) and/or NS398 (0 or 50 µM) (− and +,
respectively in the figures). The cell growth of SC-M1/pcDNA3 cells
was increased in proportion to the elevation of HGF concentration.
However, this phenomenon was not observed in the SC-M1/HA-N1IC
cells. The COX-2-specific inhibitor NS398 blocked cell growth in
both SC-M1/HA-N1IC and SC-M1/pcDNA3 cells, and HGF-stimulated cell
growth in SC-M1/pcDNA3 was also blocked by NS398 (Fig. 1C). These data showed that HGF can
enhance the proliferation ability of gastric cancer with lower
Notch1 (N1IC) activity. However, the effect of HGF treatment on
proliferation in gastric cancer cells with constitutively
overexpressed Notch1 (N1IC) was not obvious, such that the
COX-2-specific inhibitor NS398 repressed the migration ability in
gastric cancer cells with both lower and higher Notch1 (N1IC)
activity.
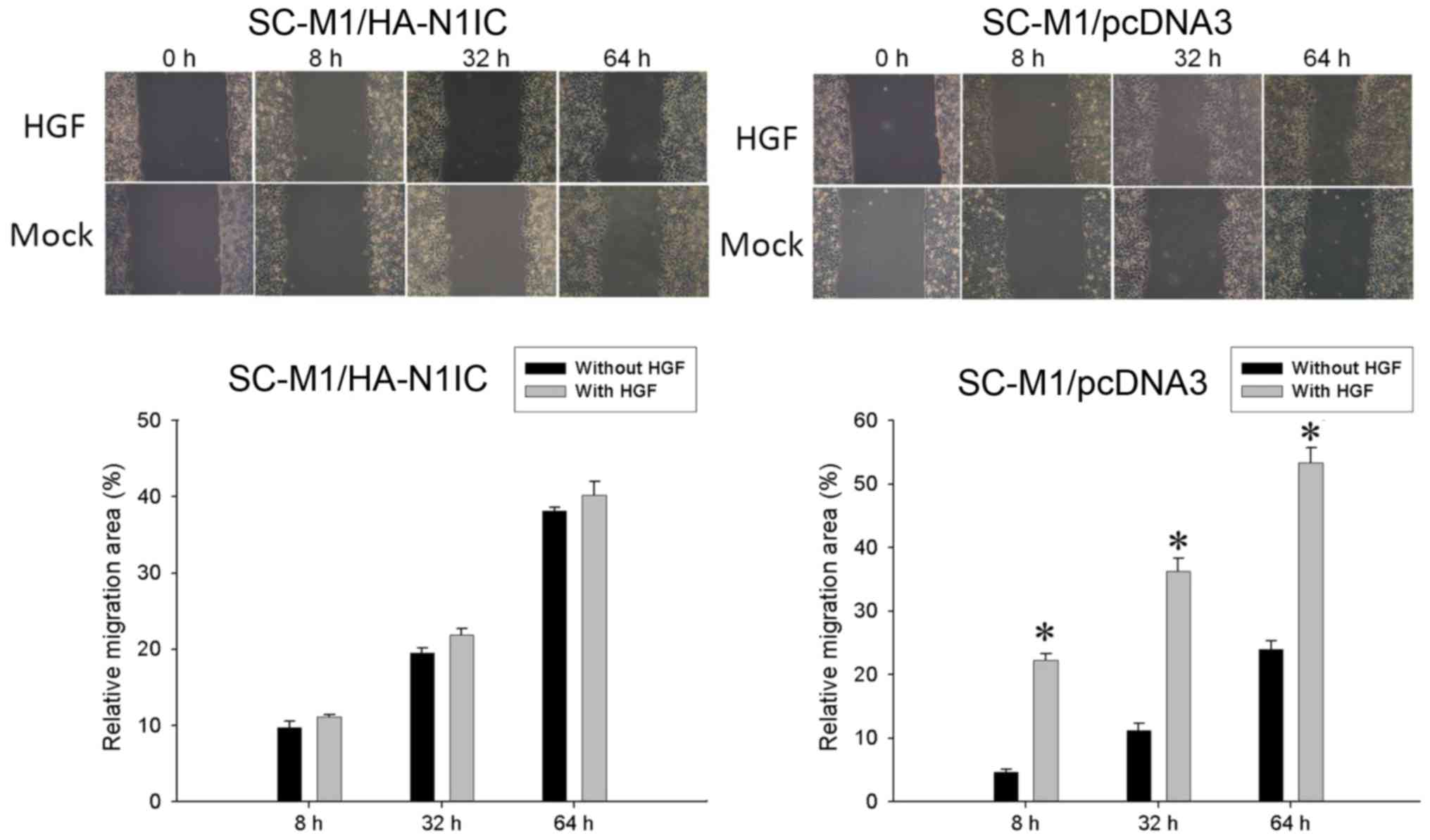
Effect of HGF treatment on wound
healing migration and Transwell migration assays in SC-M1/pcDNA3
and SC-M1/HA-N1IC cells
Next, we investigated the migration ability of
gastric cancer cells using wound healing migration and Transwell
migration assays. In the wound healing migration study,
SC-M1/HA-N1IC and SC-M1/pcDNA3 cells were treated with HGF (0 or 25
ng/ml). The relative migration area was increased after HGF
treatment in SC-M1/pcDNA3 cells compared to HGF treatment in
SC-M1/HA-N1IC cells (Fig. 2).
