Introduction
Breast cancer is a heterogeneous disease and one of
the major health issues confronting women. Its initiation and
progression could be influenced by numerous factors, including
environmental and genetic factors (1–3).
Currently, treatment with conventional chemotherapeutic drugs for
breast cancer is limited, due to issues including poor solubility
and multidrug resistance (4).
Challenges in breast cancer treatment include the combinations of
chemotherapeutic drugs that lead to adverse effects, a high level
of heterogeneity from patient to patient, and long-term drug
administration, with resistance and recurrence of cancer following
treatment (5,6).
Paclitaxel is a first-line therapeutic clinical
agent used to treat breast cancer, which exerts its antitumor
activity by promoting the polymerization of tubulin and stabilizing
the resulting microtubules, causing cell cycle arrest that leads to
apoptosis of cancer cells (7).
However, paclitaxel resistance often occurs after a short period of
treatment, causing a serious problem in chemotherapy that
eventually results in the patient succumbing to the disease due to
tumor metastasis (7,8).
In recent decades, natural products have formed an
important part of drug discovery. Among these natural products,
curcumin (diferuloylmethane), a hydrophobic polyphenol derived from
turmeric (Curcuma longa) has been used for the treatment of
numerous diseases, including cancer (9). Curcumin possesses several beneficial
properties, including anti-inflammatory, anti-metastatic and
anti-carcinogenic effects, as well as acting as an inhibitor of
tumor formation (10,11). Curcumin is considered to be a
chemopreventive and chemotherapeutic agent that exerts in
vitro anticancer activities in breast cancer cells, inducing
apoptosis through modulation of anti- and pro-apoptotic proteins
such as B-cell lymphoma 2 (Bcl-2) and Bax, respectively (10,11).
Apoptosis, a type of programmed cell death, is an
essential process for regulating homeostasis by eliminating
redundant and abnormal cells, and is associated with various
diseases, particularly cancer (12). Apoptosis is deregulated in
carcinoma, thus, is one of the most studied processes in cancer
therapy. In general, apoptosis is controlled by two principal
pathways, including the death receptor-mediated (extrinsic) and the
mitochondrial-dependent (intrinsic) pathways (13). The latter process is initiated by
loss of membrane integrity and mitochondrial depolarization, which
is regulated by members of the Bcl-2 protein family, triggering the
release of cytochrome c into the cytosol, thereby activating
caspase-3 as an effector (13).
Poly(ADP-ribose) polymerase-1 (PARP-1) is another
molecule involved in several key biological processes, including
apoptosis, cell proliferation control, replication and DNA damage
repair (14). PARP is a target of
caspase protease activity and is also associated with apoptosis.
When PARP detects DNA damage, it is auto-modified and recruits
repair factors to the DNA damage sites (15,16).
Defects in DNA repair lead to genomic instability and play a
critical role in cancer development. Abnormal PARP-1 expression has
been reported in various cancer types; increased PARP-1 expression
was noted in various cell lines exhibiting tumor progression
(14).
Nuclear factor-κB (NF-κB), a protein complex that
controls transcription of DNA, is involved in modulation of
inflammatory response through expression of multiple
pro-inflammatory proteins (17).
Tumor cells may enhance NF-κB activity through increasing cytokine
release from stromal cells and fibroblasts in the tumor
microenvironment. NF-κB overexpression has largely been manifested
in triple-negative breast cancer (17). A previous study reported
constitutive activation of NF-κB, resulting in overexpression of
anti-apoptotic genes in breast cancer. However, the precise
mechanisms for activation of NF-κB are not clear (18).
The function of proliferating cell nuclear antigen
(PCNA) is essential for the maintenance of genomic integrity in
actively growing cells. PCNA serves key functions in the metabolism
of nucleic acid. Its primary function is in DNA replication, but it
is also involved in RNA transcription, chromatin assembly, cell
cycle control and DNA excision repair (19). PCNA is expressed in actively
proliferative human cancer cells, and it is identical to a ‘cyclin’
protein identified in cell proliferation (20).
In the current study, we aimed to ascertain whether
curcumin and paclitaxel induced apoptosis, analyzed by protein
expression and flow cytometry, in MCF-10F MCF7 and MDA-MB-234 cell
lines. MCF-10F is an immortalized breast cell line that retains all
of the characteristics of normal epithelium in vitro,
including anchorage-dependence, non-invasiveness and
non-tumorigenicity in nude mice. MCF7 is an epithelial and
luminal-like adenocarcinoma cell line triple positive for estrogen
and progesterone receptor. MDA-MB-234 is a metastatic human breast
cancer cell line triple negative for such receptors. The aim of the
current study was to evaluate the effect of curcumin and paclitaxel
in human breast cancer cell lines on loss of membrane integrity,
caspase expression, PARP function and NF-κB expression, and their
associated effects on apoptosis.
Materials and methods
Breast cancer cell lines
The MCF-10F cell line [American Type Culture
Collection (ATCC), Manassas, VA, USA] was grown in DMEM/F-12 (1:1)
medium supplemented with antibiotics (100 U/ml penicillin, 2.5
µg/ml amphotericin B and 100 µg/ml streptomycin; all from Thermo
Fisher Scientific, Inc., Waltham, MA, USA), 0.5 µg/ml
hydrocortisone (Sigma-Aldrich; Merck KGaA, Darmstadt, Germany), 10
µg/ml of 5% equine serum (Biofluids, Inc., Rockville, MD, USA) and
0.02 µg/ml epidermal growth factor (Collaborative Research, Inc.,
Bedford, MA, USA). MCF7 cells (HTB-22™; ATCC) were grown in Minimum
Essential Medium supplemented with 10% fetal bovine serum (FBS).
