Introduction
Endometrial carcinoma (EC) is the most common
gynecological malignancy worldwide. Diagnosis of 63,230 new cases
in the USA, resulting in 11,350 disease-related deaths, were
predicted in 2018 (1). According to
its clinical and pathological characteristics, EC is traditionally
classified into two types (2). Type
I includes tumors of endometrioid histology graded as 1, 2 and 3
which are estrogen-dependent and usually preceded by endometrial
hyperplasia with a favorable prognosis. Conversely, type II tumors
are predominantly papillary serous carcinomas caused by atrophy of
the endometrium. They are not consistently correlated with estrogen
exposure and reveal a strong ability of aggression (3,4).
It is undeniable that estradiol (E2)
plays a critical role in the development of EC (5). E2 and its analogs regulate
the expression of selective estrogen-responsive genes by combining
to the cytoplasmic estrogen receptor α (ERα), and subsequently, the
complex migrates to the nucleus to activate the target genes
(6). However, the therapeutic
efficiency of hormones for treating EC is decreasing due to drug
insensitivity. Thus, it is essential to raise our awareness of the
molecular and cellular mechanisms underlying EC and to develop new
therapeutic strategies to prevent the progression of EC.
MicroRNAs (miRNAs) are a class of small non-coding
RNAs (~22 nt). It is well known that miRNAs induces RNA
interference by base-pairing with the 3′ untranslated region (UTR)
of a complementary messenger RNA (mRNA), thus resulting with either
mRNA translational repression or RNA degradation (7). In human cancers, specific miRNAs exist
in different tissues. The aberrant expression of miRNAs is
correlated with carcinogenesis (8),
including EC carcinogenesis (9).
Furthermore, miRNAs interact with certain transcription factors
(TFs) to regulate mutual sets of target genes, thus realizing the
coordinated modulation of gene transcriptional and
post-transcriptional expression. We previously uncovered a
regulatory loop involving TrkB/miR-204-5p that is critical in the
tumorigenesis of EC (10), and
these observations led us to hypothesize that specific miRNAs may
control ERα expression post-transcriptionally during EC
progression.
In the present study, the target gene of miR-107
containing ESR1 was predicted by bioinformatics analysis. Aberrant
expression of miR-107 has been revealed to promote the
tumorigenesis of glioma and lung cancer (11,12).
However, the mechanism by which miR-107 participates in EC is
poorly understood. Notably, our findings revealed that
anti-miR-107-5p inhibited the proliferation and invasion of EC
cells by targeting the 3′UTR of ESR1 mRNA directly. Our findings
provide new ideas for further understanding the mechanism of cell
proliferation mediated by miR-107-5p in EC.
Materials and methods
Clinical samples
Seventy-one EC patients who underwent hysterectomy
with lymph node dissection at the Shanghai General Hospital
Affiliated with Shanghai Jiao Tong University School of Medicine
from August 2014 to May 2016 were included in our study (Table I). The stages and histological
grades of these tumors were determined according to the criteria of
the Federation International of Gynecology and Obstetrics (FIGO)
surgical staging system (2009) (13). Twenty-six samples of normal
endometrium collected from October 2014 to June 2015 were obtained
from patients who underwent hysterectomy for other diseases, such
as myoma or adenomyosis. None of the patients had received hormone
therapy, radiotherapy, or chemotherapy prior to surgery. The
resected specimens were stained with hematoxylin and eosin
(H&E) for histological examination. The original tissues were
collected from each patient immediately after resection and
snap-frozen in liquid nitrogen until further use. The present study
was approved by the Human Investigation Ethical Committee of
Shanghai General Hospital Affiliated with Shanghai Jiao Tong
University. The samples of EC and normal endometrial tissues were
collected after informed consent was signed by the patients.
 | Table I.Clinicopathological parameters of EC
patients. |
Table I.
Clinicopathological parameters of EC
patients.
|
| No. patients
(%) |
|---|
|
|
|
|---|
| Variable | n | % |
|---|
| Total | 71 | 100 |
| Age (years) |
|
≤50 | 15 | 21.1 |
|
>50 | 56 | 78.9 |
| FIGO stage |
| I | 60 | 84.5 |
| II | 7 | 9.8 |
|
III | 4 | 5.7 |
| IV | 0 | 0 |
| Grade
(endometrioid, n=57) |
| G1 | 32 | 56.1 |
| G2 | 18 | 31.6 |
| G3 | 7 | 12.3 |
| Histological
type |
|
Endometrioid | 57 | 80.3 |
|
Non-endometrioid | 14 | 19.7 |
| Myometrial
invasion |
|
<1/2 | 67 | 94.4 |
|
≥1/2 | 4 | 5.6 |
| Lymph node
metastasis |
|
Negative | 63 | 88.7 |
|
Positive | 8 | 11.3 |
Laser capture microdissection
(LCM)
Each specimen was cut into 10–20 serial frozen
sections of 8-µm thickness that were then mounted on polyethylene
membrane glass slides (Applied Biosystems; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) and then stored at −80°C. The
sections were fixed at −20°C for 2 min with 70% ethanol and then
stained with 25 µl HistoGene (Applied Biosystems; Thermo Fisher
Scientific) using a frozen section staining kit. Then, they were
rinsed with ice-cold RNA nuclease-free water at −20°C, incubated in
xylene at −20°C for 2 min and air-dried for 2 min. The targeted
cells were microdissected using a UV cutting and laser capture
procedure with an LCM system (LMD7000; Leica Microsystems GmbH,
Wetzlar, Germany). The EC cells and normal epithelial cells
(Fig. 1A) were cut and captured on
CapSure Macro LCM Cap (Applied Biosystems; Thermo Fisher
Scientific, Inc.). The caps containing the captured cells were
placed into a microfuge tube (Applied Biosystems; Thermo Fisher
Scientific, Inc.) that contained 100 µl of RLT lysis buffer
(Qiagen, Inc., Valencia, CA, USA) and 1% of β-mercaptoethanol (Bio
Basic, Inc., Markham, ON, Canada). For the polymerase chain
reaction (PCR) experiments, each sample collected 10–20 serial
sections of tissue with 10 caps.
