Introduction
The most common primary malignant bone tumor is
osteosarcoma, and it is derived from primitive bone-forming
mesenchymal cells (1). There is an
association between lung metastasis and poor prognosis in
osteosarcoma, and it occurs in the initial stages of the disease
(2). Although recent innovations in
multi-method treatment, i.e., chemotherapy, surgery and
radiotherapy have improved prognosis, metastasis and recurrence
persist due to high rates of multi-drug resistance (3,4).
Therefore, novel, targeted treatment of metastasis in osteosarcoma
is urgently needed.
Cancer cells exhibit anoikis resistance when they
avoid anoikis, or programmed death in a foreign microenvironment
without extracellular matrix (ECM) adhesion (5). Similar to most tumor cells,
osteosarcoma cell metastasis is complex and includes the absence of
ECM attachment, cell migration, anoikis resistance, angiogenesis
and metastatic colonization (6). By
regulating cell homeostasis in tissues, anoikis plays a critical
role in cellular physiology; it is a type of apoptosis induced by
separating cells from their surrounding matrix and neighboring
cells (7). A crucial hallmark of
malignant tumor metastasis and progression is anchorage-independent
(AI) growth; the potential for metastasis and progression in
prostate and breast cancer, as well as melanoma, and lung cancer,
is associated with AI growth (8,9). Tumor
cells can survive in the circulatory or lymphatic system due to
resistance to anoikis, and it enables the formation of metastasis
in distant organs (10).
In recent years, the function of angiogenesis in the
invasion, growth and metastasis of tumors has attracted much
attention from the scientific community (11). Angiogenesis is a fundamental process
required for physiological and pathological processes, referring to
new blood vessel formation from the vasculature already present
(12). An important angiogenic
factor, vascular endothelial growth factor A (VEGF-A) plays a
critical role in tumor progression and prognosis (13,14).
There is an association between VEGF-A expression and the
development of lung metastases in osteosarcoma (15,16).
Neovascularization is necessary for anoikis-resistant cells
adherence to distant sites and formation of metastases. Therefore,
elucidating the molecular mechanism of angiogenesis via VEGF-A in
anoikis-resistant human osteosarcoma is vital.
Tyrosine kinase Src is an important tumor biological
factor (17). Yes, Fyn, Lck, Lyn,
Hck, Blk, Yrk, Fgr and c-Src are among the Src family kinases; it
is one of the most important cellular pathways responsible for
proliferation, apoptosis, migration, and metastasis in
osteosarcoma, chondrosarcoma and Ewing sarcoma (18). Several studies have reported that
Src kinase activation is frequently detected in a variety of
anoikis-resistant tumor cells, and there is an association between
its activation and angiogenesis through increased VEGF-A and
interleukin 8 (IL-8) expression, which are proangiogenic factors
(19,20). Previously, we found that Src kinase
plays important roles in apoptosis, metastasis, anoikis resistance,
autophagy and stem cell self-renewal of osteosarcoma (21–23).
However, it remains unclear whether Src kinase activation promotes
the production of VEGF-A to enhance angiogenesis and metastasis of
tumors in anoikis-resistant human osteosarcoma cells.
The mitogen-activated protein kinase (MAPK) pathway
is a downstream molecule of Src kinase that plays a key regulatory
role in VEGF-A expression (24,25).
The MAPK [JNK (Jun amino-terminal kinase)/ERK
(extracellular-signal-regulated kinase)/p38] pathway is another
pivotal signaling pathway that contributes to tumor cell anoikis
resistance. Nevertheless, whether the MAPK pathway is involved in
the anoikis-resistant human osteosarcoma angiogenesis cascade
requires further clarification.
In the present study, we investigated the role of
human osteosarcoma cell resistance to anoikis in angiogenesis and
aimed to uncover the underlying mechanisms. We revealed that
angiogenesis was enhanced in such cells through control of the
expression of VEGF-A by anoikis via the Src/JNK/ERK signaling
pathways. The findings revealed that Src kinase is a potential
target in anti-angiogenesis therapy of osteosarcoma.
Materials and methods
Cell culture
The U2OS human osteosarcoma cell line was obtained
from the American Type Culture Collection (ATCC; Manassas, VA,
USA). The malignant transformed hFOB1.19 cell line (MTH) was
established by treatment with an initiating agent,
N-methyl-N′-nitro-N-nitrosoguanidine (MNNG,
1.0 µg/ml) and a promoter,
12-O-tetradecanoylphorbol-13-acetate (TPA, 200 ng/ml), as
previously reported (26). Human
umbilical vein endothelial cells (HUVECs) were obtained from
Southwest Hospital, Third Military Medical University (TMMU)
(Chongqing, China). The MTH and U2OS cells were cultured in
Dulbecco's modified Eagle's medium (DMEM; HyClone; GE Healthcare
Life Sciences, Logan, UT, USA); HUVECs were cultured in RPMI-1640
medium (HyClone; GE Healthcare Life Sciences) in 5% CO2
and 95% humidified air at 37°C. Fetal bovine serum (FBS, 10%;
Biological Industries, Beit Haemek Ltd., Beit HaEmek, Israel) and
1% penicillin-streptomycin (HyClone; GE Healthcare Life Sciences)
were used to supplement the cultures.
AI culture
To establish the anoikis-resistant model, we
cultured the MTH and U2OS cells in ultra-low attachment 6-well
plates (Corning Inc., Corning, NY, USA) for 3 days, transferred to
normal plates with attachment culture until they formed monolayers,
cultured again in ultra-low attachment 6-well plates for 14 days,
and finally transferred to normal plates; the re-adherent cells
were considered anoikis-resistant cells, and were designated MTHar
and U2OSar, respectively (21). An
inverted light microscope (Olympus Corp., Tokyo, Japan) was used to
observe the cell morphology.
Flow cytometry for apoptosis
We cultured the MTH, U2OS, MTHar and U2OSar cells in
ultra-low attachment 6-well plates for 3 days before detection.
Then, 1×106 cells were harvested and an Annexin
V-fluorescein isothiocyanate (FITC)/propidium iodide (PI) Apoptosis
Detection Kit (BD Biosciences, Franklin Lakes, NJ, USA) was used to
detect the cells. We analyzed the apoptotic cell percentage using a
flow cytometer (BD Biosciences) after staining and repeated the
experiment independently 3 times.
Preparation of conditioned medium
(CM)
Complete medium (DMEM, 10% FBS, 1%
penicillin-streptomycin) was used to culture the MTH, U2OS, MTHar
and U2OSar cells, or the cells were pretreated with 10 µmol/l Src
inhibitor PP2 (Selleckchem, Houston, TX, USA) for 24 h.
Phosphate-buffered saline (PBS) was used to wash the cells three
times and the cells were transferred to serum-free medium when the
cell density was ~70–80%. Then, the CM was collected 24 h after we
had changed the medium; we stored the CM at −80°C until use.
Wound healing assay
We seeded the HUVECs in 6-well plates and cultured
in complete medium until they reached approximately 90% confluence.
A 10-µl pipette tip was used to create wounds, and the HUVECs were
treated with CM. We monitored cell migration to the wound at 0 and
20 h using an inverted light microscope, and quantified it by
measuring the distance between the start point of the scratch wound
to the migrated point, and the experiment was repeated thrice.
Cell proliferation assay
We examined the cell proliferative potential using
Cell Counting Kit-8 (CCK-8; Dojindo Laboratories, Kumamoto, Japan).
We seeded 1×103 HUVECs in 96-well plates; the cells were
treated with CM. Following 1 to 5 days of culture, we added 10
µl/well CCK-8 reagent to each plate and cultured the cells for 1 h.
A microplate reader (Bio-Rad Laboratories, Hercules, CA, USA) was
used to measure the absorbance at 450 nm.