Furthermore, we investigated the migration ability using a
Transwell migration assay. Compared to the migration ability of
SC-M1/pcDNA3 cells, SC-M1/HA-N1IC cell migration increased
independently of HGF. The migration ability of SC-M1/pcDNA3 cells
was increased after HGF treatment. However, there was no
significant difference in the migration ability of SC-M1/HA-N1IC
cells before and after HGF treatment (Fig. 3). NS398 treatment reduced the
migration ability in the SC-M1/pcDNA3 and SC-M1/HA-N1IC cells
(Fig. 4). These results suggest
that HGF can enhance the migration ability of gastric cancers with
lower Notch1 (N1IC) activity. However, the effect of HGF on gastric
cancer cells with constitutively overexpressed Notch1 (N1IC) was
not obvious. The COX-2-specific inhibitor NS398 can repress the
migration ability in gastric cancer cells with both low and
overexpressed N1IC.
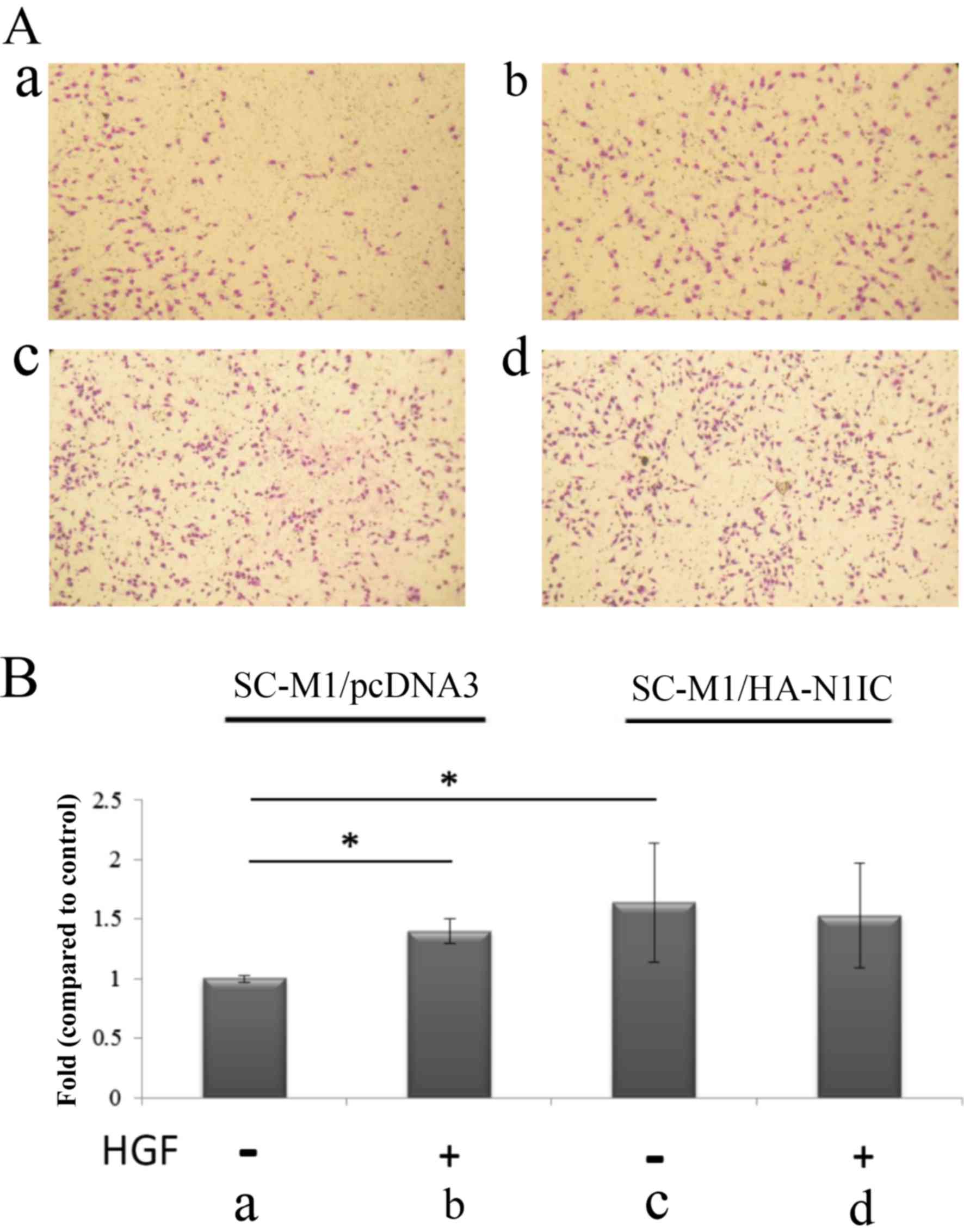
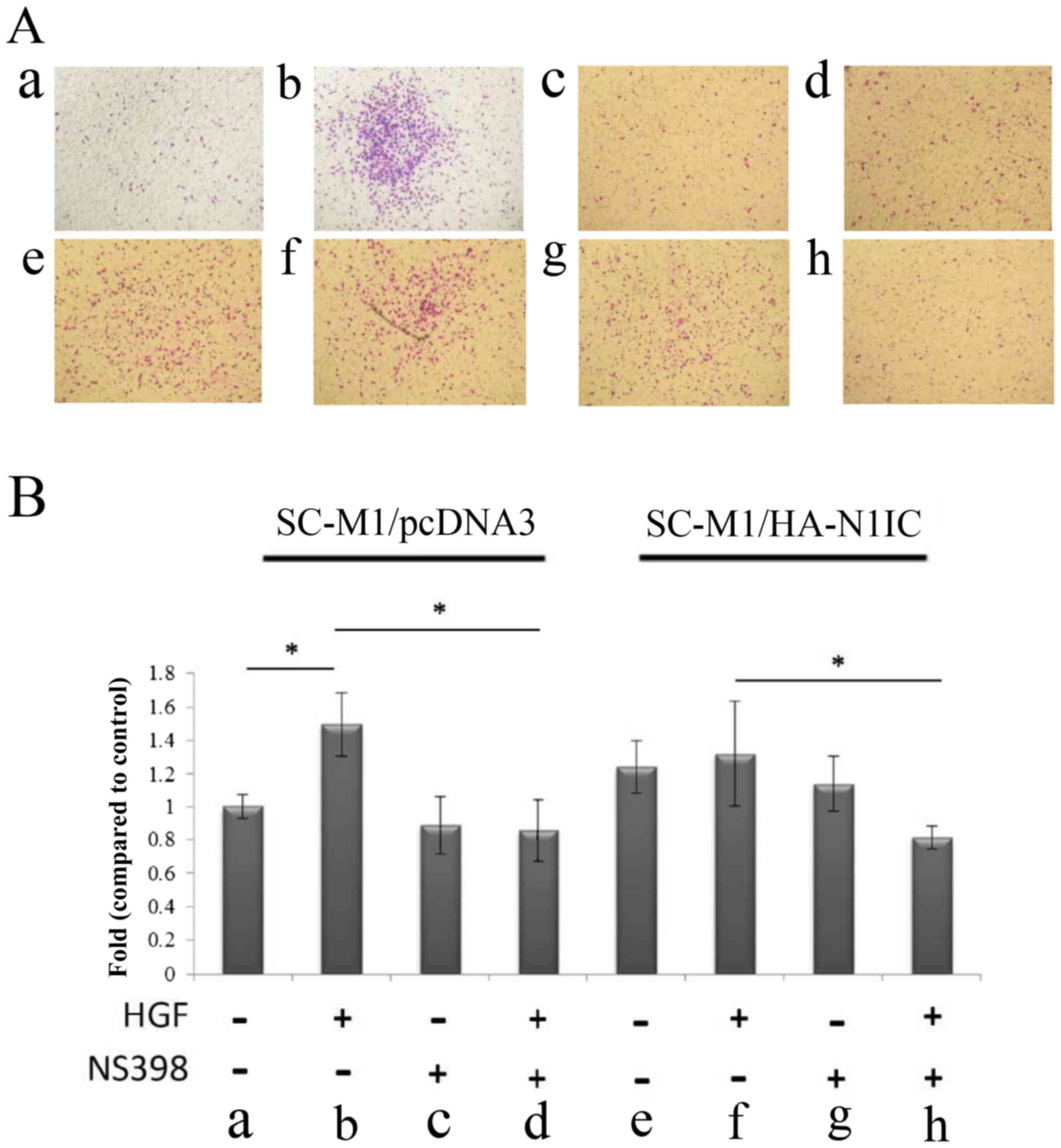 | Figure 4.Migration analysis in SC-M1/HA-N1IC
and SC-M1/pcDNA3 cells. (A) The migration of transfected SC-M1
cells was evaluated in the presence or absence of HGF (25 ng/ml),