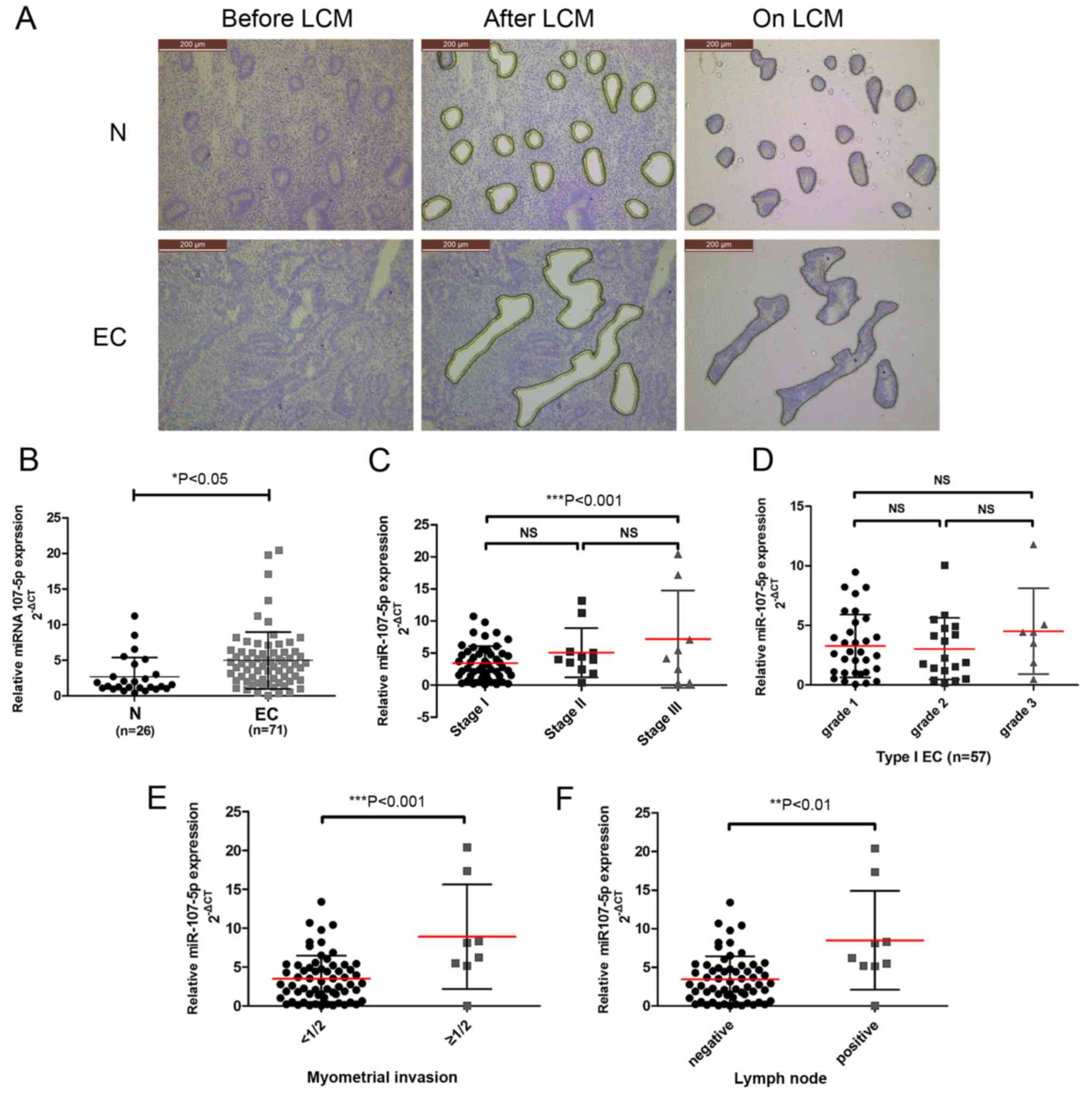 | Figure 1.miR-107-5p expression in EC specimens
and its association with clinical parameters. (A) LCM of normal
endometrial cells and EC cells. Sections were stained with
HistoGene solution. The upper panel reveals frozen sections from
normal endometrium (normal epithelial cells before LCM, after LCM
and on LCM caps) (magnification, ×100). The lower panel reveals
frozen sections from endometrioid endometrial cancer stage IA
(malignant epithelial cells before LCM, after LCM, and on LCM caps)
(magnification, ×100). (B) miR-107-5p was differentially expressed
between the EC specimens (n=71) and normal endometrium (N) (n=26)
as assessed by qRT-PCR. ΔCt describes the expression of miR-107
relative to U6B expression (−ΔCt = CtU6 -
CtmiR-107). *P<0.05, unpaired t-test. (C-F) miR-107
expression was significantly increased in (C) stage III EC patients
compared to stage I patients, (E) myometrial invasion and (F) lymph
node positive disease, but not in (C) stage I vs. stage II patients
or (E) grades (NS, not significant; **P<0.01 and ***P<0.001).
The bars represent the mean ± SD. EC, endometrial carcinoma; LCM,
laser capture microdissection. |
Cell culture
We obtained the human endometrial cancer cell lines
(Ishikawa and HEC-1B) and 293 cells from the Chinese Academy of
Sciences Committee Type Culture Collection (Shanghai, China). The
cell lines were cultured in Dulbecco's modified Eagle's medium/F12
(11030; Gibco; Thermo Fisher Scientific, Inc.) supplemented with
10% fetal bovine serum (FBS) (16000-44; Thermo Fisher Scientific,
Inc.) at 37°C in 5% CO2.
