Introduction
Colorectal cancer (CRC) is one of the most common
cancers and the leading cause of cancer-related deaths throughout
the world (1). Approximately 1.4
million new cases are identified each year and almost 694,000
deaths are reported (2). Metastasis
from CRC develops in 40–60% of patients and the most frequent sites
of metastasis for CRC are the liver and lungs (3). The 5-year survival rate with surgical
resection of liver and lung metastasis is 30–40 and 31–48%,
respectively. However, candidates for surgical resection of liver
and lung metastatic disease include 10–25% and less than 10% of
patients, respectively (4,5). Of these cases, metastatic CRC is a
crucial cause of morbidity and mortality (6).
Metastasis is a complex series of steps that
involves the spread of tumor cells or pathogens from the primary
lesion to distant parts of the body. It is simply divided into the
processes of invasion, intravasation, and extravasation. Tumor
cells that lose the cell-cell adhesion capacity and gain cellular
motility invade the surrounding stroma via degradation of the
basement membrane and extracellular matrix (ECM). The detached
cells enter the lymph nodes and blood vessels and circulate to
remote sites. Once the circulating cells have arrived at the
targeted organs, they adhere to, and penetrate into, the
endothelium and basement membrane, followed by initiation of
angiogenesis and proliferation (7).
Metastatic cancer may be treated with chemotherapy, hormone
therapy, immunotherapy, radiation therapy, resection, and
combination of these methods (8).
However, it may be difficult to treat once the cancer has spread,
resulting in poorer prognosis.
Activation of epithelial-mesenchymal transition
(EMT) plays an important role in the initial step of the metastatic
cascade by increasing cell migration and invasion. EMT is a process
by which epithelial cells undergo functional and biochemical
changes to acquire mesenchymal cell phenotypes (9). The acquisition of mesenchymal
phenotypes leads to the loss of cell polarity and adherent tight
junction, resulting in the increase of motility and invasiveness.
The characteristic features of EMT are downregulation of epithelial
markers and adherent tight junction proteins such as E-cadherin,
desmosomes, and cytokeratins as well as upregulation of mesenchymal
markers, including N-cadherin and vimentin (10). EMT is correlated with several cancer
types, including prostate, ovarian, breast and CRC (11).
Apoptosis is a form of programmed cell death
characterized by cellular changes, including cell shrinkage,
chromatin condensation, and nuclear fragmentation while avoiding
inflammation (12). Caspases are a
family of proteolytic enzymes that regulate cell death and their
activity is essential for the induction of apoptosis. Apoptotic
caspases are divided into initiator and executioner caspases.
Initiator caspases such as caspase-8 and caspase-9 activate several
executioner caspases, resulting in the induction of apoptotic
morphological changes (13). Poly
ADP-ribose polymerase (PARP) is a nuclear enzyme related to several
processes, including DNA repair, DNA stability and cell death
(14). The anti-apoptotic factor
Bcl-2 family proteins suppress apoptosis by preventing the release
of the apoptosis-inducing factor from the mitochondria. Conversely,
Bax is a pro-apoptotic protein that may trigger the activation of
caspases, resulting in cell death (15). Apoptosis reduction or resistance is
important for carcinogenesis and malignant transformation of cancer
(16). Therefore, many cancer
treatment strategies have been designed to induce apoptosis through
the regulation of apoptotic signaling pathways.
Adenosine monophosphate-activated protein kinase
(AMPK) is a major regulator that maintains energy homeostasis and
cell proliferation. Activation of AMPK can promote energy
production and conservation processes such as glycolysis and
autophagy and inhibit energy consuming reactions including lipid
synthesis (17–19). Particularly, AMPK can suppress
carcinogenesis by inducing apoptosis and autophagy in CRC cells
(20). AMPK activation is also
associated with reduction of cancer mortality and good prognosis in
CRC (21).
Galla Rhois is an insect gall produced by the
Chinese sumac aphid (Melaphis chinensis Bell) on Rhus
chinensis (22). In traditional
Korean medicine, Galla Rhois constrains the lungs to suppress cough
and excessive perspiration, astringes the intestine to check
diarrhea, secures essence, and stops bleeding (23). In addition, Galla Rhois displays
various pharmacological activities, including antioxidant,
antidiabetic, anti-inflammatory, anti-anaphylactic, antibacterial,
antiviral, and antidiarrheal effects (24,25).
Galla Rhois contains several components such as methyl gallate,
gallic acid, 1,2,3,4,6-penta-O-galloyl-β-d-glucose (PGG), and
gallotannin (GT). Previous studies have reported that these
compounds exhibit antitumor and anti-metastatic effects in breast
cancer and fibrosarcoma (26–28).
We hypothesize that Galla Rhois water extract (GRWE) may inhibit
the metastatic ability of CRC cells. The anti-metastatic effect and
related molecular mechanism of Galla Rhois in CRC are unclear. In
the present study, we investigated the anti-metastatic properties
and underlying mechanism of GRWE using metastatic CRC cell lines
and an experimental metastatic model.
Materials and methods
Preparation of GRWE
Galla Rhois was purchased from Omniherb (Uiseong,
Korea), which is a good manufacturing practices (GMP) certified
company by the Korea Food and Drug Administration. To prepare GRWE,
Galla Rhois (100 g) was boiled at 100°C for 3 h with 1 l of
distilled water (DW). The extract was filtered through Whatman
filter paper and lyophilized. The samples were used for the
treatment of cells after dissolving in DW and filtering using a
0.22-µm syringe filter. The yield of the dried extract from the
starting materials was about 12.03%.
Cell culture
The murine colorectal carcinoma cell line colon 26
(CT26) and human colorectal adenocarcinoma cell line (HT29) were
obtained from Korean Cell Line Bank (Seoul, Korea). Cells were
cultured in Dulbecco's modified Eagle's medium (DMEM) containing
10% heat-inactivated fetal bovine serum (FBS) and 1%
penicillin-streptomycin (all from Gibco-BRL; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) at 37°C in an atmosphere of 5%
CO2.
Animals
The in vivo experiment was approved and
performed in accordance with the internationally accepted
principles for the care and use of laboratory animals by the
Institutional Animal Care and Use Committee of Wonkwang University
(WKU16-11). Twenty-four female BALB/c mice (4 weeks old, 17–18 g)
were purchased from Samtako (Osan, Korea). The mice with ad
libitum access to food and water were housed (8 mice/cage) in a
laminar air-flow room with a controlled 12-h light/dark cycle at a
constant temperature of 23±1°C and humidity of 55±1%.
Assays of cell viability
Water-soluble tetrazolium salt-8 reagent (WST-8;
Enzo Life Sciences, Farmingdale, NY, USA) was used for quantifying
cell viability. CT26 cells (2×103 cells/well) and HT29
cells (1×104 cells/well) were seeded in 96-well plates
and cultured overnight. The cells were treated with GRWE (20–100
µg/ml). After 24, 48 and 72 h of incubation, WST-8 reagent was
mixed with new medium and added to each well. The absorbance was
measured by microplate reader at 450 nm wavelength.
Apoptosis analysis
After GRWE (10–100 µg/ml) treatment for 24 h, the
cells were collected and suspended in serum-containing medium.
Cells (1×105 cells/100 µl) were transferred to a new
tube and mixed with Muse™ Annexin V & Dead Cell
Reagent (EMD Millipore, Billerica, MA, USA). Samples were incubated
for 20 min in the dark and the apoptotic cells were measured by
Muse™ Cell Analyzer (EMD Millipore).
Antibodies
Anti-PARP (cat. no. 9532), caspase-3 (cat. no.
14220), cleaved caspase-3 (cat. no. 9664), caspase-8 (cat. no.
4790), caspase-9 (cat. no. 9508), Bcl-xL (cat. no. 2764),
phospho-AMPK (cat. no. 2535), AMPK (cat. no. 2532),
phospho-extracellular signal-regulated kinase (ERK) (cat. no.