MDA-MB-231 cells (HTB-26™; ATCC) were grown in RPMI supplemented
also with 10% FBS. The cells were incubated at 37°C with 5%
CO2 until reaching 70% confluence. Curcumin and
paclitaxel (Sigma-Aldrich; Merck KGaA) were dissolved in dimethyl
sulfoxide (DMSO) (0.1%). All treatments were performed at 37°C.
Cells were harvested and seeded at 4×105 cells/ml
culture medium into T75 flasks (Corning Costar, Corning, NY, USA).
After a 24-h pre-incubation period allowing cells to attach, the
culture medium was replaced by either medium without drugs or
medium with 30 µM curcumin and/or 10 nM paclitaxel for 48 h.
CellEvent Caspase-3/7 assay
To determine apoptosis by activation of caspase 3/7,
the CellEvent Caspase-3/7 green detection reagent kit (Thermo
Fisher Scientific, Inc.) was used. The kit contains a substrate for
activated caspases 3 and 7, producing a fluorogenic response
indicative of apoptosis. This was used together with the
SYTOX® AADvanced dead cell stain (Thermo Fisher
Scientific, Inc.) to discriminate necrotic from live cells. The
fluorescence emission maximum of the dye was at ~520 nm. The
acquisition process started following 30 min of incubation and
analysis was performed using Beckman Coulter FC500 Flow Cytometry
system with CXP software (Beckman Coulter, Inc., Brea, CA, USA)
within 1 h. Experiments were performed in triplicate.
7-Amino actinomycin D (7-AAD)/Annexin
V assay
Phosphatidylserine (PS) is located inside the cell
membrane in normal cells and is transferred to the surface during
the early stage of cell apoptosis. Annexin V, a
Ca2+-dependent phospholipid binding protein, has a
strong binding affinity for PS. An Annexin V-FITC/7-AAD staining
kit was used to assess curcumin or paclitaxel-induced cell
apoptosis. MCF7 and MDA-MB-231 cell lines were cultured to 70%
confluence, and then curcumin and/or paclitaxel was added. After 48
h, cells were harvested and washed twice with pre-cooled PBS and
then resuspended in 1X binding buffer at a concentration of
1×106 cells/ml. Then, 100 µl of this solution
(1×105 cells) was mixed with 10 µl Annexin V-FITC and 20
µl 7-AAD (Beckman Coulter), according to the manufacturer's
instructions. The mixed solution was incubated at room temperature
(25°C) in the dark for 15 min. Then, 400 µl 1X dilution buffer was
added to each tube. Analysis was performed using Beckman Coulter
FC500 Flow Cytometry system with CXP software (Beckman Coulter)
within 1 h. Experiments were performed in triplicate.
PARP cleavage
PARP cleavage was used to detect DNA damage in the
cell cultures (PE Mouse Anti-Cleaved PARP, Asp214; BD Biosciences,
San Diego, CA, USA). Curcumin (30 µM) and/or paclitaxel (10 nM)
were added per 1×106 proliferating breast cells. Cells
were incubated for 48 h at 37°C. Cells (treated and untreated) were
washed with cold PBS; then re-suspended in Cytofix/Cytoperm™
solution at 2×106 cells/ml. Cells were kept for 20 min
on ice. Cells were pelleted, aspirated and the Cytofix/Cytoperm
solution was discarded. Then, cells were washed twice at room
temperature with 0.5 ml Perm/Wash™ buffer per 1×106
cells, and the supernatants were discarded. Cells were re-suspended
in Perm/Wash buffer at 10×106/ml. Test samples were
aliquoted at 1×106 cells per 100-µl test. Antibody (20
µl per test) was added, and incubated for 30 min at room
temperature. Each test was washed in 1.0 ml Perm/Wash Buffer and
the supernatant was discarded. Each test was re-suspended in 0.5 ml
Perm/Wash Buffer and analyzed by flow cytometry. Analysis was
performed using Beckman Coulter FC500 Flow Cytometry system with
CXP software (Beckman Coulter) within 1 h. Experiments were
performed in triplicate.
Western blot analysis
To determine protein expression, western blotting
was performed. Cells were lysed with 1 ml lysis buffer (pH 7.2;
Tris Base, 50 mM), NaCl (100 mM), EDTA (1 mM), orthovanadate (1
mM), PMSF (1 mM) and Triton X-100 (0.1%) and centrifuged (13,200
rpm, 15 min). The supernatant with cellular proteins was dissolved
in SDS-PAGE sample solution containing 60 mM Tris (pH 6.5), 10%
(w/v) glycerol, 5% (w/v) β-mercaptoethanol, 20% (w/v) SDS, and
0.025% (w/v) bromophenol blue and denatured by boiling (2 × 5 min),
and vortex mixing (2 × 30 sec). The total amount of protein was 50
µg in each lane with standard protein markers (Bio-Rad
Laboratories, Inc., Hercules, CA, USA). Following fractionation by
SDS-PAGE on gels (7×14 cm), proteins were electro-blotted onto PVDF
membranes (GE Healthcare, Chicago, IL, USA) using a blotting
apparatus (Bio-Rad Laboratories, Inc.). Membranes were blocked for
2 h in 10% non-fat dry milk-TBS-0.1% Tween-20 and then incubated
for 2 h at room temperature with corresponding primary antibodies
(1:200): NF-κB (sc-53744), caspase-3 (sc-7148), PCNA (sc-56), and
β-actin (sc-47778). This was followed by incubation with secondary
peroxidase-conjugated mouse IgG (1:5,000; Cell Signaling
Technology, Inc., Danvers, MA, USA) in 5% non-fat dry milk-TBS-0.1%
Tween-20. All steps were performed at room temperature, and blots
were rinsed between incubation steps with TBS-0.1% Tween-20. Cell
blots were probed with mouse anti β-actin antibody as control.