and/or NS398 (50 µM) in SC-M1/pcDNA and SC-M1/HA-N1IC cells for 24
h. The migration ability of (a) SC-M1/pcDNA3 (b) SC-M1/pcDNA + HGF
(c) SC-M1/pcDNA3 + NS398 (d) SC-M1/pcDNA3 + HGF and NS398. (e)
SC-M1/HA-N1IC (f) SC-M1/HA-N1IC + HGF (g) SC-M1/HA-N1IC + NS-398
(h) SC-M1/HA-N1IC + HGF and NS398. (B) Migration of transfected
SC-M1 cells was evaluated following treatment with HGF (0 and 25
ng/ml), with/without NS-398 (50 µM) in SC-M1/pcDNA3 and
SC-M1/HA-N1IC cells for 24 h. Migration ability of (a) SC-M1/pcDNA3
cells, (b) SC-M1/pcDNA3 + HGF, (c) SC-M1/pcDNA3 + NS398, (d)
SC-M1/pcDNA3 + HGF and NS398; (e) SC-M1/HA-N1IC cells, (f)
SC-M1/HA-N1IC + HGF, (g) SC-M1/HA-N1IC + NS398, (h) SC-M1/HA-N1IC +
HGF and NS398. Fold changes in migration of gastric cancer cells
from respective groups. Cells were cultured in serum-free media for
24 h before the Transwell assay. SC-M1/pcDNA3 or SC-M1/HA-N1IC
cells were treated with/without HGF in the presence/absence of
COX-2 inhibitor NS398 for 24 h, and then the migrated cells were
counted. Total cell number were counted from nine fields.