4370), phospho-p38 (cat. no. 4511), E-cadherin (cat. no. 3195) and
N-cadherin (cat. no. 13116) antibodies were purchased from Cell
Signaling Technology, Inc. (Danvers, MA, USA). Bcl-2 (cat. no.
sc-7382), Bax (cat. no. sc-7480), ERK (cat. no. sc-94), p38 (cat.
no. sc-7149), vimentin (cat. no. sc-6260), twist (cat. no.
sc-81417), and glyceraldehyde 3-phosphate dehydrogenase (GAPDH)
(cat. no. sc-47724) antibodies were purchased from Santa Cruz
Biotechnology, Inc. (Santa Cruz, CA, USA). Anti-rabbit (cat. no.
111-035-003) and anti-mouse (cat. no. 115-035-062) secondary
antibodies were purchased from Jackson ImmunoResearch Laboratories,
Inc. (Pennsylvania, PA, USA). All antibodies were diluted 1:1,000
in 3% skim milk (BD Biosciences, San Diego, CA, USA).
Western blot analysis
CT26 cells (3×105 cells/well) were seeded
in a 6-well plate and treated with GRWE (10, 50 and 100 µg/ml).
After treatment for 24 h, the cells were lysed with ice-cold lysis
buffer (iNtRON Biotech, Seoul, Korea) for 1 h. Total lysates were
centrifuged at 14,000 × g for 10 min, and the supernatants were
collected for the determination of the extracted protein
concentration using the Lowry method. Samples were mixed with 2X
buffer, separated by sodium dodecyl sulfate polyacrylamide gel
electrophoresis (SDS-PAGE) using 10% acrylamide gels and
transferred onto polyvinylidene fluoride (PVDF) membranes. The
membranes were incubated with 5% skim milk for at least 1 h. After
washing with 0.1% PBST (0.1% Tween-20 in PBS), the membranes were
incubated with primary antibodies for 3 h, followed by their
treatment with horseradish peroxidase-conjugated secondary
antibodies for 1 h at room temperature. The protein blots were
detected using an enhanced chemiluminescence (ECL) system (Santa
Cruz Biotechnology, Inc.). The density of western blot bands was
quantified by densitometric analysis using ImageJ software
(National Institutes of Health, Bethesda, MD, USA).
Real-time reverse transcription
polymerase chain reaction (RT-PCR)
RNA-spin™ extraction Kit (iNtRON Biotech) was used
to extract total RNA in accordance with the manufacturer's
protocol. First-strand cDNA synthesis was performed using High
Capacity RNA-to-cDNA Kit (Applied Biosystems; Thermo Fisher
Scientific, Inc.). Reverse transcription was conducted at 37°C for
60 min and then at 95°C for 5 min. Real-time RT-PCR was carried out
using a Power SYBR® Green PCR Master Mix with a
StepOnePlus™ Real-Time PCR System (Applied Biosystems;
Thermo Fisher Scientific, Inc.). The PCR cycling condition was as
follows: one cycle at 95°C for 10 min, then 40 cycles of 95°C for
15 sec, 60°C for 1 min, 95°C for 15 sec and melt curve at 60°C for
1 min, 95°C for 15 sec. Sequences of the primers used for murine
genes were as follows: E-cadherin, 5′-AATGGCGGCAATGCAATCCCAAGA-3′
and 5′-TGCCACAGACCGATTGTGGAGATA-3′; N-cadherin,
5′-TGGAGAACCCCATTGACATT-3′ and 5′-TGATCCCTCAGGAACTGTCC-3′;
vimentin, 5′-CGGAAAGTGGAATCCTTGCA-3′ and
5′-CACATCGATCTGGACATGCTG-3′; Twist, 5′-AGCTACGCCTTCTCCGTCT-3′ and
5′-TCCTTCTCTGGAAACAATGACA-3′; MMP-2, 5′-CCCCATGAAGCCTTGTTTACC-3′
and 5′-TTGTAGGAGGTGCCCTGGAA-3′; MMP-9, 5′-AGACCAAGGGTACAGCCTGTTC-3′
and 5′-GGCACGCTGGAATGATCTAAG-3′; GAPDH, 5′-GACATGCCGCCTGGAGAAAC-3′
and 5′-AGCCCAGGATGCCCTTTAGT-3′. The mRNA expression of genes was
normalized to the expression of the gene encoding GAPDH. The
real-time RT-PCR data was analyzed using comparative Cq method
(29).
Immunofluorescence
CT26 cells (3×103 cells/well) were grown
on an 8-well chamber slide (Thermo Fisher Scientific, Inc.) and
treated with GRWE. The cells were fixed in 4% paraformaldehyde for
15 min and permeabilized with 0.1% Triton X-100 for 10 min. After
blocking with blocking buffer (3% bovine serum albumin and 0.3%
Triton X-100 in PBS), the cells were incubated with the primary
antibody at 4°C, overnight. Following incubation, the cells were
treated with Alexa Fluor 488-conjugated secondary antibody (Thermo
Fisher Scientific, Inc.) for 2 h and nuclei stained with
4′,6-diamidino-2-phenylindole (DAPI) (Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany). Images were acquired using a fluorescence
microscope (Zeiss Observer A1 microscope; Carl Zeiss AG,
Oberkochen, Germany).
Wound healing assay
CT26 cells (5×105 cells/well) were grown
in a 6-well plate to form a confluent monolayer. A wound was
created using a 200-µl micropipette tip and the detached cells were
removed. The cells were treated with GRWE (1, 5 and 10 µg/ml) in
the absence of FBS for 48 h. Images were captured under
EVOS® XL Core Imaging System (Thermo Fisher Scientific,
Inc.).
Invasion assay
Matrigel-coated BD Falcon cell culture chamber (BD
Biosciences) was used to estimate the invasive activity of cancer
cells. The lower wells were filled with 750 µl DMEM containing 10%
FBS. Cells (5×104 cells) were seeded in the upper part
of the transwell chambers in 200 µl FBS-free DMEM with GRWE (1, 5
and 10 µg/ml) and chambers were placed on a 24-well plate for 24 h.
The upper inserts were fixed with 3.7% formaldehyde in PBS and
washed twice with PBS. The fixed cells were permeabilized with
methanol for 25 min and stained with Giemsa solution
(Sigma-Aldrich; Merck KGaA) for 25 min. The inner sides of the
chambers were cleaned with a swab and dried. The stained cells were
observed under EVOS® XL Core Imaging System.
Zymography
For gelatin zymography, CT26 cells (5×105
cells/well) were seeded in a 6-well plate and treated with GRWE for
24 h. The supernatant from the GRWE-treated cells was mixed with 2X
zymogram sample buffer (KOMA Biotech, Seoul, Korea) and the samples
were electrophorized using SDS polyacrylamide gel containing 1%
gelatin in the zymogram running buffer (KOMA Biotech). The gel was
renatured in zymogram renaturing buffer for 30 min and incubated in
zymogram developing buffer (both from KOMA Biotech) at 37°C
overnight. The gel was stained with Brilliant Blue R staining
solution (ELPIS Biotech, Daejeon, Korea) for 30 min.
Experimental lung metastatic mouse
model
CT26 cells (1×105 cells/200 µl PBS/mouse)
were injected into mice via the lateral tail vein. GRWE and DW were
orally administrated 2 h prior to the injection of CT26 cells and
then, every day. The mice were sacrificed by cervical dislocation
after 14 days and lung tissues were stained and fixed with Bouin's
solution (Sigma-Aldrich; Merck KGaA). The weight of the lungs was
measured and the number of nodules in the lungs was counted to
evaluate CRC metastasis.