Tube formation assay
We dissolved Matrigel (BD Biosciences) at 4°C
overnight. We coated a pre-chilled 96-well plate with thawed
Matrigel (50 µl), and incubated it at 37°C for 30 min. After the
gel had solidified, 1×104 HUVECs/well were suspended in
culture medium comprised of 50% complete medium and 50% CM and
seeded in the wells. After 4–12 h of incubation at 37°C, we
photographed tube formation at 6 h using an inverted microscope. A
capillary-like tube formation assay is often used to determine
angiogenesis in vitro. Tube branches and total tube length
were measured with ImageJ software (http://rsb.info.nih.gov/ij/). Their number or total
length represents the capacity of angiogenesis (27). In this study, the number of tube
branches was used for statistical analysis.
Enzyme-linked immunosorbent assay
(ELISA)
We cultured the MTH, U2OS, MTHar, and U2OSar cells
in 6-well plates. We changed confluent cells to serum-free medium
or pretreated them with PP2 (10 µmol/l). After 24 h, we detected
supernatant VEGF-A levels using a human VEGF-A ELISA kit
(Neobioscience, Shenzhen, China).
Quantitative real-time PCR
We extracted total RNA from the osteosarcoma cells
with TRIzol (Takara Bio Inc., Shiga, Japan). We reverse-transcribed
RNA into complementary DNA (cDNA) using a QuantScript kit for
reverse transcription (RT) (Takara Bio Inc.). We performed RT-PCR
with SYBR Green using primers based on human VEGF-A and
GAPDH (glyceraldehyde-3-phosphate dehydrogenase) sequences.
The primer sequences were: VEGF-A forward,
5′-CTTCGCTTACTCTCACCTGCTTCTG-3′ and reverse,
5′-GCTGCTTCTTCCAACAATGTGTCTC-3′; GAPDH forward,
5′-CTTTGGTATCGTGGAAGGACTC-3′ and reverse,
5′-GTAGAGGCAGGGATGATGTTCT-3′. The cycling conditions were
polymerase activation at 95°C for 30 sec, and then 40 cycles at
95°C for 15 sec and 60°C for 60 sec. We performed triplicate
quantitative PCR (qPCR) with StepOnePlus (Applied Biosystems,
Foster City, CA, USA). The levels of the experiment group mRNA were
calculated with the 2−ΔΔCq method (28).
Western blotting
We performed western blotting as previously
described (21). Proteins were
extracted using RIPA buffer (cat. no. 89900; Thermo Fisher
Scientific, Inc.) and their concentrations were measured by BCA
Protein Assay kit (cat. no. P0010S; Beyotime Institute of
Biotechnology, Haimen, China). Proteins (40 µg/lane) were separated
by 10% SDS-PAGE (cat. no. P0012A; Beyotime Institute of
Biotechnology) and transferred to polyvinylidene difluoride (PVDF)
membranes (cat. no. 3010040001; Roche, Shanghai, China). Skim milk
(5%) was applied to block the membranes for 1 h at room
temperature. Then, the membranes were incubated with primary
antibodies overnight at 4°C. After 3 washes with PBST (0.05%
Tween-20 in PBS), the blots were incubated with the corresponding
secondary antibodies for 1 h at room temperature. We used the
following primary antibodies: Rabbit anti-phosphorylated (p)-Src
(dilution 1:1,000; cat. no. 2105) and Src (dilution 1:1,000; cat.
no. 2109; both from Cell Signaling Technology, Inc., Danvers, MA,
USA); mouse anti-VEGF (dilution 1:200; cat. no. sc-7269; Santa Cruz
Biotechnology, Santa Cruz, CA, USA); rabbit anti-JNK (dilution
1:1,000; cat. no. AJ518), rabbit anti-ERK (dilution 1:1,000; cat.
no. AM076), rabbit anti-p-ERK (dilution 1:1,000; cat. no. AF1891),
rabbit anti-p-JNK (dilution 1:1,000; cat. no. AF1762), mouse
anti-p38 (dilution 1:1,000; cat. no. AM065) and mouse anti-p-p38
(dilution 1:1,000; cat. no. AM063; all from Beyotime Institute of
Biotechnology). The secondary antibodies were horseradish
peroxidase (HRP)-conjugated goat anti-rabbit immunoglobulin (IgG)
(dilution 1:5,000; cat. no. BA1056) and HRP-conjugated goat
anti-mouse IgG (dilution 1:5,000; cat. no. BA1050; both from
Biodragon Immunotech, Beijing, China). GAPDH (dilution 1:1,000;
cat. no. 5174; Cell Signaling Technology, Inc.) was used as a
loading control. Finally, the blots were visualized using an ECL
kit (cat. no. P0018FFT; BeyoECL Moon; Beyotime Institute of
Biotechnology) and quantitative data were obtained using ImageJ
software (http://rsb.info.nih.gov/ij/).
Human osteosarcoma specimen
preparation
We collected specimens from 26 patients (13 males
and 13 females, aged from 8 to 49 years old, average age,
19.31±9.30 years) who had been diagnosed with osteosarcoma before
radiation therapy or chemotherapy from the Southwest Hospital, TMMU
from January 2011 to April 2014 (Table
I). We had obtained informed consent previously from the
patients or their guardians according to the standards set by the
Declaration of Helsinki. The TMMU Institutional Ethical Committee
approved the present study.
 | Table I.Correlations of the
clinicopathological features with p-Src and VEGF-A expression in
patients with osteosarcoma. |
Table I.
Correlations of the
clinicopathological features with p-Src and VEGF-A expression in
patients with osteosarcoma.
|
|
|
| p-Src |
| VEGF-A |
|
|---|
|
|
|
|
|
|
|
|
|---|
| Clinical
characteristics | Group | n | Positive | Negative |
P-valuea | Positive | Negative |
P-valuea |
|---|
| Sex | Male | 13 | 6 | 7 | 1 | 7 | 6 | 0.688 |
|
| Female | 13 | 7 | 6 |
| 9 | 4 |
|
| Age (years) | <20 | 19 | 8 | 11 | 0.378 | 10 | 9 | 0.19 |
|
| ≥20 | 7 | 5 | 2 |
| 6 | 1 |
|
| Tumor location | Femur | 14 | 5 | 9 | 0.215 | 7 | 7 | 0.397 |
|
| Tibia | 10 | 6 | 4 |
| 7 | 3 |
|
|
| Other | 2 | 2 | 0 |
| 2 | 0 |
|
| Histological
type | Osteoblastic | 12 | 7 | 5 | 0.876 | 10 | 2 | 0.185 |
|
| Chondroblastic | 5 | 2 | 3 |
| 2 | 3 |
|
|
| Fibroblastic | 2 | 1 | 1 |
| 1 | 1 |
|
|
| Other | 7 | 3 | 4 |
| 3 | 4 |
|
| Enneking stage | IIA | 3 | 0 | 3 | 0.029 | 1 | 2 | 0.05 |
|
| IIB | 13 | 5 | 8 |
| 6 | 7 |
|
|
| III | 10 | 8 | 2 |
| 9 | 1 |
|
| Tumor size
(cm) | <8 | 15 | 9 | 6 | 0.428 | 11 | 4 | 0.228 |
|
| ≥8 | 11 | 4 | 7 |
| 5 | 6 |
|
| Recurrence | Yes | 8 | 5 | 3 | 0.673 | 6 | 2 | 0.42 |
|
| No | 18 | 8 | 10 |
| 10 | 8 |
|
| Metastasis | Yes | 10 | 8 | 2 | 0.041 | 9 | 1 | 0.037 |
|
| No | 16 | 5 | 11 |
| 7 | 9 |
|
Immunohistochemistry (IHC)
analysis
p-Src, VEGF, CD34 and CD31 expression were
determined with a kit (ZSGB-BIO, Beijing, China) as previously
described (21). Two pathologists
blinded to the pathological and clinical data examined the
specimens under a light microscope. We used primary rabbit
anti-p-Src (dilution 1:200; cat. no. 2105; Cell Signaling
Technology, Inc.), mouse anti-VEGF (dilution 1:50; cat. no.
sc-7269; Santa Cruz Biotechnology, Inc.) and mouse anti-CD31
(dilution 1:100; cat. no. ZM-0044; ZSGB-BIO, Beijing, China) for
human osteosarcoma specimens. For mouse xenograft specimens,
primary rabbit anti-p-Src (dilution 1:200; cat. no. 2105; Cell
Signaling Technology, Inc.), mouse anti-VEGF (dilution 1:50; cat.
