Introduction
Breast cancer is the most frequently diagnosed
cancer and the leading cause of cancer-associated mortality in
women worldwide (1). It accounts
for 30% all novel cancer diagnoses in women (1). Triple-negative breast cancer (TNBC),
characterized by the absence of estrogen receptor and progesterone
receptor in addition to a lack of overexpression of human epidermal
growth factor receptor 2 (HER2), accounts for 15% breast cancer
cases (2,3). As a distinct subtype of breast cancer,
TNBC does not respond to the standard endocrine therapies,
including tamoxifen (an anti-estrogen agent against the estrogen
receptor) and trastuzumab (a monoclonal antibody against HER2),
thus presenting a clinical challenge, as it is associated with a
higher incidence of visceral metastases, poorer prognosis, shorter
survival and higher risk of distant recurrence compered with other
types of breast cancer (2).
Identifying novel potential targets and novel therapeutic options
are urgently required to manage this aggressive type of breast
cancer.
Increasing evidence suggested that the
aggressiveness of TNBC and its resistance to standard drug
therapies may be partially due to the presence of breast cancer
stem cells (BCSCs) within TNBC tumors in addition to the normal
tissue adjacent to TNBC tumors (4–6). In
human cancer, including breast cancer, there is a small population
of cancer stem cells, which are capable of self-renewal,
differentiation, and tumor initiation and development (7). In breast cancer, a subpopulation of
breast cancer cells [CD44 antigen (CD44+)/ signal
transducer (CD24) CD24−/low] was isolated and defined as
BCSCs, which have unique stem cell-like properties that may
contribute to chemotherapy and/or radiotherapy resistance (8). In addition to the expression of CD44
and CD24, an alternate cell surface marker, aldehyde dehydrogenase
1 (ALDH1), has been used to identify BCSCs (8–11).
Ginestier et al (11)
identified that only ALDH1+ cells may develop tumors in
mice, albeit in small numbers, whereas
CD44+/CD24− is not able to. ALDH1 is
additionally considered a predictor of prognosis in patients with
breast cancer (12–15). Therefore, ALDH1 was used as a BCSC
marker in the present study.
In BCSCs, the stem-like properties, including
self-renewal, treatment-resistance and aggressiveness, are
coordinated by a network of cellular signaling pathways, including
the Notch, Hedgehog, wingless-type MMTV integration site family
(Wnt)/β-catenin, and Janus kinase (JAK)/signal transducer and
activator of transcription 3 (STAT3) signaling pathways (16). Aberrations in one or more of these
signaling pathways have been identified in cancer stem cells,
including BCSCs (16). Therefore,
targeting these signaling pathways in BCSCs is an attractive
strategy for TNBC therapy (17).
In the present study, using triple-negative,
ALDH1+ BCSC lines HCC38 and HCC1806, in vitro and
in vivo studies were conducted to investigate the anti-tumor
effects of five signaling pathway inhibitors,
N-[N-(3,5-difluorophenacetyl)-L-alanyl]-S-phenylglycine
t-butyl ester (DAPT; Notch pathway inhibitor), vismodegib
(GDC-0449; Hedgehog pathway inhibitor), salinomycin (Wnt/β-catenin
pathway inhibitor), ruxolitinib and stattic (JAK/STAT3 pathway
inhibitors; Table I), on BCSCs in
TNBC.
 | Table I.Signaling pathways and
inhibitors. |
Table I.
Signaling pathways and
inhibitors.
| Signaling pathway
inhibitors | Signaling
pathways | Targets |
|---|
| DAPT | Notch | γ-secretase |
| GDC-0449 | Hedgehog | SMO |
| Salinomycin | Wnt/β-catenin | β-catenin |
| Ruxolitinib | JAK/STAT | JAK |
| Stattic | JAK/STAT | STAT3 |
Materials and methods
Reagents and cell culture
DAPT, salinomycin, MTT, hydrocortisone and insulin
were purchased from Sigma-Aldrich (Merck KGaA, Darmstadt, Germany).
GDC-0449, ruxolitinib and stattic were obtained from Selleck
Chemicals (Houston, TX, USA). RPMI-1640, B27, penicillin and
streptomycin were obtained from Gibco (Thermo Fisher Scientific,
Inc., Waltham, MA, USA). Epidermal growth factor (EGF) and basic
fibroblast growth factor (bFGF) were provided by Prospec-Tany
TechnoGene, Ltd. (East Brunswick, NJ, USA). The HCC38 breast cancer
cell line was obtained from The American Type Culture Collection
(Manassas, VA, USA). The HCC1806 breast cancer cell line was
provided by Dr Shibo Fu at the Transform Medical College of The
First Hospital of Jilin University (Changchun, China). The two cell
lines were grown in RPMI 1640 supplemented with 10% fetal bovine
serum (FBS; Gibco; Thermo Fisher Scientific, Inc.; cat. no.
16000-044), penicillin (6.25 µg/ml) and streptomycin (100 µg/ml) at
37°C in a humidified atmosphere of 5% CO2. The two cell
lines were tested and validated by the Department of Cell Biology,
Institute of Basic Medical Science, Chinese Academy of Medical
Science (Beijing, China) and the Cell Resource Center of the
Shanghai Institute of Biological Science (Shanghai, China).
MTT cell proliferation assay
HCC38 breast cancer cells were seeded in 96-well
plates at a density of 2×103 cells/well in serum-free
RPMI-1640. On the following day, cells were treated at 37°C with
DAPT (10, 20 and 40 µM), GDC-0449 (10, 20 and 40 µM), salinomycin
(10, 20 and 40 µM) for 24 h, or ruxolitinib (1, 10 and 20 µM) and
stattic (1, 10 and 20 µM) for 72 h. Dimethyl sulfoxide (DMSO) was
used as a vehicle control. MTT reagent was added and incubated for
1 h at 37°C. The absorbance was measured at 570 nm using a
SynergyHT microplate reader (BioTek Instruments, Inc., Winooski,
VT, USA). Data were analyzed using Excel 12.0 (Microsoft
Corporation, Redmond, WA, USA).
Apoptosis assay by flow cytometry
HCC38 cells were treated with vehicle or each
signaling pathway inhibitor for the indicated time, and
1×106 cells were subsequently trypsinized to obtain a
single-cell suspension. Apoptosis analysis was performed by flow
cytometry using an Annexin V Apoptosis Detection kit (BD
Biosciences, Franklin Lakes, NJ, USA), according to the
manufacturer's protocol. Cells were stained with Annexin
V-fluorescein isothiocyanate and propidium iodide on ice for 20 min
prior to analysis. Data acquisition was performed on an LSR-II flow
cytometer (BD Biosciences) with FACSDiva 8.0.1 software (BD
Biosciences).
Mammosphere formation assay
For primary mammosphere culture, HCC38 cells were
harvested from monolayer culture and resuspended by gentle
aspiration to obtain a single-cell suspension. The cells were
subsequently seeded at a density of 1×105 cells/well in
ultra-low attachment 6-well plates (Costar; Corning, Inc., Corning,
NY, USA), and grown in serum-free Dulbecco's modified Eagle's
medium/F12 (Hyclone; GE Healthcare Life Sciences, Logan, UT, USA)
supplemented with 2% B27, 20 ng/ml EGF, 20 ng/ml bFGF, 6.25 µg/ml
penicillin, 100 µg/ml streptomycin, 1 ng/ml hydrocortisone and 10
mg/ml insulin. Cells were divided into two groups; one group was
pretreated with DMSO or signaling pathway inhibitors on the
following day; and the other was treated immediately following
mammosphere formation. After incubation for 7 or 14 days following
the treatments, mammospheres >50 µm in diameter were counted and
imaged under an inverted light microscope (Olympus IX51; Olympus
Corporation, Tokyo, Japan; magnification, ×20).