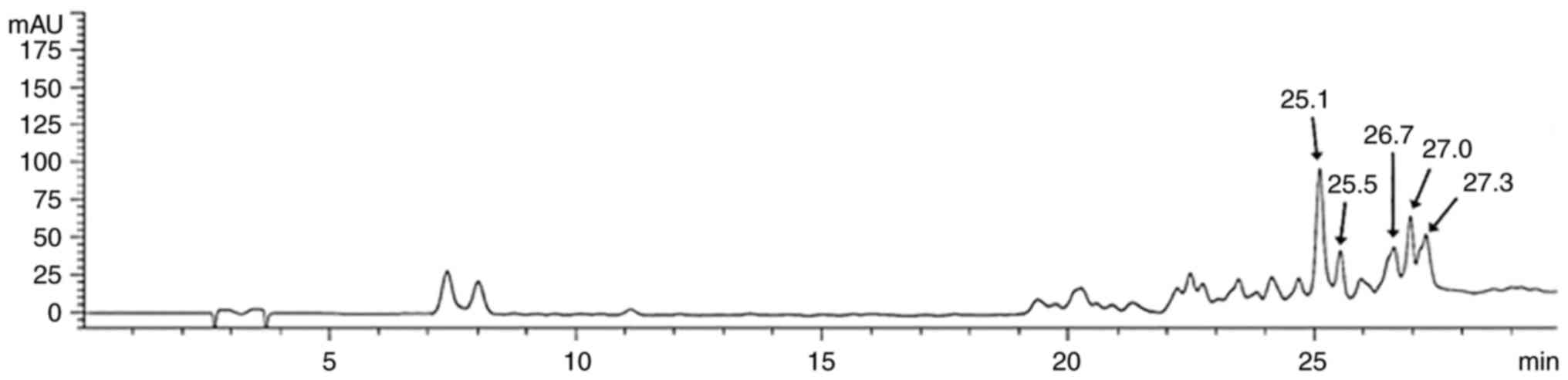
High-performance liquid chromatography
(HPLC) analysis
During analysis of GT from GRWE, GT (molecular
weight: 1701.20 g/mol; molecular formula:
C76H52O46; Santa Cruz
Biotechnology, Inc.) was used as the standard. GRWE and the
standard compound were dissolved in methanol (MeOH) and filtered
through a 0.45-µm syringe filter before injection. An HPLC system
comprised of an Agilent 1200 Series HPLC system (Agilent
Technologies, Santa Clara, CA, USA) with a UV-vis detector was
used. Separation was carried out on an YMC-Triart C18
column (150×4.6 mm I.D., 5 µm,) maintained at 40°C. The mobile
phase was comprised of solvent A (0.1% formic acid in water) and
solvent B (0.1% formic acid in acetonitrile). The gradient
condition of the mobile phase was as follows: 0–5 min, 15% A; 5–10
min, 15–20% A; 10–25 min, 20–35% A; 25–30 min, 35–15% A. Analysis
was performed at a flow rate of 1.0 ml/min with the detection
wavelength fixed at 280 nm (Fig.
1).
Statistical analysis
All data were presented as the mean ± standard
deviation (SD). ANOVA with Tukey's post hoc test was used to
determine the statistical significance. SPSS Statistics v18 (IBM
Corp., Armonk, NY, USA) was used as the statistical analysis
software. A value of P<0.05 was considered to indicate a
statistically significant difference.
Results
Effect of GRWE on the proliferation of
CRC cells
To investigate the cytotoxicity of GRWE on
metastatic CRC cells, cell viability was determined by WST-8 assay.
CT26 and HT29 cells were treated with GRWE at concentrations of
20–100 µg/ml. As shown in Fig. 2A and
B, the viability of GRWE-treated cells decreased as compared
with that of the control cells. Furthermore, treatment with GRWE
for 72 h resulted in morphological changes in CT26 and HT29 cells
(Fig. 2C).
Apoptotic effect of GRWE on CT26
cells
Annexin V assay was performed after treatment of
cells with GRWE (10, 50, and 100 µg/ml) for 24 h to determine
whether GRWE-induced cell death was related to the apoptosis of
cells. As shown in Fig. 3A and B,
treatment with GRWE resulted in an increase in the apoptosis of
CT26 cells in a dose-dependent manner. To further investigate the
mechanisms underlying GRWE-induced apoptosis, the expression of
apoptosis-related proteins was detected by western blotting.
Cleavage of caspase-3 and PARP was induced following exposure of
CT26 cells to GRWE for 24 h. In addition, GRWE treatment reduced
the expression of caspase-8, caspase-9, Bcl-2, and Bcl-xL and
increased the protein level of Bax in CT26 cells (Fig. 3C).