no. sc-7269; Santa Cruz Biotechnology, Inc.) and rabbit anti-CD34
(dilution 1:100; cat. no. Ab81289; Abcam, Cambridge, MA, USA) were
used. Semi-quantitative estimation was calculated to interpret the
IHC results based on the stained cells per 100 cells (percentage)
in >5 randomly selected fields under high-power microscopy
(magnification, ×400). We calculated the IHC scores by multiplying
the intensity of staining (0, no staining; 1, stained yellow; 2,
stained brown; and 3, stained dark brown) and staining extent
scores, i.e., percentage of positive tumor cells (0, 0–5%; 1,
6–25%; 2, 26–50%; 3, 51–75%; 4, >75%). Protein expression was
categorized using the final IHC scores as follows: 0 (−); 1–3 (+);
4–7 (++); and 8–12 (+++); positive expression was denoted by scores
≥4. We assessed microvessel density (MVD) using a CD31 antibody. We
scanned the immunostained sections at a low magnification (×40) to
detect hotspots, i.e., areas with the greatest vascular intensity.
Then, the number of microvessels were counted in 5
randomly-selected fields per hotspot under a high-power field
(magnification, ×200). The average number of microvessels in each
sample was determined and was defined as the MVD.
Mouse xenograft assay
We performed the animal experiments adhering to the
Institutional Animal Care and Use Committee-approved protocols at
Xinqiao Hospital, TMMU, according to the Directive 2010/63/EU.
Eight female 4-week-old nude mice were obtained from the Laboratory
Animal Centre of Xinqiao Hospital, TMMU (Chongqing, China). We
randomly divided mice into two groups (n=4 mice in each group) and
the average weight was ~15 g. Housing conditions included
temperature (22±2°C), humidity (40–60%), 12-h light/dark cycle and
fed ad libitum. We resuspended MTH/MTHar cells
(1×106 for each mouse) in serum-free medium, and the
cells were subcutaneously injected into each mouse. The mice were
sacrificed by cervical dislocation after 3 weeks and the harvested
tumors were fixed with formalin. The maximal volumes of xenografts
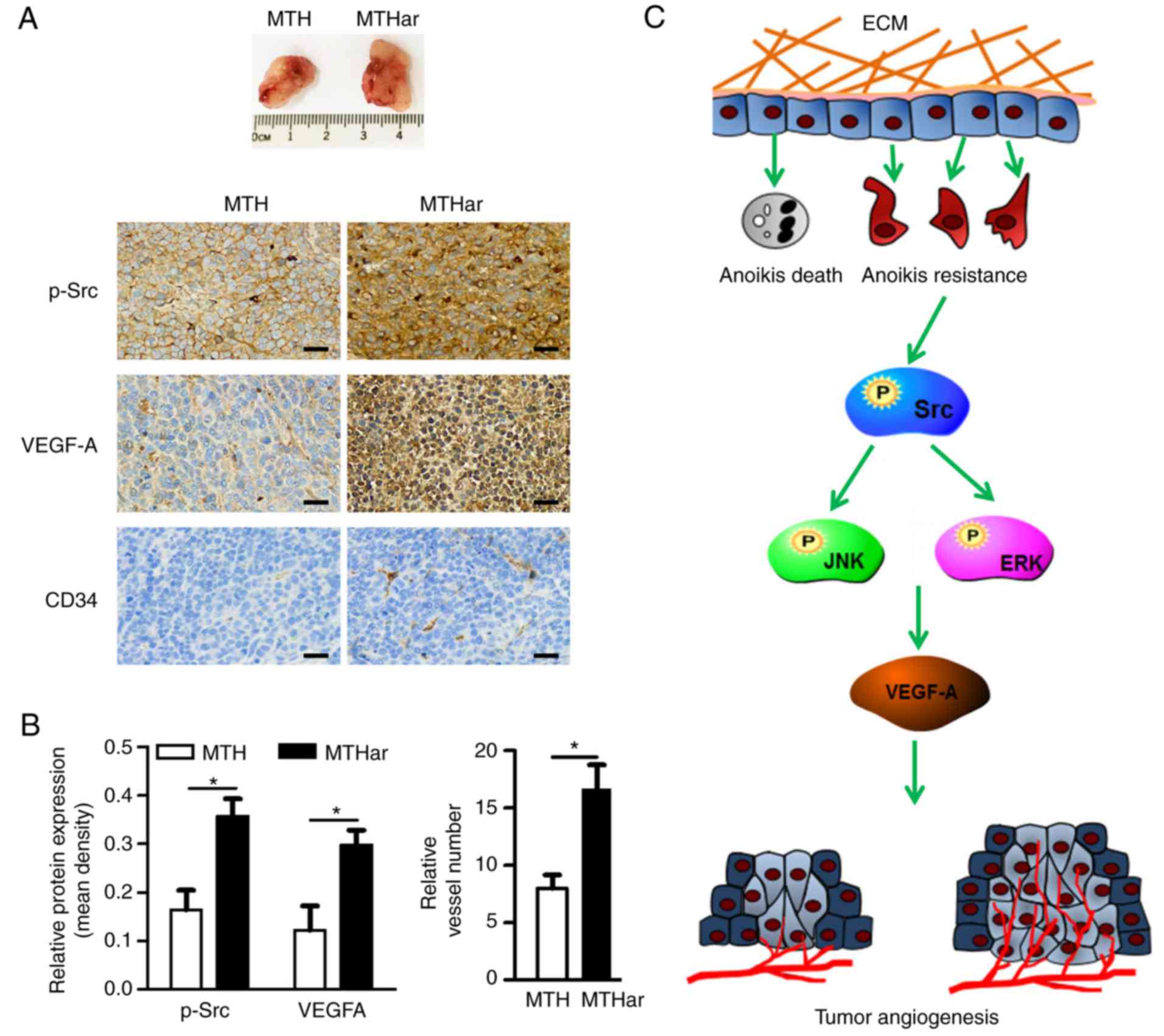
were 1,058 mm3 (MTH) and 1,573 mm3 (MTHar).
p-Src, VEGF-A and CD34 expression were determined by IHC.
Statistical analysis
We performed all statistical analyses using SPSS
19.0 (version 19.0; IBM Corp., Armonk, NY, USA). We reported the
data as the mean ± standard deviation (SD). We analyzed
between-group differences with Student's t-test of variance,
Spearman's rank correlation coefficients or Pearson's χ2
test. P<0.05 was considered to indicate a statistically
significant difference.