Cell transfections
The miR-107 mimics (miR-107m), miR-107 mimic
negative control (miR-107m NC), miR-107 inhibitor (miR-107i) and
miR-107 inhibitor negative control (miR-107i NC) are synthetic,
chemically modified short single- or double-stranded RNA
oligonucleotides, which were synthesized from Shanghai GenePharma
Co., Ltd. (Shanghai, China) (Table
II). Cells were seeded in 6-well plates at 70–80% confluence
and grown overnight before transfection. Transfection of Ishikawa
cells (or HEC-1B cells) with miR-107i (or miR-107m) or a
counterpart negative control was performed via Lipofectamine 2000
transfection reagent (Invitrogen; Thermo Fisher Scientific, Inc.)
according to the manufacturer's instructions.
| Table II.Oligo sequences used in the
study. |
Table II.
Oligo sequences used in the
study.
| Identifier | Sense primer
sequences | Antisense primer
sequences |
|---|
| miR-107i |
5′-UGAUAGCCCUGUACAAUGCUGCU-3′ |
5′-TCATTGGCATGTACCATGCAGCT-3′ |
| miR-107i NC |
5′-CAGUACUUUUGUGUAGUACAA-3′ |
|
| miR-107m |
5′-AGCAGCAUUGUACAGGGCUAUCA-3′ |
5′-AUAGCCCUGUACAAUGCUGCUUU-3′ |
| miR-107m NC |
5′-UUCUCCGAACGUGUCACGUTT-3′ |
5′-ACGUGACACGUUCGGAGAATT-3′ |
| ESR1-3′UTR-WT |
5′-TGCTCTATGCTGCAAAGACCTGAATACCAC-3′ |
5′-GCTCTATGCTGCTTGTTCCTCCCATAT-3′ |
| ESR1-3′UTR-MT |
5′-TATTACTACGACGATGAAACAAGCAATGC-3′ |
5′-TGAAAAACGATGAATTTCATAG-3′ |
Quantitative real-time reverse
transcription (RT)-PCR
Total RNA was extracted from the cultured cells and
tissues using TRI-reagent (Molecular Research Center, Inc.,
Cincinnati, OH, USA). For the miRNA analysis, TaqMan microRNA
Reverse Transcription kit (Thermo Fisher Scientific, Inc.) was used
to reverse transcribe mature miRNA from total RNA. According to the
manufacturer's instructions, real-time PCR was performed using
TaqMan MicroRNA Assay primers with TaqMan Universal PCR Master Mix
(Thermo Fisher Scientific Inc.) and analyzed with an ABI PRISM7000
Sequence Detection System (Applied Biosystems; Thermo Fisher
Scientific, Inc.). U6B was used as an internal reference for the
expression of miRNAs. The assay names were ‘hsa-miR-107-5p’ for
miR-107-5p and ‘RNU6B’ for U6B miRNA (Table III). All reagents were purchased
from Applied Biosystems; Thermo Fisher Scientific, Inc. For all the
experiments, values on the y-axis were equal 2−ΔCq,
where ΔCq was the difference between gene Cq and normalizer gene Cq
(14). The thermocycling conditions
were as follows: 95°C for 10 min, 45 cycles of 95°C for 15 sec and
60°C for 1 min, for both miRNA and mRNA PCR. The data were obtained
in triplicate in 3 independent experiments.
| Table III.Primers used for quantitative
real-time PCR analysis. |
Table III.
Primers used for quantitative
real-time PCR analysis.
| mRNA/miRNA | Primer
sequence |
|---|
| miR-107 | F:
5′-AGCAGCATTGTACAGGGCTATCA-3′ |
|
| R:
5′-GCGAGCACAGAATTAATACGAC-3′ |
| U6 | F:
5′-AGAGCCTGTGGTGTCCG-3′ |
|
| R:
5′-CATCTTCAAAGCACTTCCCT-3′ |
| ERα (ESR1) | F:
5′-CTCAACAGCGTGTCTCCG-3′ |
|
| R:
5′-GGCTCGTTCTCCAGGTAG-3′ |
| β-actin | F:
5′-CAGCCATGTACGTTGCTATCCAGG-3′ |
|
| R:
5′-AGGTCCAGACGCAGGATGGCATG-3′ |
For quantification of ERα, cDNA was generated using
a PrimeScript RT Reagent kit (Takara Biotechnology Co., Ltd.,
Dalian, China), and real-time PCR was carried out on an ABI PRISM
7000 Sequence Detection System with SYBR Premix Ex Taq (Takara
Biotechnology Co., Ltd.). The primer sequences are presented in
Table II. For all experiments,
values on the y-axis equal 2−ΔΔCq. The data were
obtained in triplicate in 3 independent experiments.
CCK-8 proliferation assays
Infected cells at 70–80% confluence were
serum-starved for 24 h and then cultured in 96-well plates at 5%
CO2 and 37°C with a density of 1,000 cells/well. At
selected time-points, a Cell Counting Kit-8 (CCK-8; Dojindo
Molecular Technologies, Inc., Kumamoto, Japan) assay was applied to
assess the rate of cell proliferation. CCK-8 reagent was added to
the wells, and the absorbance at 450 nm was measured with a
SpectraMax 190 microplate reader (Bio-Rad Model 680; Bio-Rad
Laboratories, Inc., Hercules, CA, USA).
Cell migration and invasion
assays
For wound-healing experiments, cells were seeded in
6-well plates, with 6×106 cells/well. Then, the cells
were allowed to adhere for 24 h. Cell monolayers were scratched
with a sterile micropipette tip and incubated in serum-free medium
for 24 h. For each sample, 3 defined regions were monitored, and
images were captured at 0, 24 and 48 h to assess the migration
index, a value which was obtained by calculating the distance
traveled by the cell monolayer relative to the gap made using the
pipette tip at 0 h (15).