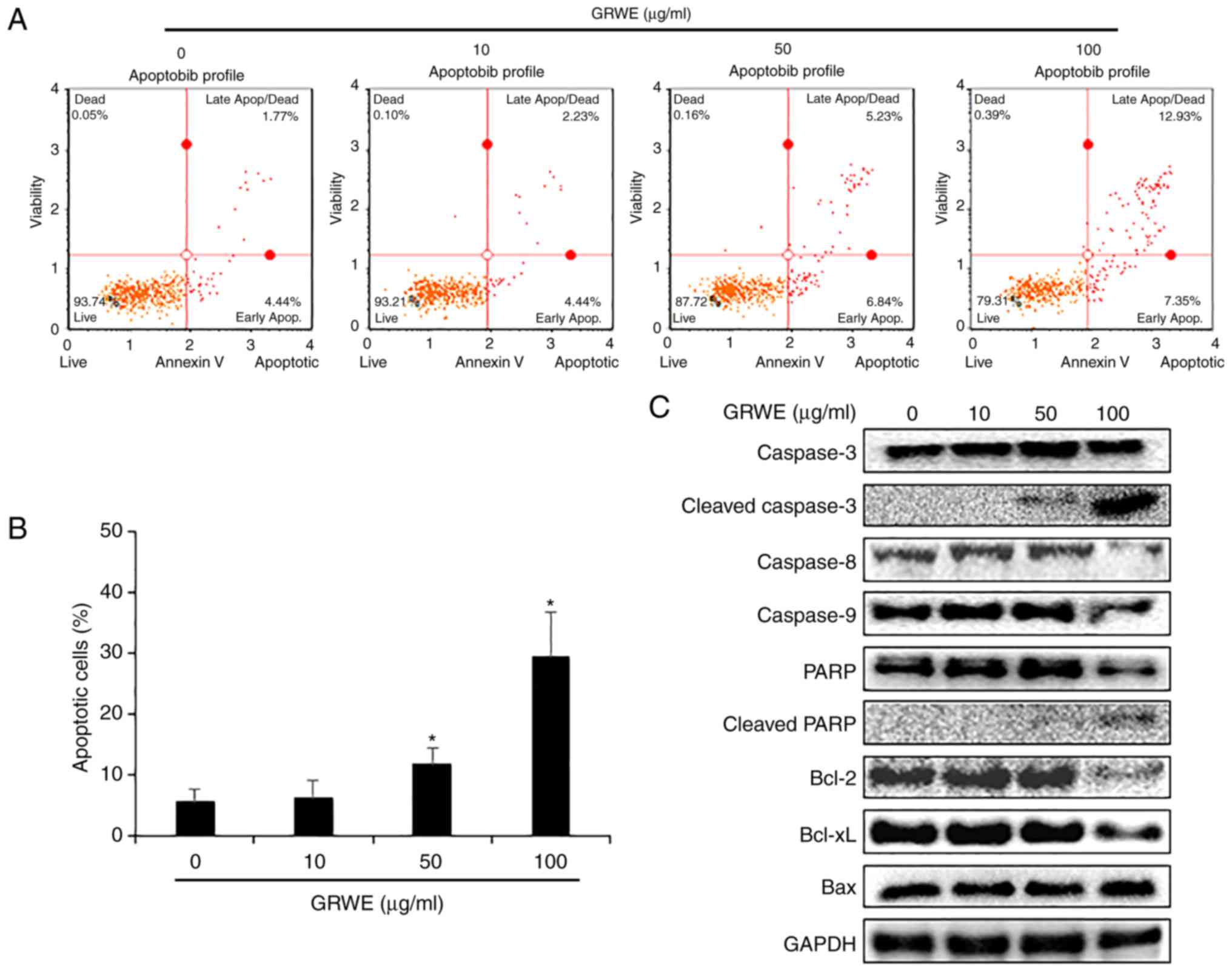 | Figure 3.GRWE induces apoptosis in CT26 cells
through extrinsic and intrinsic pathways. (A) CT26 cells were
incubated with GRWE (10, 50, and 100 µg/ml) for 24 h and stained
with Annexin V and 7-AAD. The image is a representative of three
independent experiments. (B) The statistical graph of apoptotic
cells from Annexin V staining at different concentrations of GRWE
is displayed. (C) CT26 cells were treated with the indicated
concentrations of GRWE for 24 h. The expression of caspase-3,
cleaved caspase-3, caspase-8, caspase-9, PARP, cleaved PARP, Bcl-2,
Bcl-xL and Bax were detected by western blotting using
corresponding antibodies. Results are expressed as the mean ± SD of
three independent experiments. *P<0.05. GRWE, Galla Rhois water
extract. |
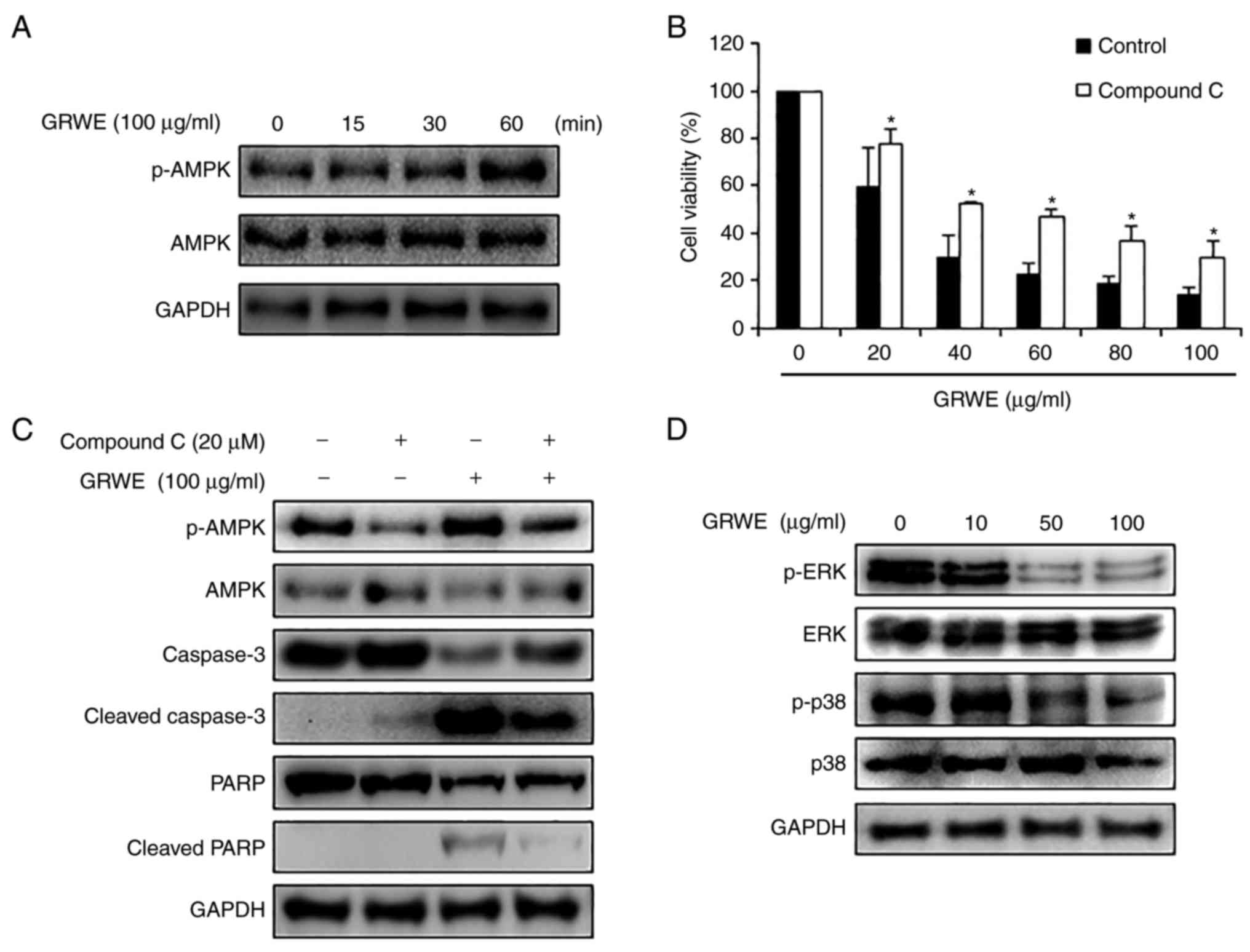
GRWE induces apoptosis via AMPK
activation in CT26 cells
Activation of AMPK inhibits cell proliferation by
promoting apoptosis of cancer cells. Various anticancer drugs and
plant extracts can induce AMPK-mediated apoptosis in cancer cells
(30). A previous study reported
that PGG, a component of Galla Rhois, induced breast cancer cell
death through phosphorylation of AMPK (31). Therefore, we hypothesized that
apoptosis of CT26 cells by GRWE was related to activation of AMPK.
As revealed in Fig. 4A,
phosphorylation of AMPK was increased after GRWE treatment in a
time-dependent manner. To confirm whether GRWE-induced AMPK
phosphorylation was involved in apoptosis of CT26 cells, AMPK
inhibitor compound C (20 µM) was pre-treated to CT26 cells for 4 h,
and then treated with GRWE (20–100 µg/ml) for 72 h. Blockage of
AMPK activation recovered GRWE-decreased viability of CT26 cells
(Fig. 4B). To further investigate
whether apoptosis-related factors were regulated by AMPK
activation, the protein levels of cleaved caspase-3 and PARP were
detected in compound C and GRWE-treated CT26 cells. Cleavage of
caspase-3 and PARP was induced following treatment with GRWE,
whereas AMPK inhibitor compound C suppressed this effect of GRWE
(Fig. 4C). Several studies have
shown that AMPK activators induce apoptosis via modulation of the
MAPK pathway in human cancer cells (32). AMPK activation induces apoptosis via
the downregulation of ERK in cancer cells, and AMPK activator AICAR
inhibits p38 MAPK (33,34). Based on these studies, we confirmed
whether GRWE could regulate the phosphorylation of ERK and p38. As
expected, GRWE reduced the phosphorylation of ERK and p38 (Fig. 4D). These results indicated that
GRWE-induced apoptosis occurred through the AMPK-ERK/p38 signaling
pathway.