Matrigel invasion assay
A cell invasion assay was performed using 24-well BD
biocoat Matrigel invasion chambers with an 8.0-µm pore size (BD
Biosciences) according to the manufacturer's protocol. In total,
4×104 HCC38 cells were loaded into the Matrigel-coated
upper chamber filled with 500 µl serum-free RPMI containing DMSO or
a signaling pathway inhibitor. To induce cell invasion, 10%
FBS-containing RPMI was loaded into the lower chamber. Following
incubation overnight, non-invading cells remaining in the upper
chamber were removed with a cotton swab. The invading cells that
were adhered to the lower surface were fixed for 20 min in 4%
paraformaldehyde at 4°C and stained in 0.1% crystal violet solution
at 25°C for 15 min using Diff-Quik (Siemens AG, Munich, Germany).
The stained cells were counted in five randomly selected fields
under an inverted light microscope (Olympus IX51; Olympus
Corporation; magnification, ×20).
Western blot analysis
HCC38 cells were treated with DMSO or different
concentrations of signaling pathway inhibitors for 24 h and lysed
with lysis buffer (Cell Signaling Technology, Danvers, MA, USA;
cat. no. 9803) on ice. Cell lysates were collected by
centrifugation at 13,800 × g at 4°C for 10 min. The protein
concentration was measured using a bicinchoninic acid protein assay
kit with bovine serum albumin, according to the manufacturer's
protocol (Beyotime Institute of Biotechnology, Haimen, China; cat.
no. P0010). Subsequently, protein samples were heated at 95°C for 5
min in loading buffer. In total, 30 µg protein was loaded in each
lane and separated by 10% SDS-PAGE, transferred to a polyvinylidene
difluoride membrane and blocked for 1 h with 5% non-fat milk in
Tris-buffered saline containing Tween-20 (TBST) at room
temperature. Subsequently, the membranes were incubated overnight
with primary antibodies against cleaved Notch1 (cat. no. 4147;
1:1,000), zinc finger protein GLI1 (cat. no. 3538; 1:1,000),
β-catenin (cat. no. 8480; 1:1,000; all Cell Signaling Technology,
Inc.), STAT3 (cat. no. ab68153; 1:1,000), phospho-STAT3 (cat. no.
ab76315; 1:2,000), JAK1 (cat. no. ab133666; 1:1,000), JAK2 (cat.
no. ab108596; 1:2,000) and β-actin (cat. no. ab8226; 1:1,000; all
Abcam, Cambridge, UK) at 4°C. The membranes were washed with TBST
three times, subsequently incubated with horseradish
peroxidase-conjugated secondary antibodies [cat. no. 7076,
anti-mouse immunoglobulin G (IgG), 1:2,000; cat. no. 7074,
anti-rabbit IgG, 1:2,000; both Cell Signaling Technology, Inc.] for
1 h at room temperature, and washed with TBST. An enhanced
chemiluminescence kit (Thermo Fisher Scientific, Inc.; cat. no.
32106) was used and the signal was detected using the ChemiScope
5300 chemiluminescence system (Clinx Science Instruments Co. Ltd.,
Shanghai, China) and quantified using Quantity One software
(v4.6.6; Bio-Rad Laboratories, Inc., Hercules, CA, USA).
Mouse xenograft and orthotopic tumor models. In
total, 54 female nonobese diabetic/severe combined immunodeficient
(NOD/SCID) mice (National Institute of Food and Drug Control of
China, Beijing, China), 5–6 weeks old and weighing 15–20 g were
bred and maintained under specific pathogen-free conditions
(temperature at 18–29°C; air changes 10–20/h; air velocity <0.18
m/sec; 12 h light/dark cycle; and free access to food and water) at
The Animal Experiment Center, Basic Medical College of Jilin
University (Changchun, China). All the animal care details and
procedures described in the present study were approved by the
Ethics Committee of The First Hospital of Jilin University. All
animal experiments were performed in accordance with guidelines for
proper conduct of animal experiments (Jilin University). The MTT
assay determined that 10 µM was the value close to the half maximal
inhibitory concentration (IC50) of the majority of the
inhibitors in the present study. The cells were more sensitive to
ruxolitinib compared with the other inhibitors; 3 µM was the value
close to the IC50 of ruxolitinib. Triple-negative and
ALDH1+ HCC1806 cells were pretreated in vitro
with DMSO, DAPT (10 µM), GDC-0449 (10 µM), salinomycin (10 µM),
ruxolitinib (3 µM) or stattic (10 µM) for 4 h prior to mixing with
an equal volume of Matrigel (BD Biosciences). The mice were
randomly divided into six groups (n=9/group) and injected with a
mixture of pretreated HCC1806 cells and Matrigel into the mammary
fat pad, as previously described (18). Following the initial appearance,
tumors were measured every 2 days using a caliper. Tumor volumes
were calculated using the formula (length × width2)/2
(19). The tumor-free survival rate
of mice was analyzed using the Kaplan-Meier method and the log rank
test.
Statistical analysis
All experiments were repeated at least three times.
Data are presented as the mean ± standard deviation. Statistical
significance was assessed using Student's t-test or one-way
analysis of variance to compare multiple groups followed by Tukey's
or Welch's t-test (variances are not equal) to conduct multiple
comparisons between the groups with SPSS 16.0 statistical software
(SPSS Inc., Chicago, IL, USA). P<0.05 was considered to indicate
a statistically significant difference.
Results
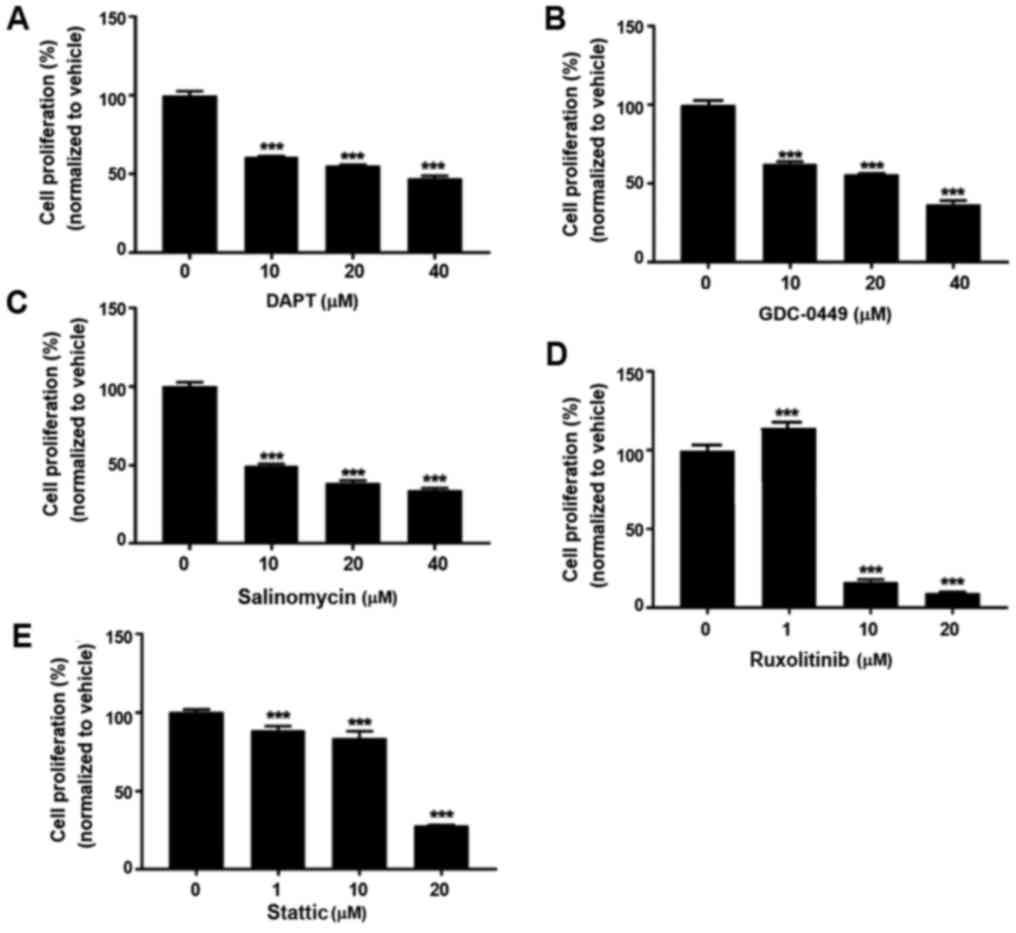
Signaling pathway inhibitors suppress
BCSC proliferation
To determine if various signaling pathway inhibitors
have effects on BCSC proliferation, an MTT assay was conducted on
HCC38 cells. As presented in Fig.