*P<0.01 compared to SC-M1/pcDNAs. (Data are presented as the
mean ± SD of 2 replicates from 3 experiments). N1IC, Notch1
intracellular domain; HGF, hepatocyte growth factor. |
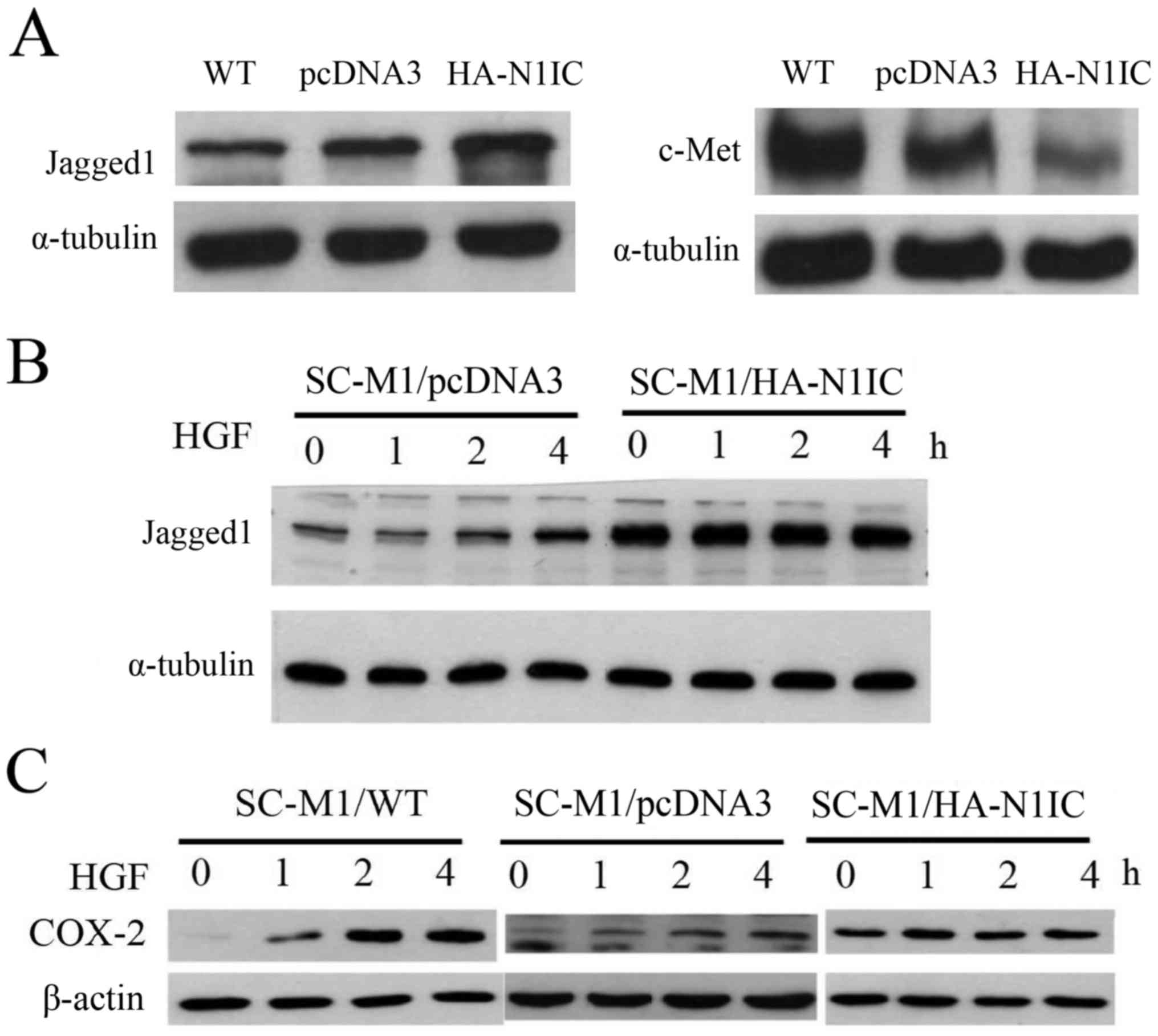
Effect of HGF treatment on the
expression of Jagged1 and COX-2 proteins in Notch1-overexpressing,
control and wild-type (WT) SC-M1 cells
First, we compared the expression of c-Met and,
Jagged1 by western blot analysis in Notch1-overexpressing, control
and WT SC-M1 gastric cancer cells. Jagged1 expression in SC-M1/WT
and SC-M1/pcDNA3 cells was lower than that in SC-M1/HA-N1IC cells,
but c-Met protein expression was lower in SC-M1/pcDNA3 and
SC-M1/HA-N1IC cells than that in the SC-M1/WT cells. The expression
of c-Met protein in the SC-M1/pcDNA3 cells was higher than that in
the SC-M1/HA-N1IC cells. Reduced expression of c-Met protein was
observed in Notch1-overexpressing SC-M1 cells (Fig. 5A). Second, we treated gastric cancer
cells with HGF and examined the effects over time. After HGF (25
ng/ml) treatment for 1, 2 or 4 h, cell lysates were collected for
western blot analysis. HGF stimulated the expression of Jagged1 and
COX-2 proteins in the SC-M1/pcDNA3 cells in a time-dependent
manner. However, Jagged1 and COX-2 proteins levels were not
obviously increased in the SC-M1/HA-N1IC cells. A time-dependent
increase of Jagged1 was observed in the HGF-stimulated SC-M1 cells
but not in the Notch1-overexpressing SC-M1 cells (Fig. 5B). COX-2 expression was elevated in
the SC-M1/WT and SC-M1/pcDNA3 cells after HGF treatment. However,
no significant change in COX-2 expression was observed in the
SC-M1/HA-N1IC cells (Fig. 5C).
