Introduction
As the fifth common human malignancy, hepatocellular
carcinoma (HCC) is the third leading cause of cancer-associated
mortality worldwide (1).
Traditional therapeutic strategies, including surgery, are often
ineffective in controlling HCC or prolonged HCC-associated
survival, partially due to late diagnoses (2,3).
Transarterial embolization (TAE), including transarterial
chemoembolization (TACE), is a common therapeutic strategy for
patients with stage B HCC who are unable to undergo surgical
resection (4). During TAE/TACE, an
emulsion of materials that includes chemotherapeutic drugs is
injected into the tumor-feeding arteries, thereby blocking the
blood supply to the tumor, leading to ischemia, hypoxia and
necrosis of the tumor tissues (5).
Studies have indicated that TAE/TACE is effective in controlling
HCC (6). However, tumor recurrence
and poor prognosis frequently occur subsequent to TAE/TACE.
Although significant advances have been achieved in the treatment
of HCC, such as gene therapy and immunotherapy (7–10),
these strategies for patients are currently limited in use due to
the high-cost and time-consuming nature. Therefore, the current
therapies, such as TAE, in combination with other novel
technologies against HCC may be an affordable and feasible
approach.
Precise gene knockout technology for editing the
human genome has been applied to translational medicine, and the
CRISPR/Cas9 (also known as clustered regularly interspaced short
palindromic repeats/CRISPR-associated protein 9) system is one of
the outstanding methods (11).
CRISPR/Cas9 is a bacterial system with endonucleases involved in
adaptive immunity. The system can effectively target intended
genomic loci by a small guide RNA (sgRNA), which is composed of a
target complementary CRISPR RNA (crRNA) and an auxiliary
trans-activating crRNA (tracrRNA) (12). Technologically, through a delivery
vector, such as a lentiviral vector, designed sgRNAs and
CRISPR/Cas9 can be expressed in a certain cell. When the
CRISPR/Cas9 endonuclease targets a specific genomic locus through
base pairing between the crRNA sequence and the target DNA
sequence, a double-stranded DNA break is formed, and a targeted
gene modification is achieved, causing gene disruption, which is
much more efficient than the effect of other nucleases, such as
Zinc finger nucleases (ZFNs) and transcription activator-like
effector nucleases (TALENs) (13).
In addition, Genetic landscape and biomarkers of HCC have a clear
delineation (14). Therefore, the
CRISPR/Cas9 system, as a powerful gene-editing tool, is promising
in the preclinical and clinical treatment of HCC, which is mainly
caused by genetic mutations and an abnormally high expression of
key oncogenic proteins.
Hypoxia inducible factor-1α (HIF-1α), a
hypoxia-induced oxygen-dependent transcription factor, play a vital
role in the aggressiveness, angiogenesis and chemotherapy
resistance of HCC, as well as in the maintenance of liver cancer
stem cells (15). Particularly,
HIF-1α activates the hypoxic pathways, leading to the
overexpression of vascular endothelial growth factor (VEGF),
triggering epithelial mesenchymal transition, tumor invasiveness
and metastasis by promoting the expression levels of matrix
metalloproteinase 2 (MMP2) and MMP9 (16,17).
Our previous study demonstrated that HIF-1α was upregulated in HCC,
and was associated with hepatic capsular invasiveness and portal
vein metastasis (18).
Additionally, the expression of multi-drug resistance protein (MDR)
is also under the control of HIF-1α, and its protein product
P-glycoprotein (P-gp) is capable of transporting chemotherapeutic
drugs out of cells. Tumor cells with an excessive expression of
P-gp are generally resistant to chemotherapeutic drugs (19). Notably, it has been reported that
TAE/TACE induced liver hypoxia, and that the expression levels of
HIF-1α in the serum and HCC tissues were significantly higher in
patients who underwent TACE compared with those in the control
group (20). High recurrence rates
and metastases of HCC following TACE treatment are also involved in
the HIF-1α-associated drug-resistance and hypoxia-tolerance
(21). Thus, the inhibition of
HIF-1α in combination with TAE may be a promising therapeutic
approach against HCC.
In the present study, a lentivirus-mediated
CRISPR/Cas9 system targeting the human HIF-1α gene was used to
analyze the role of HIF-1α in the human liver cancer cell line
SMMC-7721. The lentivirus-mediated HIF-1α disruption suppressed
cell proliferation, migration and invasiveness, and induced
apoptosis under CoCl2-induced hypoxic conditions. HIF-1α
disruption also significantly inhibited SMMC-7721 ×enograft tumor
growth in the liver and prolonged the survival of HCC-bearing mice
treated with hepatic artery ligation (HAL). These findings
suggested that CRISPR/Cas9-based HIF-1α disruption improved the
antitumor efficacy of HAL and further highlighted the possibility
that HIF-1α may be an ideal knockout target for the treatment of
HCC in combination with TAE.
Materials and methods
HCC patient data and tissue
specimens
A total of 20 matched pairs of HCC and normal
adjacent liver tissues were acquired during surgery between May
2014 and April 2016 at the Shenzhen People's Hospital (Shenzhen,
China). Patients met the following criteria according to our
previous work: i) male/female patient ratio was 1:1, patients
ranged in age from 35 to 75 years; ii) HCC patients were newly
diagnosed with liver function tests of Child-Pugh grade A; iii)
without history of anticancer therapy; iv) without any distant
metastasis; v) without other types of malignant tumors, autoimmune,
liver disease or serious heart, lung, kidney, or blood disease; and
vi) seronegative for hepatitis B surface antigen and HCV. The
samples were either snap-frozen or stored in liquid nitrogen for
protein extraction or fixed in formalin and embedded in paraffin
for immunohistochemistry analysis.
Construction of the
pLenti-CAS9-sgRNA719/720/721-egfp vectors
sgRNAs were designed using the Optimized CRISPR
Design tool (http://crispr.mit.edu/) as follows:
sgRNA719, 5′-CCTCACACGCAAATAGCTGA-3′; sgRNA720,
5′-TACTCATCCATGTGACCATG-3′; and sgRNA721,
5′-GTTATGGTTCTCACAGATGA-3′. LV-Ctrl was constructed as control
lentiviruses that did not have any sgRNAs used for targeting the
HIF-1α gene. The cDNAs encoding the sgRNAs for 3 gene
knockout sites in the first exon of the HIF-1α gene were
synthesized and purified (Invitrogen; Thermo Fisher Scientific,
Inc., Waltham, MA, USA). Different programmed lentiviral plasmids
based on the pLenti-CAS9-sgRNA-egfp (GeneChem Co., Ltd., Shanghai,
China) were constructed as previously described (22).
Packaging and purification of the
modified CRISPR/Cas9 lentiviruses
The CRISPR/Cas9 lentivirus production was performed
following previously described protocols (22). Briefly, 12 145-mm Petri dishes were
prepared and 1×107 HEK293T cells [American Type Culture
Collection (ATCC) Manassas, VA, USA] were seeded in each dish with
Dulbecco's modified Eagle's medium (Gibco; Thermo Fisher
Scientific, Inc.) containing 10% fetal bovine serum (FBS; Gibco;
Thermo Fisher Scientific, Inc.). pLenti-CAS9-sgRNA-egfp (20
µg), psPAX2 (15 µg; Addgene, Inc., Cambridge, MA, USA), pMD2.G
plasmid (10 µg; Addgene, Inc., Cambridge, MA, USA) and linear
PEImax (Polysciences, Inc., Warrington, PA, USA) were mixed and
added to the HEK293T cells in each dish. After 6 h of incubation in
a CO2 incubator at 37°C, the medium was replaced with
complete medium. After 48 h, the cell supernatants were filtered
through a 0.45-µm filter (EMD Millipore, Billerica, MA, USA).
Subsequently, supernatants were ultracentrifuged (Beckman Coulter,
Inc., Brea, CA, USA) at 20,000 × g for 2 h at 4°C, and the
lentiviruses were suspended in PBS and stored at −80°C.