1, the inhibitors generally suppressed the proliferation of
HCC38 cells in a dose-dependent manner, suggesting the
anti-proliferative roles of these inhibitors in BCSCs.
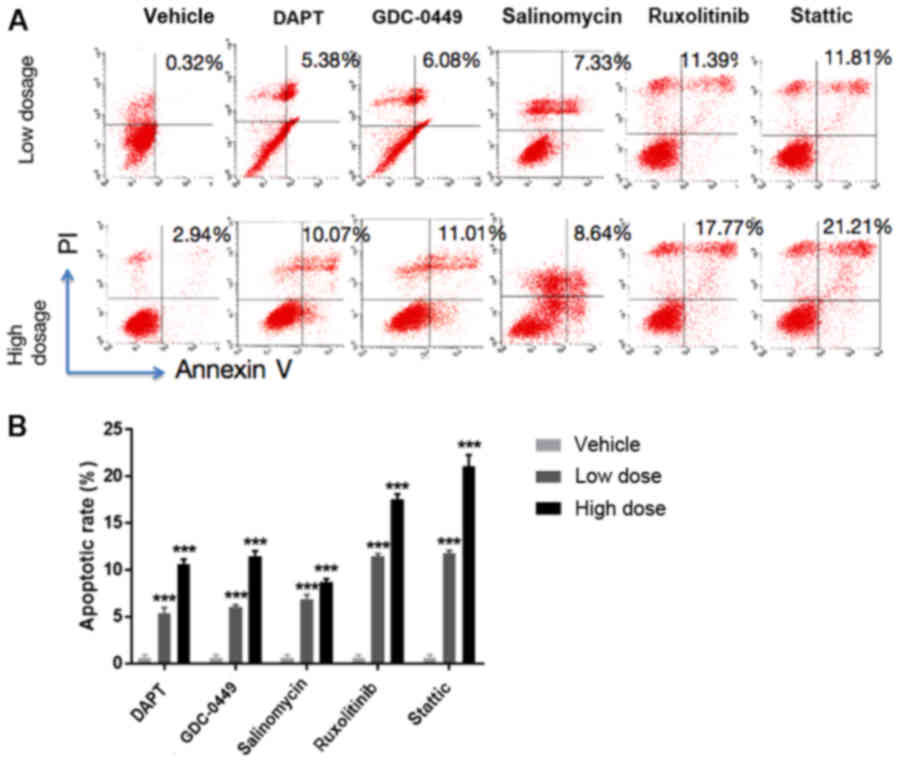
Signaling pathway inhibitors induce
apoptosis of BCSCs
To further investigate if the five signaling pathway
inhibitors induce BCSC apoptosis, which is another important
cellular event in breast cancer therapy in addition to cell
proliferation, a flow cytometry assay was performed in HCC38 cells.
As presented in Fig. 2, treatment
with DAPT, GDC-0449, salinomycin, ruxolitinib and stattic resulted
in a significantly increased apoptotic percentage ≤10.07, 11.01,
8.64, 17.77 and 21.21%, respectively, compared with the
vehicle-treated cells (P<0.001), suggesting the pro-apoptotic
roles of these inhibitors in BCSCs.
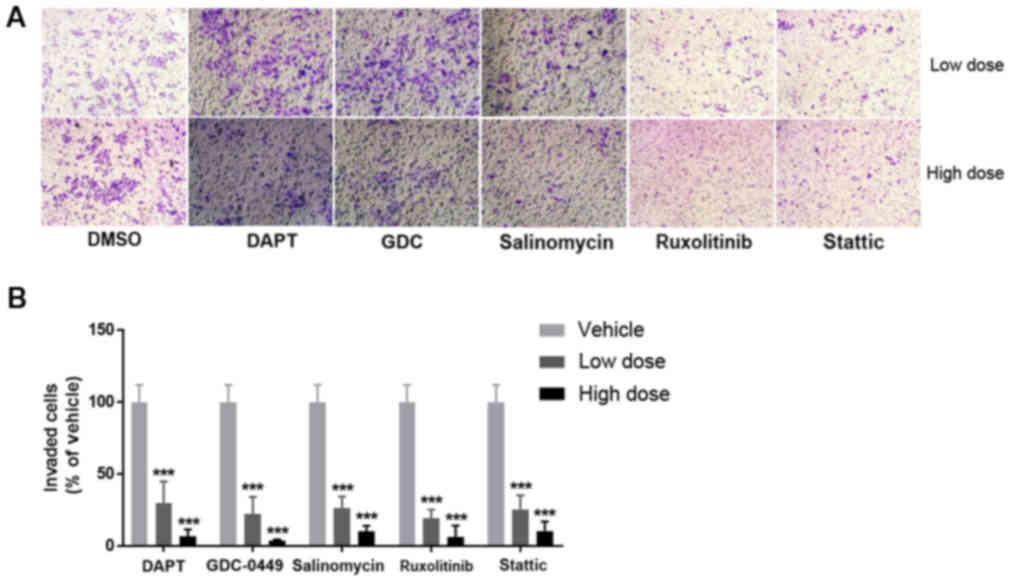
Signaling pathway inhibitors suppress
invasion of BCSCs
Cell invasion is a key process in cancer metastasis
(20). To investigate the potential
effects of the signaling pathway inhibitors on the capacity of BCSC
invasion, invasion assays were performed. As presented in Fig. 3, a significantly lower number of
invading HCC38 cells was observed with treatment with DAPT,
GDC-0449, salinomycin, ruxolitinib and stattic, compared with the
vehicle-treated group (P<0.001), suggesting that the signaling
pathway inhibitor-mediated suppression of BCSC invasion is a
possible mechanism in metastatic breast cancer therapy.