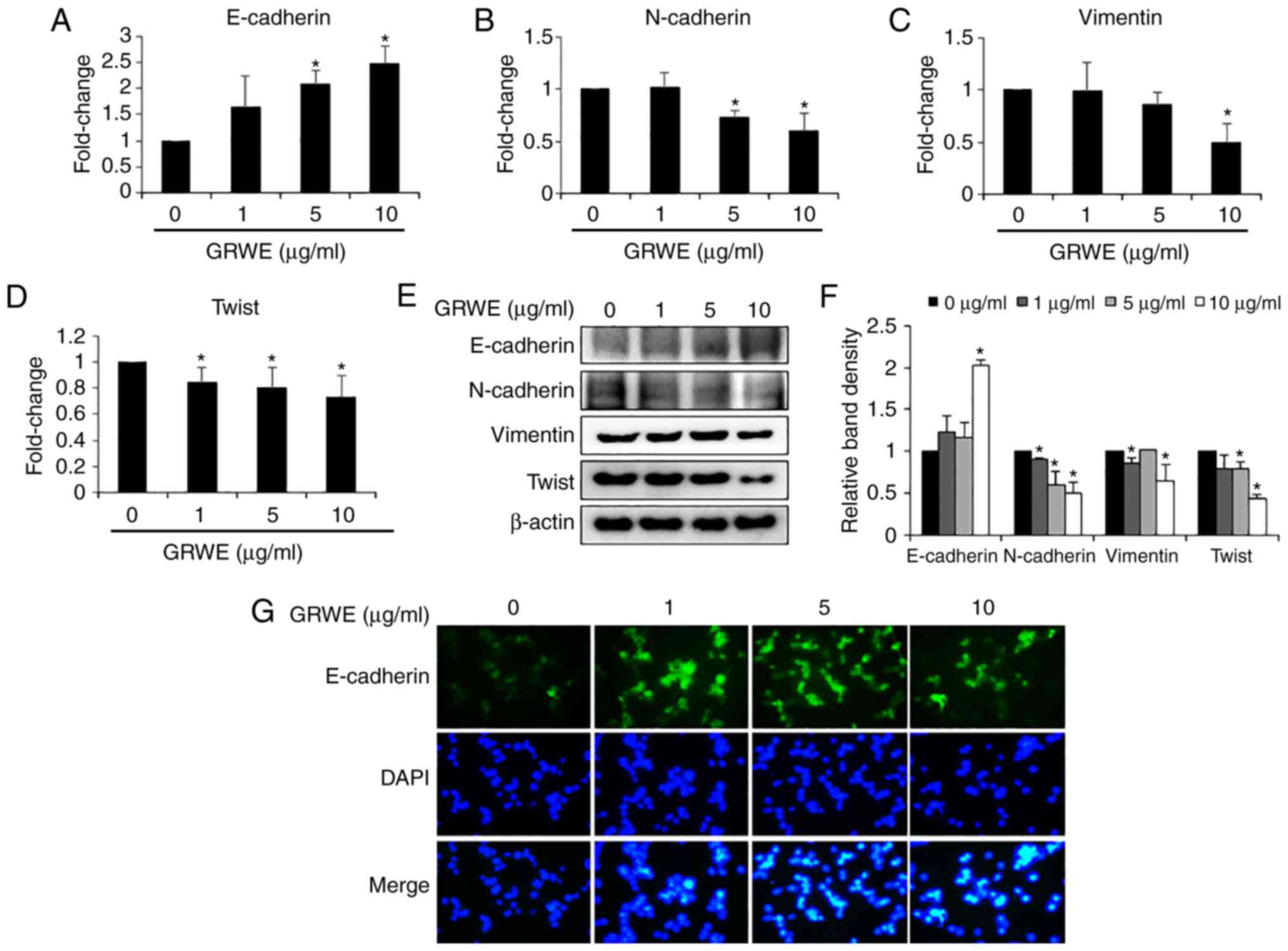
Effect of GRWE on the expression of
EMT markers in CT26 cells
Gene expression analysis was conducted to elucidate
whether GRWE regulated the expression of EMT markers, which are
involved in metastatic properties of cancer cells. The mRNA level
of epithelial phenotypic marker E-cadherin was upregulated after
GRWE treatment (Fig. 5A). In
addition, the expression of mesenchymal phenotypic markers
N-cadherin, vimentin, and Twist was downregulated in GRWE-treated
CT26 cells (Fig. 5B-D). Consistent
with the mRNA expression levels, GRWE increased the protein levels
of E-cadherin and reduced N-cadherin, vimentin, and Twist
expression (Fig. 5E and F). We
confirmed GRWE-mediated enhanced expression of E-cadherin by
immunofluorescence (Fig. 5G).
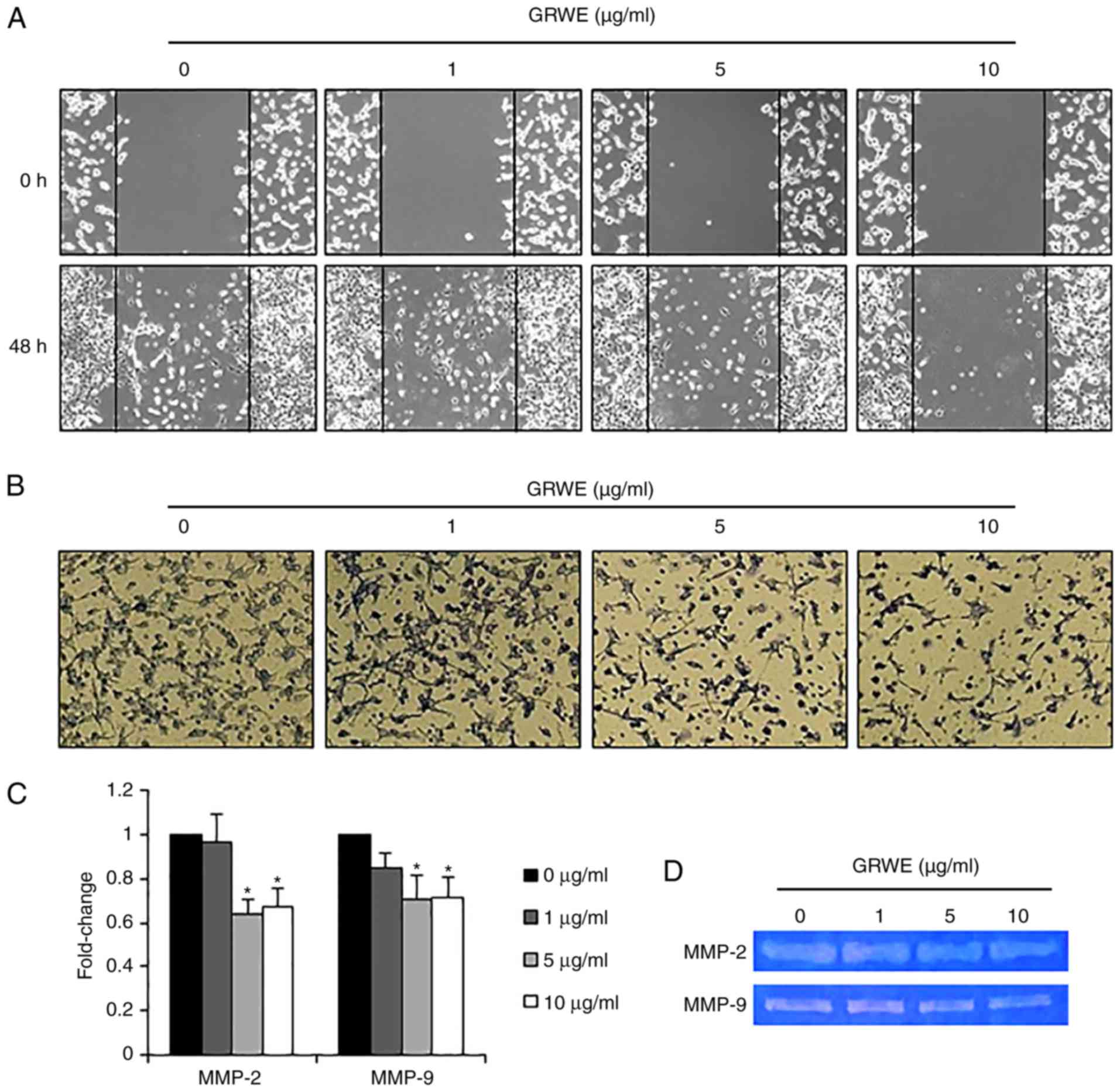
Effect of GRWE on the migratory and
invasive abilities of CT26 cells
After EMT, cancer cells acquire migratory and
invasive properties, resulting in metastasis (35). We performed a wound healing assay to
explore the effect of GRWE on the migration of CT26 cells.