Taken together, these results imply that a feedback loop exists
between the HGF/c-Met and Notch1/Jagged1 signaling pathways.
Discussion
Our results showed that patients with positive c-Met
expression had poorer overall survival than patients without
positive expression. Jagged1 expression was correlated with
HGF/c-Met expression, and HGF increased the expression of Jagged1,
which in turn enhanced the Notch1 signaling pathway, resulting in
the expression of COX-2 and increased proliferation and migration
ability in gastric cancer cells. However, the overexpression of
N1IC suppressed the expression of c-Met and the sensitivity of HGF.
The HGF/c-Met and Jagged1/Notch1 signaling pathways have the same
target that triggers COX-2 mRNA expression to further manufacture
the key product of COX-2 proteins.
Our previous studies have supported the concept of
the parallel HGF/cMET/COX-2 (2) and
Notch1/Jagged1/COX-2 (3) pathways.
Both the gain and loss of c-MET and Notch were found to regulate
COX-2 expression, and the overexpression of N1IC or Jagged1
extracellular domain in SC-M1 cells had the same effect (3,9). HGF
induced Jagged1 expression, which increased in a time-dependent
manner. Therefore, HGF could increase Jagged1 ligand activity
through the HGF/c-Met signaling pathway. Our results support
findings of a previous study by Stella et al, who reported
that constitutive activation of Notch can inhibit HGF-dependent Ras
activation in a model using Drosophila (15). Furthermore, activation of c-Met
stimulates Notch signaling by inducing Notch ligand. Hence, an
alternative loop exists in which HGF/c-Met induces the activation
of Notch signaling through Jagged1 ligand, whereas Notch
overexpression represses the expression of c-Met.
HGF plays an important role in the regulation of
growth and metastasis of tumor cells. Our previous study showed
that gastric cancer patients with high serum HGF had poorer
prognosis than those with low serum HGF (16,17).
In addition, HGF was found to bind to the c-Met receptor and
activates the tyrosine kinase signaling pathway, resulting in cell
invasion and metastasis. COX-2 inhibitor NS398 was found to repress
the proliferation and migration ability in human gastric cancer
SC-M1 cells and inhibit the expression of COX-2 protein, which is
stimulated by HGF (18). Uen et
al (19) reported that patients
with elevated c-Met mRNA expression in peripheral blood had poorer
prognosis than patients with negative c-Met expression.
Overexpression of c-Met increased the sensitization of gastric
cancer cells to HGF, which in turn resulted in cell invasion and
metastasis (20). In addition,
Yamamoto et al (21)
reported that COX-2 protein expression was significantly elevated
in human gastric cancer and associated with lymphatic invasion and
metastasis. Thus, it is conceivable that HGF/c-Met has a
transcriptional effect on the COX-2 promotor to induce the end
product COX-2 protein to modulate the behavior of gastric cancer
cells.
The Jagged1/Notch1 signaling pathway also plays an
important functional role in regulating tumor cell proliferation
and migration. Previous studies have revealed that Notch ligand
Jagged1 and c-Met expression both positively correlate with COX-2
expression (23). We found a
positive correlation between c-Met and Jagged1 in human gastric
cancer tissues. In addition to their regulation of COX-2 protein,
there is a circuit loop through which HGF increases Jagged1
expression, which in turn activates Notch1 activity. Therefore,
elucidating the mechanism involved in the downstream regulation of
c-Met and the interplay of Notch and c-Met signaling could help to
understand the transcription effect in gastric cancer.
HGF regulates cellular signaling pathways through
its interaction with c-Met. HGF was shown to elicit prolonged
phosphorylation of growth factor receptor-bound protein 2
(GRB2)-associated-binding protein 1 (GAB1) and to lead to prolonged
activation of mitogen-activated protein kinases (MAPK) (22,23).
Notch signaling, triggered by the MAPK pathway, was reported to
play an important role in tumor angiogenesis (24,25).
Jagged1 expression activates Notch signaling in head and neck
squamous cell carcinoma and promotes endothelial capillary-like
sprout formation (24). HGF was
found to induce hairy and enhancer of split-1 (HES-1) mRNA
activation, resulting in the activation of Notch (21,26).