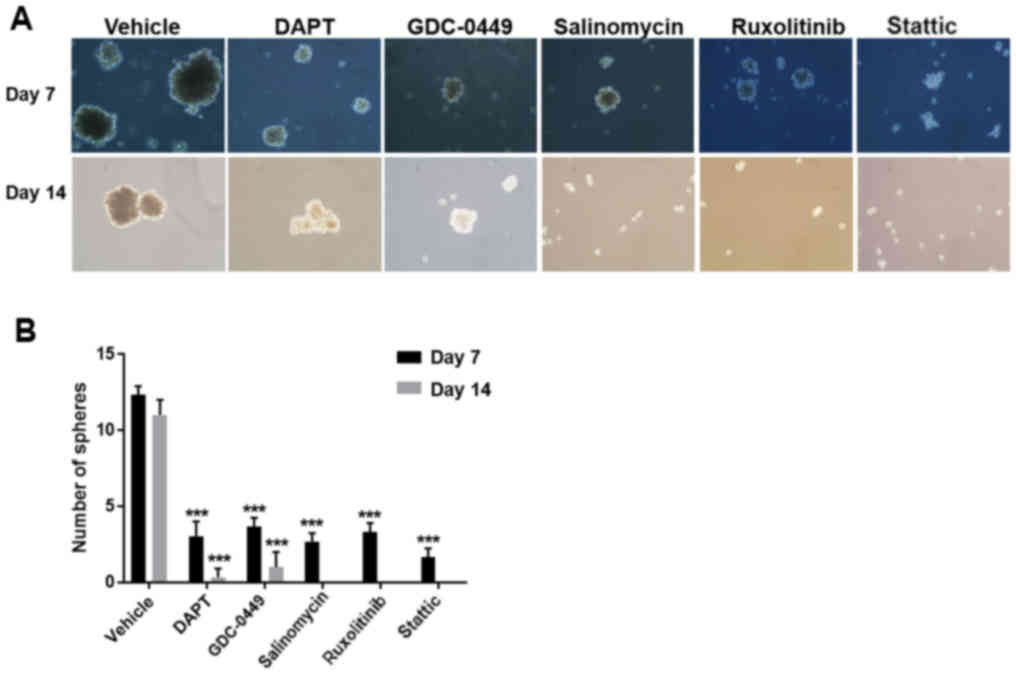
Signaling pathway inhibitors suppress
BCSC self-renewal
As stem cell self-renewal serves a critical role in
stem cell proliferation and differentiation, which are closely
associated with cancer development (21,22),
it was investigated whether the five inhibitors affect BCSC
self-renewal. A suspension mammosphere assay, which is commonly
used for measuring stem cell activity and in vitro
stem/progenitor cell frequency, was performed on HCC38 cells. The
results demonstrated that pretreatment with signaling pathway
inhibitors prior to mammosphere formation markedly decreased the
sphere size and the number of HCC38 cells, compared with the
vehicle-treated control (Fig. 4).
Among the inhibitors, stattic was the most potent one, as
demonstrated by the lack of any mammospheres (Fig. 4f-h).
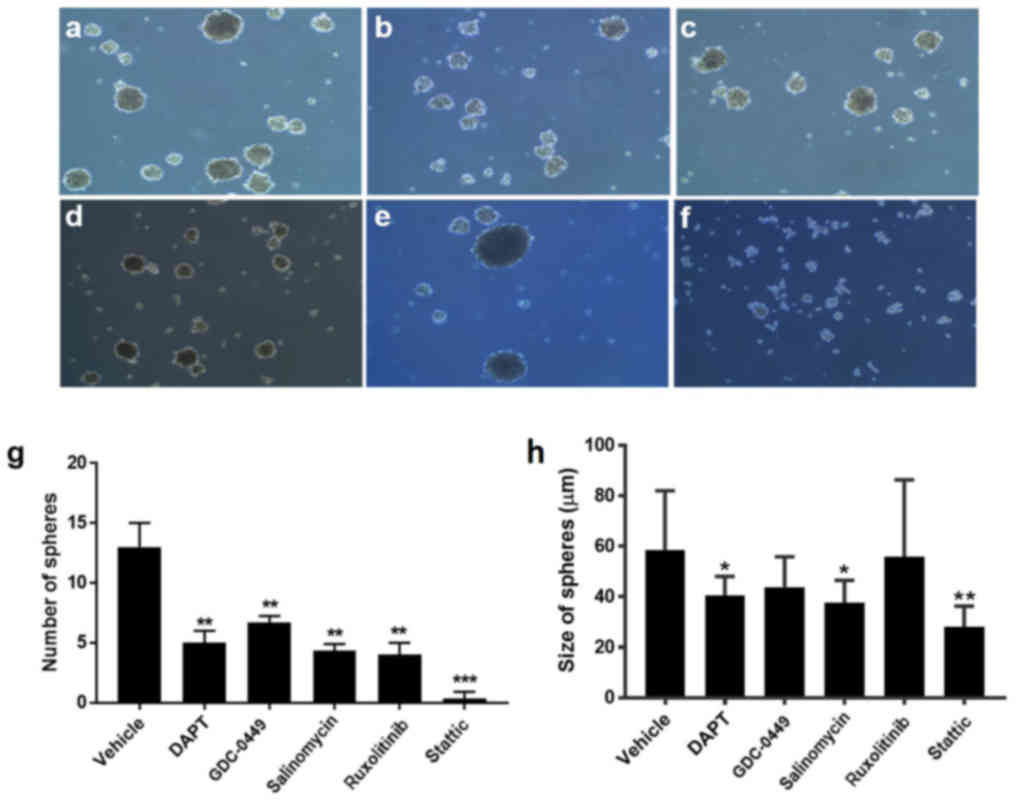 | Figure 4.Signaling pathway inhibitors suppress
mammosphere formation of HCC38 cells. Morphology of HCC38
cell-formed mammospheres after a 7-day pretreatment with (a)
vehicle, (b) 20 µM DAPT, (c) 20 µM GDC-0449, (d) 40 µM salinomycin,
(e) 10 µM ruxolitinib or (f) 5 µM stattic under a microscope.
Magnification, ×20. (g) Quantification of the number of
mammospheres formed after a 7-day incubation with the inhibitors.
(h) Quantification of the size of mammospheres formed after a 7-day
incubation with the inhibitors. All experiments were repeated at
least three times. n=3. *P<0.05, **P<0.01, ***P<0.001 vs.
vehicle. DAPT,
N-[N-(3,5-difluorophenacetyl)-L-alanyl]-S-phenylglycine
t-butyl ester; GDC-0449, vismodegib. |
Similarly, treatment with the signaling pathway
inhibitors immediately following mammosphere formation failed to
maintain the structure of the formed mammosheres (Fig. 5). Following prolonged incubation,
treatment with salinomycin, ruxolitinib and stattic even led to
further disassembly of mammospheres (Fig. 5). These results demonstrated that
the signaling pathway inhibitors diminished the tumorsphere-forming
ability of BCSCs and the maintenance of BCSC-formed mammospheres,
suggesting the negative roles of these inhibitors in the BCSC
self-renewal process.
Signaling pathway inhibitors suppress
expression and phosphorylation of downstream targets
As the inhibitors block signal transduction, they
were predicted to inhibit the activity of their downstream target
molecules. In general, these inhibitors markedly decreased the
expression or phosphorylation of their corresponding downstream
signaling molecules in a dose-dependent manner, as demonstrated in
Fig. 6.
