Introduction
Prostate cancer is among the most common types of
cancer in elderly men, and is a leading cause of cancer-associated
mortality in Western countries (1).
Treatment strategies for prostate cancer include surgery, hormone
therapy, radiotherapy and chemotherapy (2). Chemotherapy serves an important role
in the treatment of castration-resistant metastatic prostate cancer
(3). Paclitaxel has been
demonstrated to result in significant antitumor responses in
combination with other agents (4,5).
However, there are limitations to chemotherapy, including the
development of drug resistance, particularly multidrug resistance
(6). Therefore, the identification
of effective mechanisms to prevent chemotherapy resistance is
essential.
The mechanism by which paclitaxel acts on prostate
cancer cells is different from the mechanisms of action of other
anticancer drugs, and it functions by inhibition of cytoplasmic
microtubule depolymerization (7).
This inhibits normal spindle formation and causes cell cycle arrest
in the G2/M phase (8). Other
studies have suggested that the main mechanism by which paclitaxel
acts on prostate cancer cells is via Bcl-2 degradation, which
reduces the DNA-protective effect of Bcl-2. This results in DNA
cleavage into fragments by nucleic acid enzymes, and ultimately
leads to apoptosis (9,10).
Zinc is essential for maintaining the integrity of
various enzymes and transcription factors (11). Zinc also serves an important role in
gene stability and expression (12). In patients with prostate cancer,
zinc concentrations have been demonstrated to be reduced by 60–70%
in blood serum and cancerous prostate tissue, and to decrease
further as cancer progresses (13,14).
Another study reported that zinc ions are involved in tumor
occurrence and progression (15),
and a decline in the concentration of zinc in the serum and tissues
has been observed in patients with various types of malignant
tumor, including liver, gallbladder, gastrointestinal tract, blood
and prostate cancers (16,17).
Under androgen stimulation, the peripheral zone of
the prostate exhibits high zinc concentrations (18). In prostate cancer, epithelial cells
lose the ability to absorb high concentrations of zinc, and tumor
growth increases zinc consumption, thus resulting in decreased zinc
content (19,20). Previous studies have suggested that
physiological concentrations of zinc can directly induce prostate
cancer cell apoptosis via the mitochondrial pathway, release of
mitochondrial cytochrome c and activation of the
apoptosis-associated protease caspase-3, which triggers programmed
cell death (9,21). Previously, researchers have injected
zinc into prostate tumors and found that it inhibited cell growth
and prolonged patient survival. Furthermore, treatment with
paclitaxel alone has been demonstrated to be associated with the
development of drug resistance (22,23).
We hypothesized that reduced zinc concentrations may be associated
with chemotherapy resistance in prostate cancer. In the present
study, it was assessed whether zinc increased chemosensitivity of
PC-3 cells to paclitaxel.
Materials and methods
Cell cultivation
PC-3 cells were obtained from the Prostate Diseases
Prevention and Treatment Research Center (Jilin University,
Changchun, China) and were cultured in Iscove's modified Dulbecco's
medium (IMDM; HyClone; GE Healthcare Life Sciences, Little
Chalfont, UK) supplemented with 10% fetal bovine serum (HyClone; GE
Healthcare Life Sciences) and penicillin-streptomycin (1,000
µg/ml). After 48–72 h, the cells were passaged by digestion with
0.25% EDTA-free trypsin (Sangon Biotech Co., Ltd., Shanghai, China)
and re-seeded at one third of the previous density. When the cells
reached 70–75% confluency, 0, 50, 100, 150, 200, 250, 300, 350, 400
or 450 µmol/l zinc (Sigma-Adrich; Merck KGaA, Darmstadt, Germany)
and 0, 2.5, 5, 10, 20, 40, 60 or 80 nmol/l paclitaxel (Harbin
Pharmaceutical Group Holding Co., Ltd., Harbin, China) were added.
Cells were then collected for analysis of cell activity, apoptosis,
mitochondrial membrane potential and protein and gene expression
levels.
MTT assay
PC-3 cells were seeded in a 96-well plate at
approximately 8×103 cells/well and incubated at 37°C in
5% CO2. After 24 h, when the cells had reached 70–75%
confluency, 0, 50, 100, 150, 200, 250, 300, 350, 400 or 450 µmol/l
zinc chloride and 0, 2.5, 5, 10, 20, 40, 60 or 80 nmol/l paclitaxel
were added. The cells were incubated for a further 48 h, then 20 µl
of MTT (Sangon Biotech Co., Ltd., Shanghai, China) was added per
well. After 4 h, 150 µl dimethyl sulfoxide (Sangon Biotech Co.,
Ltd., Shanghai, China) was added to each well to dissolve the
formazan crystals, and the absorbance was measured at a wavelength
of 490 nm (A490) using a microplate reader (Sangon Biotech Co.,
Ltd., Shanghai, China).
Cell clone formation assay
A total of 4 groups of cells were prepared: Control
group, zinc-treated group, paclitaxel-treated group and zinc +
paclitaxel-treated group. Cells were seeded in 6-well plates (500
cells/well; 3-wells/group). The culture medium was replaced every
48 h. Colony formation was observable after 14 days of continuous
culture. The culture medium was then removed, and the cells were
washed twice with PBS and fixed in 4% formaldehyde for 30 min at
room temperature. The fixed cells were subjected to Giemsa staining
(Rapid Giemsa staining kit; Sangon Biotech Co., Ltd., Shanghai,
China) for 20 min at room temperature, and cell colonies with a
diameter >1 mm were counted.
Immunofluorescence
PC-3 cells were seeded in 24-well plates at
5×104 cells/well onto a glass coverslip. Following cell
adherence, they were exposed to different treatment conditions.
After 4 h, the cells were fixed with 4% paraformaldehyde at room
temperature for 10–15 min, then washed 2–3 times with 0.1 M PBS.
The cells were then incubated with 0.1% Triton for 6–10 min, and
washed thrice for 1 min with 0.01 M PBS. The cells were incubated
with 5% non-immune bovine serum albumin (HyClone; GE Healthcare
Life Sciences, Little Chalfont, UK) to prevent nonspecific binding,
then with a Ki-67 primary antibody (dilution, 1:1,000; cat. no.
27309-1-AP; ProteinTech Group, Inc. Chicago, IL, USA) at 37°C for
30 min. Following incubation with a goat anti-rabbit horseradish
peroxidase-conjugated IgG secondary antibody (dilution, 1:1,000;
cat. no., SA00001-2; ProteinTech Group, Inc. Chicago, IL, USA) at
room temperature for 40 min, the cells were stained with Hoechst
33342 (1 µg/ml; Sangon Biotech Co., Ltd., Shanghai, China) for 2
min at room temperature, then mounted using aqueous mounting media
(glycerin and water, 9:1). Cell staining was detected by
fluorescence microscopy at ×1,200, magnification.
Flow cytometric analysis of cell
apoptosis
Following exposure of PC-3 cells to different
treatment conditions (control group; 250 µmol/l zinc group; 10
nmol/l; paclitaxel group, and 250 µmol/l zinc + 10 nmol/l
paclitaxel group) for 48 h, they were washed twice with PBS, then
collected using EDTA-free trypsin. The cells were collected by
centrifugation at 1,000 × g for 5 min at 4°C. Annexin V-FITC (5 µl;
Beckman Coulter Inc., Brea, CA, USA) was added and the cells were
incubated in the dark for 15 min at 4°C, then 10 µl of propidium
iodide (PI; Beckman Coulter Inc.) was added and the cells were
incubated in the dark for a further 5 min at 4°C. An Epics-XL-MCL
flow cytometer (Beckman Coulter, Inc., Brea, CA, USA) was used to
detect the rate of cell apoptosis within 1 h of staining.