Following 48 h of incubation, the control cells were more
proficient in repairing wounds as compared with the GRWE-treated
cells. Treatment with GRWE (1, 5, and 10 µg/ml) resulted in the
inhibition of the migratory ability of CT26 cells (Fig. 6A). An invasion assay was carried out
using a Matrigel-coated Transwell chamber to further investigate
the effect of GRWE on the invasion ability of CT26 cells. As
revealed in Fig. 6B, the
infiltration of CT26 cells in the Matrigel-coated membrane was
decreased following GRWE treatment in a dose-dependent manner. In
addition, we assessed the mRNA expression of matrix
metalloproteinase (MMP)-2 and MMP-9 using real-time RT-PCR and
determined that MMP-2 and MMP-9 expression was downregulated by
GRWE treatment (Fig. 6C). We also
analyzed the activity of MMP-2 and MMP-9 by gelatin zymography and
found that GRWE reduced the activity of MMP-2 and MMP-9 in CT26
cells (Fig. 6D).
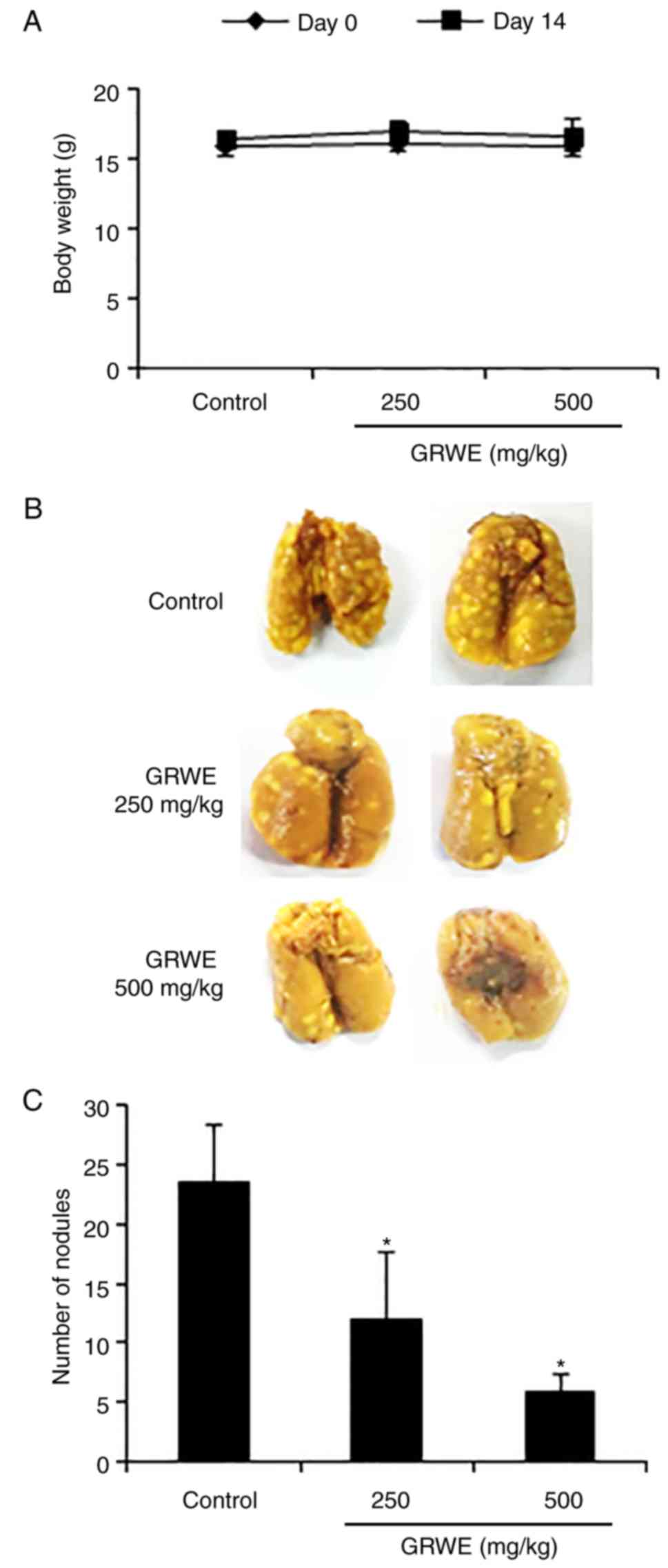
Effect of GRWE on the lung metastatic
ability of CT26 cells
To investigate whether GRWE inhibited the lung
metastatic ability of CRC cells in mice, we employed an
experimental lung metastatic model. Body weight remained unaffected
following treatment with GRWE (Fig.
7A). After intravenous injection of CT26 cells, the number of
metastatic tumor nodules in the lungs increased initially but were
reduced after the oral administration of GRWE (250 and 500 mg/kg)
for 2 weeks (Fig. 7B and C).
Discussion
This study elucidated the inhibitory effect of GRWE
on colorectal metastasis using metastatic CRC cells and an
experimental lung metastatic model. Galla Rhois has been used to
treat various diseases in East Asia. Although the pharmacological
actions of Galla Rhois are well studied, its effects on cancer as
well as the mechanism underlying the anticancer effects remain to
be addressed.
Malignant tumors develop from cancer cells that are
resistant to programmed cell death and undergo indefinite
proliferation. Therefore, induction of apoptosis contributes to the
restriction of cancer progression, and many approved cancer
therapies induce apoptosis in cancer cells (36,37).
According to recent studies, GRWE may induce cell death in various
human cancer cell lines, including HCT116, AGS, MDA-MB-231, A549,
and SK-Hep-1. In particular, ellagic acid promoted apoptosis of
human CRC cell line HCT116 via caspase-dependent pathway (25). In the present study, we observed
that GRWE significantly inhibited the proliferation of CRC cell
lines CT26 and HT29 (Fig. 2A and
B). The antiproliferative effect of GRWE was exerted through
the induction of apoptosis in CT26 cells (Fig. 3A and B).
The initiation of apoptosis occurs through strictly
controlled mechanisms. Apoptosis is mediated by two basic signaling
pathways-the extrinsic and intrinsic pathway. The extrinsic pathway
is triggered through the binding of death ligands to receptors.
Activated caspase-8 subsequently induces cleavage of caspase-3 and
PARP or activates the mitochondria-mediated intrinsic pathway. The
intrinsic pathway is activated by several stimuli such as DNA
damage and deprivation of cell survival factors (38,39).
This is controlled by a series of Bcl-2 family members. The
anti-apoptotic proteins Bcl-2 and Bcl-xL arrest the emission of
cytochrome c, while the pro-apoptotic protein Bax promotes
the release of cytochrome c from mitochondria. The release
of the cytochrome c is followed by the activation of
downstream caspases (caspase-9 and caspase-3), thereby leading to
apoptosis (40). In the present
study, GRWE induced the cleavage of caspase-3 and PARP and reduced
the expression of caspase-8 and caspase-9. Furthermore, the
expression of anti-apoptotic proteins Bcl-2 and Bcl-xL were
decreased, whereas the expression of pro-apoptotic protein Bax was
increased following GRWE treatment (Fig. 3C). These results demonstrated that
GRWE-induced apoptosis was mediated through the extrinsic and
intrinsic apoptotic pathways in CT26 cells.
Activation of AMPK can inhibit proliferation of
cancer cells by inducing apoptosis (30). In addition, AMPK activators induce
apoptosis via the MAPKs signaling pathway in human cancer cells
(32). Phosphorylation of ERK
generally suppresses apoptosis of cells by regulating apoptotic
protein expression such as the Bcl-2 family members and caspases
(41,42). p38 MAPK can also modulate survival,
growth and differentiation of various cancer cells (43–45).
Several studies have reported that p38 MAPK inhibited autophagy and
apoptosis, thereby promoting survival of CRC cells (34,46).
In the present study, GRWE induced the phosphorylation of AMPK and
inhibited the phosphorylation of ERK and p38 (Fig. 4A and D). Inhibition of AMPK activity
partially recovered cell viability and the expression of
apoptosis-related proteins in GRWE-treated CT26 cells (Fig. 4B and C). Based on these results,
GRWE induced apoptosis of CRC cells via the AMPK-ERK/p38 signaling
pathway.