Lentivirus infection in SMMC-7721
cells and the T7 endonuclease 1 assay (T7E1) of
HIF-1α-knockout
The titer of the concentrated lentiviruses was
determined by a quantitative polymerase chain reaction (qPCR)
method as described previously with 293T cells (23). Subsequently, SMMC-7721 cells (ATCC)
were cultured in Dulbecco's modified Eagle's medium (Gibco; Thermo
Fisher Scientific, Inc.) with 10% FBS and 1%
penicillin-streptomycin (Gibco; Thermo Fisher Scientific, Inc.) in
a CO2 incubator at 37°C. SMMC-7721 cells were infected
with the lentivirus at a multiplicity of infection (MOI) of 2.5. To
check the total percentage of GFP-positive cells, single-cell
suspensions were prepared in PBS plus 2% FBS from trypsinized cells
at 72-h after infection. After resuspension, cells were subjected
to fluorescence activated cell sorting (FACS) analysis with a BD
Accuri C6 flow cytometer (BD Biosciences, San Jose, CA, USA) as
described (22). Next, SMMC-7721
cell genomic DNA was extracted using a DNeasy Blood & Tissue
kit (Qiagen, Hilden, Germany). A mismatch-sensitive T7E1 Assay kit
(New England BioLabs, Inc., Ipswich, MA, USA) was then used to
confirm CRISPR/Cas9 cleavage and targeted sequence disruption,
according to the manufacturer's protocol. Subsequently, the PCR
fragments containing each knockout gene locus were cloned with
primers in different group using a Takara PCR amplification kit
(cat. no. DR011; Takara Bio, Inc., Otsu, Japan) according to the
manufacturer's protocols. The following primers were used: i)
LV-H719 + 7721 cells group: 5′-TCTAATCCTTCTGTGATAAGCAG-3′ (forward)
and 5′-CAAAATCAAAACATTGCGACCAC-3′ (reverse); ii) LV-H720 + 7721
cells group: 5′-ACATGAAAGCACAGAAATTGC-3′ (forward) and
5′-TGCCTTGGGTAAGTACAATAGC-3′ (reverse); and iii) LV-H721 + 7721
cells group: H721, 5′-TCTTCTTGTGCCCTTTTTAGGTG-3′ (forward) and
5′-CTTACCATTTCTGTGTGTAAGC-3′ (reverse). The cloned DNA sequences
were then inserted into a plasmid using pMD19-T (Takara Bio, Inc.,
Otsu, Japan) for DNA sequencing (Invitrogen; Thermo Fisher
Scientific, Inc.) and comparison. The SMMC-7721 cell line where
HIF-1α has been successfully knockout after LV-H721
infection at MOI=2.5 was established and was named as
7721-HIF-1α-KO cells for further testing.
Reverse transcription-qPCR analysis of
the mRNA expression levels of vegf and mdr1 genes
The total cell RNA was extracted using the TRIzol
reagent (Invitrogen; Thermo Fisher Scientific, Inc.) according to
the manufacturer's protocol. The concentration and the quality of
RNAs were measured with a NanoDrop 8000 (Thermo Fisher Scientific,
Inc.). A PrimeScript® II First Strand cDNA synthesis kit
(cat. no. H6210B; Takara Bio, Inc.) was used to synthesize the
cDNA. Subsequently, the gene mRNA expression levels were detected
by qPCR using Thunderbird SYBR-Green qPCR Mix (cat. no. QPS-201;
Toyobo Life Science, Osaka, Japan) according to the manufacturer's
protocol. Primers used in this experiment were as follows:
vegf, 5′-TGCTCTACCTCCACCATGCCA-3′ (forward) and
5′-GAAGATGTCCACCAGGGTCTCG-3′ (reverse); mdr1,
5′-TGATGCTGCTCAAGTTAAAGGG-3′ (forward) and
5′-TTGCCAACCATAGATGAAGGATAT-3′ (reverse); β-actin,
5′-GTCCACCGCAAATGCTTCTA-3′ (forward) and 5′-TGCTGTCACCTTCACCGTTC-3′
(reverse); and gfp, 5′-TGCTTCAGCCGCTACCC-3′ (forward) and
5′-AGTTCACCTTGATGCCGTTC-3′ (reverse).
Protein sample preparation and western
blotting
Protein samples from patient tissues and SMMC-7721
cells were prepared using radio-immunoprecipitation assay lysis
buffer (RIPA; Beyotime Institute of Biotechnology, China), and the
protein concentration was detected by the BCA protein quantitation
kit (Bio-Rad Laboratories, Inc., Hercules, CA, USA). Western blot
analysis was then performed based on protocols reported in a
previous study (22). The following
primary and secondary antibodies were used: Anti-VEGF (cat. no.
19003-1-AP; 1:2,000, ProteinTech, Inc., Chicago, IL, USA),
anti-MDR1 (cat. no. PA5-28810; 1:2,000, Thermo Fisher Scientific,
Inc.), anti-HIF-1α (cat. no. 20960-1-AP; 1:2,000, ProteinTech,
Inc.), anti-β-actin (cat. no. MA1-140; 1:2,000, Thermo Fisher
Scientific, Inc.), horseradish peroxidase (HRP)-conjugated
anti-rabbit IgG (cat. no. 7074; 1:2,000, Cell Signaling Technology,
Inc., Danvers, MA, USA) and HRP-conjugated anti-mouse IgG (cat. no.
7076; 1:2,000; Cell Signaling Technology, Inc.) antibodies.
MTT assay, apoptosis and cell cycle
detection
SMMC-7721 cells were cultured in 96-well
(104 cells per well) or 6-well (3×105 cells
per well) plates. A total of 150 µM cobalt chloride
(CoCl2; Sigma-Aldrich; Merck KGaA, Darmstadt, Germany),
a well-known hypoxia mimetic (24,25),
was used to mimic tumor hypoxic conditions. In order to examine the
cell proliferation, at 8 h after the lentivirus infection, 20 µl
MTT (5 mg/ml) was added to each sample and incubated for 3 h. The
formazan crystals were dissolved in dimethyl sulfoxide (150
ml/well), and the absorbance of each sample at 490 nm was
measured.
For cell apoptosis analysis, a PE-Annexin V
Apoptosis Detection kit (cat. no. 559763; BD Biosciences) was used.
Briefly, 5 µl of Annexin V-PE and 5 µl of 7-AAD were added in 100
µl cell suspension with 1× binding buffer plus 5% FBS, and
incubated for 15 min in the dark. Finally, cells were washed twice
and diluted with buffer and immediately subjected to FACS analysis
(22). For the detection of cell
cycle progression, PI-RNase solution (cat. no. 550825; BD
Biosciences) was used, according to the manufacturer's protocol.
Flow cytometry was then performed using a BD FACS Arial flow
cytometer (BD Biosciences), and the results were analyzed with
FlowJo software (Tree Star, Inc., Ashland, OR, USA).
Transwell and scratch wound assay
A Transwell system with a 6.5-mm diameter and an 8.0
µm pore polycarbonate filter membrane (Corning Inc., Corning, NY,
USA) was used, and Matrigel (cat. no. 356234; Corning Inc.) was
used to form a thin gel layer on the wells. At 12 h after
infection, the SMMC-7721 cells (1×105 per well) were
added to the upper chamber. In the lower chambers, 20% FBS was
added as a chemoattractant. CoCl2 (150 µM) was added to
simulate hypoxia in both upper and lower chambers. After 24 h, the
cells that had penetrated to the lower surface of the filter were
stained with crystal violet and counted using a Nikon microscope
(Nikon Corporation, Tokyo, Japan).
A scratch wound assay was also performed. Briefly,
the SMMC-7721 cells were seeded into 6-well plates
(3×105 per well). Following infection with lentivirus
after 12 h, a linear wound was created in the monolayer with a
sterile 100-µl plastic pipette tip, and cells were then gently
washed with PBS immediately. Cells were cultured under the
CoCl2-simulated hypoxic conditions and then observed
under the microscope.
Establishment of SMMC-7721-Fluc
cells
The overlapping gene mFluc2, which included
the mCherry fluorophore and Fluc2 gene (the gene that
expresses the firefly luciferase protein), was cloned from the
pmCherry-C1 (Clontech Laboratories, Inc., Mountainview, CA, USA)
and pGL4.17 plasmids (Promega Corp., Madison, WI, USA) using a
Takara PCR amplification kit. The PCR cycling conditions were set
as follows: 1 min at 95°C for denaturation, 45 cycles of 95°C for
30 sec, and 55°C for 30 sec, then 72°C for 1 min. Lastly 1 h s at
4°C. Next, the pWPXLd-mFluc2 plasmid was constructed, and the
LV-mFluc lentivirus was packaged according to previous methods
(22). The SMMC-7721 cells were
infected with LV-mFluc with an MOI of 2.5, and the mCherry-positive
SMMC-7721 cells were purified by flow cytometry (BD FACS Arial
device) and cloned. Subsequently, a dual-luciferase reporter assay
in the intended SMMC-7721-Fluc cells was conducted using the
Dual-Luciferase® Reporter Assay system according to the
manufacturer's protocol (cat. no. E1910; Promega Corp. Madison, CA,
USA). The primers used in this experiment are as follows: Clone for
mCherry gene: 5′-GGGGATCCATGGTGAGCAAGGGCGAGGAGGATA-3′
(forward) and
5′-TCTTTATGTTTTTGGCGTCTTCCATCTTGTACAGCTCGTCCATGCCGCCG-3′ (reverse);
and clone for Fluc gene,
5′-CGGCGGCATGGACGAGCTGTACAAGATGGAAGACGCCAAAAACATAAAGA-3′ (forward)
and 5′-ACGGAATTCTCACTCGAGCAATTTGGACTTTCCG-3′ (reverse).