Flow cytometric analysis of
mitochondrial membrane potential
Around 2.7×105 cells/well were cultured
in 6-well plates. The cells were collected for flow cytometric
analysis by washing twice with PBS, followed by treatment with
0.25% EDTA-free trypsin and centrifugation at 900 × g for 5 min at
4°C. Serum-free IMDM (1 ml) and JC-1 (1 µl; Beckman Coulter Inc.)
were added and the cells were incubated at 37°C for 20 min, then
centrifuged at 1,500–2,000 × g for 5 min. The culture medium was
discarded and the cells were washed twice with PBS, and centrifuged
at 2,000 × g for 5 min. Finally, the cells were resuspended in 400
µl IMDM. Mitochondrial membrane potential was analyzed using an
Epics-XL-MCL flow cytometer.
Reverse transcription PCR
(RT-PCR)
mRNA was extracted using TRIzol (Invitrogen; Thermo
Fisher Scientific, Inc., Waltham, MA, USA), according to the
manufacturer's protocol. PC-3 cells were cultured in a 6-well plate
and harvested by washing twice with PBS, 500 µl TRIzol was added
per well and incubated at room temperature for 5 min. The cells
were transferred to RNase-free Eppendorf tubes and trichloromethane
was added prior to centrifugation. Isopropanol (cat. no., A507048;
Sangon Biotech Co., Ltd.) was added to the aqueous phase at room
temperature for 5 min. Finally, RNA was washed in 75% ethanol and
resuspended in RNase-free water. Reverse transcription was
conducted using Promega M-MLV reverse transcriptase (Thermo Fisher
Scientific, Inc., Waltham, MA, USA), according to the
manufacturer's instructions. PCR was conducted using 2X EasyTaq PCR
SuperMix (cat. no. AS111-02; Beijing Transgen Biotech Co., Ltd.,
Beijing, China). The primer sequences and thermocycling conditions
are presented in Tables I and
II. The images were captured using
the Tanon, 1600R image processing system (Tanon Science and
Technology Co., Ltd., Shanghai, China). The gray ratio between the
target genes and GAPDH was analyzed to measure the expression level
of target genes by image processing system (Tanon, 1600R; Tanon
Science and Technology Co., Ltd.).
 | Table I.Primer sequences used reverse
transcription-polymerase chain reaction analysis. |
Table I.
Primer sequences used reverse
transcription-polymerase chain reaction analysis.
| Gene | Forward primer
sequence | Reverse primer
sequence |
|---|
| GAPDH |
5′-AGAAGGCTGGGGCTCATTTG-3′ |
5′-AGGGGCCATCCACAGTCTTC-3′ |
| Bcl-2 |
5′-GACTTCGCCGAGATGTCCAGC-3′ |
5′-GCGTTACGATCGCCTCCATCA-3′ |
| Bax |
5′-CGGCGAATTGGAGATGAACTG-3′ |
5′-AGCAAAGTAGAAGAGGGCAACC-3′ |
| Caspase-9 |
5′-GGCCCTTCCTCGCTTCATCTC-3′ |
5′-GGTCCTTGGGCCTTCCTGGTAT-3′ |
| Caspase-3 |
5′-ATGGACAACAACGAAACCTCCGTG-3′ |
5′-CCACTCCCAGTCATTCCTTTTAGTG-3′ |
| Table II.Polymerase chain reaction
thermocycling conditions. |
Table II.
Polymerase chain reaction
thermocycling conditions.
| Gene | Denaturation | Annealing | Extension | Number of
cycles |
|---|
| GAPDH | 94°C, 30 sec | 55°C, 30 sec | 72°C, 30 sec | 28 |
| Bcl-2 | 94°C, 30 sec | 55°C, 30 sec | 72°C, 30 sec | 30 |
| Bax | 94°C, 30 sec | 54°C, 30 sec | 72°C, 30 sec | 30 |
| Caspase-9 | 94°C, 30 sec | 55°C, 30 sec | 72°C, 30 sec | 28 |
| Caspase-3 | 94°C, 30 sec | 56°C, 30 sec | 72°C, 30 sec | 28 |
Western blotting
PC-3 cells were lysed by incubation with 100–150 µl
of radioimmunoprecipitation assay buffer supplemented with
phenylmethylsulfonyl fluoride (Roche Diagnostics, Basel,
Switzerland), then sonicated 3–5 times for 3–4 sec using ultrasound
pyrolysis apparatus (Tomy Seiko Co., Ltd., Tokyo, Japan).
Subsequently, the cells were centrifuged at 12,000 × g, for 20 min
at 4°C and the protein concentration was measured using Bradford
reagent (BioRad Laboratories, CA, USA). The lysates were subjected
to 12% SDS-PAGE and transferred onto polyvinylidene fluoride
membranes (EMB Millipore, Billerica, MA, USA). The membraned was
wished with TSB-Tween (Sangon Biotech Co., Ltd.) on the horizontal
rocking bed 3 times for 5 min, and then blocked with 5% skimmed
milk (Sangon Biotech Co., Ltd.) on the horizontal rocking bed for 1
h at room temperature. Bcl-2-associated X, apoptosis regulator
(Bax), Bcl-2, caspase-3, caspase-9, cleaved caspase-3 and cleaved
caspase-9 were detected by western blotting. Bax (dilution,
1:1,000; cat. no., 23931-1-AP) and Bcl-2 (dilution, 1:1,000; cat.
no. 12789-1-AP) primary antibodies were purchased from ProteinTech
Group, Inc. (Chicago, IL, USA). Cleaved caspase-3 (dilution,
1:1,000; cat. no., 9664), cleaved caspase-9 (dilution, 1:1,000;
cat. no., 9509), caspase-3 (dilution, 1:1,000; cat. no., 9662),
caspase-9 (dilution, 1:1,000; cat. no., 9504) and actin (dilution,
1:1,000; cat. no., 8456) primary antibodies were purchased from
Cell Signaling Technology Inc. (Danvers, MA, USA). All the primary
antibodies were incubated at room temperature for 4 h prior to
washing with TBS-Tween. Next, membranes were incubated with a goat
anti-rabbit horseradish peroxidase-conjugated IgG secondary
antibody (dilution, 1:1,000; cat. no., SA00001-2; ProteinTech
Group, Inc.) at room temperature for 1 h. Proteins were detected
using an enhanced chemiluminescence kit (cat. no. 120702-74;
Advansta, Inc., Menio Park, CA, USA). The Syngene Bio Imaging
system (GeneGnome HR, Synoptics Ltd., Cambridge, UK) was used to
capture the images. And the Image-Pro Plus (Media Cybernetics,
Inc., Rockville, MD, USA) was used to quantify the protein
expression.
Statistical analysis
All data are presented as the mean ± standard
deviation. SPSS 19.0 (IBM Corp., Armonk, NY, USA) was used to
perform one-way analysis of variance and the least significant
difference test for multiple comparisons. P<0.05 was considered
to indicate a statistically significant difference.