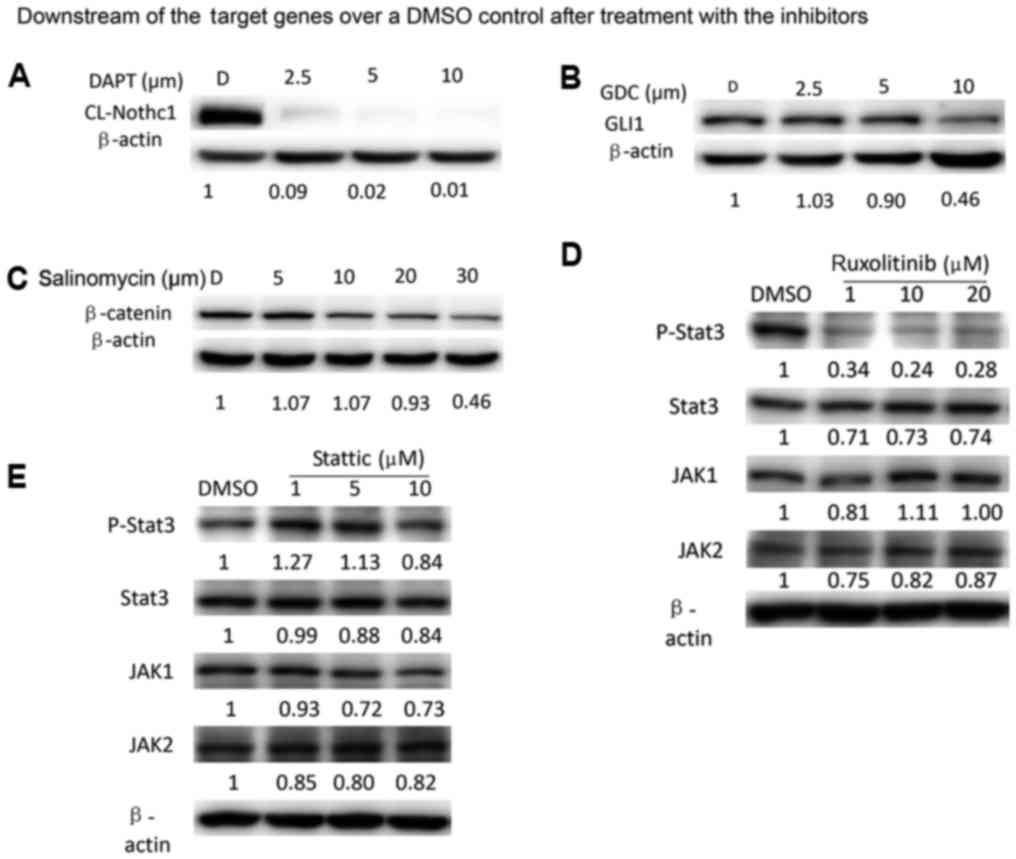 | Figure 6.Signaling pathway inhibitors suppress
expression or phosphorylation of downstream targets. Western blot
analysis for the expression of (A) cl-Notch1, (B) GLI1, (C)
β-catenin. Expression of p-STAT3, total STAT3, JAK1 and JAK2 in
HCC38 cells treated with different concentrations of the (D)
ruxolitinib and (E) stattic for 24 h. All experiments were repeated
at least three times. STAT3, signal transducer and activator of
transcription; JAK, Janus kinase; p, phosphorylated; cl, cleaved;
DMSO, dimethyl sulfoxide; GLI1, zinc finger protein GLI1. |
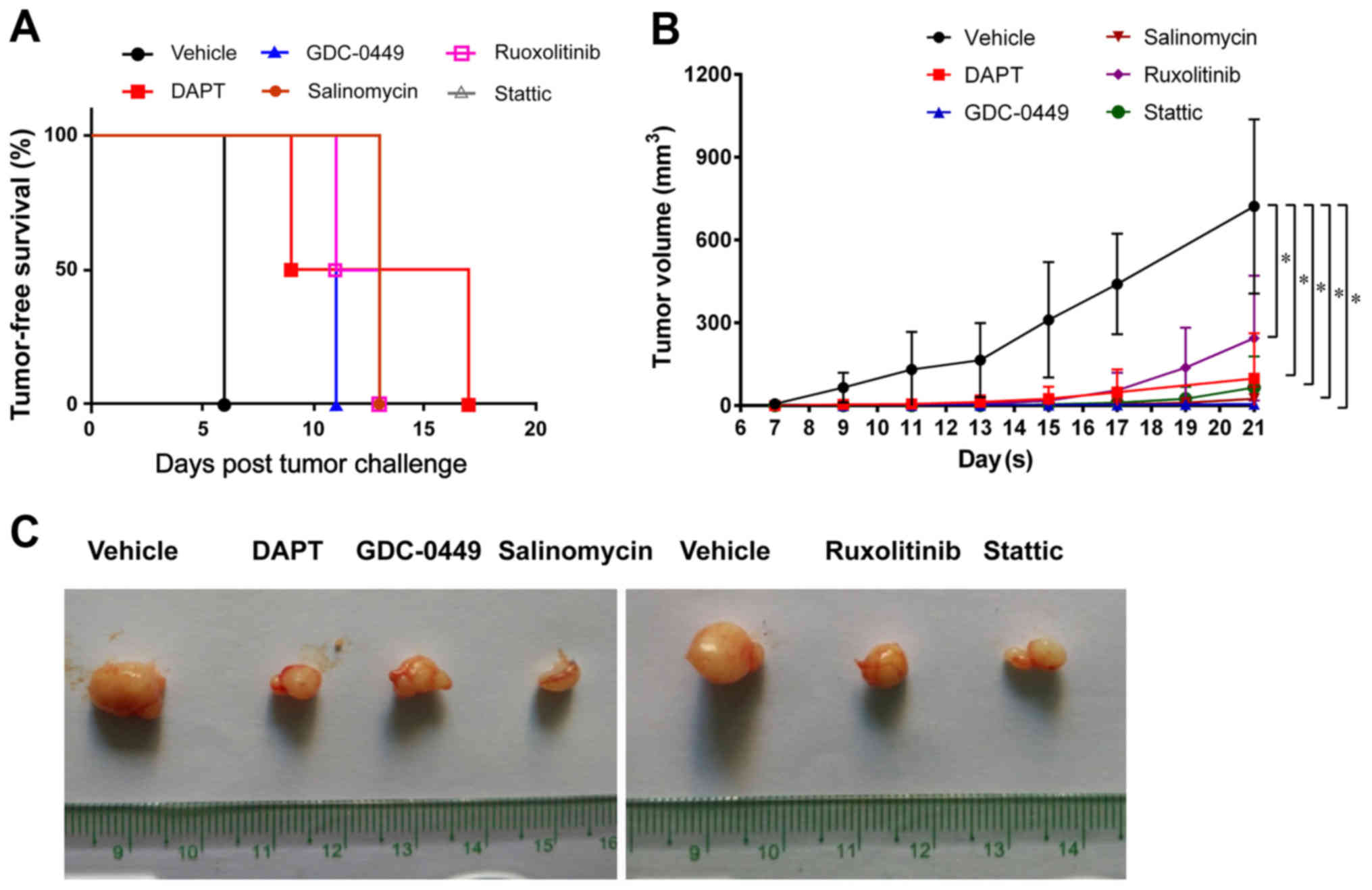
Signaling pathway inhibitors suppress
the tumor-forming ability of TNBC
To determine the effects of the signaling pathway
inhibitors on the breast tumor-forming ability in vivo,
HCC1806 cells pretreated with vehicle or inhibitors were injected
into NOD/SCID mice. It was observed that all mice injected with
vehicle-treated HCC1806 cells developed mammary tumors at 7 days
following injection (Fig. 7A). In
contrast, mice injected with inhibitor-treated HCC1806 cells
exhibited a delay in tumor formation and a decrease in tumor
incidence (Fig. 7A). At 21 days
after injection, all the mice in the treatment groups exhibited a
significant decreased tumor volume compared with the control group
(Fig. 7B and C; P<0.05),
suggesting that these inhibitors suppress the growth of
BSBC-derived tumors in vivo.
Discussion
The present study aimed to determine if inhibitors
of the Notch, Hedgehog, Wnt and JAK/STAT signaling pathways may be
used as potential therapeutic agents targeting BCSCs in TNBC. It
was demonstrated that the five signaling pathway inhibitors, DAPT,
GDC-0449, salinomycin, ruxolitinib and stattic, individually
suppressed the proliferation and promoted the apoptosis of HCC38
cells, a TNBC cell line with stem cell-like characteristics
(CD44+/CD24low/− and ALDH+) that
exhibits the features of BCSCs. The invasion assay demonstrated
that the five inhibitors significantly decreased HCC38 invasion
compared with the control group, suggesting their suppressive
effects on the breast cancer invasive capacity. In addition, these
inhibitors blocked the BCSC mammosphere process by preventing
mammosphere formation and promoting mammosphere disassembly,
suggesting that these inhibitors may simultaneously inhibit the
proliferation and differentiation of cancer stem cells. However, it
was observed that GDC-0449 and ruxolitinib had no significant
inhibitory effects on the mammosphere size. A possible explanation
for this discrepancy is that the mammospheres were no longer
sensitive to the present drug concentrations when they grew to a
given size (40 µm). The mammospheres >40 µm had limited drug
contact with interior HCC38 cells due to their large size.