Animals and CRISPR/Cas9 treatment in
vivo
Seventy male BALB/c nu/nu mice (4–6 weeks old, 16–20
g) were purchased from the Shanghai SLAC Laboratory Animal Co.,
Ltd. [license no. SCXK (HU) 2007-0003; Shanghai, China]. Mice were
housed in specific pathogen-free (SPF) conditions, with a 12-hours
light cycle and food and water at ad libitum. In order to
establish a subcutaneous HCC model, 4×106 SMMC-7721
cells were injected in the right flank of the mouse subcutaneously
(n=4/group). After 9 days, 5×107 LV-H721, which was a
lentivirus-mediated CRISPR/Cas system with small guide RNA-721,
were injected into the tumor tissues. Tumor tissues injected with
PBS or LV-Ctrl (a lentivirus without CRISPR/Cas system targeting
HIF-1α) were used as control groups. The tumors were
harvested 3 days post LV-infection for immunohistochemical
examination.
The procedure for the establishment of the
orthotopic liver tumor model was conducted according to a
previously described method (26).
Briefly, pentobarbital was dissolved in saline to obtain a 10 mg/ml
solution. Mice were anesthetized with pentobarbital (50 mg/kg)
through intraperitoneal injection and placed in a supine position.
A small transverse incision was made below the sternum to expose
the liver. A total of 5×106 control lentivirus-infected
SMMC-7721-Fluc cells (7721) or LV-H721-infected SMMC-7721-Fluc
cells at 48 h after infection were suspended in 50 µl PBS and
slowly injected into each mouse liver. At 5 days after
implantation, mice were subdivided into four groups as follows: i)
7721-induced HCC + sham surgery (Ctrl; n=8); ii) 7721-induced HCC +
HAL surgery (n=8); iii) control lentivirus-infected 7721-induced
HCC + HAL (HAL + LV-Ctrl; n=8); and iv) LV-H721-infected
7721-induced HCC + HAL (HAL + LV-H721; n=8). HAL was performed in
the mice by ligating the main branch of the hepatic artery. The
tumor volume was measured by bioluminescence imaging with 200 µl
D-luciferin potassium salt solutions (15 mg/ml; Sigma-Aldrich;
Merck KGaA) after HAL during 2 weeks. After 2 weeks, the mice
(n=5/group) were sacrificed and mouse tumor samples were collected
for immunohistochemistry analysis. Furthermore, an extended
observation of tumor-bearing mice with four different treatments
was carried out again, and the mouse overall survivals were
recorded.
Immunohistochemical staining for
HIF-1α and CD31 expression in tissues
Human or SMMC-7721-induced mouse tumor tissues were
fixed with 4% neutral paraformaldehyde. Next, the paraffin-embedded
sections (human and mouse tumor tissues) were incubated with the
anti-HIF-1α antibody (cat. no. 20960-I-AP; 1:100; ProteinTech,
Inc., Rosemont, IL, USA) overnight at 4°C, followed by incubation
with 50–100 µl 1× the secondary antibody solution for 1-h (cat. no.
KIHC-5, 1:1, ProteinTech, Inc.). The signals were detected by
staining the sections with 3,3′-diaminobenzidine (DAB; ProteinTech,
Inc.) and hematoxylin. Furthermore, to monitor the microvascular
density (MVD), the mouse tumor sections were stained with an
anti-CD31 monoclonal antibody overnight at 4°C (cat. no.
14-0319-80; 1:300, Thermo Fisher Scientific, Inc.), and the rest of
the test was carried out as previously described.
TUNEL staining assay for cell
apoptosis in vivo
The DeadEnd™ Colorimetric TUNEL System (cat. no.
G7130, Promega Corp.) was used for the TUNEL staining assay.
Briefly, the tumor slides were incubated with terminal
deoxynucleotide transferase (TdT) and a biotinylated nucleotide
mixture at 37°C for 30 min. Subsequently, the endogenous
peroxidases was blocked by immersing the slides in 0.3% hydrogen
peroxide in PBS for 3–5 min at room temperature. The slides were
incubated with streptavidin-HRP and visualized with DAB. Negative
controls were set up by substituting distilled water for TdT in the
working solution. The results are presented as the ratio of the
TUNEL-positive cells to the total number of cells.
Statistics analysis
The statistical analysis was performed with SPSS
version 17.0 software (SPSS, Inc., Chicago, IL, USA). All the data
were analyzed using one-way analysis of variance followed by least
significant difference post-hoc analysis, or using an unpaired
two-tailed Student's t-test (as appropriate). The survival curves
were obtained by Kaplan-Meier analysis, and the prognostic
significance was analyzed by the log-rank test. Statistically
significant differences were denoted by P<0.05. All data were
acquired from four repeated experiments.
Results
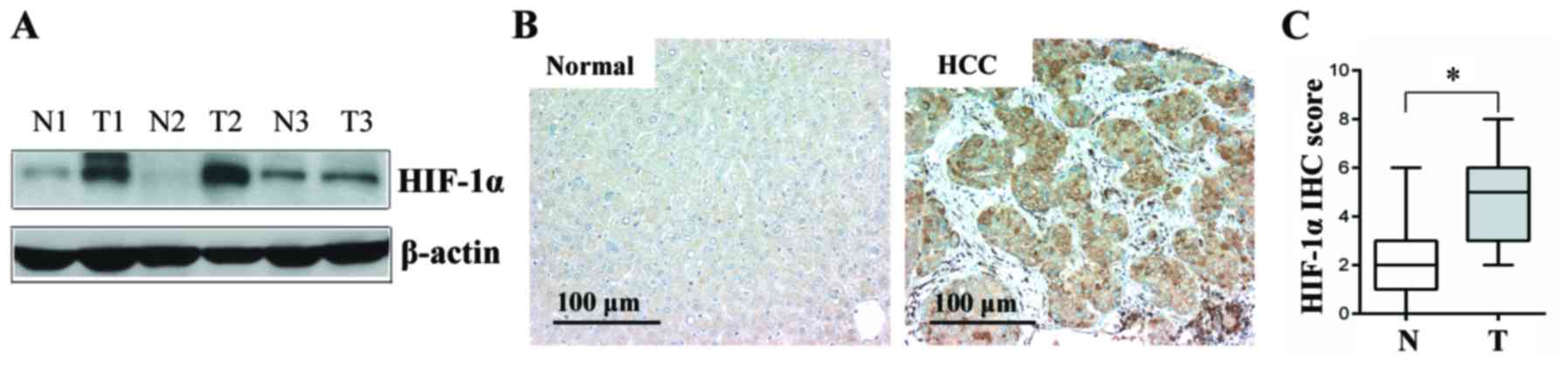
HIF-1α is overexpressed in human HCC
tissues and in SMMC-7721-induced HCC xenograft tissues in mice
The expression levels of HIF-1α protein were first
analyzed in 20 paired HCC and normal adjacent liver tissues by
western blot and immunohistochemical analyses. As shown in Fig. 1A and B, the HIF-1α protein levels in
the HCC tissues were higher in comparison with those in the normal
adjacent liver tissues, and the difference was found to be
statistically significant (P<0.05; Fig. 1C).
SMMC-7721 cells, a human liver cancer cell line,
were also used to establish an HCC model in BALB/c nu/nu mice. The
results revealed that the implanted HCC cells had a significantly
higher expression of HIF-1α protein as compared with the control
groups (Fig. 2A). Subsequently, the
SMMC-7721-induced HCC model was used in further
CRISPR/Cas9-mediated HIF-1α-knockout experiments.