Moreover, the activation of c-Met was previously shown to stimulate
Notch function in Drosophila (15). We found that Jagged1/Notch1
signaling could be triggered by HGF/c-Met signaling. Taken
together, these findings suggest that, through MAPK and Hes-1
signal transduction, Jagged1/Notch1 signaling functions downstream
of c-Met.
The identification of patients with specific genetic
mutations or amplifications has been applied in clinical target
therapy for lung and breast cancer, and gastrointestinal stromal
tumor. The Cancer Genome Atlas (TCGA) project divided gastric
cancer into four molecular subtypes: Epstein-Barr virus
(EBV)-positive, microsatellite instability (MSI), genomically
stable (GS), and chomosomal instability (CIN) (27). Targeted therapy toward human
epidermal growth factor receptor 2 (Her-2 receptor) is applied to
specific advanced gastric cancer patients with positive expression
of Her-2/Neu (28). Recent studies
have described carcinogenesis and the development of targeted
therapy for c-Met signaling in gastric cancer (6,29).
Nickoloff et al also reported the biopharmacological
potential of the Notch receptor as a targeted therapy for cancer
(30). Notch ligand Jagged1 is also
a potential pharmacogenomic target for cancer therapy (31). Inhibitory antibodies for c-Met and
Notch receptors or inhibitors for Notch ligand Jagged1 may provide
a therapeutic strategy for cancer on the basis of this signaling
pathway. However, the interaction between HGF/c-Met and
Jagged1/Notch1 signaling pathways should be considered. Both
HGF/Notch1 and Jagged1/Notch1 signaling pathways target COX-2 mRNA,
resulting in the production of COX-2 protein and the secretion of
prostaglandin E2 (PGE2), and the elevation of
the serum HGF level could induce the expression of Jagged1.
Therefore, it is insufficient for the treatment of gastric cancer
to involve the use of a single agent for a unique target receptor.
Multi-modality targeted therapies should be considered for the
therapeutic strategy for gastric cancer.
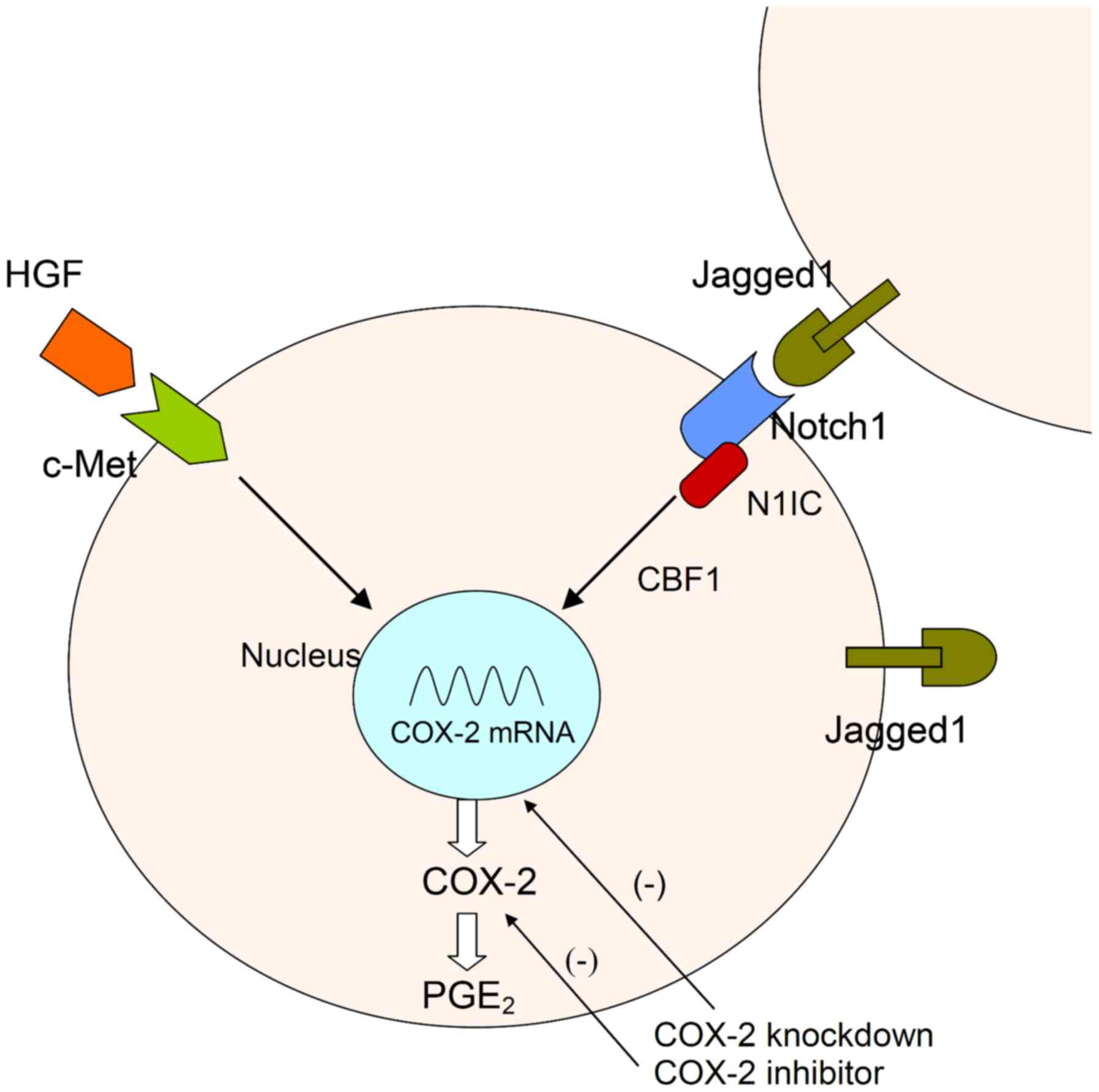
In conclusion, Jagged1 expression is positively
correlated with c-Met expression in human gastric cancer specimens.