Therefore, higher concentrations may be required to increase the
sensitivity of the mammospheres to these two inhibitors; further
investigation in a future study is required. In the present study,
only HCC38 cells (100% ALDH+) were used for the in
vitro study as they are considered an ideal candidate cell line
of BCSCs (23). To the best of the
authors' knowledge, at present, no other cell line shares more
characteristic features of BCSCs than HCC38. Therefore, data from
HCC38 cells are representative of an in vitro BCSC
study.
DAPT is used as a γ-secretase inhibitor to block
Notch signaling (24). The present
study demonstrated that DAPT downregulated the expression of
cl-Notch1, the activated form of the Notch1 receptor, ≤99% in HCC38
cells, demonstrating that DAPT prevents the final cleavage step of
the Notch1 receptor by inactivating γ-secretase and subsequently
decreasing the expression level of cl-Notch1 (25). As the Notch signaling pathway is
dysregulated due to the overexpression of Notch receptors and their
ligands in human breast cancer (26,27),
the expression of cl-Notch1 is a notable biomarker for therapeutic
efficacy of drugs or agents in breast cancer (28,29).
Additionally, it was demonstrated that DAPT suppressed BCSC
proliferation and tumor growth in vitro and in vivo.
Therefore, DAPT may serve as a promising therapeutic agent in
breast cancer by targeting the Notch signaling pathway in
BCSCs.
GDC-0449 was used in the present study to target the
Hedgehog signaling pathway, which is normally in a resting state;
however, is activated in response to carcinogenic stimuli by
hedgehog ligands binding to a transmembrane receptor called Patched
(PTCH) (30). The hedgehog
signaling pathway serves a significant role in cancer development
and progression in various malignancies, including breast cancer
(31–34). Following the hedgehog ligand-PTCH
binding, smoothened (SMO) protein initiates the signaling cascade
by activating GLI transcription factors, which in turn drive the
expression of a variety of target genes that are associated with
carcinogenesis (35,36). In the present study, treatment with
40 µM GDC-0449 led to ~50% decrease in the GLI1 protein expression
level in HCC38 cells, which is likely due to the direct inhibition
of SMO by GDC-0449. Consistently, GDC-0449 inhibited BCSC
proliferation, invasion and mammosphere formation, while inducing
BCSC apoptosis. Based on the present in vivo data, it was
hypothesized that SMO is undetectable in normal breast tissue
however, is increased in breast cancer tissues. Consistently, SMO
has been identified to be ectopically expressed in 70% ductal
carcinoma in situ and 30% invasive breast cancer in an
animal model (37), suggesting that
inhibition of SMO is a valuable therapeutic strategy against BCSCs.
Previously, GDC-0449 was approved by the US Food and Drug
Administration for the treatment of advanced basal-cell carcinoma
(38–40), which may result in the future
clinical application of GDC-0449 in breast cancer therapy.
Salinomycin was originally developed as a commercial
antibiotic in veterinary medicine (41). A previous study demonstrated that
salinomycin may additionally be used as an effective drug against
breast cancer by targeting drug-resistant BCSCs (41). Previously, Lu et al (42) observed that salinomycin inhibits
Wnt/β-catenin signaling, a key signaling pathway supporting
self-renewal of normal and malignant mammary stem cells (43). Enhanced Wnt signaling contributes to
cell proliferation in the majority of breast cancer by
downregulating the expression of secreted Frizzled-related protein
1, a negative Wnt pathway regulator (44). These results suggested that
salinomycin is a promising anticancer drug by inhibiting
Wnt/β-catenin signaling. The present results demonstrated that
salinomycin, by markedly decreasing the expression of β-catenin,
significantly suppressed BCSC proliferation and mammosphere
formation in vitro in addition to tumor formation in
vivo.
Ruxolitinib (Jakafi) and stattic are inhibitors of
the JAK/STAT3 pathway, another important pathway in normal and
cancer stem cells, which is considered a promising therapeutic
target (45). JAKs phosphorylate
STATs, thus subsequently activating the signaling pathway and
various target genes (46).
Ruxolitinib and stattic inhibit JAKs and STATs, respectively
(47,48). In the present study, ruxolitinib
markedly downregulated the expression of phospho-STAT3; however,
appeared to have no marked effects on JAK1 and JAK2 expression. A
possible explanation for this discrepancy is that the activities of
JAK1 and JAK2 are determined by their phosphorylation levels, not
by their protein expression levels. However, stattic did not appear
to markedly alter the expression of all four proteins.
Phosphorylation levels of JAK1 and JAK2 require examination to
further conform the role of stattic in suppressing breast cancer in
future studies. Furthermore, further investigation is required to
examine the expression pattern of SMO in the inhibitors-treated
BCSCs, which may provide novel insight for the underlying
mechanisms.
Although a number of previous studies demonstrated
that these five signaling inhibitors exert inhibitory effects on
TNBC in animal models and in clinical trials (49–53),
in the present study, it was demonstrated that all five signaling
inhibitors suppressed stemness of BCSCs in extensive aspects,
including proliferation, invasion, apoptosis, self-renewal,
mammosphere formation and tumorigenesis. In addition, the
inhibition efficacy of these five inhibitors against TNBC was
compared. The present in vitro and in vivo data
demonstrated that these five signaling inhibitors, individually or
in combination, may be applied clinically with high efficacy.
In conclusion, it was demonstrated that the
signaling pathway inhibitors suppressed BCSC proliferation,
invasion and mammosphere number, while inducing apoptosis. As these
signaling pathway inhibitors, with the exception of stattic, have
been applied clinically or in clinical trials for the intervention
of disorders other than TNBC, including operable basal cell
carcinoma, myelofibrosis and coccidial infection (54–60),
they may provide novel therapeutic options for TNBC.
Acknowledgements
The authors would like to thank Dr Shibo Fu of the
Transforming Medical College of First Hospital, Jilin University
(Changchun, China), who revised the manuscript and conducted the
western blot experiments.
Funding
The present study was financially supported by the
Bethune Plan B (Jilin, China; grant no. 2012217), the National
Natural Scientific Fund of China Grant (grant no. 30300336) and the
Health Specific Programme of Jilin Province (grant no.
2018SCZWSZX-035).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
WL, HY and XL performed the examination. XL and LH
prepared the figures. LH and NX conducted the statistical analyses.