Immunoreactive bands were visualized using the ECL™ Western
Blotting Detection Reagent detection method (GE Healthcare) and
exposure of the membrane to X-ray film. Protein determination was
performed using the Bicinchoninic Acid Method (Bio-Rad
Laboratories, Inc.) with BSA as the standards. Experiments were
performed in triplicate.
Statistical analysis
Data are expressed as the average ± standard error
of the mean. Comparisons between untreated and treated groups were
analyzed by analysis of variance and Dunnett's test. P<0.05 was
considered to indicate a statistically significant difference.
Assays were performed at least three times independently.
Results
The present study evaluated the apoptotic effect of
curcumin, paclitaxel and their combined administration in human
breast cancer cell lines MCF-10F, MCF7 and MDA-MB-231. Apoptosis
and necrosis were analyzed by flow cytometry using the Cell Event
Caspase-3/7 kit and 7-AAD/Annexin V assay, and by western blotting,
analysis of PARP protein activation, and evaluation of cell
proliferation via PCNA.
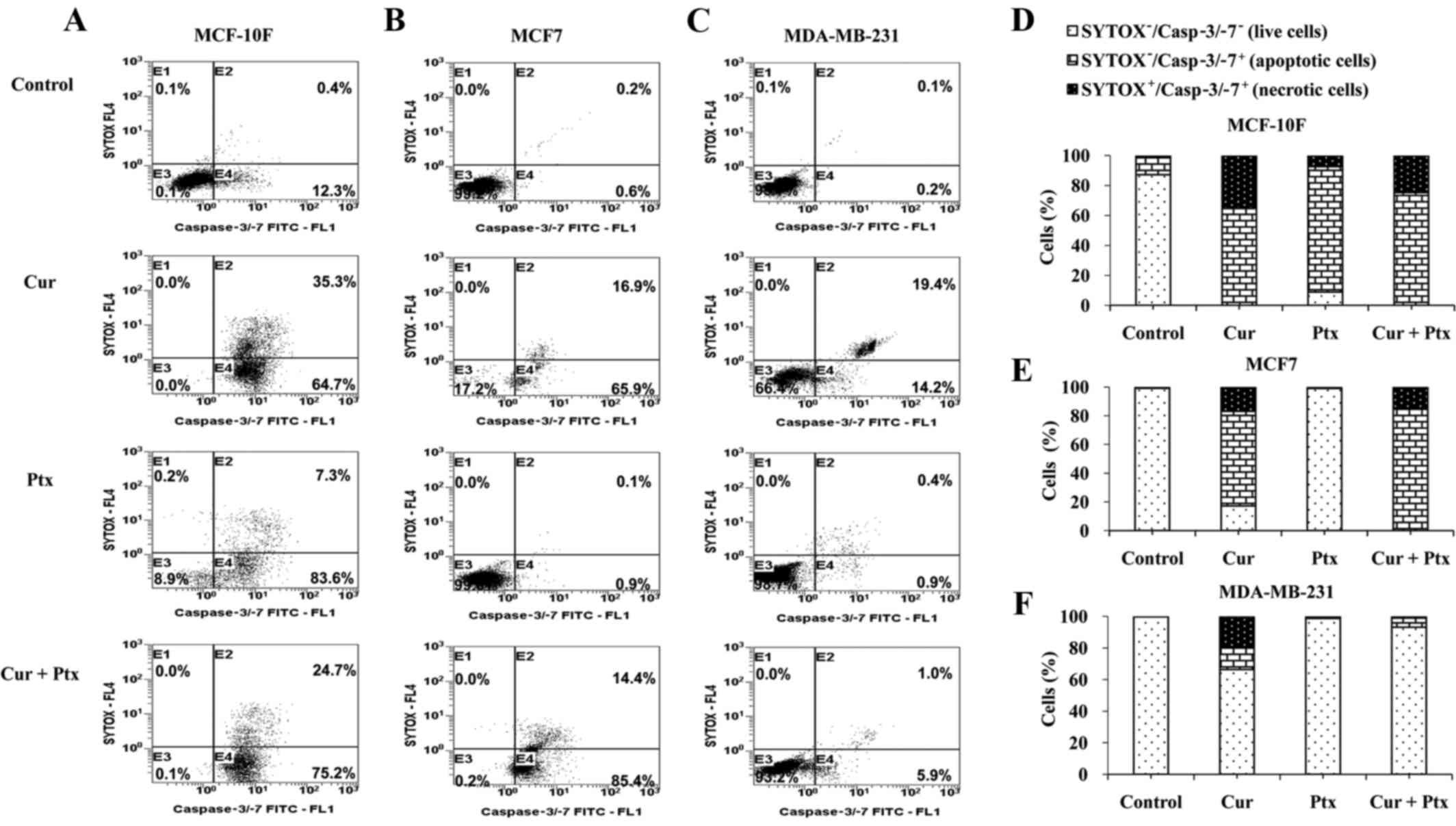
Analysis of caspase 3/7 activity
A distinctive feature of the early stages of
apoptosis is the activation of caspases. This was evaluated in the
current study using the Cell Event Caspase-3/7 kit. Fig. 1A-F shows dot-plot and representative
graphs with results that indicate the effect of curcumin,
paclitaxel and the combined treatment on caspase 3/7 activity, as
analyzed by flow cytometry. Fig. 1A
indicates that paclitaxel induced a higher level of apoptosis
(83.6%) compared with curcumin (64.7%) or the combined treatment
(75.2%) in the MCF-10F cell line. Paclitaxel alone induced only
7.3% of necrosis. However, curcumin alone induced 35.3% and the
combined treatment induced 24.7% necrosis in the MCF-10F cell line
(Fig. 1A and D). However, the
results in Fig. 1B indicate that
curcumin (65.9%) and the combined treatment (85.4%) induced a
higher level of apoptosis compared with paclitaxel alone (0.9%) in
the MCF7 cell line and there was no significant induction of
necrosis by any treatment given (Fig.