Results
Zinc increases the sensitivity of PC-3
cells to paclitaxel
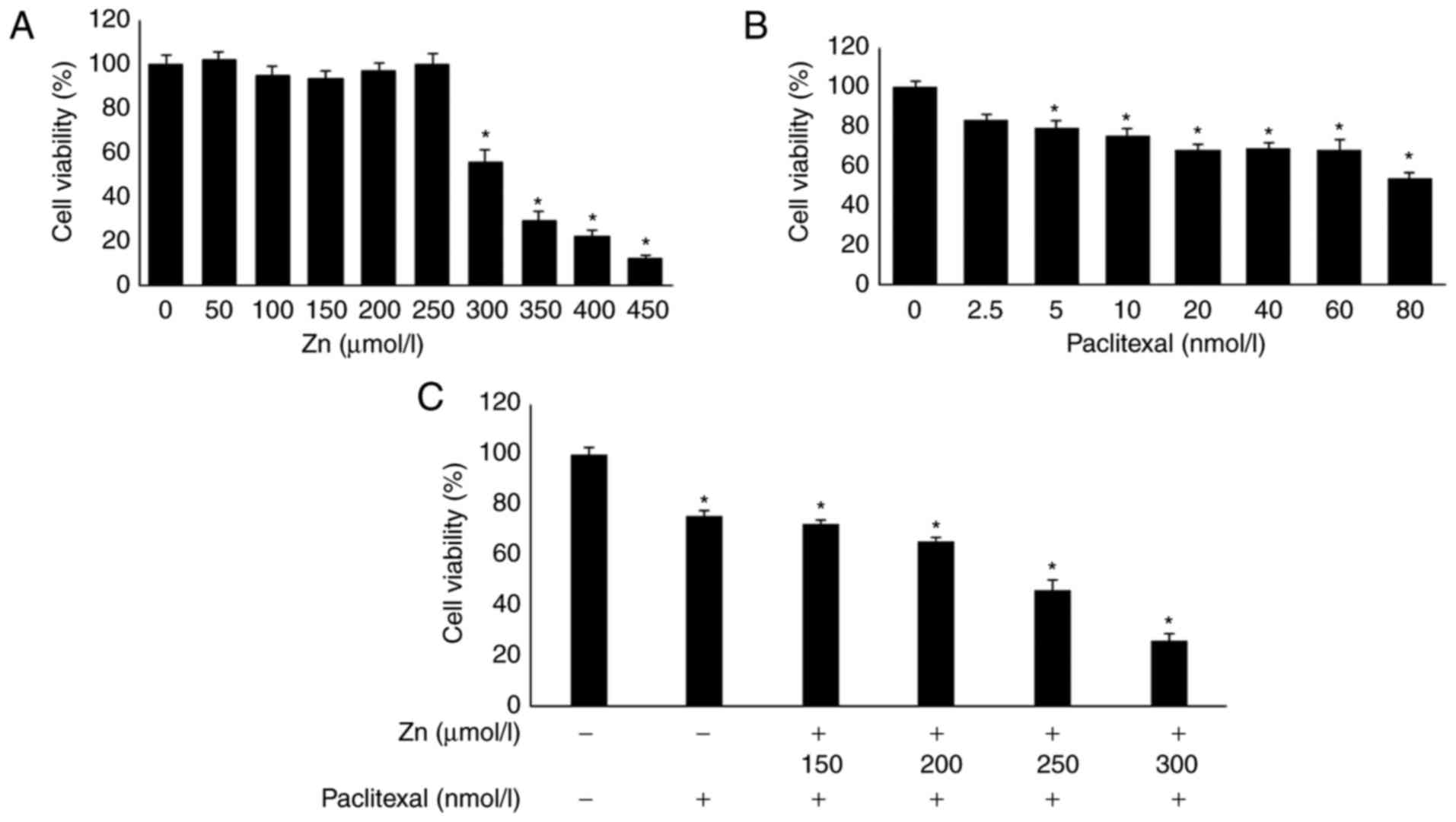
First, an MTT assay was used to determine the
optimal concentration combination of zinc and paclitaxel. PC-3
cells were treated with different concentrations of zinc and
paclitaxel for 48 h. The results showed that 0–250 µmol/l zinc had
no significant effect on PC-3 cells compared with untreated cells
(Fig. 1A). However, zinc
concentrations >250 µmol/l significantly decreased the activity
of PC-3 cells. PC-3-cell viability was reduced by ~25% by 10 nmol/l
paclitaxel compared with untreated cells (Fig. 1B). The effect of a 48-h incubation
with 10 nmol/l paclitaxel and different concentrations of zinc was
investigated. Treatment with 10 nmol/l paclitaxel combined with 250
µmol/l zinc reduced PC-3 cell proliferation by ~50% compared with
untreated cells (P<0.05; Fig.
1C). Therefore, 10 nmol/l paclitaxel combined with 250 µmol/l
zinc was used in subsequent experiments.
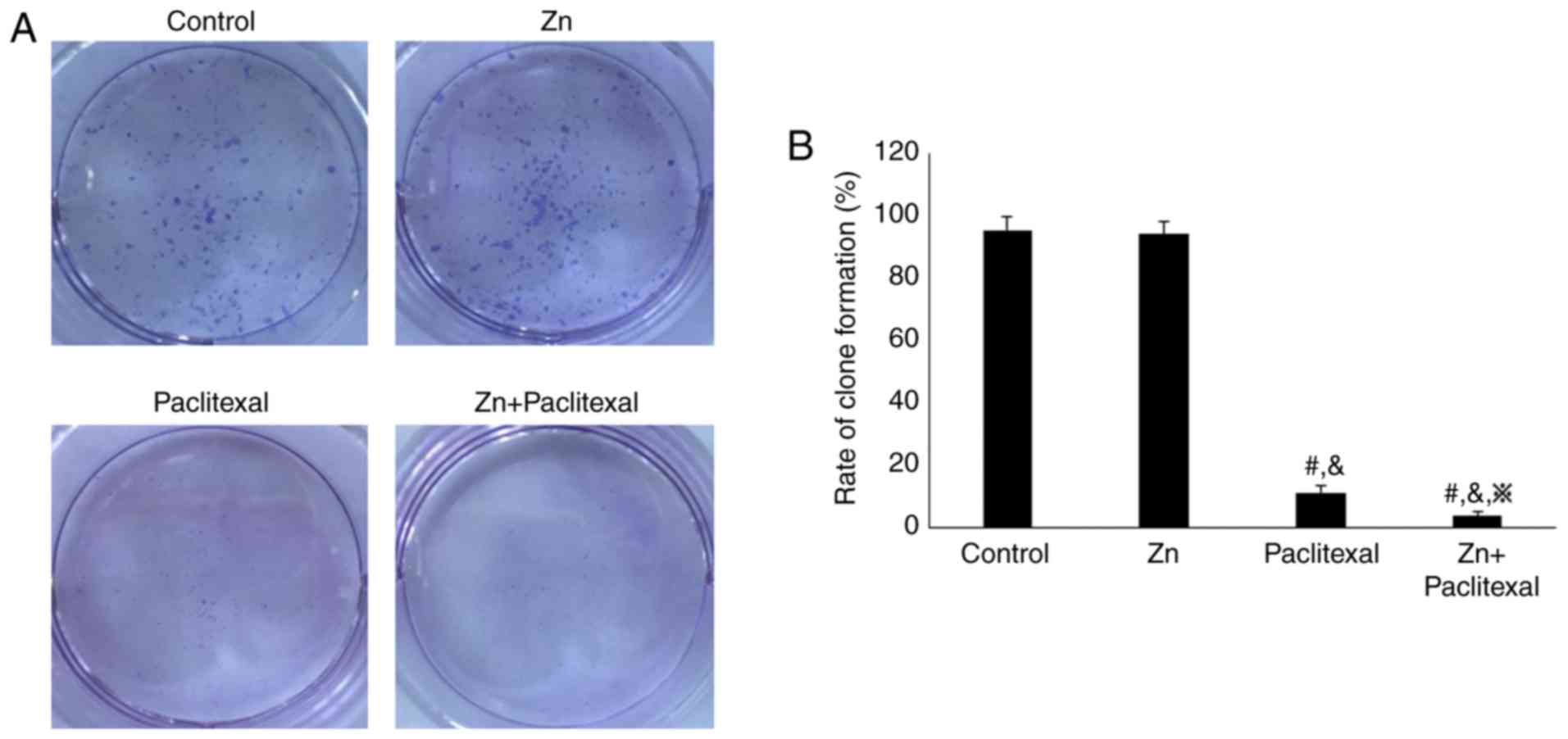
To explore the effect of zinc combined with
paclitaxel on the proliferative ability of PC-3 cells, a clone
formation assay was performed. The cell clone formation rate was
significantly reduced in the cells treated with paclitaxel only or
zinc combined with paclitaxel, compared with the control cells and
cells treated with zinc only (Fig.