For invasion assays, cells were trypsinized,
centrifuged at 1,000 × g and resuspended in serum-free medium. The
cells were then plated at a density of 2×105 cells/well
in invasion chambers (8-µm pore size; BD Biosciences, Franklin
Lakes, NJ, USA) with Matrigel coating for the invasion assays. The
medium containing 10% FBS was added to 24-well plates as a
chemoattractant. After 6 h of incubation for the migration assay or
after 24 h of incubation for the invasion assay, the cells were
fixed with 4% paraformaldehyde for 1 h. The cells on the apical
side of each insert were removed by mechanical scraping. The cells
that migrated to the basal side of the membrane were stained with
0.1% crystal violet and visualized under a Leica DMI 3000B
microscope (Leica Microsystems GmbH). The cells were counted at a
200-fold magnification.
Western blot analysis
The protein samples were washed with ice-cold PBS
and then lysed in lysis buffer containing 10 mM Tris (pH 7.5), 150
mM NaCl, 10 mM ethylenediaminetetraacetic acid (EDTA), 1% sodium
dodecyl sulfate (SDS), 1 mM sodium orthovanadate, and a mixture of
protease inhibitors (1 mM phenylmethylsulfonyl fluoride, 1 µg/ml
pepstatin A and 2 µg/ml aprotinin). After protein extraction
(Beyotime Institute of Biotechnology, Shanghai, China), all the
sample were detected with BCA protein determination method
according to the protocol (Beyotime Institute of Biotechnology).
Equal amounts (25 µg) of the cell lysates were separated via 8%
sodium dodecyl sulfate-polyacrylamide gel electrophoresis
(SDS-PAGE), and then transferred onto a polyvinylidene difluoride
(PVDF) membrane. Following blocking and washing 4 times for 15 min
with Tris-buffered saline with Tween-20 (TBST) at room temperature,
the PVDF membrane was incubated with rabbit polyclonal primary
antibodies at 1:1,000 to detect ERα (cat. no. ab75635; Abcam,
Cambridge, MA, USA). Following extensive washing, the membranes
were incubated with secondary peroxidase-conjugated goat
anti-rabbit IgG (dilution 1:5,000; cat. no. 7074; Cell Signaling
Technology, Inc., Danvers, MA, USA) for 1 h. The results were
visualized using an enhanced chemiluminescence kit (ECL Kit;
Pierce; Thermo Fisher Scientific, Inc.) using Kodak XAR-5 film
(Sigma-Aldrich; Merck KGaA, Darmstadt, Germany). β-actin was used
as the internal reference.
Construction of reporter plasmids and
luciferase assays
A DNA fragment comprised ofa partial wild-type 3′UTR
of ESR1 or a corresponding mutant 3′UTR of ESR1 was chemically
synthesized and cloned into the pGL3-REPORT luciferase vector
containing Renilla luciferase and firefly luciferase
(Ambion; Thermo Fisher Scientific, Inc.) downstream of the
luciferase gene (designated pGL3-ESR1-3′UTR-WT and
pGL3-ESR1-3′UTR-MT, respectively). The nucleotide sequences of the
plasmid constructs were confirmed by DNA sequencing. All steps of
the luciferase reporter assay were performed in accordance with
procedures previously described (16,17).
Briefly, 293 cells (5×104) were seeded into 24-well
plates and transfected with 0.2 µg of either pGL3-ESR1-3′UTR-WT or
pGL3-ESR1-3′UTR-MT, with or without 50 nM miR-107m or miR-107m NC
using Lipofectamine 2000 transfection reagent according to the
manufacturer's protocol. After 48 h, we used the dual luciferase
reporter assay system to assess the luciferase activity (Promega
Corp., Madison, WI, USA) and the results were expressed as the
relative luciferase activity (firefly LUC/Renilla LUC).
Meanwhile, ERα was the predicted target of miR-107 using the miR
target prediction algorithm TargetScan (http://targetscan.org/), Pictar and miRanda.
Xenograft tumor formation assays
Animal research was conducted in accordance with the
recommendations in the Guidelines for the Care and Use of
Laboratory Animals of China strictly. The protocol was approved by
the Ethics Committee of Animal Experiments of Shanghai General
Hospital Affiliated with Shanghai Jiaotong University School of
Medicine (Permit no. SYXK (hu) 2009-0086). All efforts were made to
minimize suffering. Ten female BALB/c nude mice (5 weeks of age,
~20 g weight) were obtained from the Chinese Academy of Sciences
(Shanghai, China). The mice were housed in an environmentally
controlled room (22±2°C; 40–60% humidity and a 12-h light/dark
cycle). Ishikawa cells were harvested and resuspended at a density
of 5×106 cells/200 µl in sterile saline. Five mice/group
were subcutaneously injected with Ishikawa cells in the subdermal
space on the medial side of the neck. One week after treatment,
when the tumors reached an average volume of ~30 mm3,
they were directly injected with a cocktail of antagomiRs
(Dharmacon, Milan, Italy) targeting miR-107 or with a control
antagomiR [40 ml of phosphate-buffered saline (PBS) containing 1 µg
of each anti-miR-107 or control antagomiR] at days 0, 5 and 9, for
a total of 3 injections/tumor (18). The tumor volume was assessed every 7
days until the end of the experiment, using the formula: largest
diameter × smallest diameter2 ×0.5. The tumor weight was
determined after the animals were sacrificed with cervical
dislocation at the completion of the xenograft experiments.
Immunohistochemistry (IHC)
The 4-µm-thick sections of paraformaldehyde-fixed
and paraffin-embedded xenograft tumor tissue were used for
immunohistochemical examination of ERα expression. The standard
avidin-biotin immunohistochemical techniques with the use of
anti-Ki-67 (dilution 1:100; Wuhan Boster Biological Engineering
Co., Ltd., Wuhan, China) was used to detect Ki-67 expression in the
tumor tissues according to the manufacturer's instructions.
Statistical analyses
Each experiment was performed at least 3 times.
Independent samples t-tests were used to compare the 2 groups and
one-way analysis of variance (ANOVA) followed by post hoc Tukey's
test were performed to compare multiple groups. Data are presented
as the means ± SD. All P-values were two-sided, and P<0.05 was
considered to indicate a statistically significant result. All
statistical analyses were performed using SPSS 16.0 software (SPSS,
Inc., Chicago, IL, USA).