Results
Establishment of anoikis-resistant
model
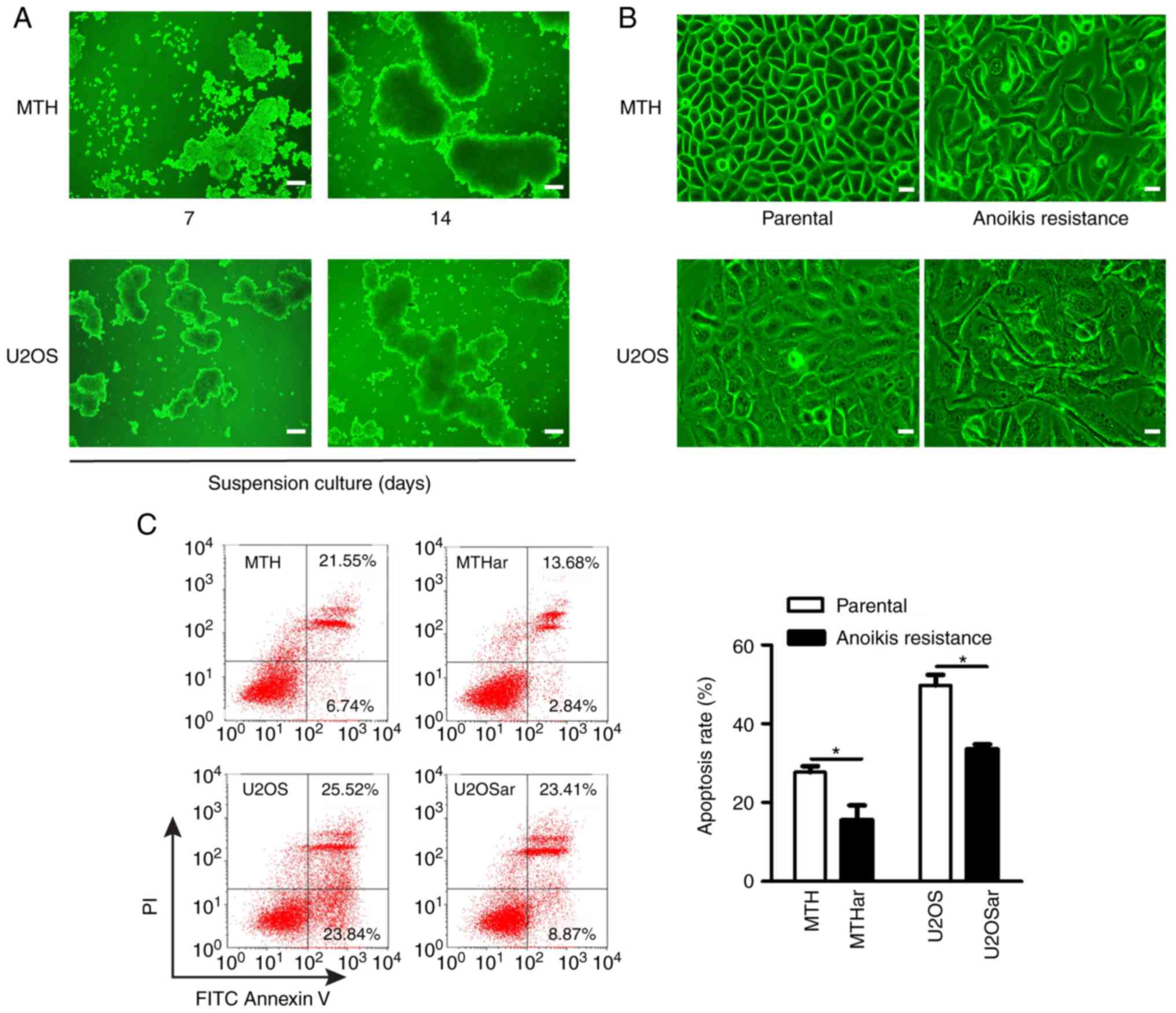
The anoikis-resistant human osteosarcoma cell model
was established by cell suspension culture according to a previous
study (21). The osteosarcoma cells
were round and gathered into clusters gradually in the suspension
culture. The clusters grew bigger and tighter over time (Fig. 1A). The regular polygonal
osteosarcoma cells became irregular, elongated and spindle-shaped
when they were transferred into normal plates and these cells (the
re-adherent cells) were considered anoikis-resistant cells
(Fig. 1B). Flow cytometry
demonstrated significantly less apoptosis in the anoikis-resistant
osteosarcoma cells under suspension conditions compared with the
parental osteosarcoma cells (Fig.
1C).
Anoikis-resistant human osteosarcoma
cells promote angiogenesis by controlling the expression of
VEGF-A
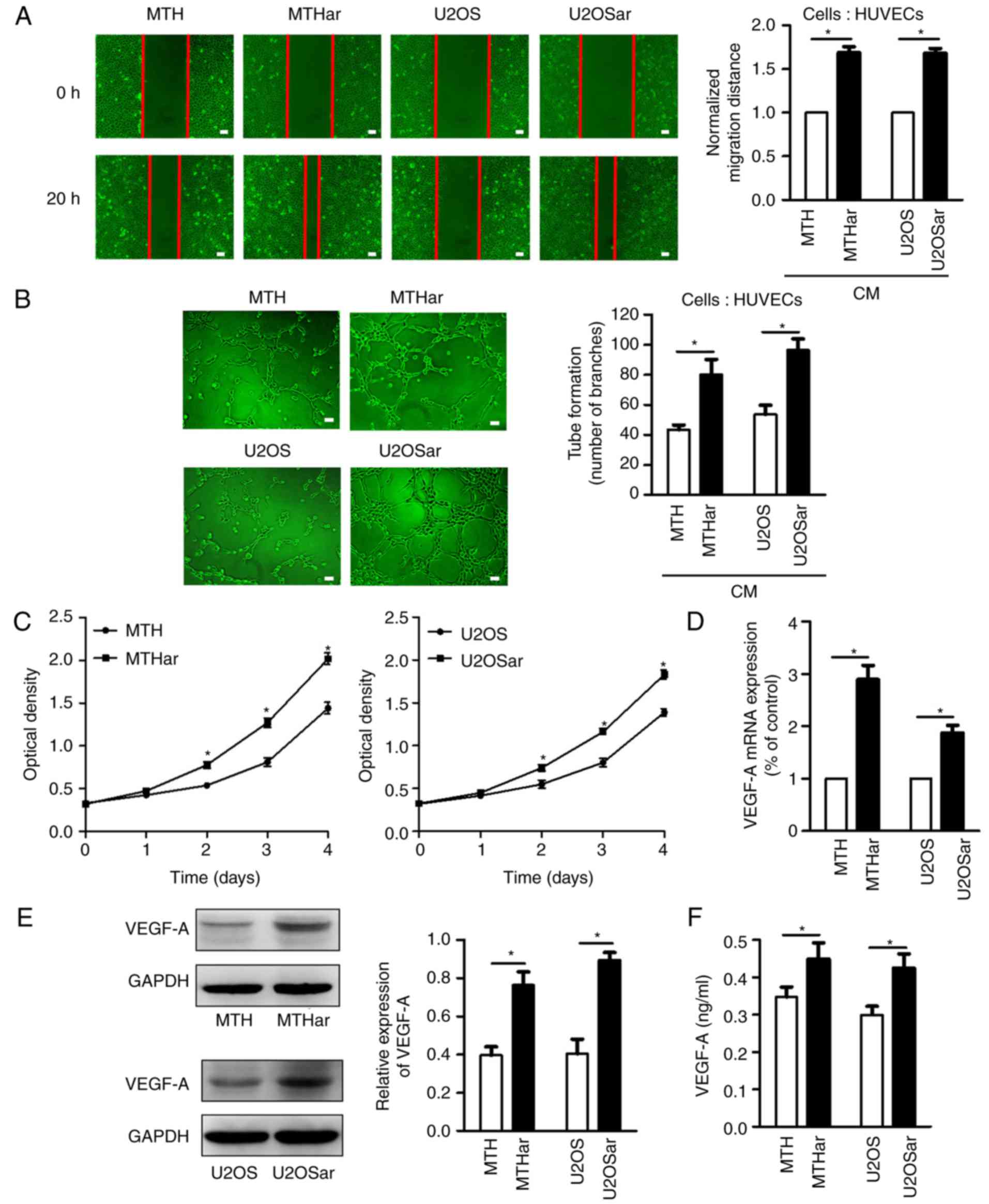
Tumor metastasis requires angiogenesis, a critical
factor in controlling cancer progression (27,29).
Anoikis-resistant cells play a key role in metastasis; however, the
effects of resistance to anoikis in human osteosarcoma cells on
angiogenesis remain largely unknown. Angiogenesis includes HUVEC
migration, proliferation, and tube formation for the formation of
new blood vessels (30). Therefore,
we investigated the effects of resistance to anoikis in
osteosarcoma cells from humans on HUVEC angiogenesis using in
vitro migration, tube formation and proliferation assays. CM
from anoikis-resistant osteosarcoma cells promoted HUVEC migration,
tube formation and proliferation (Fig.
2A-C). RT-PCR, western blotting and ELISA revealed increased
mRNA and protein expression of VEGF-A in the anoikis-resistant
osteosarcoma cells (Fig. 2D-F).
These data demonstrated that osteosarcoma cells that were resistant
to anoikis had increased expression of VEGF-A and angiogenesis.