1B and E). As indicated in Fig.
1C, curcumin induced 14.2% apoptosis and 19.4% necrosis in the
MDA-MB-231 cell line. There was no evidence of necrosis following
paclitaxel or combined treatment in the MDA-MB-231 cell line
(Fig. 1C and F).
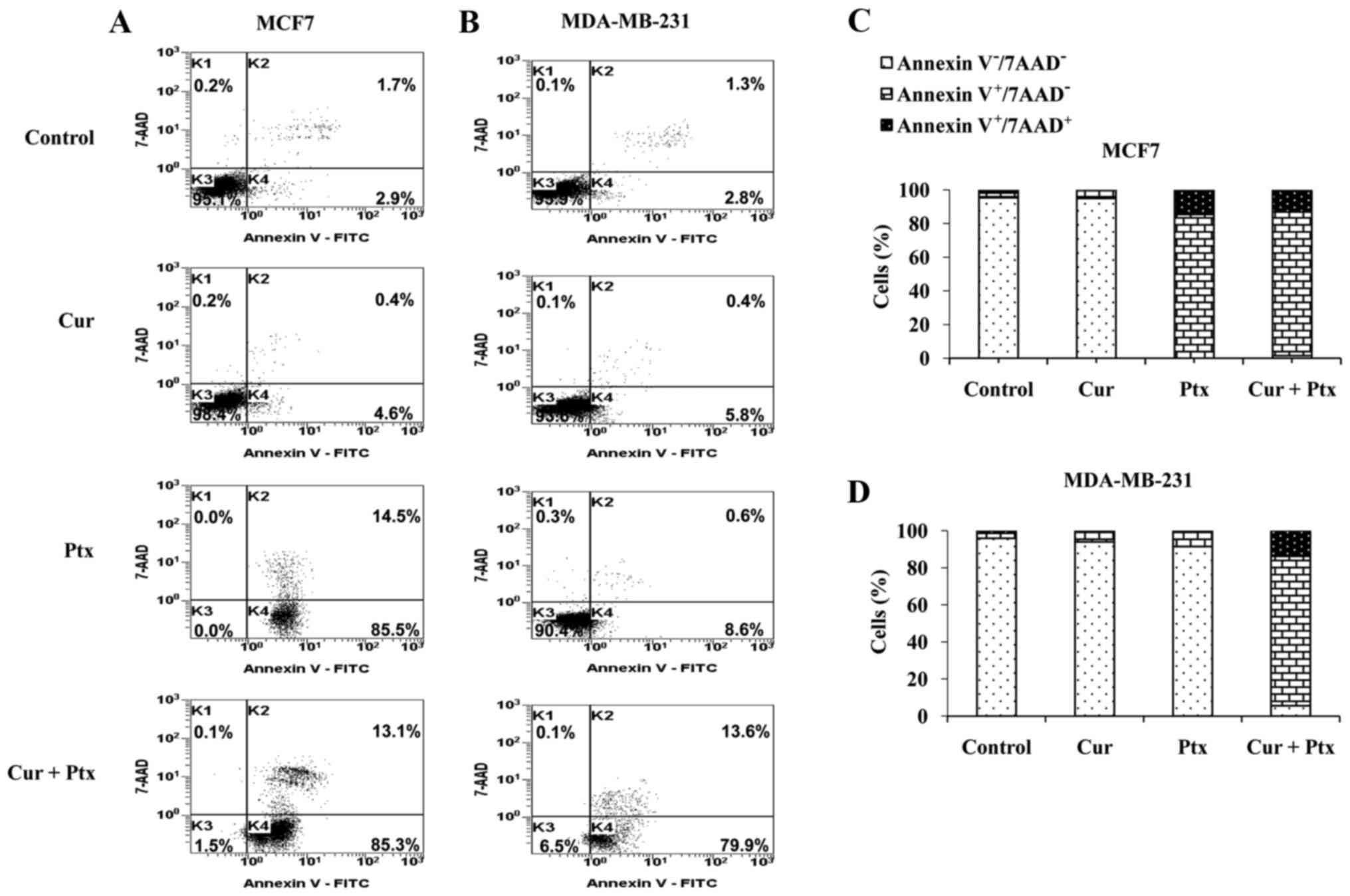
Analysis of cell membrane
integrity
The loss of membrane integrity was analyzed by
7-AAD/Annexin V assay (Fig. 2A-D).
As shown in Fig. 2A and C,
paclitaxel or the combined treatment induced increased levels of
apoptosis (85.5 and 85.3%, respectively) compared with curcumin
alone (4.6%) in the MCF7 cell line. In contrast, no significant
effect on apoptosis was observed following paclitaxel or curcumin
treatment in the MDA-MB-231 cell line compared with the control
(Fig. 2B). However, combined
treatment with paclitaxel and curcumin induced 79.9% apoptosis
(Fig. 2B and D). However, necrosis
was not observed in these two cell lines.