AS analyzed and interpreted the data, and was the principal
contributor in writing the manuscript. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
All the animal care details and procedures described
in the present study were approved by the Ethics Committee of The
First Hospital of Jilin University (Changchun, China). All animal
experiments were performed in accordance with guidelines for proper
conduct of animal experiments (Jilin University).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
ALDH1
|
aldehyde dehydrogenase 1
|
|
BCSCs
|
breast cancer stem cells
|
|
bFGF
|
basic fibroblast growth factor
|
|
cl-Notch1
|
cleaved Notch1 intracellular
domain
|
|
DAPT
|
N-[N-(3,5-difluorophenacetyl)-L-alanyl]-S-phenylglycine
t-butyl ester
|
|
DMSO
|
dimethyl sulfoxide
|
|
EGF
|
epidermal growth factor
|
|
FBS
|
fetal bovine serum
|
|
GDC-0449
|
vismodegib
|
|
HER2
|
human epidermal growth factor receptor
2
|
|
JAK
|
Janus kinase
|
|
NOD/SCID
|
non-obese diabetic/severe combined
immunodeficient
|
|
PTCH
|
Patched
|
|
SMO
|
smoothened
|
|
STAT3
|
signal transducer and activator of
transcription 3
|
|
TBST
|
Tris-buffered saline containing
Tween-20
|
|
TNBC
|
triple-negative breast cancer
|
|
Wnt
|
wingless-type MMTV integration site
family
|
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
Statistics, 2017. CA Cancer J Clin. 67:7–30. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bauer KR, Brown M, Cress RD, Parise CA and
Caggiano V: Descriptive analysis of estrogen receptor
(ER)-negative, progesterone receptor (PR)-negative, and
HER2-negative invasive breast cancer, the so-called triple-negative
phenotype: A population-based study from the California cancer
Registry. Cancer. 109:1721–1728. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Dent R, Trudeau M, Pritchard KI, Hanna WM,
Kahn HK, Sawka CA, Lickley LA, Rawlinson E, Sun P and Narod SA:
Triple-negative breast cancer: Clinical features and patterns of
recurrence. Clin Cancer Res. 13:4429–4434. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Atkinson RL, Yang WT, Rosen DG, Landis MD,
Wong H, Lewis MT, Creighton CJ, Sexton KR, Hilsenbeck SG, Sahin AA,
et al: Cancer stem cell markers are enriched in normal tissue
adjacent to triple negative breast cancer and inversely correlated
with DNA repair deficiency. Breast Cancer Res. 15:R772013.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lehmann BD, Bauer JA, Chen X, Sanders ME,
Chakravarthy AB, Shyr Y and Pietenpol JA: Identification of human
triple-negative breast cancer subtypes and preclinical models for
selection of targeted therapies. J Clin Invest. 121:2750–2767.
2011. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Burstein MD, Tsimelzon A, Poage GM,
Covington KR, Contreras A, Fuqua SA, Savage MI, Osborne CK,
Hilsenbeck SG, Chang JC, et al: Comprehensive genomic analysis
identifies novel subtypes and targets of triple-negative breast
cancer. Clin Cancer Res. 21:1688–1698. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Dalerba P, Cho RW and Clarke MF: Cancer
stem cells: Models and concepts. Annu Rev Med. 58:267–284. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Geng SQ, Alexandrou AT and Li JJ: Breast
cancer stem cells: Multiple capacities in tumor metastasis. Cancer
Lett. 349:1–7. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Collina F, Di Bonito M, Li Bergolis V, De
Laurentiis M, Vitagliano C, Cerrone M, Nuzzo F, Cantile M and Botti
G: Prognostic value of cancer stem cells markers in triple-negative
breast cancer. BioMed Res Int. 2015:1586822015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Al-Hajj M, Wicha MS, Benito-Hernandez A,
Morrison SJ and Clarke MF: Prospective identification of
tumorigenic breast cancer cells. Proc Natl Acad Sci USA.
100:3983–3988. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ginestier C, Hur MH, Charafe-Jauffret E,
Monville F, Dutcher J, Brown M, Jacquemier J, Viens P, Kleer CG,
Liu S, et al: ALDH1 is a marker of normal and malignant human
mammary stem cells and a predictor of poor clinical outcome. Cell
Stem Cell. 1:555–567. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Resetkova E, Reis-Filho JS, Jain RK, Mehta
R, Thorat MA, Nakshatri H and Badve S: Prognostic impact of ALDH1
in breast cancer: A story of stem cells and tumor microenvironment.
Breast Cancer Res Treat. 123:97–108. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Charafe-Jauffret E, Ginestier C, Bertucci
F, Cabaud O, Wicinski J, Finetti P, Josselin E, Adelaide J, Nguyen
TT, Monville F, et al: ALDH1-positive cancer stem cells predict
engraftment of primary breast tumors and are governed by a common
stem cell program. Cancer Res. 73:7290–7300. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Alamgeer M, Ganju V, Kumar B, Fox J, Hart
S, White M, Harris M, Stuckey J, Prodanovic Z, Schneider-Kolsky ME,
et al: Changes in aldehyde dehydrogenase-1 expression during
neoadjuvant chemotherapy predict outcome in locally advanced breast
cancer. Breast Cancer Res. 16:R442014. View
Article : Google Scholar : PubMed/NCBI
|
|
15
|
Marcato P, Dean CA, Liu RZ, Coyle KM,
Bydoun M, Wallace M, Clements D, Turner C, Mathenge EG, Gujar SA,
et al: Aldehyde dehydrogenase 1A3 influences breast cancer
progression via differential retinoic acid signaling. Mol Oncol.
9:17–31. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Pires BR, DE Amorim ÍS, Souza LD,
Rodrigues JA and Mencalha AL: Targeting cellular signaling pathways
in breast cancer stem cells and its implication for cancer
treatment. Anticancer Res. 36:5681–5691. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Takebe N, Miele L, Harris PJ, Jeong W,
Bando H, Kahn M, Yang SX and Ivy SP: Targeting Notch, Hedgehog, and
Wnt pathways in cancer stem cells: Clinical update. Nat Rev Clin
Oncol. 12:445–464. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Frank NY, Schatton T and Frank MH: The
therapeutic promise of the cancer stem cell concept. J Clin Invest.
120:41–50. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Naito S, von Eschenbach AC, Giavazzi R and
Fidler IJ: Growth and metastasis of tumor cells isolated from a
human renal cell carcinoma implanted into different organs of nude
mice. Cancer Res. 46:4109–4115. 1986.PubMed/NCBI
|
|
20
|
Krakhmal NV, Zavyalova MV, Denisov EV,
Vtorushin SV and Perelmuter VM: Cancer invasion: Patterns and
mechanisms. Acta Naturae. 7:17–28. 2015.PubMed/NCBI
|
|
21
|
Shaw FL, Harrison H, Spence K, Ablett MP,
Simões BM, Farnie G and Clarke RB: A detailed mammosphere assay
protocol for the quantification of breast stem cell activity. J
Mammary Gland Biol Neoplasia. 17:111–117. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Harrison H, Farnie G, Howell SJ, Rock RE,
Stylianou S, Brennan KR, Bundred NJ and Clarke RB: Regulation of
breast cancer stem cell activity by signaling through the Notch4
receptor. Cancer Res. 70:709–718. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Londoño-Joshi AI, Oliver PG, Li Y, Lee CH,
Forero-Torres A, LoBuglio AF and Buchsbaum DJ: Basal-like breast
cancer stem cells are sensitive to anti-DR5 mediated cytotoxicity.
Breast Cancer Res Treat. 133:437–445. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Jiang LY, Zhang XL, Du P and Zheng JH:
γ-Secretase inhibitor, DAPT inhibits self-renewal and stemness
maintenance of ovarian cancer stem-like cells in vitro. Chin J
Cancer Res. 23:140–146. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Olsauskas-Kuprys R, Zlobin A and Osipo C:
Gamma secretase inhibitors of Notch signaling. Onco Targets Ther.
6:943–955. 2013.PubMed/NCBI
|
|
26
|
Imatani A and Callahan R: Identification
of a novel NOTCH-4/INT-3 RNA species encoding an activated gene
product in certain human tumor cell lines. Oncogene. 19:223–231.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Stylianou S, Clarke RB and Brennan K:
Aberrant activation of notch signaling in human breast cancer.