Results
Increased miR-107-5p expression in EC
is associated with myometrial invasion and lymph node
metastasis
To investigate the potential clinicopathological
implications of altered miR-107-5p expression, the expression
levels of miR-107-5p in 26 normal endometrium samples (N) and 71 EC
tissue samples (EC) were compared by TaqMan PCR. While collecting
the endometrial cells captured by LCM (Fig. 1A), it was concluded that the
expression of the miR-107-5p in the EC tissues was significantly
higher than that in the normal endometrium (P<0.05) (Fig. 1B).
To further examine the clinical implications of
miR-107-5p in EC, the association between miR-107-5p expression
levels and clinicopathological characteristics of EC were assessed.
Higher miR-107-5p expression levels were observed in FIGO stage III
tumors than in stage I tumors (P<0.05) (Fig. 1C). There were no statistical
associations found with respect to tumor grade (P>0.05)
(Fig. 1D). However, a statistically
significant association was observed between miR-107-5p expression
and EC myometrial invasion and lymph node metastasis (P<0.001
and P<0.01) (Fig. 1E and F).
These results indicated that reduced miR-107-5p expression was
closely related to advanced FIGO stage, myometrial invasion and
lymph node metastasis in EC.
miR-107-5p promotes the proliferation
of EC cells
To determine the biological role of miR-107-5p, the
effect of modulation of miR-107-5p expression on the proliferation
of EC cells was assessed using miR-107 inhibitor in Ishikawa and
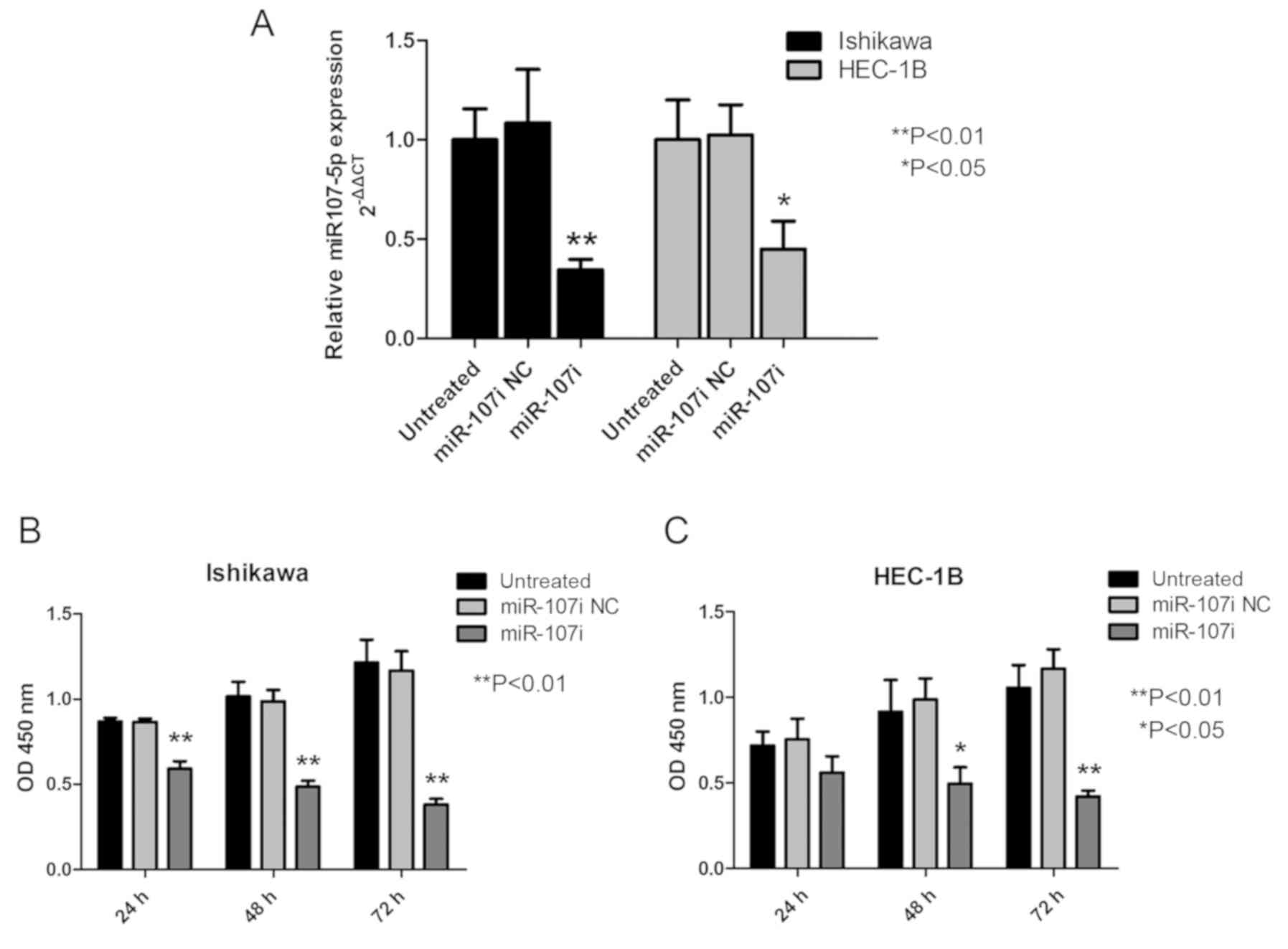
HEC-1B cells. Transfection efficiency was verified at 24 h
post-transfection (Fig. 2A). As
revealed in Fig. 2B, compared with
miR-107i NC and untreated cells, treatment with miR-107i
significantly inhibited the growth of the Ishikawa cells as
assessed by the CKK-8 assay (Fig.