Src inhibitor reduces the expression
of VEGF-A and angiogenesis and inhibits JNK and ERK pathway
activity
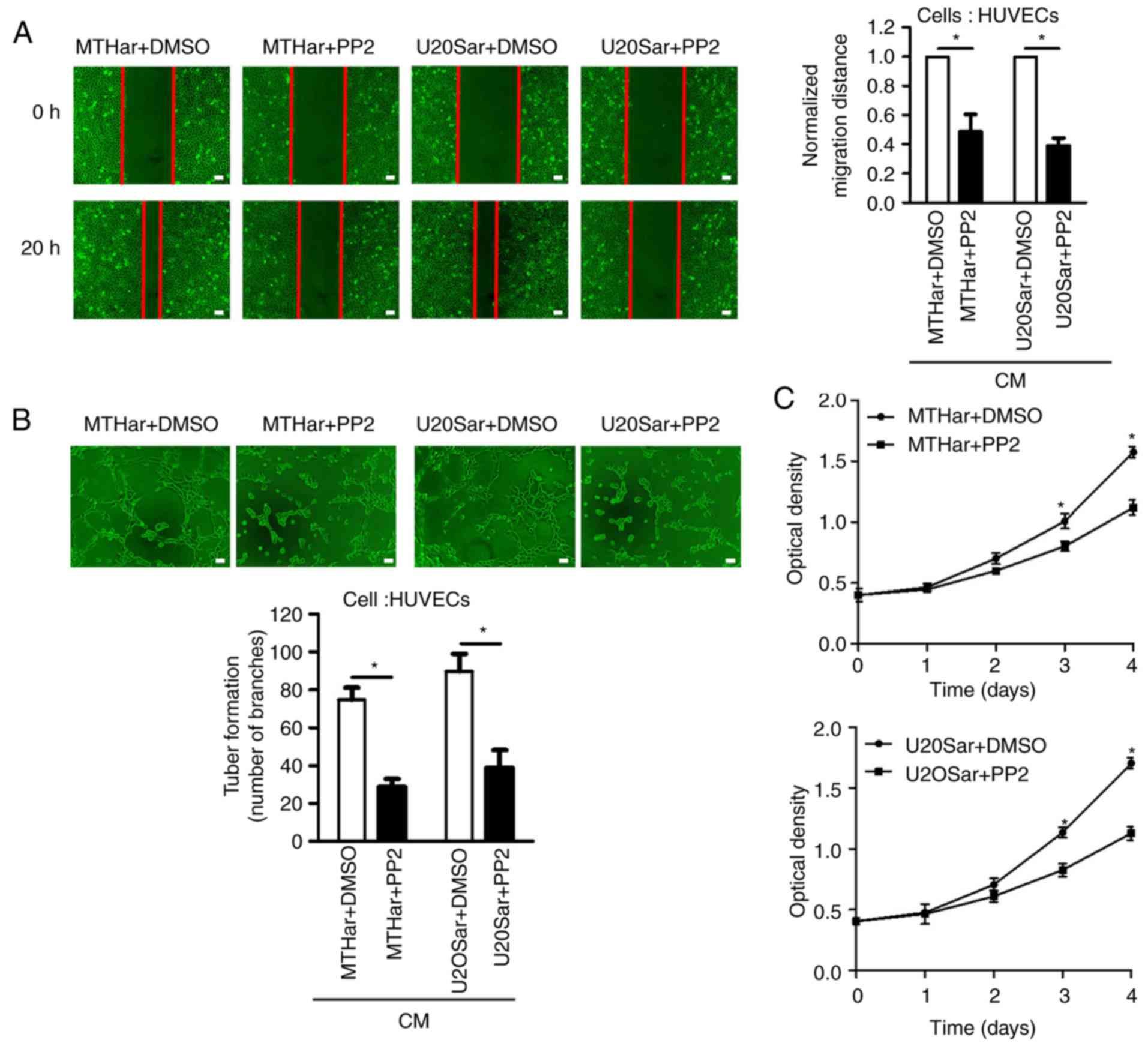
Src kinase activationis frequently detected in a
variety of anoikis-resistant tumor cells as demonstrated in
Fig. 3G. Recent research has
focused on kinases directly modulating the apoptosis machinery in
anoikis resistance. However, the relationship between p-Src and
VEGF-A has been poorly explored in anoikis-resistant osteosarcoma
cells in humans. To verify whether the expression of VEGF-A
involved Src kinase activation in anoikis-resistant osteosarcoma
cells, we pretreated anoikis-resistant cells with an Src inhibitor
(PP2) for 24 h. PP2 inhibited the expression of p-Src and VEGF-A in
the osteosarcoma cells resistant to anoikis (Fig. 3D-G). Compared with the control
group, PP2 also reduced HUVEC proliferation, migration and tube
formation (Fig. 3A-C). In various
cancers, the MAPK (JNK/ERK/p38) signaling pathway is a downstream
molecule of Src kinase (24).
Accordingly, we investigated whether MAPK (JNK/ERK/P38) signaling
pathway molecules were involved in the expression of VEGF-A induced
by p-Src in anoikis-resistant osteosarcoma cells. We investigated
JNK and ERK phosphorylation in anoikis-resistant cells and p-p38
was not altered; PP2 decreased JNK and ERK phosphorylation in
anoikis-resistant cells (Fig. 3G).
These results revealed that JNK/ERK may be an Src target downstream
in controlling VEGF-A expression. Altogether, our findings
indicated that Src kinase activation induced the expression of
VEGF-A and angiogenesis via the JNK/ERK pathway activation in
anoikis-resistant osteosarcoma cells.
VEGF-A, Src and MVD clinical
importance in osteosarcoma
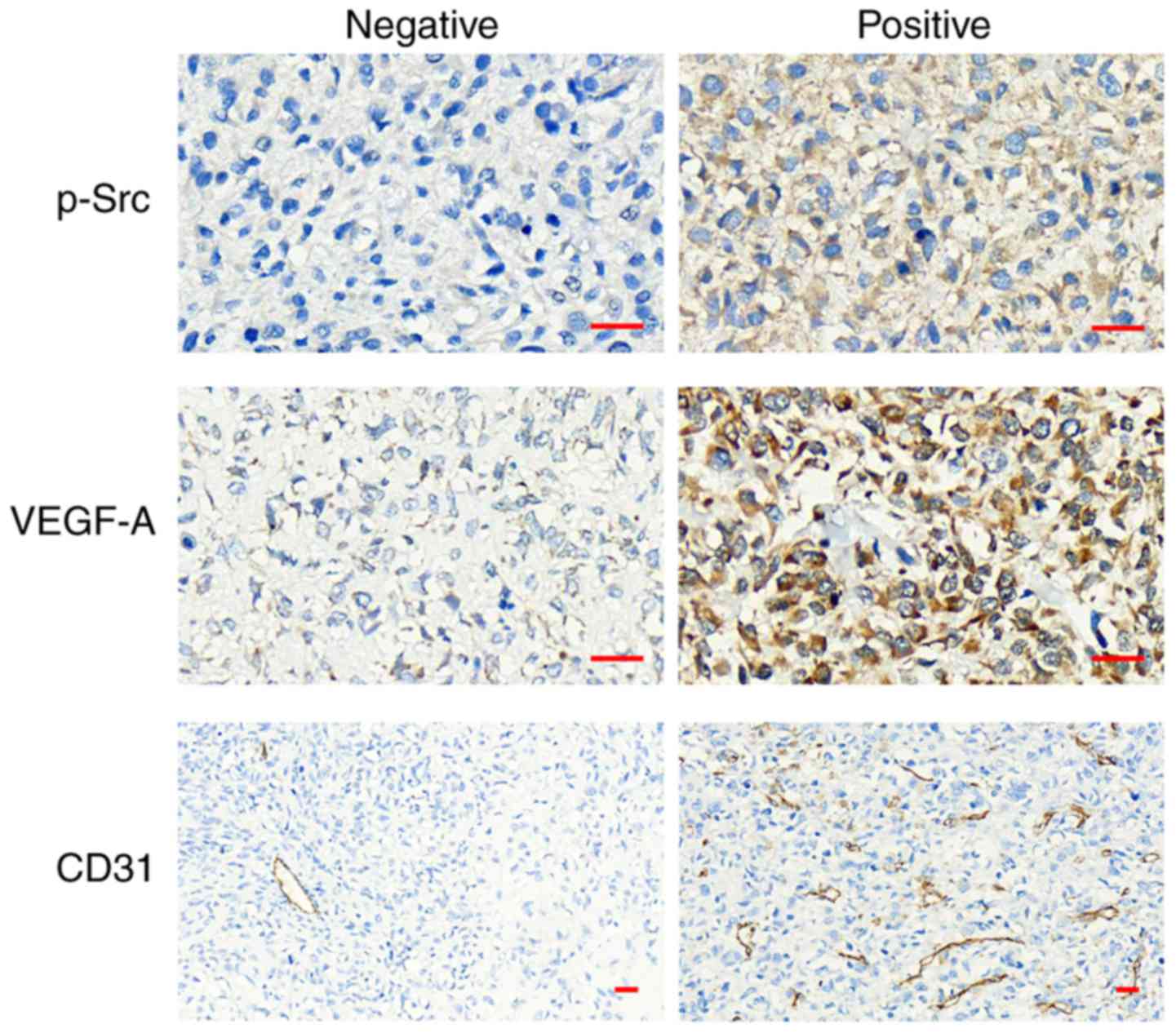
We quantified the expression of P-Src and VEGF-A in
osteosarcoma tissues from humans using IHC staining, and the areas
of positive expression were mainly localized in the (Fig. 4). To elucidate their clinical
significance, we assessed the correlation between P-Src and VEGF-A
expression and the patients' available clinicopathological
parameters (Table I). Both P-Src
and VEGF-A expression was significantly associated with lung
metastasis, and there was an association between P-Src expression
and Enneking stage, but not with age, sex, tumor size, pathological
type and recurrence. The positive rate of P-Src and VEGF-A
expression was 50% (13/26) and 61.5% (16/26), respectively, in the
osteosarcoma samples (Table I). The
relationship between P-Src and VEGF-A expression was calculated and
showed that high P-Src expression correlated with expression of
VEGF-A (r=0.474; P=0.014; Table
II). To assess the P-Src-angiogenesis association, we detected
the MVD via CD31 IHC staining in osteosarcoma tissues (Fig. 4). MVD was significantly higher in
tumors with positive expression of P-Src (r=0.545; P=0.004;
Table II). Taken together, these
data demonstrate that P-Src and VEGF-A may be important clinical
markers in human osteosarcoma with lung metastasis, and Src kinase
may promote angiogenesis via VEGF-A expression in osteosarcoma
development.
| Table II.Associations between p-Src expression
and VEGF-A expression and MVD in patients with osteosarcoma. |
Table II.
Associations between p-Src expression
and VEGF-A expression and MVD in patients with osteosarcoma.
|
|
| VEGF-A | MVD |
|---|
|
|
|
|
|
|---|
| p-Src | n | Positive | Negative | High | Low |
|---|
| Positive | 13 | 11 | 2 | 11 | 2 |
| Negative | 13 | 5 | 8 | 4 | 9 |
| r |
| 0.474 |
| 0.545 |
|
| P-value |
| 0.014a |
| 0.004a |
|
Resistance to anoikis in osteosarcoma
cells from humans promotes angiogenesis in a mouse xenograft
model
We found that VEGF-A expression and HUVEC
proliferation, migration, and tube formation were enhanced in
anoikis-resistant osteosarcoma cells via Src kinase activation
in vitro. To verify our experimental results, a xenograft
model in mice was used to assess the association between anoikis
resistance and angiogenesis. p-Src, VEGF-A and CD34 expression in
the xenograft tumors formed in the nude mice were detected using
IHC (Fig. 5A). p-Src and VEGF-A
expression was higher in anoikis-resistant cells, as was MVD
(Fig. 5B). Collectively, these data
indicated that anoikis-resistant osteosarcoma cells enhanced
osteosarcoma angiogenesis via p-Src and VEGF-A overexpression.
Discussion
Osteosarcoma is a highly malignant bone cancer whose