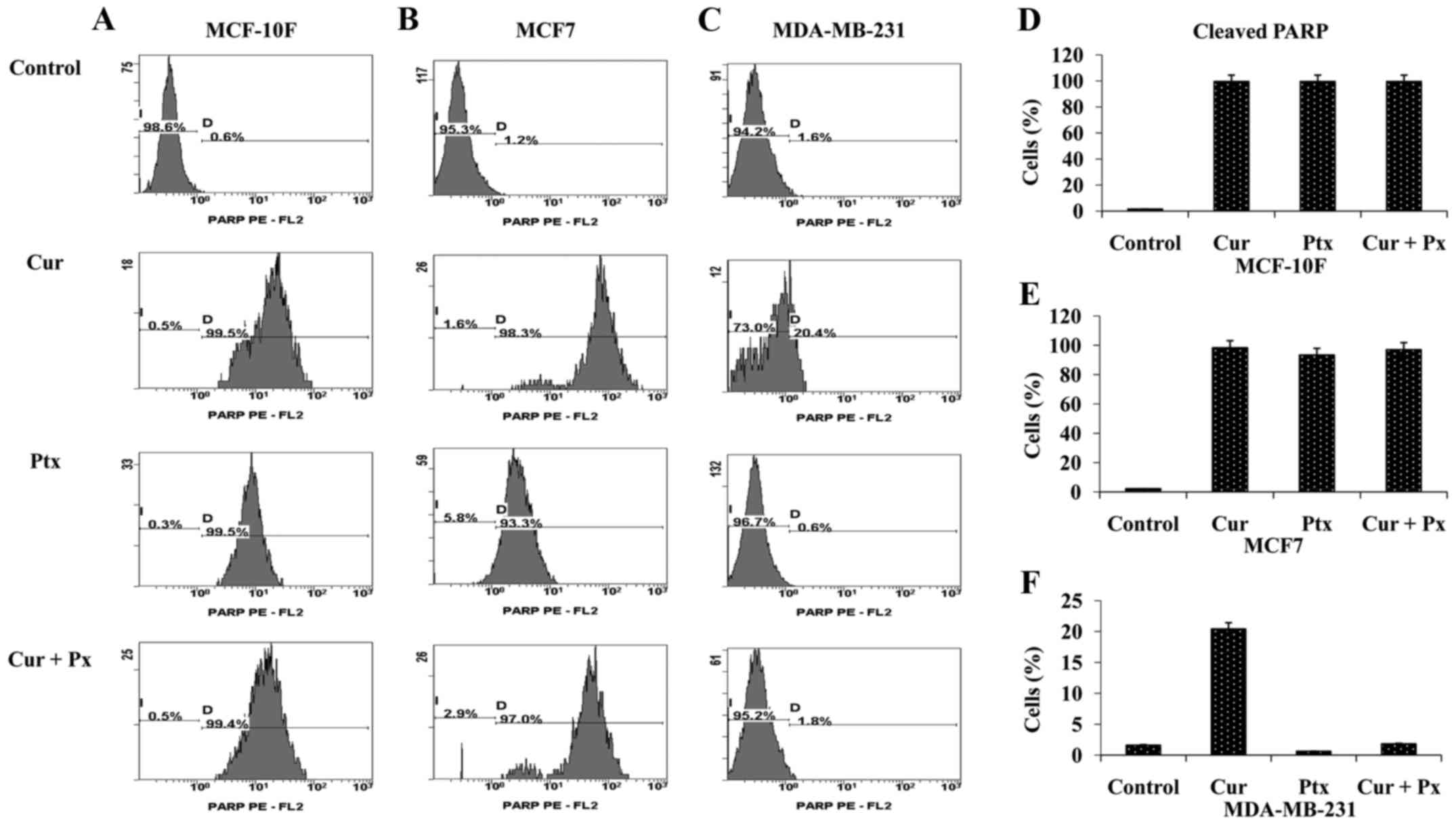
Analysis of PARP activation
Since PARP is a target of caspase protease activity
and is also associated with apoptosis, its activation was evaluated
in the present study (Fig. 3A-F). A
total of 99.5% of cells treated with curcumin, paclitaxel and the
combined treatment had induced PARP-cleavage activation in the
MCF-10F cell line (Fig. 3A). A
graph of these results is presented in Fig. 3D. A similar percentage of cells
exhibited PARP activation in the MCF7 cell line, as indicated in
Fig. 3B. A graph of these results
is presented in Fig. 3E. On the
other hand, only curcumin treatment induced PARP activation in the
MDA-MB-231 cell line (Fig. 3C).
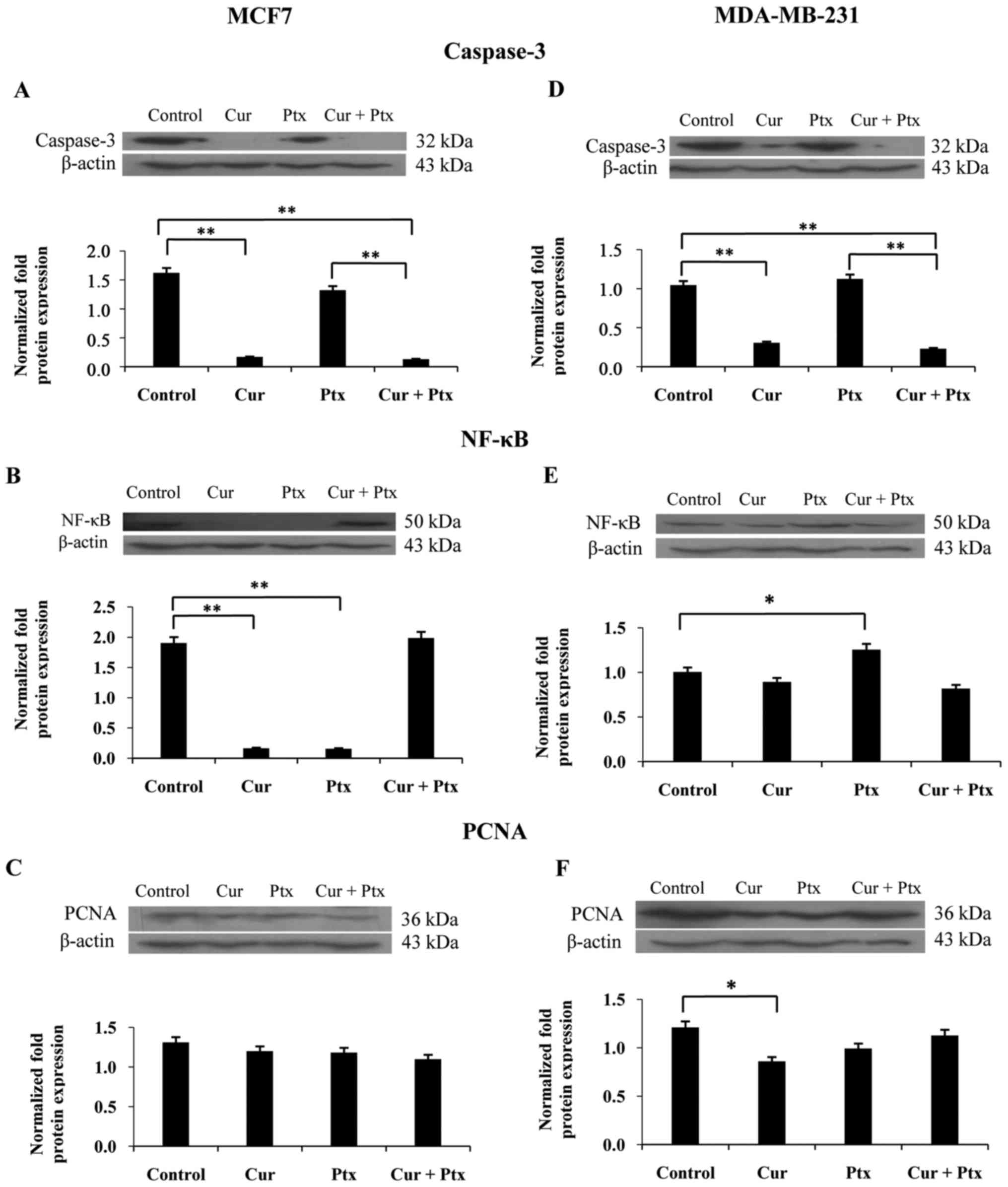
Western blot analysis
The results of the western blot analysis of the MCF7
and MDA-MB-231 cell lines following treatment with curcumin alone,
paclitaxel alone and combined with paclitaxel are shown in Fig. 4A-F. Curcumin alone and combined with
paclitaxel significantly decreased caspase-3 protein expression in
the MCF7 cells (P<0.05; Fig.
4A), as analyzed by western blotting. Curcumin alone and the
combined treatment significantly decreased caspase-3 expression in
the MDA-MB-231 cell line (P<0.01; Fig. 4D).
NF-κB is a transcription factor that regulates the
expression of hundreds of genes that are involved in regulating
cell growth and apoptosis pathways in the cytoplasm. As indicated
in Fig. 4B, curcumin and paclitaxel
alone significantly decreased NF-κB protein expression in the MCF7
cell line (P<0.01). By contrast, paclitaxel alone significantly
increased NF-κB protein expression in the MDA-MB-231 cell line
(P<0.05; Fig. 4E).
PCNA was analyzed in the current study since it is
involved in numerous molecular pathways responsible for the life
and death of mammalian cells. As indicated in Fig. 4C, curcumin or paclitaxel did not
induce any significant changes in PCNA protein expression in the
MCF7 cell line. However, as shown in Fig. 4F, curcumin alone significantly
inhibited PCNA protein expression in comparison with the control in
MDA-MB-231 cells (P<0.05).
Discussion
In the present study, the induction of apoptosis was
studied through several processes that indicate cell death, cell