2B; P<0.01). HEC-1B cells treated with miR-107i grew more
slowly than untreated HEC-1B cells and miR-107i NC (Fig. 2B; P<0.05). All the aforementioned
results revealed that miR-107-5p promoted EC cell
proliferation.
miR-107-5p enhances the migration and
invasion of EC cells
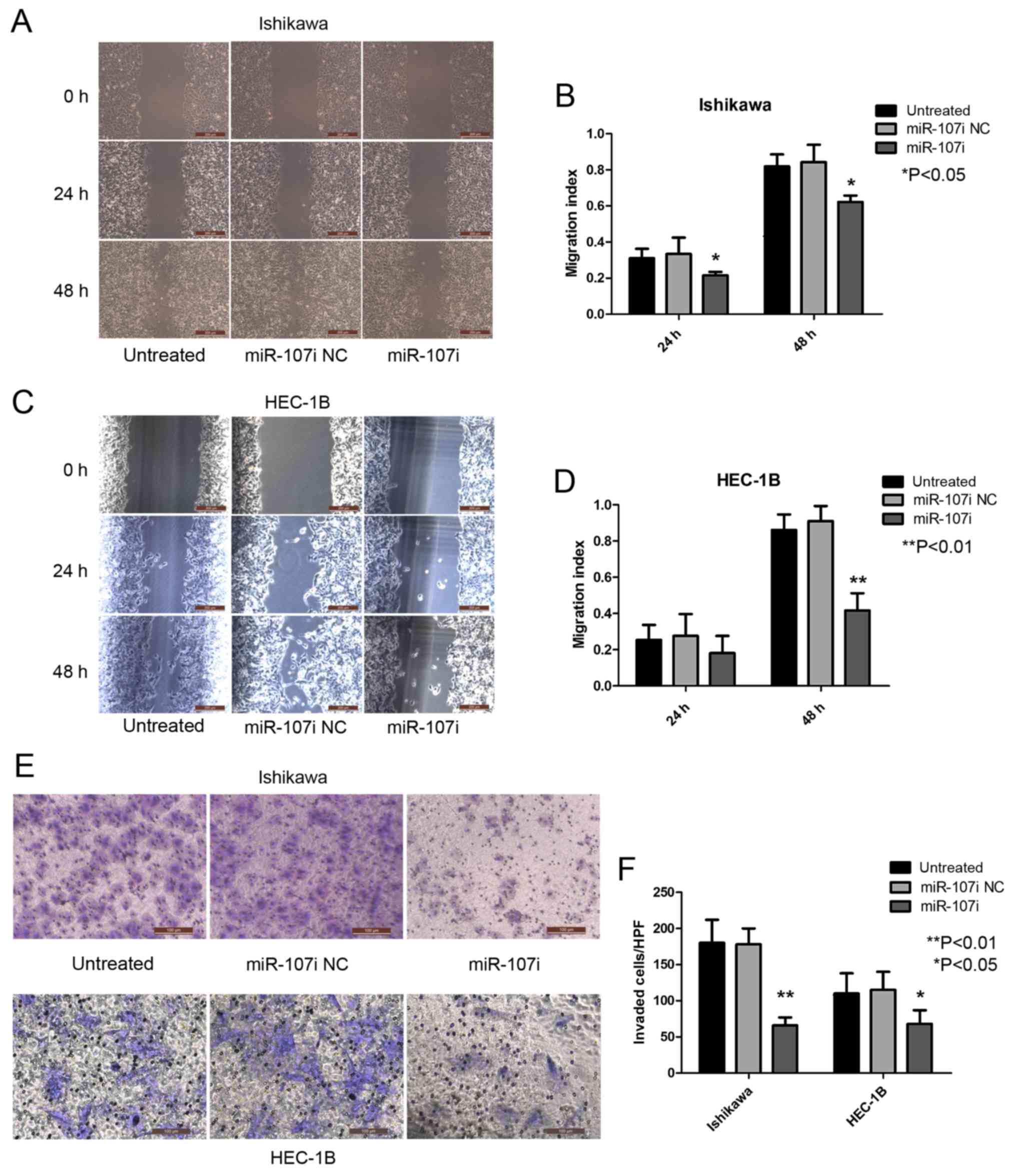
To explore the role of miR-107-5p in the regulation
of metastatic function, we used wound-healing and Transwell
invasion assays to examine the migration and invasion abilities of
Ishikawa and HEC-1B cells after transfection with miR-107
inhibitor. Untreated Ishikawa and miR-107 NC cells displayed an
enhanced rate of migration compared with Ishikawa cells transfected
with miR-107i (Fig. 3A and B;
P<0.05). HEC-1B cells transfected with miR-107i exhibited a low
migration rate compared with untreated HEC-1B cells and miR-107i NC
cells (Fig. 3C and D; P<0.01).
These results indicated that miR-107-5p enhanced EC cell
migration.
In accordance with the aforementioned results, the
invasion rate of Ishikawa and HEC-1B cells transfected with
miR-107i was significantly lower than that of untreated cells and
miR-107i NC cells (Fig. 3E and F;
P<0.01 and P<0.05). These results demonstrate a functional
role for miR-107-5p in regulating migration and invasion in EC
cells.