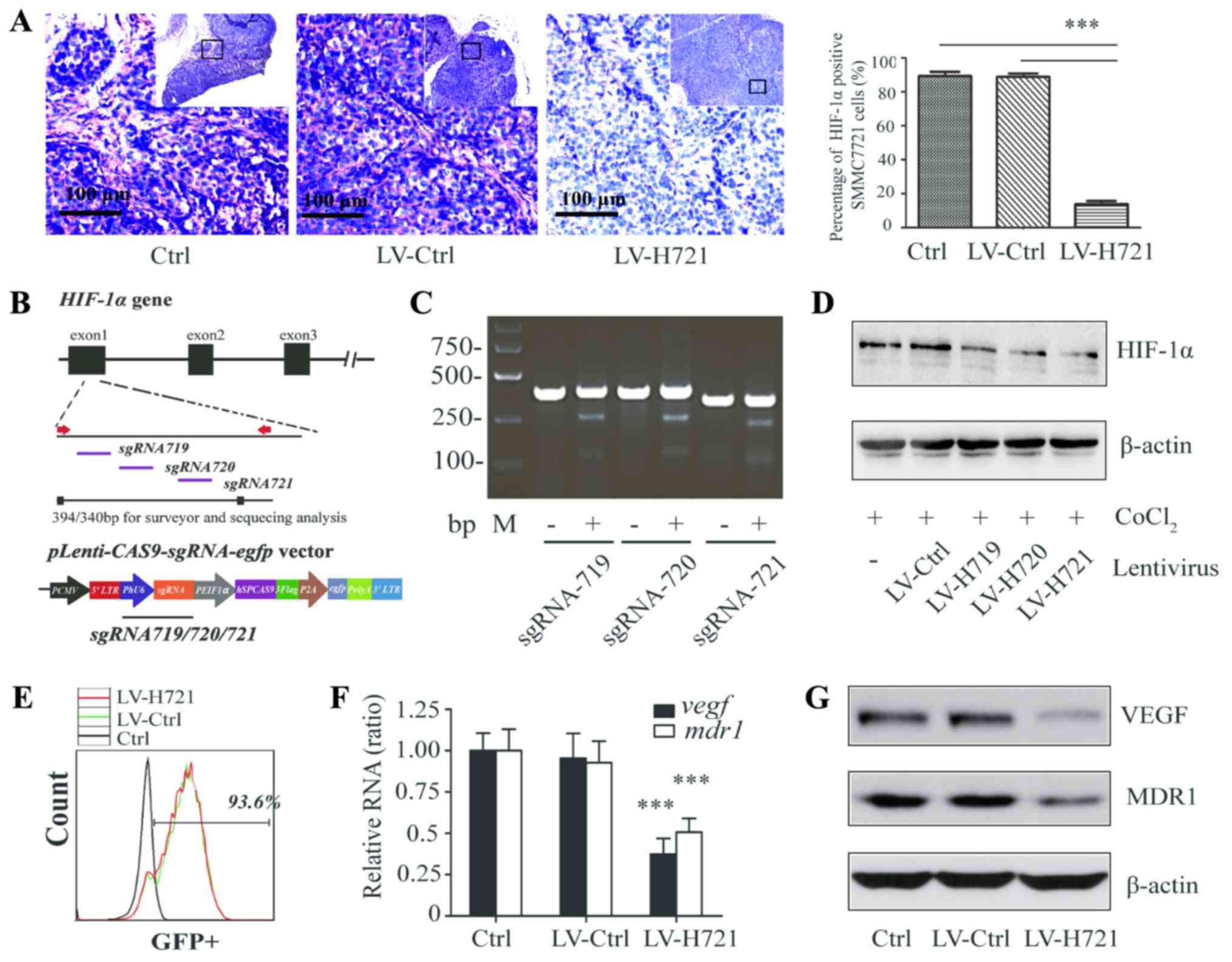 | Figure 2.Generation of the CRISPR/Cas9-based
lentivirus and HIF-1α knockout in SMMC-7721 cells and in the
SMMC-7721-induced tumor tissues of mice. (A) Immunohistochemical
analysis for the detection of HIF-1α expression in hepatocellular
carcinoma tissues following infection with LV-Ctrl or LV-H721 in
the subcutaneous animal model (bars, 100 µm). (B) Diagram
illustration of the lentiviral vector
(Lenti-CAS9-sgRNA-egfp). Three sgRNAs (sgRNA719, sgRNA720
and sgRNA721) targeting HIF-1α were designed, and
Lenti-CAS9-sgRNA719/720/721 plasmids were
constructed. (C) Gel electrophoresis and DNA analysis of the HIF-1α
genomic frame shift mutation conducted after the T7E1 endonuclease
assay in SMMC-7721 cells infected with LV-Ctrl or LV-H719/720/721.
(D) Western blot analysis of HIF-1α expression in the different
lentivirus-infected SMMC-7721 cells with CoCl2-simulated
hypoxia. (E) GFP-positive cells analyzed by flow cytometry
following infection with LV-Ctrl and LV-H721. (F) Relative mRNA and
(G) protein expression levels of VEGF and MDR1 in different cell
groups were examined by reverse transcription-quantitative
polymerase chain reaction and western blot analysis. β-actin served
as an internal control. Data are a representation of four repeated
experiments, and histograms represent the mean ± standard
deviation. ***P<0.001. sgRNA, small guide RNA; Ctrl, control;
LV, lentivirus; HIF-1α, hypoxia inducible factor-1α; VEGF, vascular
endothelial growth factor; MDR1, multi-drug resistance protein
1. |
HIF-1α-knockout via CRISPR/Cas9
suppressed HIF-1α expression
To identify sgRNAs specifically targeting the gene
loci of HIF-1α, three sgRNAs were designed to target the
first exon of HIF-1α according to guidelines of the
Optimized CRISPR Design online tool (Fig. 2B). The
pLenti-CAS9-sgRNA719/720/721-egfp plasmids, based on the
pLenti-CAS9 vector, were established (Fig. 2B). The CRISPR/Cas9 lentiviruses with
different sgRNAs were labeled as LV-H719, LV-H720 and LV-H721.
Subsequently, SMMC-7721 cells were infected with the lentiviruses
at an MOI of 2.5, and the gene disruption efficacies were assessed,
followed by the T7E1 endonuclease assay and DNA sequencing. The
results demonstrated that all three sgRNAs led to DNA frameshift
mutations and had high gene disruption efficiencies (Fig. 2C). Furthermore, the results of the
western blot analysis revealed that LV-H721 infection significantly
reduced the expression of HIF1-α with the maximum efficiency
(Fig. 2D). The percentage of total
GFP-positive cells in LV-H721-infected SMMC-7721 cells was also
measured to assess the lentiviral transfection efficiency, and was
found to be 93.6% (Fig. 2E).
LV-Ctrl (a lentivirus without CRISPR/Cas system targeting
HIF-1) could also express GFP in the infected SMMC-7721
cells and had equal lentiviral transfection. It was observed that
HIF-1α knockout caused by LV-H721 transfection in the cells
was observed to significantly downregulate the mRNA levels of
vegf and mdr1 as compared with those in the control
groups (all P<0.001; Fig. 2F).
As shown in Fig. 2G, the expression
levels of the VEGF and MDR1 protein, which has been reported to be
under the control of HIF-1α (17),
were also inhibited in the present study. Thus, LV-H721 was used as
a CRISPR/Cas9-mediated vector for further experiments.
The efficiency of LV-H721-mediated HIF-1α
knockout was also examined in the mice. The SMMC-7721 ×enograft HCC
tissues were harvested 3 days post LV infection.
Immunohistochemical examination revealed that HIF-1α was highly
expressed in the control and LV-Ctrl-treated HCC tissues.
Furthermore, the expression of HIF-1α was significantly reduced in
the LV-H721-treated tissues when compared with those in the control
groups (both P<0.001; Fig. 2A).
Collectively, these data indicated that the lentivirus-mediated
CRISPR/Cas9 efficiently disrupted the expression levels of
HIF-1α gene and its targets, including VEGF and MDR1, in
liver cancer cells and xenograft tumor tissues.
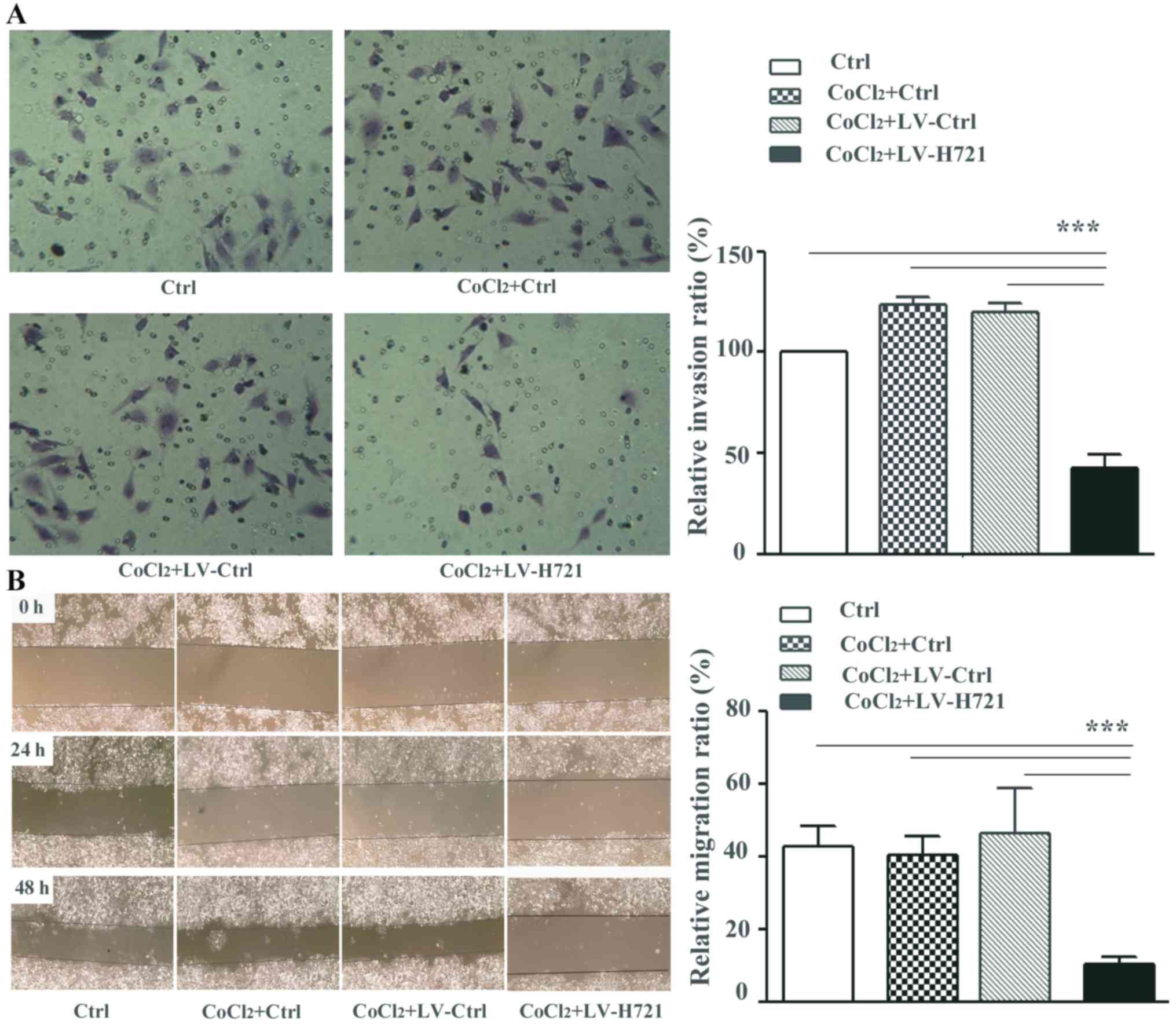
HIF-1α knockout impairs cell invasion
and migration
First, the effect of the CRISPR/Cas9 based
HIF-1α knockout on migration and invasion was determined in
the SMMC-7721 cells (7721-HIF-1α-KO). The SMMC-7721 cells
were infected with the lentiviruses, and CoCl2, a
well-known hypoxia mimetic, was used to simulate the hypoxic
conditions in vitro. In the transwell invasion assay, the
number of invasive 7721-HIF-1α-KO cells (namely the
CoCl2 + LV-H721 group) that penetrated the polycarbonate
filter was significantly reduced, and this difference was
statistically significant (both P<0.001; Fig. 3A). Meanwhile, 7721-HIF-1α-KO
cells exhibited further reduced relative migration rates at 24 and
48 h post LV infection compared with the control groups in the
scratch-wound assay, and the difference was statistically
significant (all P<0.001; Fig.