necrosis, activation of proteins and protein expression. Among the
substances analyzed, curcumin and the combined treatment with
curcumin and paclitaxel induced a lower rate of apoptosis compared
with paclitaxel alone in the MCF-10F cell line. However, curcumin
and the combined treatment induced a higher rate of apoptosis
compared with paclitaxel alone in the MCF7 cell line, and there was
no induction of necrosis by any treatment. It was notable that
there was no evidence of necrosis following paclitaxel or combined
treatment in the MDA-MB-231 cell line.
In apoptosis, caspase-3 is the most important
effector. This molecule is inactive inside the cell. Once the
apoptotic cascade is triggered, it undergoes cleavage and
subsequent activation (21). The
current study indicated that curcumin alone and combined with
paclitaxel decreased caspase-3 protein expression in the MCF7 and
MDA-MB-231 cell lines. Similar results have been obtained with
curcumin in the Tumor2 cell line (22). On the other hand, drugs such as 5-FU
were found to increase caspase-3 expression in Tumor2 and
MDA-MB-231 cell lines as an indication of apoptosis (23). The ability of curcumin to induce
apoptosis in tumor cells, and/or potentiate apoptosis induction by
classical chemotherapeutic drugs, supports its potential in
anticancer therapies. It has previously been indicated that
curcumin activates caspase-3 (24).
Others have found that the combination of Trichostatin A with 20 mM
of curcumin increased caspase-3 cleavage products (24). It has been demonstrated that
dimethoxy curcumin, an analogue of curcumin, has good chemical and
metabolic stability, influencing the activation of caspase-3 in
vitro and in vivo (25).
It has also been reported that combined paclitaxel and curcumin
enhances caspase-3/7 activities in SKOV3 and SKOV3TR cells
(26).
It is interesting to note that apoptosis may be
evaluated via two methods involving caspase 3/7 and Annexin V;
however, in principle these two processes detect apoptosis in
different ways. Caspase 3/7 determines apoptosis through activation
of effector caspases, while Annexin V is related to loss of cell
membrane integrity evidenced by phosphatidylserine migration from
the inner to outer cellular surface of the membrane.