2A). Furthermore, the cell clone formation rate was
significantly lower in cells treated with zinc combined with
paclitaxel than those treated with paclitaxel only (P<0.05;
Fig. 2B). This suggested that zinc
combined with paclitaxel significantly reduced the proliferation
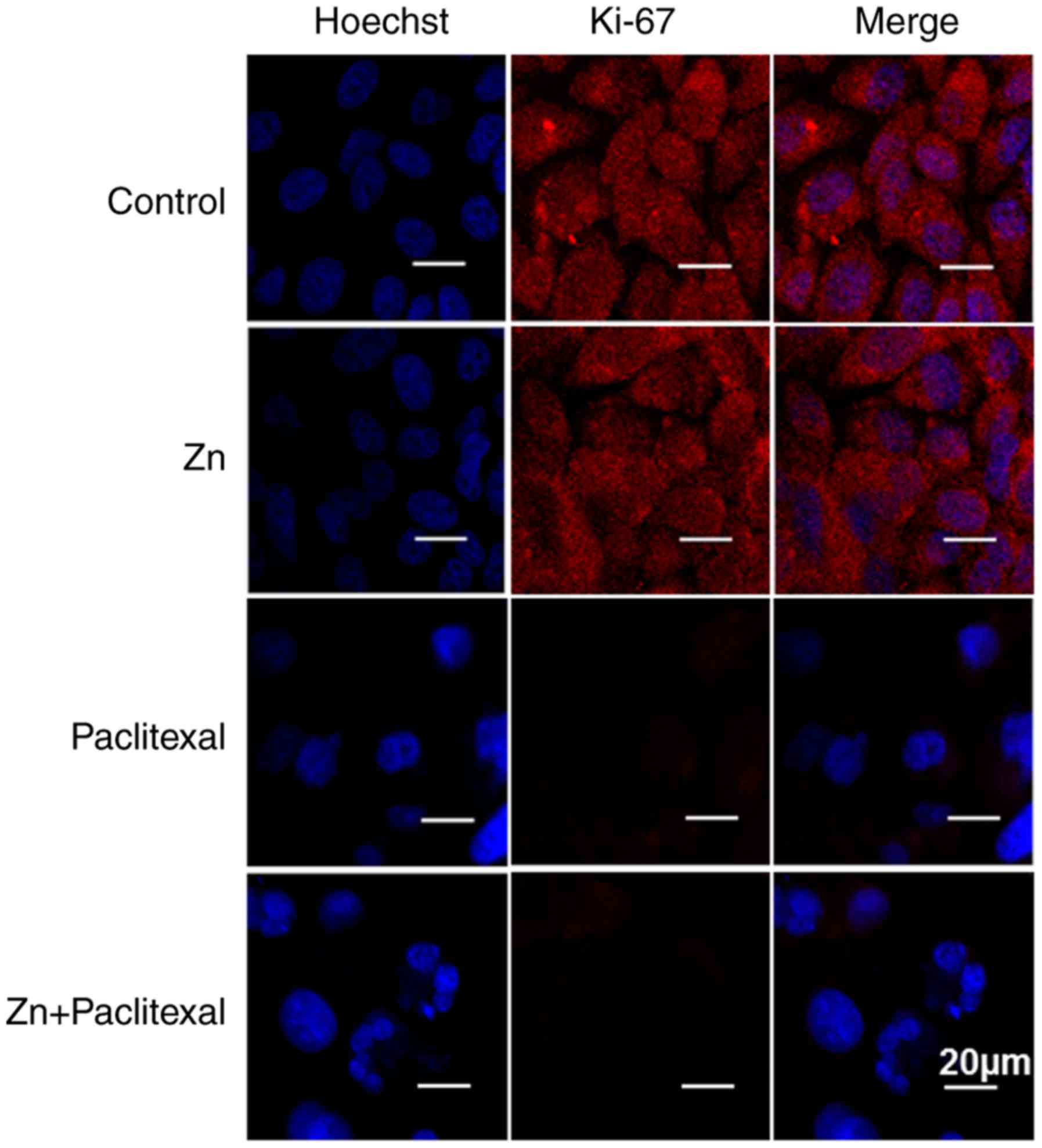
ability of PC-3 cells. Cell proliferation was further investigated
by immunofluorescence staining of Ki-67, visualized by confocal
microscopy (Fig. 3). Zinc combined
with paclitaxel significantly decreased the expression of Ki-67
protein, suggesting significant inhibition of the proliferative
ability of PC-3 cells.