Constitutive activation of Notch1 limits HGF activity by repressing
the c-Met oncogene. HGF/c-Met plays a minor role in the
constitutive Notch1 signaling pathway. HGF/c-Met signaling induced
an increase in Jagged1 expression in the control group in a
time-dependent manner. Therefore, the reciprocal loop regulation
between the HGF/c-Met and Jagged 1/Notch signaling pathways plays
an important role in modulating a biological response in gastric
cancer cells (Fig. 6).
Acknowledgements
We thank Dr Shin-Shian Hsu for the suggestions
regarding our manuscript.
Funding
The present study was supported by the Division of
Experimental Surgery of the Department of Surgery in Taipei
Veterans General Hospital, grants from Taipei Veterans General
Hospital (V101B-035, V107C-043) and grants from the Ministry of
Science and Technology (NSC 101-2314-B-075-074, MOST
104-2314-B-075-032-, 105-2314-B-075-005-MY2). No sources of funding
played a role in the study design, data collection, analysis and
interpretation of data, writing of the manuscript, and in the
decision to submit the manuscript for publication.
Availability of data and materials
The datasets used during the present study are
available from the corresponding author upon reasonable
request.
Authors' contributions
CWC, MHY and YMS conceived and designed the study.
CWW and WLF collected surgical specimens. KHH, ICS and AFYL
performed the experiments. KHH wrote the study. MHY, CWC, TSY, HCL
and PHY reviewed and edited the manuscript. All authors read and
approved the manuscript and agree to be accountable for all aspects
of the research in ensuring that the accuracy or integrity of any
part of the work are appropriately investigated and resolved.
Ethics approval and consent to
participate
This study was approved by the Institutional Review
Board of Taipei Veterans General Hospital and informed consent was
provided by all subjects.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Ferlay J, Soerjomataram I, Dikshit R, Eser
S, Mathers C, Rebelo M, Parkin DM, Forman D and Bray F: Cancer
incidence and mortality worldwide: Sources, methods and major
patterns in GLOBOCAN 2012. Int J Cancer. 136:E359–E386. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chen JH, Wu CW, Kao HL, Chang HM, Li AFY,
Liu TY and Chi CW: Effects of COX-2 inhibitor on growth of human
gastric cancer cells and its relation to hepatocyte growth factor.
Cancer Lett. 239:263–270. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Yeh TS, Wu CW, Hsu KW, Liao WJ, Yang MC,
Li AFY, Wang AM, Kuo ML and Chi CW: The activated Notch1 signal
pathway is associated with gastric cancer progression through
cyclooxygenase-2. Cancer Res. 69:5039–5048. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Jones MK, Sasaki E, Halter F, Pai R,
Nakamura T, Arakawa T, Kuroki T and Tarnawski AS: HGF triggers
activation of the COX-2 gene in rat gastric epithelial cells:
Action mediated through the ERK2 signaling pathway. FASEB J.
13:2186–2194. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Amemiya H, Kono K, Itakura J, Tang RF,
Takahashi A, An FQ, Kamei S, Iizuka H, Fujii H and Matsumoto Y:
c-Met expression in gastric cancer with liver metastasis. Oncology.
63:286–296. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Christensen JG, Burrows J and Salgia R:
c-Met as a target for human cancer and characterization of
inhibitors for therapeutic intervention. Cancer Lett. 225:1–26.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Chappell WH, Green TD, Spengeman JD,
McCubrey JA, Akula SM and Bertrand FE: Increased protein expression
of the PTEN tumor suppressor in the presence of constitutively
active Notch-1. Cell Cycle. 4:1389–1395. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lindsell CE, Shawber CJ, Boulter J and
Weinmaster G: Jagged: A mammalian ligand that activates Notch1.