Paclitaxel (10 nM) combined with curcumin (30 µM)
induced loss of membrane integrity, indicating increased apoptosis
in comparison with curcumin alone in the MCF7 cell line. However,
only combined treatment induced apoptosis in the MDA-MB-231 cell
line. Previous results indicated that the concentrations used were
similar (27–29). Other studies have demonstrated the
apoptotic effect of curcumin combined with paclitaxel via decreased
gene expression of Bcl-xL in comparison with the control or with
either substance alone in MCF7 and MDA-MB-231 cell lines. The
results indicated that curcumin or paclitaxel alone, as well as
combined treatment, decreased Bcl-2 and increased Bax protein
expression in comparison with the control in MCF7 cells (27).
Activation of PARP was evaluated in the current
study as a target of caspase protease activity, and is associated
with apoptosis. It was notable to observe that 99.5% of cells
treated with curcumin, paclitaxel or combined treatment induced
PARP activation in the MCF-10F cell line. A similar percentage of
cells was observed to induce PARP activation in the MCF7 cell line.
Previous results found that PARP-1 was cleaved upon curcumin
treatment in the malignant and tumorigenic Tumor2 cell line from an
experimental breast cancer model exposed to ionizing radiation
(alpha particles) and estradiol (28–30).
This is similar to the results for the metastatic MDA-MB-231 cell
line in the current study, in which only curcumin treatment induced
PARP activation or cleavage.
Previous studies have indicated that curcumin is a
pharmacologically safe compound, and it has therapeutic potential
in preventing breast cancer metastasis, possibly through
suppression of NF-κB, an important transcription factor. It has
been demonstrated to be involved in regulating apoptosis. The
current results indicated that either curcumin or paclitaxel alone
significantly decreased NF-κB protein expression in the MCF7 cell
line, and combined treatment slightly decreased NF-κB protein
expression in the MDA-MB-231 cell line. However, paclitaxel alone
increased NF-κB protein expression in the MDA-MB-231 cell line,
indicating possible resistance to this drug that is counteracted by
curcumin when they are combined. It has previously been observed
that curcumin suppresses NF-κB activation, and most
chemotherapeutic agents activate genes that mediate proliferation,
which is why it was ascertained whether curcumin would potentiate
the effect of chemotherapy in breast cancer cell lines in the
current study. Curcumin decreased expression of NF-κB in the
current study, supporting the conclusion of other reports that the
mechanism of proliferation and inhibition induced by combined
effect of paclitaxel and curcumin is through NF-κB inhibition
(27).
Previous results indicated that NF-κB is altered by
curcumin (29). Since curcumin
suppresses NF-κB activation and most chemotherapeutic agents
activate genes that mediate cell survival, proliferation, invasion
and metastasis, it was analyzed whether curcumin would potentiate
the effect of chemotherapy in breast cancer cell lines (27). Curcumin decreased expression of
NF-κB, supporting the conclusion of other reports that the
mechanism of growth inhibition induced by combined effect of
paclitaxel and curcumin is through NF-κB inhibition. It was
previously demonstrated that curcumin suppresses the
paclitaxel-induced NF-κB pathway in breast cancer cells by
inhibiting lung metastasis of human breast cancer in nude mice
(27). The current results
indicated that the NF-κB gene and protein expression increased in
the MCF-7 and MDA-MB-231 cells compared with controls.
In the current study, PCNA protein expression was
not altered by curcumin or paclitaxel in the MCF7 cell line, but
was inhibited in MDA-MB-231 by curcumin alone. These results are
similar to those published by Calaf and Hei (28), in which curcumin inhibited PCNA
protein expression in an in vitro breast cancer progression
model. It may be concluded that both curcumin and paclitaxel induce
apoptosis by regulating NF-κB expression in malignant breast cell
lines, since this molecule is a master modulator of several
mechanisms, including apoptosis and cell proliferation.
Acknowledgements
The technical support of Georgina Vargas Marchant,
Guiliana Rojas and Leodán A. Crispin is greatly appreciated.
Funding
This study was supported by a grant from Universidad
de Tarapacá, Convenio de desempeño UTA1117 (GMC).
Availability of data and materials
The datasets used during the present study are
available from the corresponding author upon reasonable
request.
Authors' contributions
GMC, RPC, FC conceived, designed and performed the
study. GMC, RPC, FC wrote, reviewed and edited the manuscript. All
authors read and approved the manuscript and agree to be
accountable for all aspects of the research in ensuring that the
accuracy or integrity of any part of the work are appropriately
investigated and resolved.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Ferlay J, Soerjomataram I, Dikshit R, Eser
S, Mathers C, Rebelo M, Parkin DM, Forman D and Bray F: Cancer
incidence and mortality worldwide: Sources, methods and major
patterns in GLOBOCAN 2012. Int J Cancer. 136:E359–E386. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Jafari SH, Saadatpour Z, Salmaninejad A,
Momeni F, Mokhtari M, Nahand JS, Rahmati M, Mirzaei H and Kianmehr
M: Breast cancer diagnosis: Imaging techniques and biochemical
markers. J Cell Physiol. 233:5200–5213. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Mustacchi G and De Laurentiis M: The role
of taxanes in triple-negative breast cancer: Literature review.
Drug Des Devel Ther. 9:4303–4318. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Bidkar AP, Sanpui P and Ghosh SS:
Efficient induction of apoptosis in cancer cells by
paclitaxel-loaded selenium nanoparticles. Nanomedicine (Lond).
12:2641–2651. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Karimi G, Vahabzadeh M, Lari P, Rashedinia
M and Moshiri M: ‘Silymarin’, a promising pharmacological agent for
treatment of diseases. Iran J Basic Med Sci. 14:308–317.