3B). These results suggested that the disruption of
HIF-1α with the CRISPR/Cas9 system, particularly under the
hypoxic microenvironment, inhibited liver cancer cell migration and
invasion.
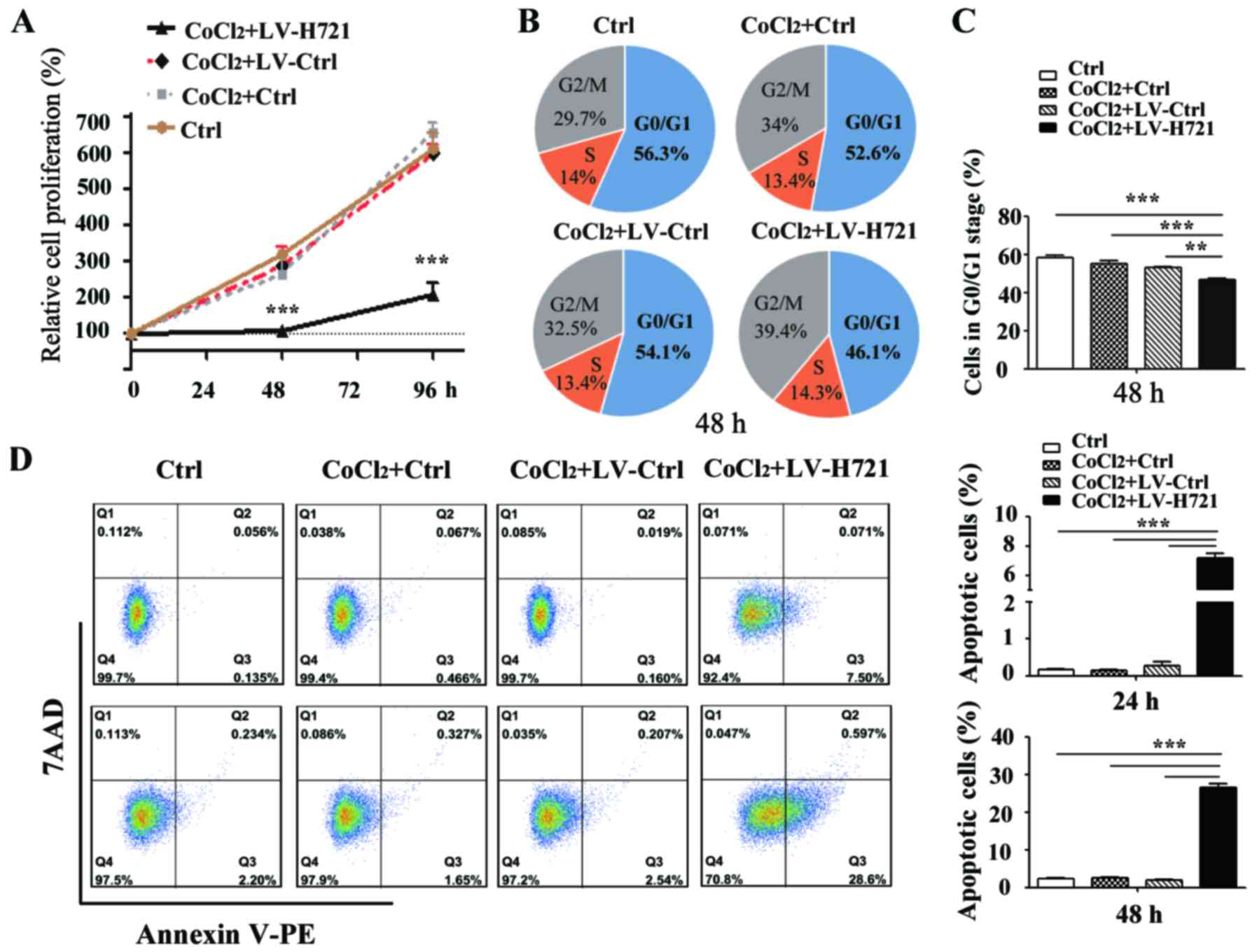
HIF-1α disruption suppresses cell
proliferation and induces cell apoptosis
Hypoxic conditions are reported in the majority of
advanced HCC tissues, in particular in patients who undergo TAE.
Therefore, the growth characteristics of the 7721-HIF-1α-KO
cells under the hypoxic microenvironment were analyzed using an MTT
assay. The optical density, an indicator of the number of viable
cells, was monitored subsequent to infection, and a growth curve
was plotted (Fig. 4A). The results
demonstrated that cell proliferation was profoundly suppressed at
48 and 96 h post infection between the LV-H721-treated group and
the control groups (all P<0.001).
Subsequently, the cell cycle progression under
hypoxic conditions was assessed at 48 h post infection in
vitro. SMMC-7721 cells infected with LV-H721 exhibited marked
changes in their cell cycle progression, and there was an ~10%
decrease in the number of cells in the G0/G1-phase, as well as a
concomitant increase in the G2/M-phase (Fig. 4B). However, the S-phase ratio,
serving as the marker of cell division, was almost consistent.
These data suggested that HIF-1α knockout did not induce
evident cell cycle arrest (Fig. 4B and
C). Furthermore, HIF-1α knockout led to a high
percentage of early cell apoptosis under hypoxic conditions, and
the rates of cell apoptosis were 7.5% at 24 h and 28.6% at 48 h
post infection, with a statistically significant difference
observed (P<0.001; Fig. 4D).
These results suggested that disruption of HIF-1α by the
CRISPR/Cas9 system inhibited cell proliferation and induced cell
apoptosis in the SMMC-7721 cells.
CRISPR/Cas9-mediated HIF-1α disruption
with HAL inhibits liver tumor growth
The SMMC-772-Fluc cells were established and cloned
(data not shown). On days 5, 10, 15 and 20 after implantation of
these cells into the mice, the tumor volume was measured by
bioluminescence imaging (Fig. 5A).
On day 20, the representative features of the tumors in mice with
live bioluminescence imaging are shown in Fig. 5B and tumor outlines in mouse livers
are shown in Fig. 5C. The results
demonstrated that HAL inhibited the progression of HCC when
compared with that observed in the control group (P<0.05;
Fig. 5A). Notably, the combination
of HAL and LV-H721 exhibited a more significant anti-tumor effect
on day 20 as compared with that in the control, HAL and HAL +
LV-Ctrl groups (P<0.001, P<0.05 and P<0.05, respectively;
Fig. 5A). Furthermore, the overall
median survival results revealed that the combination of HAL +
LV-H721 significantly prolonged the survival of tumor-bearing mice
from ~40 to 60 days, as compared with the other groups (P<0.05;
Fig. 5D), indicating that
HIF-1α disruption enhanced the efficacy of HAL in the
treatment of HCC.