Epithelial cells are closely connected to
surrounding cells by tight junctions, adherens junctions and gap
junctions. In addition, these cells display cell polarity and are
attached by a basal lamina. Conversely, mesenchymal cells rarely
form cell-cell junctions, interplay only focally, and lack
polarity. During EMT, the stationary cancer cells gain motility and
invasiveness. Therefore, the EMT process plays an important role in
promoting metastasis. EMT is induced by the interaction of
extracellular signals. The activation of the signaling pathway
enhances transcriptional regulators, including Snail, Slug, and
Twist. The main target of transcriptional regulators is the
suppression of E-cadherin, the cell-cell adhesion molecule
(47,48). The downregulation of E-cadherin is
often accompanied by the upregulation of N-cadherin, the invasion
promoter molecule (49). In the
present study, GRWE at non-cytotoxic concentrations increased the
expression of the epithelial marker E-cadherin and reduced the
expression of mesenchymal markers, including N-cadherin, vimentin,
and twist (Fig. 5). These results
indicated that GRWE may suppress colorectal metastasis through the
inhibition of the EMT process.
MMPs play a critical role in many events during
malignant transformation, including tumor growth, angiogenesis,
invasion, and metastasis. EMT of cancer cells contributes to the
production of MMPs, and increased MMPs promote the pathogenic EMT
process (35). In particular, MMP-2
and MMP-9 may induce degradation of ECM, thereby facilitating
migration and invasion in most cancers (50). The non-cytotoxic concentrations of
GRWE inhibited the motility and invasiveness of CT26 cells
(Fig. 6A and B). Moreover, the
expression and activity of MMP-2 and MMP-9 were suppressed by GRWE
treatment (Fig. 6C and D). These
results confirmed that GRWE may inhibit the migration and invasion
ability through the reduction in MMP-2 and MMP-9 expression.
In the present study, HPLC chromatogram of GRWE
revealed characteristic peaks for GT (25.1, 25.5, 26.7, 27.0 and
27.3 min) (Fig. 1), indicative of
the presence of GT in GRWE. The GT-rich Caesalpinia spinosa
fraction is known to exert antitumor and anti-metastatic effects in
murine breast cancer models through the reduction of serum
interleukin (IL)-6 level and cell cycle arrest in the S phase
(51,52). Moreover, GT inhibited primary CRC
tumors by suppressing the expression of nuclear factor-κB
(NF-κB)-regulated inflammatory cytokines and proliferation of HT29
and HCT116 cells (53). GT present
in GRWE was expected to exert an anti-metastatic effect on CRC
cells. Further studies are required to evaluate the potential of GT
and other GRWE contents to inhibit metastatic abilities of CRC
cells and colorectal metastasis in in vivo models.
To the best of our knowledge, this is the first
study to demonstrate the anti-metastatic potential of GRWE against
metastatic CRC cells in vitro and in vivo. In
summary, GRWE decreased the lung metastasis of CRC cells by
inducing apoptosis and suppressing metastatic phenotypes, including
EMT, migration, and invasion. Thus, GRWE may be used as a potential
therapeutic agent for the treatment of colorectal metastasis.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Wonkwang
University in 2016.
Availability of data and materials
The datasets used during the present study are
available from the corresponding author upon reasonable
request.
Authors' contributions
JYK and SHH designed the experiments. JGM and JYK
wrote the manuscript. JGM, JYK, YHH, SHP and HDJ carried out the
experiments. SL performed the HPLC analysis. SHH supervised the
entire experiments and analyses. All authors read and approved the
manuscript and agree to be accountable for all aspects of the
research in ensuring that the accuracy or integrity of any part of
the work are appropriately investigated and resolved.
Ethics approval and consent to
participate
The in vivo experiment was approved and
performed in accordance with the internationally accepted
principles for the care and use of laboratory animals by the
Institutional Animal Care and Use Committee of Wonkwang University
(WKU16-11).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Haggar FA and Boushey RP: Colorectal
cancer epidemiology: Incidence, mortality survival, and risk
factors. Clin Colon Rectal Surg. 22:191–197. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Dromain C, Caramella C, Dartigues P, Goere
D, Ducreux M and Deschamps F: Liver, lung and peritoneal metastases
in colorectal cancers: Is the patient still curable? What should
the radiologist know. Diagn Interv Imaging. 95:513–523. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Tsoulfas G, Pramateftakis MG and Kanellos
I: Surgical treatment of hepatic metastases from colorectal cancer.
World J Gastrointest Oncol. 3:1–9. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Dahabre J, Vasilaki M, Stathopoulos GP,
Kondaxis A, Iliadis K, Papadopoulos G, Stathopoulos J, Rigatos S,
Vasilikos K and Koutantos J: Surgical management in lung metastases
from colorectal cancer. Anticancer Res. 27:4387–4390.
2007.PubMed/NCBI
|
|
6
|
Kim HJ, Kye BH, Lee JI, Lee SC, Lee YS,
Lee IK, Kang WK, Cho HM, Moon SW and Oh ST: Surgical resection for
lung metastases from colorectal cancer. J Korean Soc Coloproctol.
26:354–358. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Martin TA, Ye L, Sanders AJ, Lane J and
Jiang WG: Cancer invasion and metastasis: Molecular and cellular
perspective. Metastatic Cancer: Clinical and Biological
Perspectives. Jandial R: Landes Bioscience; Austin: pp. 135–168.
2013
|
|
8
|
Mishra J, Drummond J, Quazi SH, Karanki
SS, Shaw JJ, Chen B and Kumar N: Prospective of colon cancer
treatments and scope for combinatorial approach to enhanced cancer
cell apoptosis. Crit Rev Oncol Hematol. 86:232–250. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kalluri R and Weinberg RA: The basics of
epithelial-mesenchymal transition. J Clin Inv est. 119:1420–1428.
2009. View
Article : Google Scholar
|
|
10
|
Garg M: Epithelial-mesenchymal
transition-activating transcription factors-multifunctional
regulators in cancer. World J Stem Cells. 5:188–195. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Heerboth S, Housman G, Leary M, Longacre
M, Byler S, Lapinska K, Willbanks A and Sarkar S: E MT and tumor
metastasis. Clin Transl Med. 4:62015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lowe SW and Lin AW: Apoptosis in cancer.
Carcinogenesis. 23:485–495. 2000. View Article : Google Scholar
|
|
13
|
Olsson M and Zhivotovsky B: Caspases and
cancer. Cell Death Differ. 18:1441–1449. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Herceg Z and Wang ZQ: Functions of
poly(ADP-ribose) polymerase (PARP) in DNA repair, genomic integrity
and cell death. Mutat Res. 477:97–110. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Tsujimoto Y: Role of Bcl-2 family proteins
in apoptosis: Apoptosomes or mitochondria? Genes Cells. 3:697–707.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wong RS: Apoptosis in cancer: From
pathogenesis to treatment. J Exp Clin Cancer Res. 30:872011.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Monteverde T, Muthalagu N, Port J and
Murphy DJ: Evidence of cancer-promoting roles for AMPK and related
kinases. FEBS J. 282:4658–4671. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Schultze SM, Hemmings BA, Niessen M and
Tschopp O: P I3K/AKTMA PK and AMPK signalling: Protein kinases in
glucose homeostasis. Expert Rev Mol Med. 14:e12012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hardie DG, Ross FA and Hawley SA: AMPK: A
nutrient and energy sensor that maintains energy homeostasis. Nat
Rev Mol Cell Biol. 13:251–262. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
20
|
Rehman G, Shehzad A, Khan AL and Hamayun
M: Role of AMP-activated protein kinase in cancer therapy. Arch
Pharm. 347:457–468. 2014. View Article : Google Scholar
|
|
21
|
Baba Y, Nosho K, Shima K, Meyerhardt JA,
Chan AT, Engelman JA, Cantley LC, Loda M, Giovannucci E, Fuchs CS
and Ogino S: Prognostic significance of AMP-activated protein
kinase expression and modifying effect of MAPK3/1 in colorectal
cancer. Br J Cancer. 103:1025–1033. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ahn YJ, Lee CO, Kweon JH, Ahn JW and Park
JH: Growth-inhibitory effects of Galla Rhois-derived tannins on
intestinal bacteria. J Appl Microbiol. 84:439–443. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Go J, Kim JE, Koh EK, Song SH, Seong JE,
Park CK, Lee HA, Kim HS, Lee JH, An BS, et al: Hepatotoxicity and
nephrotoxicity of gallotannin-enriched extract isolated from Galla
Rhois in ICR mice. Lab Anim Res. 31:101–110. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kim SH, Park HH, Lee S, Jun CD, Choi BJ,
Kim SY, Kim SH, Kim DK, Park JS, Chae BS, et al: The
anti-anaphylactic effect of the gall of Rhus javanica is mediated
through inhibition of histamine release and inflammatory cytokine
secretion. Int Immunopharmacol. 5:1820–1829. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yim NH, Gu MJ, Hwang YH, Cho WK and Ma JY:
Water extract of Galla Rhois with steaming process enhances
apoptotic cell death in human colon cancer cells. Integr Med Res.