Cell. 80:909–917. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hsu KW, Hsieh RH, Huang KH, Li Fen-Yau A,
Chi CW, Wang TY, Tseng MJ, Wu KJ and Yeh TS: Activation of the
Notch1/STAT3/Twist signaling axis promotes gastric cancer
progression. Carcinogenesis. 33:1459–1467. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ohno R, Yoshinaga K, Fujita T, Hasegawa K,
Iseki H, Tsunozaki H, Ichikawa W, Nihei Z and Sugihara K: Depth of
invasion parallels increased cyclooxygenase-2 levels in patients
with gastric carcinoma. Cancer. 91:1876–1881. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Shi H, Xu JM, Hu NZ and Xie HJ: Prognostic
significance of expression of cyclooxygenase-2 and vascular
endothelial growth factor in human gastric carcinoma. World J
Gastroenterol. 9:1421–1426. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lim HY, Joo HJ, Choi JH, Yi JW, Yang MS,
Cho DY, Kim HS, Nam DK, Lee KB and Kim HC: Increased expression of
cyclooxygenase-2 protein in human gastric carcinoma. Clin Cancer
Res. 6:519–525. 2000.PubMed/NCBI
|
|
13
|
Huang TT, Ping YH, Wang AM, Ke CC, Fang
WL, Huang KH, Lee HC, Chi CW and Yeh TS: The reciprocal regulation
loop of Notch2 pathway and miR-23b in controlling gastric
carcinogenesis. Oncotarget. 6:18012–18026. 2015.PubMed/NCBI
|
|
14
|
Hsu KW, Fang WL, Huang KH, Huang TT, Lee
HC, Hsieh RH, Chi CW and Yeh TS: Notch1 pathway-mediated
microRNA-151-5p promotes gastric cancer progression. Oncotarget.
7:38036–38051. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Stella MC, Trusolino L, Pennacchietti S
and Comoglio PM: Negative feedback regulation of Met-dependent
invasive growth by Notch. Mol Cell Biol. 25:3982–3996. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wu CW, Chi CW, Su TL, Liu TY, Lui WY and
P'eng FK: Serum hepatocyte growth factor level associate with
gastric cancer progression. Anticancer Res. 18:3657–3659.
1998.PubMed/NCBI
|
|
17
|
Wu CW, Li AFY, Chi CW, Chung WW, Liu TY,
Lui WY and P'eng FK: Hepatocyte growth factor and Met/HGF receptors
in patients with gastric adenocarcinoma. Oncol Rep. 5:817–822.
1998.PubMed/NCBI
|
|
18
|
Lin JS, Lu CW, Huang CJ, Wu PF, Robinson
D, Kung HJ, Chi CW, Wu CW, Yang WK, Whang-Peng JJ, et al:
Protein-tyrosine kinase and protein-serine/threonine kinase
expression in human gastric cancer cell lines. J Biomed Sci.
5:101–110. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Uen YH, Lin SR, Wu CH, Hsieh JS, Lu CY, Yu
FJ, Huang TJ and Wang JY: Clinical significance of MUC1 and c-Met
RT-PCR detection of circulating tumor cells in patients with
gastric carcinoma. Clin Chim Acta. 367:55–61. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Boccaccio C and Comoglio PM: Invasive
growth: A MET-driven genetic programme for cancer and stem cells.
Nat Rev Cancer. 6:637–645. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yamamoto H, Itoh F, Fukushima H, Hinoda Y
and Imai K: Overexpression of cyclooxygenase-2 protein is less
frequent in gastric cancers with microsatellite instability. Int J
Cancer. 84:400–403. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Gual P, Giordano S, Williams TA, Rocchi S,
Van Obberghen E and Comoglio PM: Sustained recruitment of
phospholipase C-γ to Gab1 is required for HGF-induced branching
tubulogenesis. Oncogene. 19:1509–1518. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Maroun CR, Naujokas MA, Holgado-Madruga M,
Wong AJ and Park M: The tyrosine phosphatase SHP-2 is required for
sustained activation of extracellular signal-regulated kinase and
epithelial morphogenesis downstream from the met receptor tyrosine
kinase. Mol Cell Biol. 20:8513–8525. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zeng Q, Li S, Chepeha DB, Giordano TJ, Li
J, Zhang H, Polverini PJ, Nor J, Kitajewski J and Wang CY:
Crosstalk between tumor and endothelial cells promotes tumor
angiogenesis by MAPK activation of Notch signaling. Cancer Cell.
8:13–23. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
King AM, Van der Put E, Blomberg BB and
Riley RL: Accelerated Notch-dependent degradation of E47 proteins
in aged B cell precursors is associated with increased ERK MAPK
activation. J Immunol. 178:3521–3529. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Gude NA, Emmanuel G, Wu W, Cottage CT,
Fischer K, Quijada P, Muraski JA, Alvarez R, Rubio M, Schaefer E,
et al: Activation of Notch-mediated protective signaling in the
myocardium. Circ Res. 102:1025–1035. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
The Cancer Genome Atlas Research Network,
. Bass AJ, Thorsson V, Shmulevich I, Reynolds SM, Miller M, Bernard
B, Hinoue T, Laird PW, Curtis C, Shen H, et al: Comprehensive
molecular characterization of gastric adenocarcinoma. Nature.
513:202–209. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Gravalos C and Jimeno A: HER2 in gastric
cancer: A new prognostic factor and a novel therapeutic target. Ann
Oncol. 19:1523–1529. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Peruzzi B and Bottaro DP: Targeting the
c-Met signaling pathway in cancer. Clin Cancer Res. 12:3657–3660.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Nickoloff BJ, Osborne BA and Miele L:
Notch signaling as a therapeutic target in cancer: A new approach
to the development of cell fate modifying agents. Oncogene.
22:6598–6608. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Katoh M and Katoh M: Notch ligand, JAG1,
is evolutionarily conserved target of canonical WNT signaling
pathway in progenitor cells. Int J Mol Med. 17:681–685.
2006.PubMed/NCBI
|