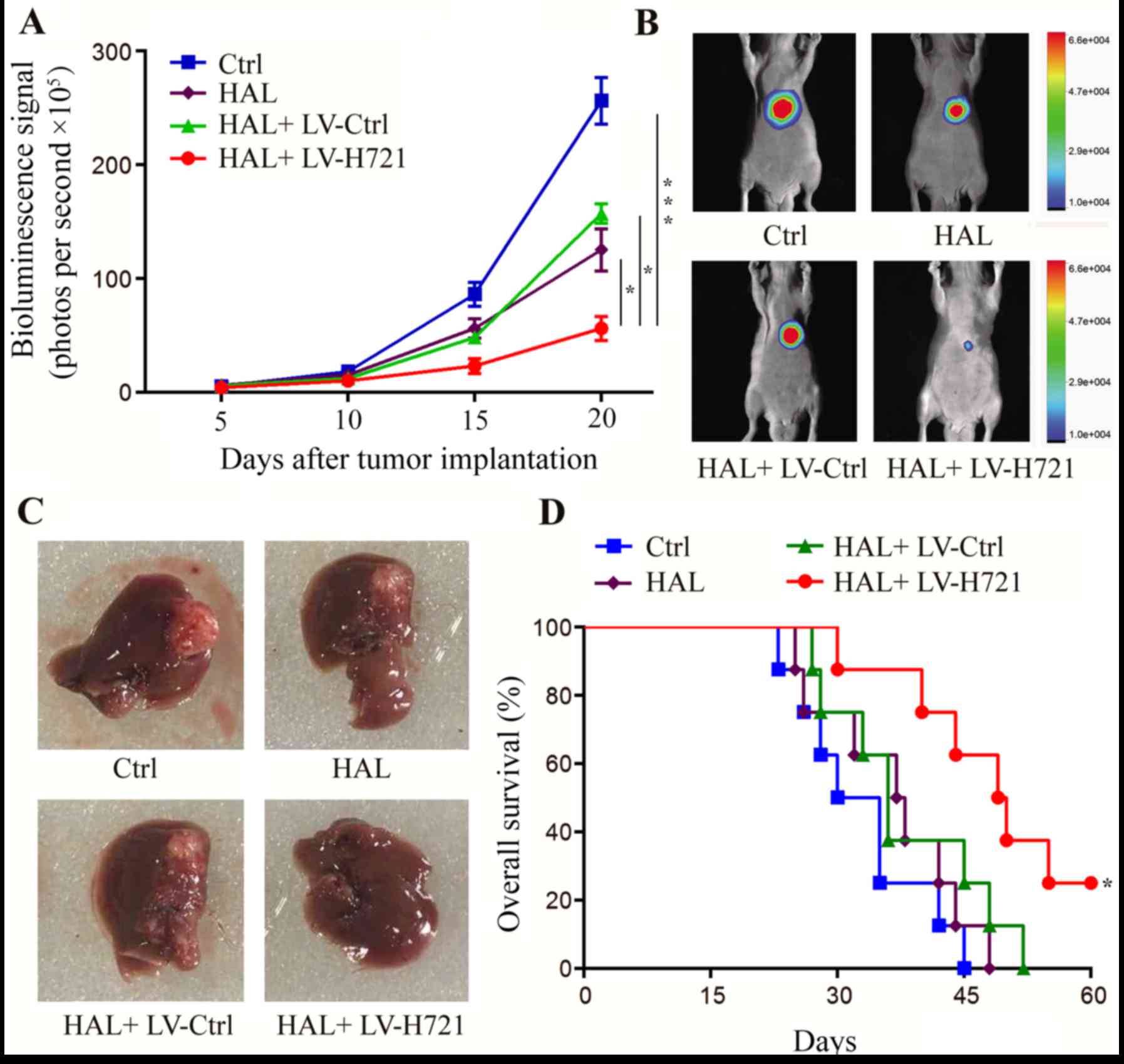 | Figure 5.Effect of hypoxia inducible factor-1α
disruption on tumor growth and progression in the mouse orthotopic
model. At 5 days after tumor cell implantation, the animals were
randomly divided into four groups. (A) Longitudinal monitoring was
plotted by the detection of the mean bioluminescence signals on
days 5, 10, 15 and 20 after cell implantation (error bars represent
the standard error of the mean). (B) Live imaging of the
representative tumors on day 20. (C) Tumors of mice sacrificed on
day 20, indicating that the tumor volume was evidently decreased in
the HAL + LV-H721 group. (D) Animal median survival in each group
plotted by the log-rank test. Statistically significant differences
were denoted between HAL + LV-H721 group and the other three groups
correspondingly (Ctrl, HAL, and HAL + LV-Ctrl groups, all
*P<0.05). Data are representative of four independent
experiments. *P<0.05 and ***P<0.001. HAL, hepatic artery
ligation; Ctrl, control; LV, lentivirus. |
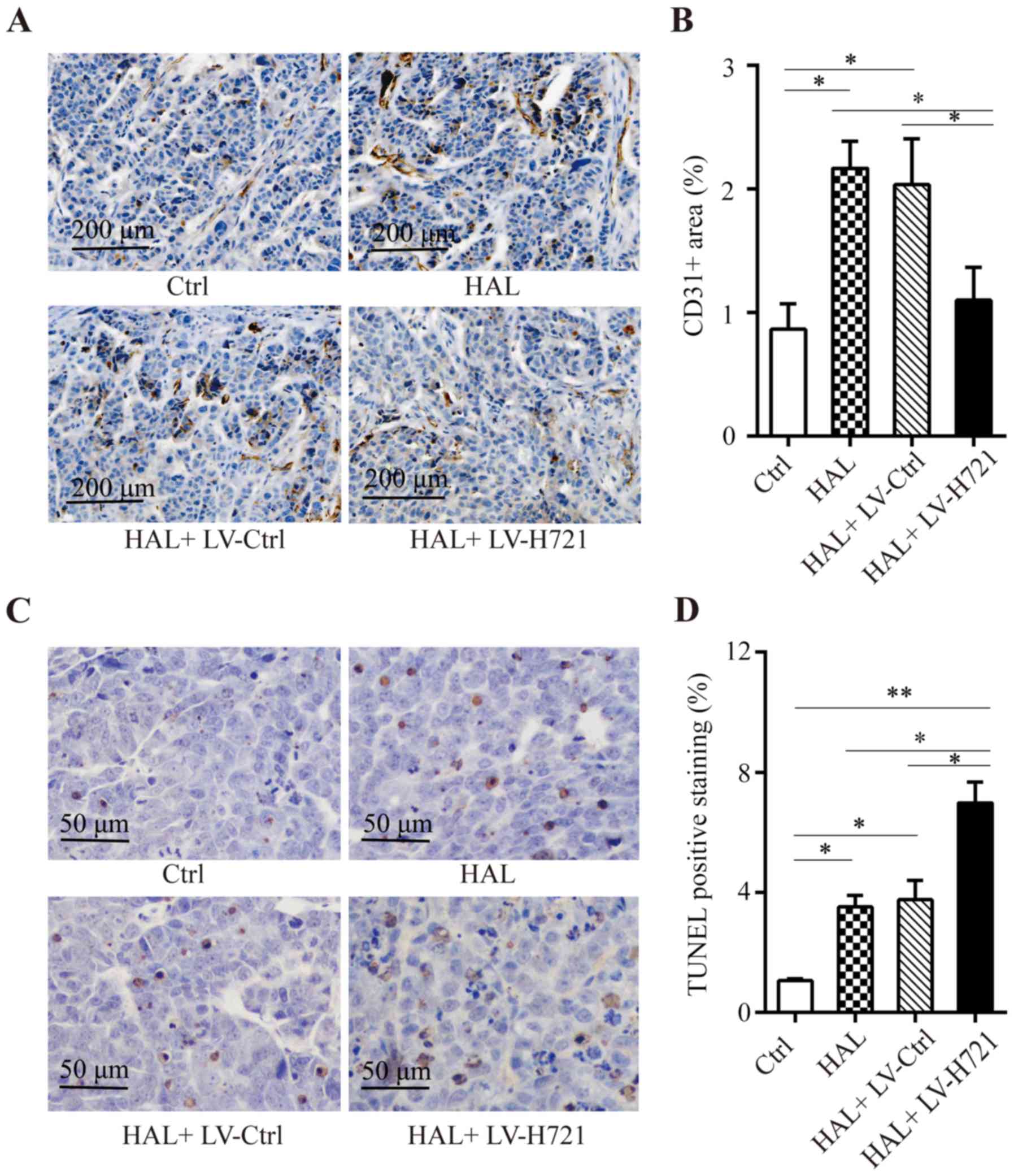
Combination of the
CRISPR/Cas9-mediated HIF-1α disruption with HAL inhibits
angiogenesis and induces apoptosis in tumor tissues
The expression of CD31 was used as a marker of
vascular endothelial cells, or tumor microvascular density (MVD).