leading cause of death is metastasis, and treatment options are
unsatisfactory (31). Resistance to
anoikis is a vital aspect of metastasis in several human cancers,
including osteosarcoma, in which an indispensable role is played by
angiogenesis in its metastasis and growth (31,32).
Anti-angiogenesis enables the normalization of tumor blood vessels
to improve oxygen transport and enhance chemotherapy efficiency,
thereby prolonging survival (33).
Anti-angiogenesis is a potential osteosarcoma treatment target. In
the present study, we determined the effect of anoikis resistance
on the expression of VEGF-A in human osteosarcoma, i.e., it
subsequently increased angiogenesis. Anoikis-resistant
subpopulations of human osteosarcoma cells had upregulated VEGF-A
and increased angiogenesis via the Src/JNK/ERK signaling pathways
(Fig. 5C).
Due to the high pulmonary metastasis in
osteosarcoma, it is vital to elucidate the metastasis-controlling
mechanisms in order to improve poor prognosis. Metastasis involves
cancer cells migrating from primary sites, invading through the
basement membrane, surviving in the vascular system, and
proliferating in a distant target organ (7,10). A
barrier is presented when tumor cells typically detach from the
primary site before entering the bloodstream; apoptosis resulting
from the loss of cell connection to the ECM, or cell adhesion in an
inappropriate location, is termed anoikis (Greek for homelessness)
(34). The intrinsic or extrinsic
apoptosis pathway mediates anoikis initiation and execution, and it
likely prevents ectopic growth of cells at unsuitable sites.
Anoikis is simply a special type of apoptosis, there is no special
marker. Some studies have reported that the overexpression of cell
adhesion molecules CEACAM6 and CD44V6 were associated with cancer
cell mobility, poor overall survival and lung metastasis in
osteosarcoma (35,36). However, no studies have reported
that these molecules were regarded as the specific markers to
predict anoikis in osteosarcoma tissues. Anoikis suppression
following cancer cells disassociating from the ECM is an important
step in tumor malignancy and metastatic progression. Therefore,
acquiring the ability to resist anoikis is a critical step in these
metastatic cell subpopulations. Many investigations have indicated
the involvement of multiple molecular pathways in conferring
anoikis resistance in cancer cells, including activation of the
pathways of phosphatidylinositol 3-kinase (PI3K)/AKT, Src and
Wnt/β-catenin signaling, caveolin-1 downregulation, c-Met
activation, interaction of ezrin/β4 integrin and the Bcl family
(37,38). Previously, we determined that the
Src/AKT pathway played vital roles in osteosarcoma anoikis
resistance and metastasis (21).
Osteosarcoma is a highly malignant, blood
vessel-rich bone tumor, and the maintenance of tumor growth and
metastasis rely on angiogenesis (6,39).
Without the formation of a new vascular network, tumor cells would
not acquire the necessary oxygen and nutrients for tumor invasion,
metastasis and growth (6). The
pro-angiogenic and anti-angiogenic factor imbalance results in
tumor angiogenesis (40). In
particular, there is an important function of VEGF-A in tumor
angiogenesis, including in osteosarcoma.
Commonly, osteosarcoma has a tendency to develop
distant metastasis, particularly to the lungs, by blood
circulation, resulting in death. Angiogenesis is essential for
anoikis-resistant cells adherence to distant sites and
establishment of metastatic lesions. VEGF-A was revealed to be
upregulated in anoikis-resistant sublines in intestinal epithelial
cells and prostate cancer cells (32,41).
Research has reported VEGF-A upregulation in osteosarcoma and
positive correlation with poor prognosis and lung metastases
(15,16). In the present study, VEGF-A
expression was significantly higher in anoikis-resistant human
osteosarcoma cells in contrast to that of parental cells (Fig. 2D-F), subsequently promoting HUVEC
migration, tube formation and proliferation in vitro
(Fig. 2A-C). In addition, we
determined that VEGF-A expression was significantly associated with
lung metastasis in osteosarcoma specimens (Table I). Our findings indicated that human
osteosarcoma cells resistant to anoikis had significant
angiogenesis by increasing VEGF-A expression, and VEGF-A expression
may be an important clinical predictive marker of pulmonary
metastasis in osteosarcoma.