Cancer Res. 66:1517–1525. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Stoeck A, Lejnine S, Truong A, Pan L, Wang
H, Zang C, Yuan J, Ware C, MacLean J, Garrett-Engele PW, et al:
Discovery of biomarkers predictive of GSI response in
triple-negative breast cancer and adenoid cystic carcinoma. Cancer
Discov. 4:1154–1167. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhong Y, Shen S, Zhou Y, Mao F, Lin Y,
Guan J, Xu Y, Zhang S, Liu X and Sun Q: NOTCH1 is a poor prognostic
factor for breast cancer and is associated with breast cancer stem
cells. OncoTargets Ther. 9:6865–6871. 2016. View Article : Google Scholar
|
|
30
|
Gupta S, Takebe N and Lorusso P: Targeting
the Hedgehog pathway in cancer. Ther Adv Med Oncol. 2:237–250.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Beachy PA, Hymowitz SG, Lazarus RA, Leahy
DJ and Siebold C: Interactions between Hedgehog proteins and their
binding partners come into view. Genes Dev. 24:2001–2012. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Varjosalo M and Taipale J: Hedgehog:
Functions and mechanisms. Genes Dev. 22:2454–2472. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Mullor JL, Sánchez P and Ruiz i Altaba A:
Pathways and consequences: Hedgehog signaling in human disease.
Trends Cell Biol. 12:562–569. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Ng JMY and Curran T: The Hedgehog's tale:
Developing strategies for targeting cancer. Nat Rev Cancer.
11:493–501. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Odoux C, Fohrer H, Hoppo T, Guzik L, Stolz
DB, Lewis DW, Gollin SM, Gamblin TC, Geller DA and Lagasse E: A
stochastic model for cancer stem cell origin in metastatic colon
cancer. Cancer Res. 68:6932–6941. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Amakye D, Jagani Z and Dorsch M:
Unraveling the therapeutic potential of the Hedgehog pathway in
cancer. Nat Med. 19:1410–1422. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Moraes RC, Zhang X, Harrington N, Fung JY,
Wu MF, Hilsenbeck SG, Allred DC and Lewis MT: Constitutive
activation of smoothened (SMO) in mammary glands of transgenic mice
leads to increased proliferation, altered differentiation and
ductal dysplasia. Development. 134:1231–1242. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Sandhiya S, Melvin G, Kumar SS and Dkhar
SA: The dawn of hedgehog inhibitors: Vismodegib. J Pharmacol
Pharmacother. 4:4–7. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Rudin CM, Hann CL, Laterra J, Yauch RL,
Callahan CA, Fu L, Holcomb T, Stinson J, Gould SE, Coleman B, et
al: Treatment of medulloblastoma with hedgehog pathway inhibitor
GDC-0449. N Engl J Med. 361:1173–1178. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Von Hoff DD, LoRusso PM, Rudin CM, Reddy
JC, Yauch RL, Tibes R, Weiss GJ, Borad MJ, Hann CL, Brahmer JR, et
al: Inhibition of the hedgehog pathway in advanced basal-cell
carcinoma. N Engl J Med. 361:1164–1172. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Gupta PB, Onder TT, Jiang G, Tao K,
Kuperwasser C, Weinberg RA and Lander ES: Identification of
selective inhibitors of cancer stem cells by high-throughput
screening. Cell. 138:645–659. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Lu D, Choi MY, Yu J, Castro JE, Kipps TJ
and Carson DA: Salinomycin inhibits Wnt signaling and selectively
induces apoptosis in chronic lymphocytic leukemia cells. Proc Natl
Acad Sci USA. 108:13253–13257. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Smalley MJ and Dale TC: Wnt signalling in
mammalian development and cancer. Cancer Metastasis Rev.
18:215–230. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Schlange T, Matsuda Y, Lienhard S, Huber A
and Hynes NE: Autocrine WNT signaling contributes to breast cancer
cell proliferation via the canonical WNT pathway and EGFR
transactivation. Breast Cancer Res. 9:R632007. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Hernandez-Vargas H, Ouzounova M, Le
Calvez-Kelm F, Lambert MP, McKay-Chopin S, Tavtigian SV, Puisieux
A, Matar C and Herceg Z: Methylome analysis reveals Jak-STAT
pathway deregulation in putative breast cancer stem cells.
Epigenetics. 6:428–439. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Buchert M, Burns CJ and Ernst M: Targeting
JAK kinase in solid tumors: Emerging opportunities and challenges.
Oncogene. 35:939–951. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Schust J, Sperl B, Hollis A, Mayer TU and
Berg T: Stattic: A small-molecule inhibitor of STAT3 activation and
dimerization. Chem Biol. 13:1235–1242. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Tavallai M, Booth L, Roberts JL,
Poklepovic A and Dent P: Rationally repurposing ruxolitinib (Jakafi
(®)) as a solid tumor therapeutic. Front Oncol.
6:1422016. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Aktas CC, Zeybek ND and Piskin AK: In
vitro effects of phenytoin and DAPT on MDA-MB-231 breast cancer
cells. Acta Biochim Biophys Sin (Shanghai). 47:680–686. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Kai M, Kanaya N, Wu SV, Mendez C, Nguyen
D, Luu T and Chen S: Targeting breast cancer stem cells in
triple-negative breast cancer using a combination of LBH589 and
salinomycin. Breast Cancer Res Treat. 151:281–294. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Stover DG, Gil Del Alcazar CR, Brock J,
Guo H, Overmoyer B, Balko J, Xu Q, Bardia A, Tolaney SM, Gelman R,
et al: Phase II study of ruxolitinib, a selective JAK1/2 inhibitor,
in patients with metastatic triple-negative breast cancer. NPJ
Breast Cancer. 4:102018. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Rai G, Suman S, Mishra S and Shukla Y:
Evaluation of growth inhibitory response of Resveratrol and
Salinomycin combinations against triple negative breast cancer
cells. Biomed Pharmacother. 89:1142–1151. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Habib JG and O'Shaughnessy JA: The
hedgehog pathway in triple-negative breast cancer. Cancer Med.
5:2989–3006. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Robarge KD, Brunton SA, Castanedo GM, Cui
Y, Dina MS, Goldsmith R, Gould SE, Guichert O, Gunzner JL, Halladay
J, et al: GDC-0449-a potent inhibitor of the hedgehog pathway.
Bioorg Med Chem Lett. 19:5576–5581. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Luistro L, He W, Smith M, Packman K,
Vilenchik M, Carvajal D, Roberts J, Cai J, Berkofsky-Fessler W,
Hilton H, et al: Preclinical profile of a potent gamma-secretase
inhibitor targeting notch signaling with in vivo efficacy and
pharmacodynamic properties. Cancer Res. 69:7672–7680. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Fan X, Khaki L, Zhu TS, Soules ME, Talsma
CE, Gul N, Koh C, Zhang J, Li YM, Maciaczyk J, et al: NOTCH pathway
blockade depletes CD133-positive glioblastoma cells and inhibits
growth of tumor neurospheres and xenografts. Stem Cells. 28:5–16.
2010.PubMed/NCBI
|
|
57
|
Antoszczak M and Huczyński A: Anticancer
Activity of Polyether Ionophore-Salinomycin. Anticancer Agents Med
Chem. 15:575–591. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Zhang Q, Zhang C, He J, Guo Q, Hu D, Yang
X, Wang J, Kang Y, She R, Wang Z, et al: STAT3 inhibitor stattic
enhances radiosensitivity in esophageal squamous cell carcinoma.
Tumour Biol. 36:2135–2142. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Plosker GL: Ruxolitinib: A review of its
use in patients with myelofibrosis. Drugs. 75:297–308. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Vannucchi AM, Kiladjian JJ, Griesshammer
M, Masszi T, Durrant S, Passamonti F, Harrison CN, Pane F, Zachee
P, Mesa R, et al: Ruxolitinib versus standard therapy for the
treatment of polycythemia vera. N Engl J Med. 372:426–435. 2015.
View Article : Google Scholar : PubMed/NCBI
|