5:284–292. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lee HJ, Seo NJ, Jeong SJ, Park Y, Jung DB,
Koh W, Lee HJ, Lee EO, Ahn KS, Ahn KS, et al: Oral administration
of penta-O-galloyl-β-D-glucose suppresses triple-negative breast
cancer xenograft growth and metastasis in strong association with
JAK1-STAT3 inhibition. Carcinogenesis. 32:804–811. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Deiab S, Mazzio E, Eyunni S, McTier O,
Mateeva N, Elshami F and Soliman KF:
1,2,3,4,6-Penta-O-galloylglucose within Galla Chinensis inhibits
human LDH-A and attenuates cell proliferation in MDA-MB-231 breast
cancer cells. Evid Based Complement Alternat Med. 2015:2769462015.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ata N, Oku T, Hattori M, Fujii H, Nakajima
M and Saiki I: Inhibition by galloylglucose (GG6-10) of tumor
invasion through extracellular matrix and gelatinase-mediated
degradation of type IV collagens by metastatic tumor cells. Oncol
Res. 8:503–511. 1996.PubMed/NCBI
|
|
29
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2−ΔΔCT method. Methods. 25:402–408. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Chen MB, Zhang Y, Wei MX, Shen W, Wu XY,
Yao C and Lu PH: Activation of AMP-activated protein kinase (AMPK)
mediates plumbagin-induced apoptosis and growth inhibition in
cultured human colon cancer cells. Cell Signal. 25:1993–2002. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Chai Y, Lee HJ, Shaik AA, Nkhata K, Xing
C, Zhang J, Jeong SJ, Kim SH and Lu J:
Penta-O-galloyl-beta-D-glucose induces G1 arrest and DNA
replicative S-phase arrest independently of cyclin-dependent kinase
inhibitor 1A, cyclin-dependent kinase inhibitor 1B and P53 in human
breast cancer cells and is orally active against triple negative
xenograft growth. Breast Cancer Res. 12:R672010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Li W, Saud SM, Young MR, Chen G and Hua B:
Targeting AMPK for cancer prevention and treatment. Oncotarget.
6:7365–7378. 2015.PubMed/NCBI
|
|
33
|
Plews RL, Mohd Yusof A, Wang C, Saji M,
Zhang X, Chen CS, Ringel MD and Phay JE: A novel dual AMPK
activator/mTOR inhibitor inhibits thyroid cancer cell growth. J
Clin Endocrinol Metab. 100:E748–E756. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Kim MJ, Yun H, Kim DH, Kang I, Choe W, Kim
SS and Ha J: AMP-activated protein kinase determines apoptotic
sensitivity of cancer cells to ginsenoside-Rh2. J Ginseng Res.
38:16–21. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Son H and Moon A: Epithelial-mesenchymal
transition and cell invasion. Toxicol Res. 26:245–252. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Plati J, Bucur O and Khosravi-Far R:
Apoptotic cell signaling in cancer progression and therapy. Integr
Biol. 3:279–296. 2011. View Article : Google Scholar
|
|
37
|
Takeda K, Stagg J, Yagita H, Okumura K and
Smyth MJ: Targeting death-inducing receptors in cancer therapy.
Oncogene. 26:3745–3757. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Su Z, Yang Z, Xu Y, Chen Y and Yu Q:
Apoptosis, autophagy, necroptosis, and cancer metastasis. Mol
Cancer. 14:482015. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Hideshima T and Anderson KC: Molecular
mechanisms of novel therapeutic approaches for multiple myeloma.
Nat Rev Cancer. 2:927–937. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
40
|
Han YH, Kee JY, Kim DS, Mun JG, Jeong MY,
Park SH, Choi BM, Park SJ, Kim HJ, Um JY, et al: Arctigenin
inhibits lung metastasis of colorectal cancer by regulating cell
viability and metastatic phenotypes. Molecules. 21:E11352016.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Huang T, Xiao Y, Yi L, Li L, Wang M, Tian
C, Ma H, He K, Wang Y, Han B, et al: Coptisine from rhizoma
coptidis suppresses hct-116 cells-related tumor growth in vitro and
in vivo. Sci Rep. 7:385242017. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Lu Z and Xu S: ERK1/2 MAP kinases in cell
survival and apoptosis. IUBMB Life. 58:621–631. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Grossi V, Peserico A, Tezil T and Simone
C: p38α MAPK pathway: A key factor in colorectal cancer therapy and
chemoresistance. World J Gastroenterol. 20:9744–9758. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Sui X, Kong N, Ye L, Han W, Zhou J, Zhang
Q, He C and Pan H: p38 and JNK MAPK pathways control the balance of
apoptosis and autophagy in response to chemotherapeutic agents.
Cancer Lett. 344:174–179. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Chen L, Mayer JA, Krisko TI, Speers CW,
Wang T, Hilsenbeck SG and Brown PH: Inhibition of the p38 kinase
suppresses the proliferation of human ER-negative breast cancer
cells. Cancer Res. 69:8853–8861. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Thornton TM and Rincon M: Non-classical
p38 map kinase functions: Cell cycle checkpoints and survival. Int
J Biol Sci. 5:44–51. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Thiery JP and Sleeman JP: Complex networks
orchestrate epithelial-mesenchymal transitions. Nat Rev Mol Cell
Biol. 7:131–142. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Lindsey S and Langhans SA: Crosstalk of
oncogenic signaling pathways during epithelial-mesenchymal
transition. Front Oncol. 4:3582014. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Derycke LD and Bracke ME: N-cadherin in
the spotlight of cell-cell adhesion, differentiation,
embryogenesis, invasion and signalling. Int J Dev Biol. 48:463–476.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Gialeli C, Theocharis AD and Karamanos NK:
Roles of matrix metalloproteinases in cancer progression and their
pharmacological targeting. FEBS J. 278:16–27. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Urueña C, Mancipe J, Hernandez J,
Castañeda D, Pombo L, Gomez A, Asea A and Fiorentino S:
Gallotannin-rich Caesalpinia spinosa fraction decreases the primary
tumor and factors associated with poor prognosis in a murine breast
cancer model. BMC Complement. Altern Med. 13:742013.
|
|
52
|
Zhao T, Sun Q, del Rincon SV, Lovato A,
Marques M and Witcher M: Gallotannin imposes S phase arrest in
breast cancer cells and suppresses the growth of triple-negative
tumors in vivo. PLoS One. 9:e928532014. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Al-Halabi R, Bou Chedid M, Abou Merhi R,
El-Hajj H, Zahr H, Schneider-Stock R, Bazarbachi A and
Gali-Muhtasib H: Gallotannin inhibits NFκB signaling and growth of
human colon cancer xenografts. Cancer Biol Ther. 12:59–68. 2011.
View Article : Google Scholar : PubMed/NCBI
|