ERα is a functional target of
miR-107-5p
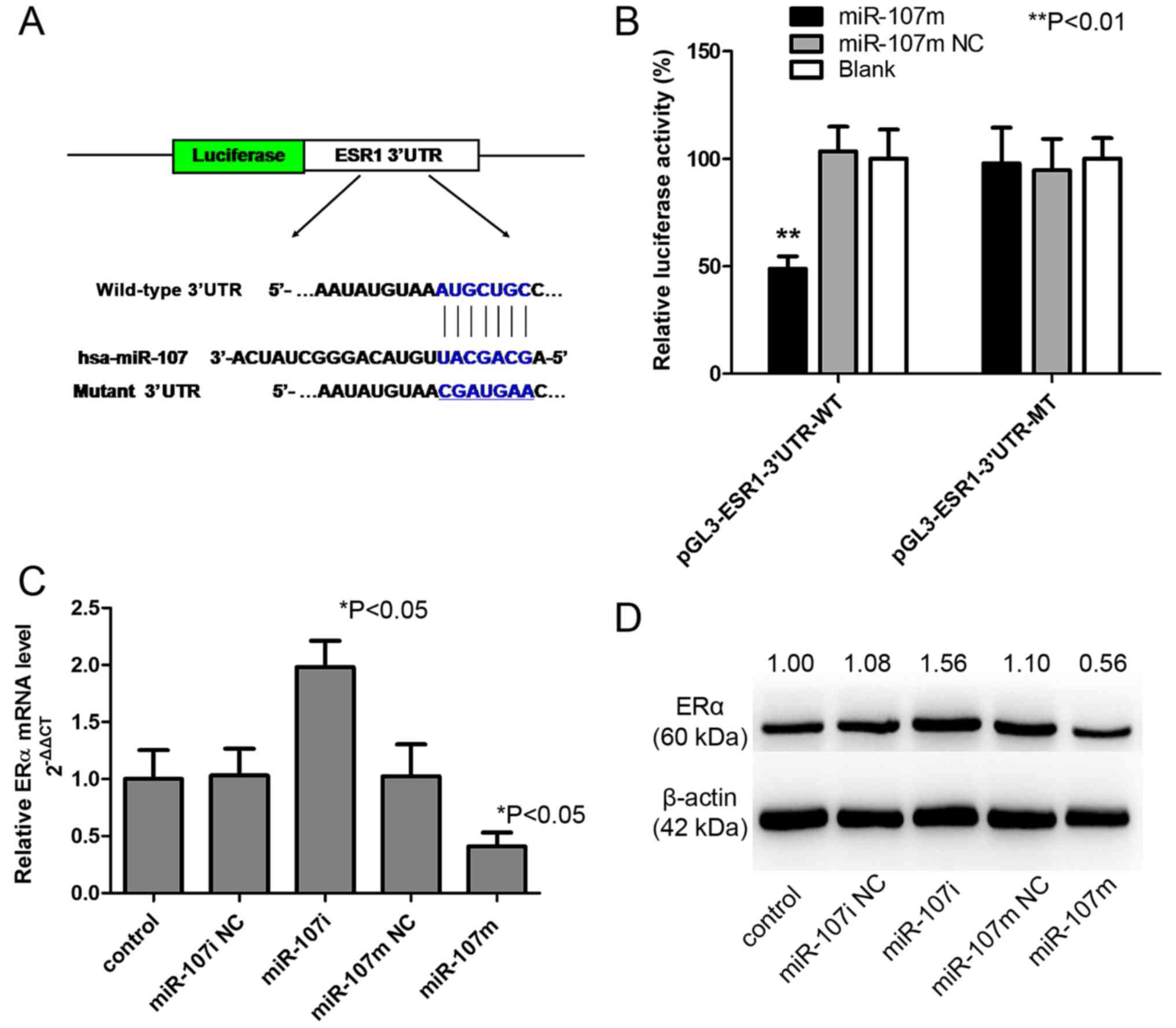
To further illuminate the underlying mechanism of
miR-107-5p in EC carcinogenesis, the target of miR-107-5p must be
determined. Notably, by searching for the potential targets of
miR-107, we found the miR-107-5p targeting site within the 3′UTR of
ESR1, as detected by 3 software algorithms (TargetScan, Pictar and
miRanda) (data not shown). To directly examine whether this site
was responsible for the regulation of ERα expression by miR-107-5p,
a vector containing wild-type or mutant ESR1 3′UTR was constructed
and directly fused downstream of firefly luciferase reporter gene
(Fig. 4A). The wild-type or mutant
vector was co-transfected into 293 cells with miR-107m or miR-107m
NC. As revealed in Fig. 4B, miR-107
significantly decreased the relative luciferase activity of the
wild-type ESR1 3′UTR (>50% inhibition, P<0.01), whereas the
miR-107 NC had no significant effect. This inhibition was specific
to the wild-type 3′UTR sequence, given that no reduction in
luciferase activity was observed for the mutant ESR1 3′UTR.
Collectively, these results indicated that ERα is a direct
downstream target of miR-107-5p in 293 cells.
To further investigate the functional significance
of the deregulated miR-107-5p observed in EC cell lines, Ishikawa
cells were transfected with a control mimic/inhibitor (miR-107m
NC/miR-107i NC) or miR-107 mimic/miR-107 inhibitor
(miR-107m/miR-107i), and the effects on the expression of ERα were
examined. Expression of miR-107m significantly decreased the
expression of ERα at both the mRNA (Fig. 4C) and protein levels (Fig. 4D). In contrast, inhibition of
miR-107-5p significantly increased the expression of ERα mRNA
(Fig. 4C) and protein (Fig. 4D), further supporting its role as a
functional suppressor of ERα.