Hoechst staining was used to analyze apoptosis
(Fig. 3). Zinc combined with
paclitaxel significantly enhanced the fluorescence intensity, and
nuclear fragmentation was evident. This indicated that zinc
combined with paclitaxel significantly induced PC-3 cell apoptosis
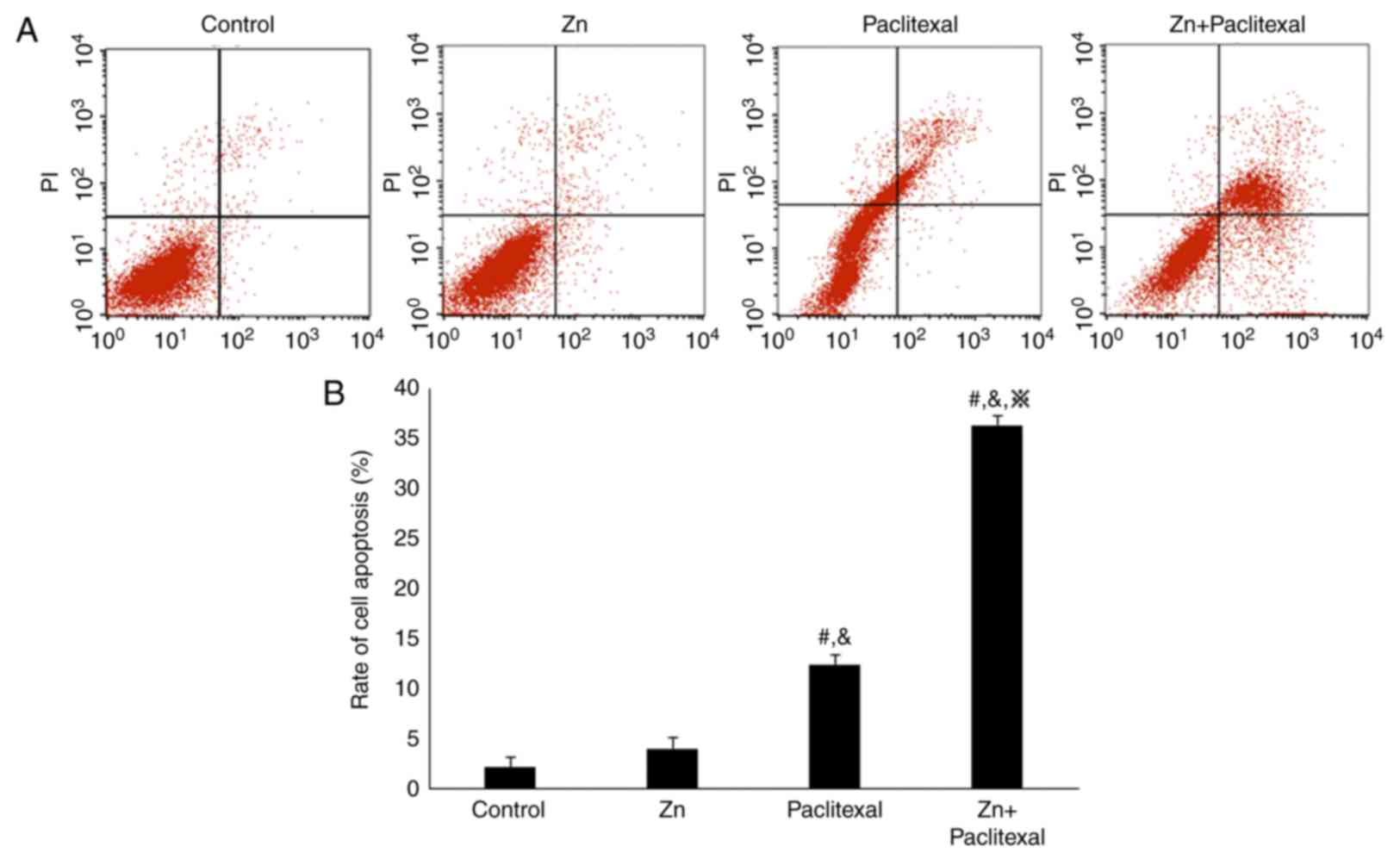
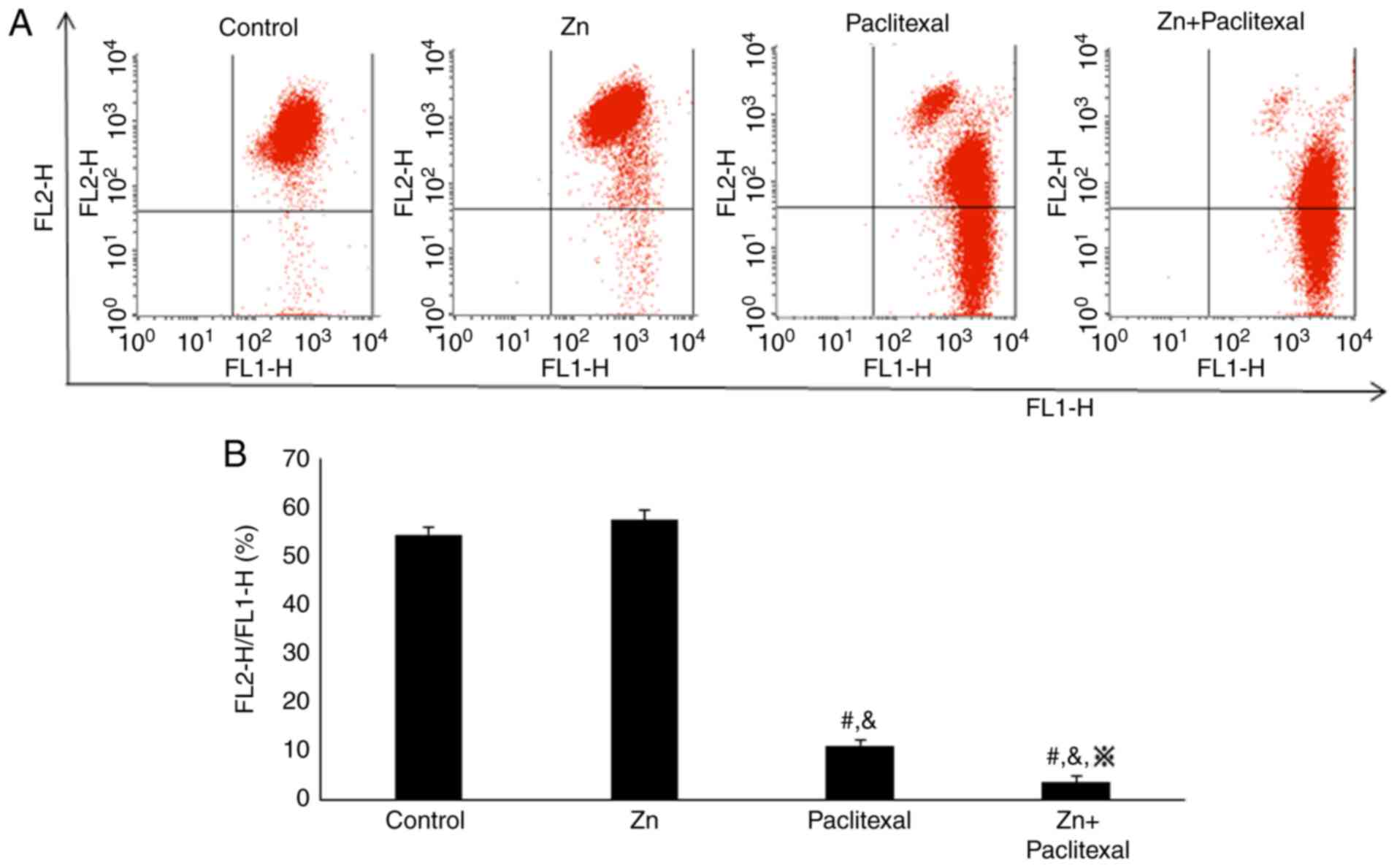
(P<0.05). Annexin V and PI staining and flow cytometry revealed
that, compared with control treatment (2.28±3.34%) and treatment
with zinc only (4.12±2.52%), treatment with paclitaxel alone
(12.48±4.34%) and zinc combined with paclitaxel (36.30±3.68%) both
increased the rate of PC-3-cell apoptosis, with the combination
treatment having most significant effect (P<0.05; Fig. 4A and B). These results suggested
that zinc enhanced the sensitivity of PC-3 prostate cancer cells to
paclitaxel chemotherapy.
A decrease in mitochondrial membrane potential is a
key feature of early-stage apoptosis. A change in JC-1 fluorescence
from red to green reflects a change in cell membrane potential.