From immunohistochemical analysis of CD31 protein expression
(Fig. 6A), the CD31 was
significantly higher in the mice treated with HAL and HAL+ LV-Ctrl
as compared with the control group, whereas CD31 was significantly
lower in the combined HAL + LV-H721 treatment group in comparison
with the HAL and HAL+ LV-Ctrl groups (all P<0.05; Fig. 6B). A TUNEL assay was also performed
to detect the apoptotic cells in the tumor tissues. The results
demonstrated that combined treatment with HAL + LV-H721
significantly enhanced cell apoptosis compared with that observed
in the control, HAL and HAL + LV-Ctrl groups (P<0.01, P<0.05
and P<0.05, respectively; Fig. 6C
and D). Collectively, these results indicated that HAL
treatment may promote the microvascular growth. However,
HIF-1α knockout in tumor tissues significantly inhibited the
tumor hypoxia-mediated increase in CD31 expression and the
microvascular growth, while it promoted tumor cell apoptosis in
vivo. These results further highlight the relevance of HIF-1α
inhibition in combination with HAL in the treatment of advanced,
unresectable HCC.
Discussion
TAE, including TACE, is one of the most important
treatment strategies for advanced HCC; however, these methods have
various side effects. TAE has been suggested to aggravate the
hypoxia condition of solid tumors and inevitably activate various
pathways that promote cell invasion, tumor metastasis and
angiogenesis (27,28). Previously, Hanahan and Weinberg
(29) proposed ten hallmarks of
cancer and therapeutic targets for these hallmarks. In the present
study, HIF-1α was proposed as a potentially effective therapeutic
knockout target of the hallmarks of HCC during TAE. The results
demonstrated that HIF-1α knockout in SMMC-7721 cells
significantly suppressed cell invasiveness and migration, and
induced cell apoptosis under CoCl2-simulated hypoxic
conditions. Hepatic artery ligation (HAL) was used to mimic human
transarterial chemoembolization in mice. Results showed that the
combination method of HAL and Cas9-mediated genome editing
exhibited an anti-tumor effect in an orthotopic HCC model, as well
as prolonged the survival of tumor-bearing mice. The current
preclinical study indicates that the combination of HAL and
HIF-1α knockout may have a huge potential for further
clinical applications.
Enhanced tumor invasion, distant metastasis and
neovascularization subsequent to TAE treatment are the most severe
complications observed (30).
Furthermore, the development, progression, metastasis and
recurrence of HCC are known to be closely associated with
angiogenesis. HIF-1α is the key regulator of angiogenesis under
hypoxic conditions and a useful predictor of prognosis in patients
with HCC. Thus, HIF-1α is an ideal target for improving the
therapeutic efficacy of TAE/TACE, and previous studies have
reported that RNA interference targeting HIF-1α resulted in an
ameliorative efficacy compared with that observed in the TACE group
(31,32). In the present study, the lentiviral
delivery (LV-H721) of the CRISPR/Cas9 protein and an
HIF-1α-specific sgRNA resulted in highly efficient
HIF-1α modification (Fig.
2C), and the suppression of VEGF and MDR1 in SMMC-7721 liver
tumor cells, indicating that HIF-1α was efficiently
disrupted by the CRISPR/Cas9 system (Fig. 2D and G). These findings are
consistent with the observations of previous studies on HIF-1α,
where HIF-1α downregulation was reported to inhibit the expression
levels of VEGF and MDR1 in cancer cells (17,33).
Notably, LV-H721 infection led to HIF-1α knockout in the
SMMC-7721 ×enograft tissues (Fig.
2A). In addition, CoCl2 was used to mimic a hypoxic
tumor microenvironment in vitro (34), and the in vitro anti-tumor
effect of the HIF-1α knockout in the SMMC-7721 cells was
exhibited by the impairment of the liver cancer cell migration and
invasiveness capacities, particularly under hypoxic conditions
(Fig. 3). Under hypoxic conditions,
HIF-1α knockout reduced cell proliferation and was
accompanied by increased cell apoptosis in the SMMC-7721 cells
(Fig. 4), indicating that combining
the CRISPR/Cas9 system targeting HIF-1α with TAE in the
hypoxic tumor microenvironment may more efficient in HCC treatment.
Notably, this combination therapy of CRISPR/Cas9-mediated
HIF-1α knockout and TAE (namely the HAL + LV-H721 treatment)
conferred a more significant tumor suppressive effect and a
prolonged survival time in the HCC-bearing mice (Fig. 5). Furthermore, the reduced
expression of CD31 in the HAL + LV-H721 tumor tissues indicated
that tumor angiogenesis was effectively inhibited by the
HIF-1α knockout (Fig. 6).
Thus, HIF-1α, a key regulator of angiogenesis, cell migration and
cell invasiveness capacity, may be an effect therapeutic knockout
target during TAE in the treatment of HCC.
The emerging CRISPR/Cas9 system has several
advantages and has rapidly changed the landscape of cancer biology
(35). By simply combining the
expression of CRISPR/Cas9 and sgRNAs, highly efficient tumor genome
engineering was achieved. Relevant research studies have reported
that CRISPR/Cas9 interventions targeting hepatitis B virus (HBV)
cellular genes in cancer resulted in the cleavage of the HBV genome
and viral clearance in a mouse model (36,37).
Nevertheless, the application of the lentiviral CRISPR/Cas9 system
in clinical cases is limited by the lack of efficient delivery
methods and the toxicity of the Cas9 nuclease (38). Notably, lentivirus infection and the
CRISPR/Cas9 system mediating the knockout of a certain gene is
permanent, and causes insertion inactivation of genes and
off-target events. Therefore, the development of a delivery vector
and a CRISPR/Cas9 system that mainly targets only the cancer cell
genome is highly desirable. In the current study, SMMC-7721 cells
were first infected with LV-H721 lentivirus and then used to
establish an HCC mouse model. However, for the combination of TAE
and the lentivirus-mediated CRISPR/Cas9 system, it may be better to
inject an emulsion of materials that include the LV-H721 lentivirus
into the HCC feeding arteries, thereby blocking the blood supply to
the tumor. Furthermore, compared with the use of an intravenous
systemic injection, a more efficient HIF-1α gene knockout
may be achieved by hepatic transarterial embolization with LV-H721
due to the close encounters between the lentiviruses, HCC tissues
and a desirable HIF-1α target. Thus, an extension of the
current study could further utilize this combination strategy in
rats. Thus far, it appears that using the lentivirus-mediated
CRISPR/Cas9 system to correct an error in a certain human gene or
to remove an alien gene may be more practical and achievable during
TAE. Further studies are required to elucidate the application of
the CRISPR/Cas9 system in treating human malignancies with a
lentivirus vector.
In conclusion, the present study revealed that the
CRISPR/Cas9-mediated HIF-1α knockout enhanced the antitumor
effect of TAE in HCC, suggesting that HIF-1α may serve as a
potential clinical knockout target, in combination with TAE/TACE,
for the treatment of HCC.
Acknowledgements
We are grateful to Qingshuang Zou, Jianlong Kang and
Shiyou Chen for their contributions to this project. We thank Jie
Zhang, Wei Ma, Yang Li, Shang Chen and Weilin Lu for their animal
management and technical supports.
Funding
This study was funded by the National Natural
Science Foundation of Guangdong (grant no. 2015A030310409,
2017A030313846), and the Science and Technology Innovation
Foundation of Shenzhen (grant no. JCYJ20160422152408705).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
LPL and SYB conceived and designed the study; QL and
DA wrote the manuscript, carried out cellular and animal-associated
experiments and performed statistical analyses; DF collected the
clinical samples and analyzed the clinical information; ZW, YZ and
ZYD assisted with the collection of clinical tumor samples,
Transwell and TUNEL staining assay; DW and YL assisted with PCR,
western blotting and packaging and purification of CRISPR/Cas9
lentiviruses; RL was involved in the conception of the study; QTY
was responsible for cell culture and IHC staining. All authors read
and approved the manuscript and agreed to be accountable for all
aspects of the research in ensuring that the accuracy or integrity
of any part of the work are appropriately investigated and
resolved.
Ethics approval and consent to
participate
The present study conformed to the Ethical
Guidelines of the 2013 revision of the Declaration of Helsinki. All
the HCC patients provided written informed consent for the use of
clinical specimens for medical research. The studies using the
human specimens and the mouse experiments were approved by the
Ethics Committee of the Jinan University (Guangzhou, Guangdong,
China). All the animal experiments were conducted with the standard
guidelines for the care of animals, which were approved by the
Welfare Committee of the Center of Experimental Animals (Jinan
University, Guangzhou, Guangdong, China).