Src kinase activation is considered a prometastatic
pathway in several types of tumors, since vital oncogenic
mechanisms such as cell proliferation, resistance to apoptosis,
invasion and adhesion are involved in it (42). p-Src expression was revealed to be
high in osteosarcoma tissue (64.7%) and was significantly
associated with metastasis and clinical stage (43). Notably, Src kinase activation was
frequently detected in anoikis-resistant osteosarcoma cells, and
its activation was associated with angiogenesis through increased
proangiogenic factor expression, e.g., VEGF-A and IL-8 (19,21).
Using IHC, we determined a significant association between the
expression of p-Src and lung metastasis and Enneking stage in
osteosarcoma specimens (Table I).
Furthermore, the expression of p-Src and VEGF-A was positively
correlated (r=0.474; P=0.014; Table
II), and there was an association between positive expression
of p-Src and significantly higher MVD in vivo (r=0.545;
P=0.004; Table II). In the present
study, Src activation was detected in anoikis-resistant human
osteosarcoma cells, and the Src inhibitor reduced the expression of
VEGF-A and angiogenesis (Fig.
3A-F). We determined that Src kinase activation played a key
part in stimulating the expression of VEGF-A and angiogenesis in
anoikis-resistant osteosarcoma cells in humans, indicating that, in
osteosarcoma, Src kinase may be a novel treatment target against
angiogenesis and metastasis.
As a downstream molecule of Src kinase, the MAPK
cascade pathway is a key regulator of oncogenic characteristics
such as invasion, proliferation, apoptosis, angiogenesis and
metastasis in osteosarcoma, Ewing sarcoma and chondrosarcomas
(44). The MAPK pathway is
comprised of 3 families: JNK, ERK and p38/SAPK (stress-activated
protein kinase) (45). ERK and p38
signaling pathway activation was detected in ovarian cancer under
AI growth conditions (46). In this
study, JNK and ERK phosphorylation in anoikis-resistant
osteosarcoma cells was investigated; p38 was not activated in this
cancer type, and Src inhibitor inhibited JNK and ERK pathway
activity (Fig. 3G). p38 activation
is believed to regulate apoptosis (anoikis) and/or inflammation
(47). However, a number of studies
have shown that p38 plays an important role in the developmental
processes and progression of manysolid tumors (48,49).
These seemingly contradictory biological properties may beexplained
by the fact that genetic and epigenetic alterations involve cancer
onset and progression. The data indicated that the MAPK pathway may
be important in resistance to anoikis in osteosarcoma and that it
has an important function in the expression of VEGF-A.
In conclusion, VEGF-A expression and angiogenesis
were enhanced in anoikis-resistant subpopulations of osteosarcoma
cells from humans via the Src/JNK/ERK signaling pathways.
Therefore, Src kinase may be a new treatment target against
metastasis and angiogenesis in osteosarcoma. Our findings
demonstrated the underlying mechanisms by which resistance to
anoikis in osteosarcoma cells from humans promoted
angiogenesis.
Acknowledgements
We thank Ms. Ya-Li Wang and Mr. Qiu-Lin Tan (Xinqiao
Hospital, Army Medical University, Chongqing, China) for their
assistance with the IHC techniques.
Funding
The present study was supported by grants from the
National Natural Science Foundation Project of China (nos. 81672653
and 81372864) and the Chongqing Research Program of Basic Research
and Frontier Technology (no. cstc2015jcyjBX0067).
Availability of data and materials
The datasets used during the present study are
available from the corresponding author upon reasonable
request.
Authors' contributions
The conceptualization and the formal analysis was
conducted by QNG. ZG was responsible for the conceptualization,
formal analysis, data analysis and interpretation, the figure
preparation and was also involved in the writing. GSZ was involved
in the conceptualization, data analysis and interpretation, and
performed the mouse xenograft model experiments. YL was involved in
the conceptualization and data analysis. DP, HS and XT performed
the clinical sample collection, the IHC assay and the data
analysis. All authors read and approved the manuscript and agree to
be accountable for all aspects of the research in ensuring that the
accuracy or integrity of any part of the work are appropriately
investigated and resolved.
Ethics approval and consent to
participate
All experiments were approved by the Ethical
Committee of the Third Military Medical University (Army Medical
University, Chongqing, China).
Patient consent for publication
Not applicable.
Competing interests
The authors state that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
AI
|
anchorage-independent
|
|
ECM
|
extracellular matrix
|
|
ERK
|
extracellularsignal-regulated
kinase
|
|
HUVECs
|
human umbilical vein endothelial
cells
|
|
IL-8
|
interleukin 8
|
|
JNK
|
Jun amino-terminal kinase
|
|
MAPK
|
mitogen-activated protein kinase
|
|
MVD
|
microvessel density
|
|
VEGF-A
|
vascular endothelial growth
factor-A
|
References
|
1
|
Messerschmitt PJ, Garcia RM, Abdul-Karim
FW, Greenfield EM and Getty PJ: Osteosarcoma. J Am Acad Orthop
Surg. 17:515–527. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ottaviani G and Jaffe N: The etiology of
osteosarcoma. Cancer Treat Res. 152:15–32. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Allison DC, Carney SC, Ahlmann ER,
Hendifar A, Chawla S, Fedenko A, Angeles C and Menendez LR: A
meta-analysis of osteosarcoma outcomes in the modern medical era.
Sarcoma. 2012:7048722012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Chou AJ and Gorlick R: Chemotherapy
resistance in osteosarcoma: Current challenges and future
directions. Expert Rev Anticancer Ther. 6:1075–1085. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Paoli P, Giannoni E and Chiarugi P:
Anoikis molecular pathways and its role in cancer progression.
Biochim Biophys Acta. 1833:3481–3498. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhu L, McManus MM and Hughes DP:
Understanding the biology of bone sarcoma from early initiating
events through late events in metastasis and disease progression.
Front Oncol. 3:2302013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kim YN, Koo KH, Sung JY, Yun UJ and Kim H:
Anoikis resistance: An essential prerequisite for tumor metastasis.
Int J Cell Biol. 2012:3068792012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Guadamillas MC, Cerezo A and Del PMA:
Overcoming anoikis-pathways to anchorage-independent growth in
cancer. J Cell Sci. 124:3189–3197. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Mori S, Chang JT, Andrechek ER, Matsumura
N, Baba T, Yao G, Kim JW, Gatza M, Murphy S and Nevins JR:
Anchorage-independent cell growth signature identifies tumors with
metastatic potential. Oncogene. 28:2796–2805. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Strauss SJ, Ng T, Mendoza-Naranjo A,
Whelan J and Sorensen PH: Understanding micrometastatic disease and
Anoikis resistance in ewing family of tumors and osteosarcoma.
Oncologist. 15:627–635. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Giner F, Lopez-Guerrero JA, Machado I,
Garcia-Casado Z, Peydro-Olaya A and Llombart-Bosch A: The early
stages of tumor angiogenesis in human osteosarcoma: A nude mice
xenotransplant model. Virchows Arch. 467:193–201. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Walia A, Yang JF, Huang YH, Rosenblatt MI,
Chang JH and Azar DT: Endostatin's emerging roles in angiogenesis,
lymphangiogenesis, disease, and clinical applications. Biochim
Biophys Acta. 1850:2422–2438. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kerbel RS: Tumor angiogenesis: Past,
present and the near future. Carcinogenesis. 21:505–515. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Matsumoto K and Ema M: Roles of VEGF-A
signalling in development, regeneration, and tumours. J Biochem.