Flow cytometry was used to assess the effect of zinc combined with
paclitaxel on the mitochondrial membrane potential of PC-3 cells
(Fig. 5A). Treating cells with zinc
combined with paclitaxel significantly decreased the ratio of red
to green fluorescence (P<0.05; Fig.
5B). This indicates that PC-3 cells treated with zinc and
paclitaxel exhibited significantly reduced mitochondrial function
compared with control cells, which may reflect early apoptosis. In
summary, zinc combined with paclitaxel promoted apoptosis and
inhibited cell proliferation. Hence, zinc increased the
chemosensitivity of PC-3 cells to paclitaxel.
Zinc combined with paclitaxel induced
the mitochondria-mediated apoptosis pathway
To investigate the mechanism by which zinc improved
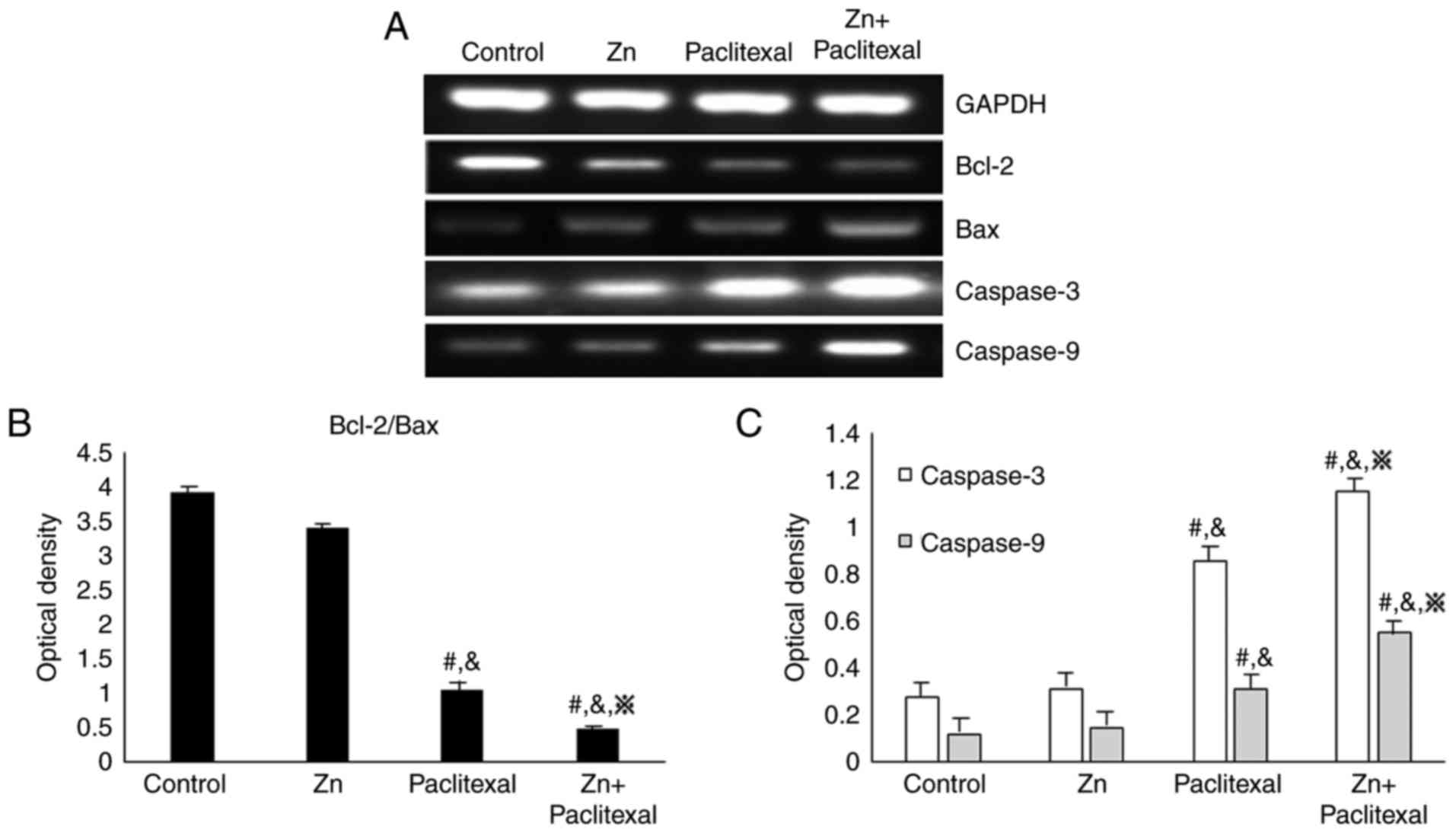
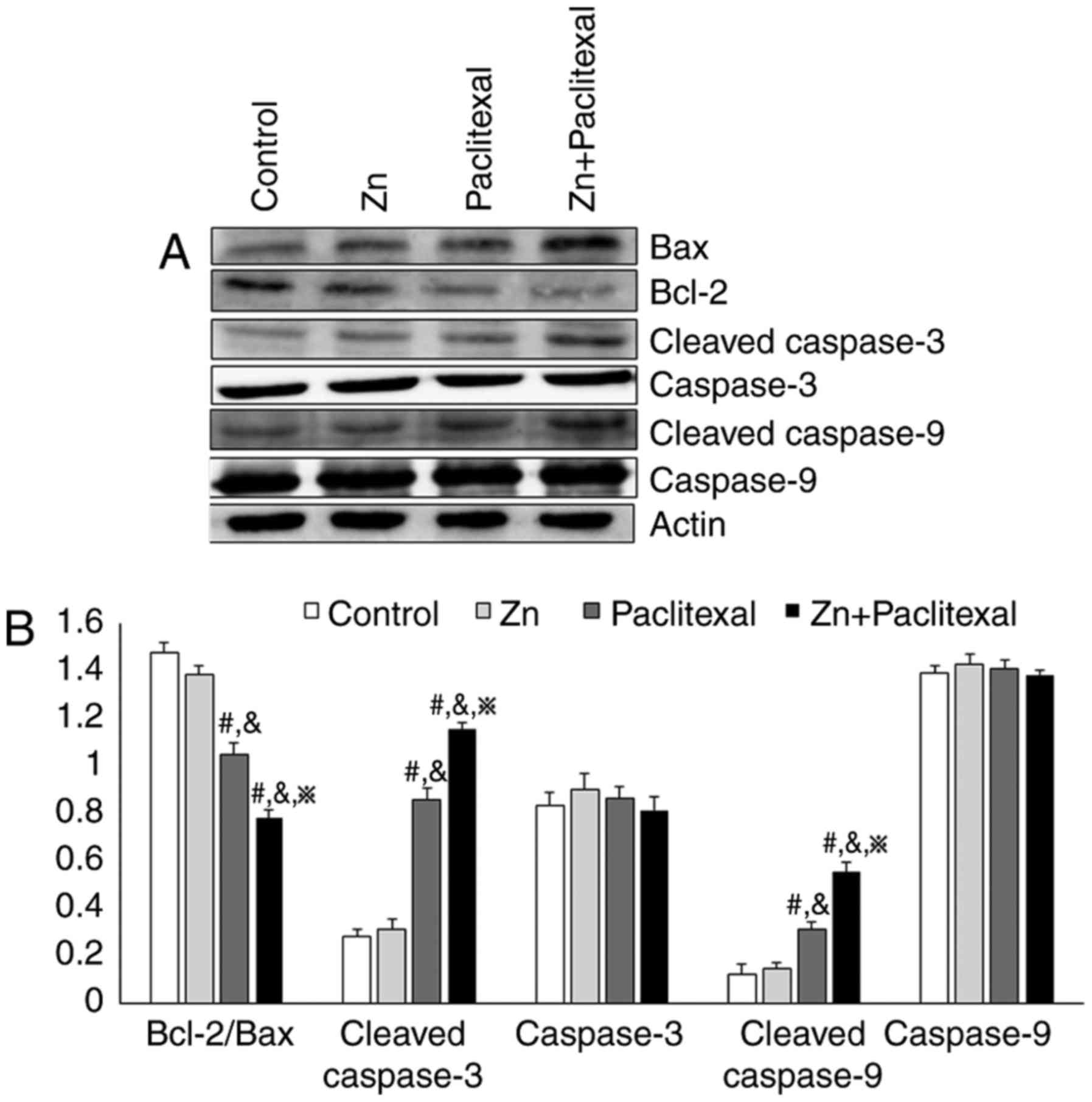
the sensitivity of PC-3 cells to paclitaxel chemotherapy, the mRNA
levels of Bax, Bcl-2, caspase-3 and caspase-9 by RT-PCR (Fig. 6A) and the protein expression levels
were analyzed by western blotting (Fig.