2011.PubMed/NCBI
|
|
6
|
Razavi-Azarkhiavi K, Iranshahy M, Sahebkar
A, Shirani K and Karimi G: The protective role of phenolic
compounds against doxorubicin-induced cardiotoxicity: A
comprehensive review. Nutr Cancer. 68:892–917. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Stage TB, Bergmann TK and Kroetz DL:
Clinical pharmacokinetics of paclitaxel monotherapy: An updated
literature review. Clin Pharmacokinet. 57:7–19. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zheng X, Wang C, Xing Y, Chen S, Meng T,
You H, Ojima I and Dong Y: SB-T-121205, a next-generation taxane,
enhances apoptosis and inhibits migration/invasion in MCF-7/PTX
cells. Int J Oncol. 50:893–902. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wang Y, Yu J, Cui R, Lin J and Ding X:
Curcumin in treating breast cancer: A review. J Lab Autom.
21:723–731. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Robles-Escajeda E, Das U, Ortega NM, Parra
K, Francia G, Dimmock JR, Varela-Ramirez A and Aguilera RJ: A novel
curcumin-like dienone induces apoptosis in triple-negative breast
cancer cells. Cell Oncol (Dordr). 39:265–277. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ko EY and Moon A: Natural products for
chemoprevention of breast cancer. J Cancer Prev. 20:223–231. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Księżakowska-Łakoma K, Żyła M and
Wilczyński JR: Mitochondrial dysfunction in cancer. Prz
Menopauzalny. 13:136–144. 2014.PubMed/NCBI
|
|
13
|
Cui L, Bu W, Song J, Feng L, Xu T, Liu D,
Ding W, Wang J, Li C, Ma B, et al: Apoptosis induction by
alantolactone in breast cancer MDA-MB-231 cells through reactive
oxygen species-mediated mitochondrion-dependent pathway. Arch Pharm
Res. 41:299–313. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Li J, Dou D, Li P, Luo W, Lv W, Zhang C,
Song X, Yang Y, Zhang Y, Xu Y, et al: PARP-1 serves as a novel
molecular marker for hepatocellular carcinoma in a Southern Chinese
Zhuang population. Tumour Biol. 39:10104283177069142017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lucarini L, Durante M, Lanzi C, Pini A,
Boccalini G, Calosi L, Moroni F, Masini E and Mannaioni G:
HYDAMTIQ, a selective PARP-1 inhibitor, improves bleomycin-induced
lung fibrosis by dampening the TGF-β/SMAD signalling pathway. J
Cell Mol Med. 21:324–335. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Schuhwerk H, Atteya R, Siniuk K and Wang
ZQ: PARPing for balance in the homeostasis of
poly(ADP-ribosyl)ation. Semin Cell Dev Biol. 63:81–91. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Darvishi B, Farahmand L, Eslami-S Z and
Majidzadeh-A K: NF-κB as the main node of resistance to receptor
tyrosine kinase inhibitors in triple-negative breast cancer. Tumour
Biol. 39:10104283177069192017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Baskari S, Govatati S, Madhuri V,
Nallabelli N; K PM, ; Naik S, Poornachandar Balka S, Tamanam RR and
Devi VR: Influence of autocrine growth hormone on NF-κB activation
leading to epithelial-mesenchymal transition of mammary carcinoma.
Tumour Biol. 39:10104283177191212017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Juríková M, Danihel Ľ, Polák Š and Varga
I: Ki67, PCNA, and MCM proteins: Markers of proliferation in the
diagnosis of breast cancer. Acta Histochem. 118:544–552. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wang SC: PCNA: A silent housekeeper or a
potential therapeutic target? Trends Pharmacol Sci. 35:178–186.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lossi L, Cocito C, Alasia S and Merighi A:
Ex vivo imaging of active caspase 3 by a FRET-based molecular probe
demonstrates the cellular dynamics and localization of the protease
in cerebellar granule cells and its regulation by the
apoptosis-inhibiting protein survivin. Mol Neurodegener. 11:342016.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Gallardo M and Calaf GM: Curcumin and
epithelial-mesenchymal transition in breast cancer cells
transformed by low doses of radiation and estrogen. Int J Oncol.
48:2534–2542. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ponce-Cusi R and Calaf GM: Apoptotic
activity of 5-fluorouracil in breast cancer cells transformed by
low doses of ionizing α-particle radiation. Int J Oncol.
48:774–782. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yan G, Graham K and Lanza-Jacoby S:
Curcumin enhances the anticancer effects of trichostatin a in
breast cancer cells. Mol Carcinog. 52:404–411. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chen D, Dai F, Chen Z, Wang S, Cheng X,
Sheng Q, Lin J and Chen W: Dimethoxy curcumin induces apoptosis by
suppressing survivin and inhibits invasion by enhancing E-cadherin
in colon cancer cells. Med Sci Monit. 22:3215–3222. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ganta S and Amiji M: Coadministration of
paclitaxel and curcumin in nanoemulsion formulations to overcome
multidrug resistance in tumor cells. Mol Pharm. 6:928–939. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Quispe-Soto ET and Calaf GM: Effect of
curcumin and paclitaxel on breast carcinogenesis. Int J Oncol.
49:2569–2577. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Calaf GM and Hei TK: Establishment of a
radiation- and estrogen-induced breast cancer model.
Carcinogenesis. 21:769–776. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Calaf GM, Echiburú-Chau C, Wen G, Balajee
AS and Roy D: Effect of curcumin on irradiated and
estrogen-transformed human breast cell lines. Int J Oncol.
40:436–442. 2012.PubMed/NCBI
|
|
30
|
Calaf GM, Echiburú-Chau C, Roy D, Chai Y,
Wen G and Balajee AS: Protective role of curcumin in oxidative
stress of breast cells. Oncol Rep. 26:1029–1035. 2011.PubMed/NCBI
|