156:1–10. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kaya M, Wada T, Akatsuka T, Kawaguchi S,
Nagoya S, Shindoh M, Higashino F, Mezawa F, Okada F and Ishii S:
Vascular endothelial growth factor expression in untreated
osteosarcoma is predictive of pulmonary metastasis and poor
prognosis. Clin Cancer Res. 6:572–577. 2000.PubMed/NCBI
|
|
16
|
Hu F, Shang XF, Wang W, Jiang W, Fang C,
Tan D and Zhou HC: High-level expression of periostin is
significantly correlated with tumour angiogenesis and poor
prognosis in osteosarcoma. Int J Exp Pathol. 97:86–92. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chen Q, Zhou Z, Shan L, Zeng H, Hua Y and
Cai Z: The importance of Src signaling in sarcoma. Oncol Lett.
10:17–22. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yeatman TJ: A renaissance for SRC. Nat Rev
Cancer. 4:470–480. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
19
|
Aleshin A and Finn RS: S RC A century of
science brought to the clinic. Neoplasia. 12:599–607. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kanda S, Miyata Y, Kanetake H and
Smithgall TE: Non-receptor protein-tyrosine kinases as molecular
targets for antiangiogenic therapy (Review). Int J Mol Med.
20:113–121. 2007.PubMed/NCBI
|
|
21
|
Dai H, Lv YF, Yan GN, Meng G, Zhang X and
Guo QN: RanBP9/TSSC3 complex cooperates to suppress anoikis
resistance and metastasis via inhibiting Src-mediated Akt signaling
in osteosarcoma. Cell Death Dis. 7:e25722016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yan GN, Tang XF, Zhang XC, He T, Huang YS,
Zhang X, Meng G, Guo DY, Lv YF and Guo QN: TSSC3 represses
self-renewal of osteosarcoma stem cells and Nanog expression by
inhibiting the Src/Akt pathway. Oncotarget. 8:85628–85641. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhao GS, Gao ZR, Zhang Q, Tang XF, Lv YF,
Zhang ZS, Zhang Y, Tan QL, Peng DB, Jiang DM, et al: TSSC3 promotes
autophagy via inactivating the Src-mediated PI3K/Akt/mTOR pathway
to suppress tumorigenesis and metastasis in osteosarcoma, and
predicts a favorable prognosis. J Exp Clin Cancer Res. 37:1882018.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Cui S, Wang J, Wu Q, Qian J, Yang C and Bo
P: Genistein inhibits the growth and regulates the migration and
invasion abilities of melanoma cells via the FAK/paxillin and MAPK
pathways. Oncotarget. 8:21674–21691. 2017.PubMed/NCBI
|
|
25
|
Huang D, Ding Y, Luo WM, Bender S, Qian
CN, Kort E, Zhang ZF, VandenBeldt K, Duesbery NS, Resau JH, et al:
Inhibition of MAPK kinase signaling pathways suppressed renal cell
carcinoma growth and angiogenesis in vivo. Cancer Res. 68:81–88.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Li Y, Meng G and Guo QN: Changes in
genomic imprinting and gene expression associated with
transformation in a model of human osteosarcoma. Exp Mol Pathol.
84:234–239. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Tsai HC, Tzeng HE, Huang CY, Huang YL,
Tsai CH, Wang SW, Wang PC, Chang AC, Fong YC and Tang CH: WISP-1
positively regulates angiogenesis by controlling VEGF-A expression
in human osteosarcoma. Cell Death Dis. 8:e27502017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2−ΔΔCT method. Methods. 25:402–408. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Folkman J: Role of angiogenesis in tumor
growth and metastasis. Semin Oncol. 29:15–18. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Mund JA and Case J: The role of
circulating endothelial progenitor cells in tumor angiogenesis.
Curr Stem Cell Res Ther. 6:115–121. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhou W, Hao M, Du X, Chen K, Wang G and
Yang J: Advances in targeted therapy for osteosarcoma. Discov Med.
17:301–307. 2014.PubMed/NCBI
|
|
32
|
Zhang P, Chen L, Song Y, Li X, Sun Y, Xiao
Y and Xing Y: Tetraiodothyroacetic acid and transthyretin silencing
inhibit pro-metastatic effect of L-thyroxin in anoikis-resistant
prostate cancer cells through regulation of MAPK/ERK pathway. Exp
Cell Res. 347:350–359. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Jain RK: Antiangiogenesis strategies
revisited: From starving tumors to alleviating hypoxia. Cancer
Cell. 26:605–622. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Frisch SM and Screaton RA: Anoikis
mechanisms. Curr Opin Cell Biol. 13:555–562. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zhang Y, Ding C, Wang J, Sun G, Cao Y, Xu
L, Zhou L and Chen X: Prognostic significance of CD44V6 expression
in osteosarcoma: A meta-analysis. J Orthop Surg Res. 10:1872015.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wang Z, Luo C, Wang H, Yan X, Liu W and
Meng Z: CEACAM6 is associated with osteosarcoma metastasis and
facilitates epithelial-mesenchymal transition in osteosarcoma
cells. Onco Targets Ther. 11:3159–3166. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Cantiani L, Manara MC, Zucchini C, De
Sanctis P, Zuntini M, Valvassori L, Serra M, Olivero M, Di Renzo
MF, Colombo MP, et al: Caveolin-1 reduces osteosarcoma metastases
by inhibiting c-Src activity and met signaling. Cancer Res.
67:7675–7685. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Wan X, Kim SY, Guenther LM, Mendoza A,
Briggs J, Yeung C, Currier D, Zhang H, Mackall C, Li WJ, et al:
Beta4 integrin promotes osteosarcoma metastasis and interacts with
ezrin. Oncogene. 28:3401–3411. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Peng N, Gao S, Guo X, Wang G, Cheng C, Li
M and Liu K: Silencing of VEGF inhibits human osteosarcoma
angiogenesis and promotes cell apoptosis via VEGF/PI3K/AKT
signaling pathway. Am J Transl Res. 8:1005–1015. 2016.PubMed/NCBI
|
|
40
|
Carmeliet P and Jain RK: Molecular
mechanisms and clinical applications of angiogenesis. Nature.
473:298–307. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Rak J, Mitsuhashi Y, Sheehan C, Krestow
JK, Florenes VA, Filmus J and Kerbel RS: Collateral expression of
proangiogenic and tumorigenic properties in intestinal epithelial
cell variants selected for resistance to anoikis. Neoplasia.
1:23–30. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Summy JM and Gallick GE: Src family
kinases in tumor progression and metastasis. Cancer Metastasis Rev.
22:337–358. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Hu C, Deng Z, Zhang Y, Yan L, Cai L, Lei J
and Xie Y: The prognostic significance of Src and p-Src expression
in patients with osteosarcoma. Med Sci Monit. 21:638–645. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Chandhanayingyong C, Kim Y, Staples JR,
Hahn C and Lee FY: MAPK/ERK signaling in osteosarcomas, Ewing
sarcomas and chondrosarcomas: Therapeutic implications and future
directions. Sarcoma. 2012:4048102012. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Morrison DK: MAP kinase pathways. Cold
Spring Harb Perspect Biol. 4:a0112542012. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Carduner L, Picot CR, Leroy-Dudal J, Blay
L, Kellouche S and Carreiras F: Cell cycle arrest or survival
signaling through alphav integrins, activation of PKC and ERK1/2
lead to anoikis resistance of ovarian cancer spheroids. Exp Cell
Res. 320:329–342. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Walsh MF, Thamilselvan V, Grotelueschen R,
Farhana L and Basson M: Absence of adhesion triggers differential
FAK and SAPKp38 signals in SW620 human colon cancer cells that may
inhibit adhesiveness and lead to cell death. Cell Physiol Biochem.
13:135–146. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Leelahavanichkul K, Amornphimoltham P,
Molinolo AA, Basile JR, Koontongkaew S and Gutkind JS: A role for
p38 MAPK in head and neck cancer cell growth and tumor-induced
angiogenesis and lymphangiogenesis. Mol Oncol. 8:105–118. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Rodríguez-Berriguete G, Fraile B,
Martínez-Onsurbe P, Olmedilla G, Paniagua R and Royuela M: MAP
kinases and prostate cancer. J Signal Transduct.
2012:1691702012.PubMed/NCBI
|