Patient consent for publication
All author consent to the publication of this
work.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
HCC
|
hepatocellular carcinoma
|
|
TAE
|
transarterial embolization
|
|
TACE
|
transarterial chemoembolization
|
|
HIF-1α
|
hypoxia inducible factor-1α
|
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2016. CA Cancer J Clin. 66:7–30. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Forner A, Llovet JM and Bruix J:
Hepatocellular carcinoma. The Lancet. 379:1245–1255. View Article : Google Scholar
|
|
3
|
Omyla-Staszewska J and Deptala A:
Effective therapeutic management of hepatocellular carcinoma-on the
basis of a clinical case. Contemp Oncol. 16:60–63. 2012.
|
|
4
|
Song DS, Nam SW, Bae SH, Kim JD, Jang JW,
Song MJ, Lee SW, Kim HY, Lee YJ, Chun HJ, et al: Outcome of
transarterial chemoembolization-based multi-modal treatment in
patients with unresectable hepatocellular carcinoma. World J
Gastroenterol. 21:2395–2404. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wang YX, De Baere T, Idee JM and Ballet S:
Transcatheter embolization therapy in liver cancer: An update of
clinical evidences. Chin J Cancer Res. 27:96–121. 2015.PubMed/NCBI
|
|
6
|
Lo CM, Ngan H, Tso WK, Liu CL, Lam CM,
Poon RT, Fan ST and Wong J: Randomized controlled trial of
transarterial lipiodol chemoembolization for unresectable
hepatocellular carcinoma. Hepatology. 35:1164–1171. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Pecot CV, Calin GA, Coleman RL,
Lopez-Berestein G and Sood AK: RNA interference in the clinic:
Challenges and future directions. Nat Rev Cancer. 11:59–67. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Sharma P and Allison JP: The future of
immune checkpoint therapy. Science. 348:56–61. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Liu Q, Yang Y, Tan X, Tao Z, Adah D, Yu S,
Lu J, Zhao S, Qin L, Qin L and Chen X: Plasmodium parasite as an
effective hepatocellular carcinoma antigen glypican-3 delivery
vector. Oncotarget. 8:24785–24796. 2017.PubMed/NCBI
|
|
10
|
Rosenberg SA and Restifo NP: Adoptive cell
transfer as personalized immunotherapy for human cancer. Science.
348:62–68. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Doudna JA and Charpentier E: Genome
editing. The new frontier of genome engineering with CRISPR-Cas9.
Science. 346:12580962014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Cong L, Ran FA, Cox D, Lin S, Barretto R,
Habib N, Hsu PD, Wu X, Jiang W, Marraffini LA and Zhang F:
Multiplex genome engineering using CRISPR/Cas systems. Science.
339:819–823. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Eid A and Mahfouz MM: Genome editing: The
road of CRISPR/Cas9 from bench to clinic. Exp Mol Med. 48:e2652016.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zucman-Rossi J, Villanueva A, Nault JC and
Llovet JM: Genetic landscape and biomarkers of hepatocellular
carcinoma. Gastroenterology. 149(1226–1239): e12242015.
|
|
15
|
Favaro E, Lord S, Harris AL and Buffa FM:
Gene expression and hypoxia in breast cancer. Genome Med. 3:1–12.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Unruh A, Ressel A, Mohamed HG, Johnson RS,
Nadrowitz R, Richter E, Katschinski DM and Wenger RH: The
hypoxia-inducible factor-1 alpha is a negative factor for tumor
therapy. Oncogene. 22:3213–3220. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kang MJ, Jung SA, Jung JM, Kim SE, Jung
HK, Kim TH, Shim KN, Yi SY, Yoo K and Moon IH: Associations between
single nucleotide polymorphisms of MMP2, VEGF, and HIF1A genes and
the risk of developing colorectal cancer. Anticancer Res.
31:575–584. 2011.PubMed/NCBI
|
|
18
|
Yang SL, Liu LP, Jiang JX, Xiong ZF, He QJ
and Wu C: The Correlation of Expression Levels of HIF-1α and HIF-2α
in hepatocellular carcinoma with capsular invasion, portal vein
tumor thrombi and patients' clinical outcome. Jpn J Clin Oncol.
44:159–167. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Chen M, Huang SL, Zhang XQ, Zhang B, Zhu
H, Yang VW and Zou XP: Reversal effects of pantoprazole on
multidrug resistance in human gastric adenocarcinoma cells by
down-regulating the V-ATPases/mTOR/HIF-1α/P-gp and MRP1 signaling
pathway in vitro and in vivo. J Cell Biochem. 113:2474–2487. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Jia ZZ, Jiang GM and Feng YL: Serum
HIF-1alpha and VEGF levels pre- and post-TACE in patients with
primary liver cancer. Chin Med Sci J. 26:158–162. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Liu K, Min XL, Peng J, Yang K, Yang L and
Zhang XM: The Changes of HIF-1alpha and VEGF expression after TACE
in patients with hepatocellular carcinoma. J Clin Med Res.
8:297–302. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yu S, Yao Y, Xiao H, Li J, Liu Q, Yang Y,
Adah D, Lu J, Zhao S, Qin L, et al: Simultaneous knockout of CXCR4
and CCR5 genes in CD4+ T cells via CRISPR/Cas9 confers resistance
to both X4- and R5-tropic human immunodeficiency virus type 1
infection. Hum Gene Ther. 29:51–67. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang S, Zeng X, Liu Y, Liang C, Zhang H,
Liu C, Du W and Zhang Z: Construction and characterization of a
PDCD5 recombinant lentivirus vector and its expression in tumor
cells. Oncol Rep. 28:91–98. 2012.PubMed/NCBI
|
|
24
|
Lopez-Sanchez LM, Jimenez C, Valverde A,
Hernandez V, Peñarando J, Martinez A, Lopez-Pedrera C,
Muñoz-Castañeda JR, De la Haba-Rodríguez JR, Aranda E, et al:
CoCl2, a mimic of hypoxia, induces formation of polyploid giant
cells with stem characteristics in colon cancer. PLoS One.
9:e991432014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhang YB, Wang X, Meister EA, Gong KR, Yan
SC, Lu GW, Ji XM and Shao G: The effects of CoCl2 on HIF-1α protein
under experimental conditions of autoprogressive hypoxia using
mouse models. Int J Mol Sci. 15:10999–11012. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ma SH, Chen GG, Yip J and Lai PB:
Therapeutic effect of alpha-fetoprotein promoter-mediated tBid and
chemotherapeutic agents on orthotopic liver tumor in mice. Gene
Ther. 17:905–912. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ikeda M, Maeda S, Shibata J, Muta R,
Ashihara H, Tanaka M, Fujiyama S and Tomita K: Transcatheter
arterial chemotherapy with and without embolization in patients
with hepatocellular carcinoma. Oncology. 66:24–31. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Soeda A, Park M, Lee D, Mintz A,
Androutsellis-Theotokis A, McKay RD, Engh J, Iwama T, Kunisada T,
Kassam AB, et al: Hypoxia promotes expansion of the CD133-positive
glioma stem cells through activation of HIF-1alpha. Oncogene.
28:3949–3959. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: The next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Liu K, Min XL, Peng J, Yang K, Yang L and
Zhang XM: The changes of HIF-1α and VEGF expression after TACE in
patients with hepatocellular carcinoma. J Clin Med Res. 8:297–302.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Chen CS, Zhao Q, Qian S, Li HL, Guo CY,
Zhang W, Yan ZP, Liu R and Wang JH: Ultrasound-guided RNA
interference targeting HIF-1 alpha improves the effects of
transarterial chemoembolization in rat liver tumors. Onco Targets
Ther. 8:3539–3548. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Chen C, Wang J, Liu R and Qian S: RNA
interference of hypoxia-inducible factor-1 alpha improves the
effects of transcatheter arterial embolization in rat liver tumors.
Tumour Biol. 33:1095–1103. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Shan JZ, Xuan YY, Zhang Q and Huang JJ:
Ursolic acid sensitized colon cancer cells to chemotherapy under
hypoxia by inhibiting MDR1 through HIF-1α. J Zhejiang Univ Sci B.
17:672–682. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wang M, Zhao X, Zhu D, Liu T, Liang X, Liu
F, Zhang Y, Dong X and Sun B: HIF-1α promoted vasculogenic mimicry
formation in hepatocellular carcinoma through LOXL2 up-regulation
in hypoxic tumor microenvironment. J Exp Clin Cancer Res.
36:602017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Sanchez-Rivera FJ and Jacks T:
Applications of the CRISPR-Cas9 system in cancer biology. Nat Rev
Cancer. 15:387–395. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zhen S, Hua L, Liu YH, Gao LC, Fu J, Wan
DY, Dong LH, Song HF and Gao X: Harnessing the clustered regularly
interspaced short palindromic repeat (CRISPR)/CRISPR-associated
Cas9 system to disrupt the hepatitis B virus. Gene Therapy.
22:404–412. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Seeger C and Ji AS: Complete spectrum of
CRISPR/Cas9-induced mutations on HBV cccDNA. Mol Ther.
24:1258–1266. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
White MK and Kamel K: CRISPR/Cas9 and
cancer targets: Future possibilities and present challenges.
Oncotarget. 7:12305–12317. 2016. View Article : Google Scholar : PubMed/NCBI
|