7A). There was a significant increase in the mRNA and protein
levels of cleaved caspase-3 and cleaved caspase-9 in PC-3 cells
treated with paclitaxel alone and paclitaxel combined with zinc,
compared with control cells or those treated with zinc alone
(P<0.05; Figs. 6C and 7B). There was no significant change in
protein levels of caspase-3 and caspase-9 (Fig. 7B). There was also a significant
decrease in the Bcl-2/Bax ratio (P<0.05; Figs. 6B and 7B). These results indicate that zinc
improved the sensitivity of PC-3 cells to chemotherapy, possibly
via effects on the mitochondria-mediated apoptosis pathway.
Discussion
Current prostate cancer treatment comprises surgery,
endocrine therapy, radiotherapy and chemotherapy. Seeing as
prostate cancer is often diagnosed at a late stage, the optimal
time for surgery is often missed (8,24),
making effective chemotherapy important. Paclitaxel has been
approved by the Food and Drug Administration as a first-line
treatment for hormone-refractory prostate cancer, and has been
reported to be the only current drug that can prolong survival and
improve quality of life in patients with tumors of the prostate
(4,25). Paclitaxel has been widely used in
the treatment of breast, ovarian, head and neck, and lung cancers.
However, chemotherapy with a single drug has been associated with
the development of drug resistance over time (9). Therefore, research based on
chemotherapeutic combination strategies is important.
Functional p53 enhances the sensitivity of prostate
cancer cells to chemotherapeutic agents, and loss of p53 function
in the PC-3 prostate cancer cell line increases paclitaxel
resistance (26). Zinc deficiency
is a cause of p53 mutation and inactivation (27). Zinc deficiency in prostate cells is
closely associated with the occurrence and development of prostate
cancer (2,28). Zinc is an essential trace element
for the function of ~200 enzymes in humans, thus serving an
important role in genome stability and processes, including gene
expression (9,20). Zinc concentrations have been
demonstrated to be decreased by 60–70% in the serum and cancerous
prostate tissue of patients with prostate cancer, and to decrease
further as the cancer progresses (29). We speculate that the reduction in
zinc concentration in prostate cancer may be associated with
chemotherapy resistance. Therefore, in the present study, PC-3
prostate cancer cells were treated with zinc to investigate whether
zinc could enhance sensitivity to chemotherapy.
Previous research has reported that physiological
concentrations of zinc can directly induce apoptosis of prostate
cancer cells via the mitochondrial pathway. The main mechanism of
this is the release of mitochondrial cytochrome c and the
activation of caspase-3, which triggers apoptosis (20,30).
In the mitochondria-mediated apoptosis pathway, in the presence of
deoxyadenosine triphosphate, cytochrome c binds with and
activates apoptotic protease activating factor-1 (Apaf-1), and this
complex further combines with procaspase-9 to form an apoptosis
complex that activates procaspase-9. Activated caspase-9 activates
other caspases, including caspase-3, thus inducing apoptosis
(31). The Bcl-2 gene family serves
an important role in this pathway. Some death receptor-independent
stimuli (including ultraviolet ray irradiation and chemotherapy)
activate the Bcl-2 gene family. Bcl-2 regulates the opening and
closing of the permeability transition pore (32,33).
To study the mechanism by which zinc enhances the chemotherapeutic
effectiveness of paclitaxel, the expression of apoptosis-associated
proteins was investigated by western blotting. The results indicate
that the mechanism may involve a decrease in Bcl-2 protein
expression levels, and an increase in Bax, cleaved caspase-3 and
cleaved caspase-9 protein expression levels (Fig. 7). It is likely that the
mitochondria-mediated apoptosis pathway was activated, causing
apoptosis via activation of caspases by a cascade reaction.
Although single-agent chemotherapy can have a
therapeutic effect against prostate cancer, patients with prostate
cancer patients are prone to chemotherapy resistance. This is
possibly due to severe zinc deficiency in prostate cancer tissues.
The present study suggests that zinc enhances the sensitivity of
the PC-3 prostate cancer cell line to the chemotherapeutic agent,
paclitaxel. Combining paclitaxel with zinc enhanced therapeutic
efficacy, and may have potential for overcoming issues associated
with single-agent treatment. The present study provides a basis for
novel strategies for tumor combination therapy.
Acknowledgements
We are grateful to Ruth Tunn, PhD, for editing the
English text of a draft of this manuscript.
Funding
The present study was supported by the Research Fund
of the National Natural Science Foundation of China (grant nos.
81572927, 81501982, 81772794, 81472419 and 81672948), the Jilin
Province Research Foundation for the Development of Science and
Technology Project (grant nos. 20150414025GH, 20170623021TC and
20160414005GH) and the Jilin University Bethune Plan B Projects
(grant no., 2015222).
Availability of data and materials
Some datasets are not available as the experimental
group is still performing further studies. However, other data is
available from the corresponding author on reasonable request.
Authors' contributions
YLi, YLiu, LS and PZ participated in the research
design. PZ, XT, HG, JL and RG conducted the experiments and
contributed to data acquisition and analysis. PZ, JS and YYC
performed the data analysis. PZ and YLiu contributed to the writing
of the manuscript. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Attard G, Parker C, Eeles RA, Schröder F,
Tomlins SA, Tannock I, Drake CG and de Bono JS: Prostate cancer.
Lancet. 387:70–82. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lavery HJ and Cooperberg MR: Clinically
localized prostate cancer in 2017: A review of comparative
effectiveness. Urol Oncol. 35:40–41. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Strano S, Dell'Orso S, Di Agostino S,
Fontemaggi G, Sacchi A and Blandino G: Mutant p53: An oncogenic
transcription factor. Oncogene. 26:2212–2219. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Gan L, Wang J, Xu H and Yang X: Resistance
to docetaxel-induced apoptosis in prostate cancer cells by
p38/p53/p21 signaling. Prostate. 71:1158–1166. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Giannakakou P, Gussio R, Nogales E,
Downing KH, Zaharevitz D, Bollbuck B, Poy G, Sackett D, Nicolaou KC
and Fojo T: A common pharmacophore for epothilone and taxanes:
Molecular basis for drug resistance conferred by tubulin mutations
in human cancer cells. Proc Natl Acad Sci USA. 97:2904–2909. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Li Y, Mizokami A, Izumi K, Narimoto K,
Shima T, Zhang J, Dai J, Keller ET and Namiki M: CTEN/tensin 4
expression induces sensitivity to paclitaxel in prostate cancer.
Prostate. 70:48–60. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Li Y, Zeng Y, Mooney SM, Yin B, Mizokami
A, Namiki M and Getzenberg RH: Resistance to paclitaxel increases
the sensitivity to other microenvironmental stresses in prostate
cancer cells. J Cell Biochem. 112:2125–2137. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Takeda M, Mizokami A, Mamiya K, Li YQ,
Zhang J, Keller ET and Namiki M: The establishment of two
paclitaxel-resistant prostate cancer cell lines and the mechanisms
of paclitaxel resistance with two cell lines. Prostate. 67:955–967.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Gan L, Chen S, Wang Y, Watahiki A, Bohrer
L, Sun Z, Wang Y and Huang H: Inhibition of the androgen receptor
as a novel mechanism of taxol chemotherapy in prostate cancer.
Cancer Res. 69:8386–8394. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ting HJ, Hsu J, Bao BY and Lee YF:
Docetaxel-induced growth inhibition and apoptosis in androgen
independent prostate cancer cells are enhanced by
1alpha,25-dihydroxyvitamin D3. Cancer Lett. 247:122–129. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ding WQ, Yu HJ and Lind SE: Zinc-binding
compounds induce cancer cell death via distinct modes of action.
Cancer Lett. 271:251–259. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kim S, Seo JW, Oh SB, Kim SH, Kim I, Suh N
and Lee JY: Disparate roles of zinc in chemical hypoxia-induced
neuronal death. Front Cell Neurosci. 9:12015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhao Y, Tan Y, Dai J, Wang B, Li B, Guo L,
Cui J, Wang G, Li W and Cai L: Zinc deficiency exacerbates diabetic
down-regulation of Akt expression and function in the testis:
Essential roles of PTEN, PTP1B and TRB3. J Nutr Biochem.
23:1018–1026. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kayatekin C, Zitzewitz JA and Matthews CR:
Zinc binding modulates the entire folding free energy surface of
human Cu, Zn superoxide dismutase. J Mol Biol. 384:540–555. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Fanzo JC, Reaves SK, Cui L, Zhu L, Wu JY,
Wang YR and Lei KY: Zinc status affects p53, gadd45, and
c-fos expression and caspase-3 activity in human bronchial
epithelial cells. Am J Physiol Cell Physiol. 281:C751–C757. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kelleher SL, McCormick NH, Velasquez V and
Lopez V: Zinc in specialized secretory tissues: Roles in the
pancreas, prostate, and mammary gland. Adv Nutr. 2:101–111. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Margalit O, Simon AJ, Yakubov E, Puca R,
Yosepovich A, Avivi C, Jacob-Hirsch J, Gelernter I, Harmelin A,
Barshack I, et al: Zinc supplementation augments in vivo antitumor
effect of chemotherapy by restoring p53 function. Int J Cancer.
131:E562–E568. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kloubert V and Rink L: Zinc as a
micronutrient and its preventive role of oxidative damage in cells.
Food Funct. 6:3195–3204. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Carraway RE and Dobner PR: Zinc pyrithione
induces ERK- and PKC-dependent necrosis distinct from TPEN-induced
apoptosis in prostate cancer cells. Biochim Biophys Acta.
1823:544–557. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Gallus S, Foschi R, Negri E, Talamini R,
Franceschi S, Montella M, Ramazzotti V, Tavani A, Dal Maso L and La
Vecchia C: Dietary zinc and prostate cancer risk: A case-control
study from Italy. Eur Urol. 52:1052–1056. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Seth R, Corniola RS, Gower-Winter SD,
Morgan TJ Jr, Bishop B and Levenson CW: Zinc deficiency induces
apoptosis via mitochondrial p53- and caspase-dependent pathways in
human neuronal precursor cells. J Trace Elem Med Biol. 30:59–65.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Gu J, Wang B, Liu Y, Zhong L, Tang Y, Guo
H, Jiang T, Wang L, Li Y and Cai L: Murine double minute 2
siRNA and wild-type p53 gene therapy interact positively
with zinc on prostate tumours in vitro and in vivo. Eur J Cancer.
50:1184–1194. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Shao Y, Liu Y, Shao C, Hu J, Li X, Li F,
Zhang L, Zhao D, Sun L, Zhao X, et al: Enhanced tumor suppression
in vitro and in vivo by co-expression of survivin-specific siRNA
and wild-type p53 protein. Cancer Gene Ther. 17:844–854. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Bataineh ZM, Hani Bani IH and Al-Alami JR:
Zinc in normal and pathological human prostate gland. Saudi Med J.
23:218–220. 2002.PubMed/NCBI
|
|
25
|
Wang C, Huang SB, Yang MC, Lin YT, Chu IH,
Shen YN, Chiu YH, Hung SH, Kang L, Hong YR, et al: Combining
paclitaxel with ABT-263 has a synergistic effect on paclitaxel
resistant prostate cancer cells. PLoS One. 10:e01209132015.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Liu C, Zhu Y, Lou W, Nadiminty N, Chen X,
Zhou Q, Shi XB, deVere White RW and Gao AC: Functional p53
determines docetaxel sensitivity in prostate cancer cells.
Prostate. 73:418–427. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Krone CA and Harms LC: Re: Zinc supplement
use and risk of prostate cancer. J Natl Cancer Inst. 95:1556–1557.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Costello LC and Franklin RB: The clinical
relevance of the metabolism of prostate cancer; zinc and tumor
suppression: Connecting the dots. Mol Cancer. 5:172006. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Stepien M, Hughes DJ, Hybsier S, Bamia C,
Tjonneland A, Overvad K, Affret A, His M, Boutron-Ruault MC, Katzke
V, et al: Circulating copper and zinc levels and risk of
hepatobiliary cancers in Europeans. Br J Cancer. 116:688–696. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Costello LC, Franklin RB and Feng P:
Mitochondrial function, zinc, and intermediary metabolism
relationships in normal prostate and prostate cancer.
Mitochondrion. 5:143–153. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Yang Y, Zong M, Xu W, Zhang Y, Wang B,
Yang M and Tao L: Natural pyrethrins induces apoptosis in human
hepatocyte cells via Bax- and Bcl-2-mediated mitochondrial pathway.
Chem Biol Interact. 262:38–45. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Shan M and Fan TJ: Cytotoxicity of
carteolol to human corneal epithelial cells by inducing apoptosis
via triggering the Bcl-2 family protein-mediated mitochondrial
pro-apoptotic pathway. Toxicol In Vitro. 35:36–42. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Gibson CJ and Davids MS: BCL-2 Antagonism
to target the intrinsic mitochondrial pathway of apoptosis. Clin
Cancer Res. 21:5021–5029. 2015. View Article : Google Scholar : PubMed/NCBI
|