Blocking miR-107-5p inhibits
tumorigenicity in vivo
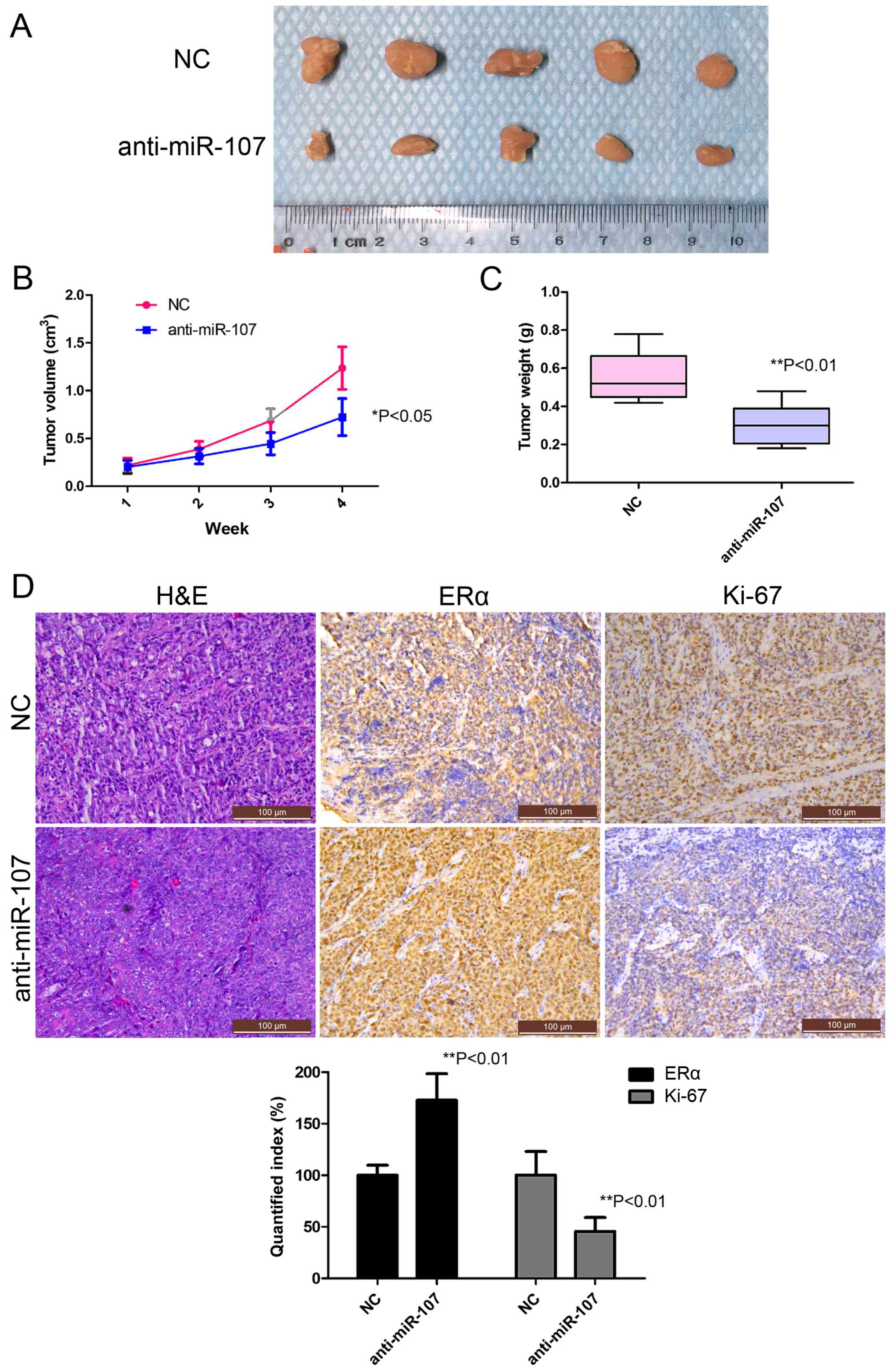
To examine the potential role of miR-107-5p in
tumorigenicity, we further assessed the feasibility of direct
anti-miR-107 treatment in vivo. Ishikawa cells were
subcutaneously injected into the flanks of nude mice in order to
yield tumors that were then treated by direct intratumoral
injection as soon as they became clearly palpable. For the treated
group, the tumor on the flank was injected with a mixture of
anti-miR-107 antagomiR, while for the control group, the tumor was
injected with a control antagomiR. Tumor progression was monitored
over a 28-day course. Inhibition of miR-107-5p caused a marked
reduction in tumor size and tumor weight compared with the NC group
over the time period tested (Fig.
5A-C). To ascertain the effects on protein expression, tumor
tissue was embedded in paraffin and then stained with H&E for
histological examination. As expected, Ki-67 levels were detectable
in the control xenografts, while they were significantly decreased
in the treated group (Fig. 5D),
while ERα levels were significantly increased in the treated group
(Fig. 5D, lower panel), thus
confirming the role of miR-107-5p as a negative regulator of the
miR-107/ERα pathway in vivo.
Discussion
In Western countries, endometrial carcinoma (EC) is
the most common cancer of the female reproductive system. EC occurs
mainly in peri- and postmenopausal women, although it may also be
present in premenopausal women, particularly in the context of
hyperestrogenism. From a clinical point of view, EC is classified
into two different types: types I and II. Type I tumors are
low-grade and estrogen-associated endometrioid ECs, which usually
develop in perimenopausal women and coexists with or prior to
endometrial hyperplasia (19).
These tumors often occur in the environment of estrogen
overexposure and early diagnosis has a good prognosis. However, the
incidence rate of EC has risen over the last decade (20). In addition, due to obesity and
physical inactivity, the onset age of patients is earlier. For
early disease, surgical intervention with or without adjuvant whole
pelvic radiation is the standard treatment, whereas the use of
progestogen-based endocrine therapy is mainly restricted to
patients with terminal or recurrent cancer. Non-invasive targeted
therapies without side-effects or resistance have yet to be found
(21).
ERα is an important mediator of estrogenic action in
the female genital tract. E2 and its receptors are
closely related to a variety of human diseases, including
hormone-dependent tumors (22,23).
Estrogens promote physiological actions after binding to their
estrogen receptor (ER) subtypes (ERα and ERβ). The subtype ERα
exists on the surface of the cell membrane and activates rapid
E2-ER signaling. Amplification of the ERα gene is
regarded as a common phenomenon in EC (24). Notably, presently, there is
increasing coherent evidence supporting the view that ERα
expression decreases in late stage and poorly differentiated ECs
(25,26). Multiple hypotheses have been
proposed to explain the ERα deletion, including ERα gene
polymorphism (27),
hypermethylation of CpG islands (28), and overexpression of estrogen
receptor-associated receptors (29). However, the underlying regulatory
mechanisms and the downstream biological effects of decreased
expression of ERα on EC malignancy remain unclear.
Epigenetic effects are a key aspect of the
relationship between miRNAs and carcinogenesis (30). Modulated expression of miRNAs has
recently been related to the carcinogenesis of EC (31–33).
Numerous studies have demonstrated the distinct miRNA expression
profiles between EC and normal cells (32,34).
Boren et al studied miRNA expression in EC, normal
endometrium and complex atypical hyperplasia and found 13 miRNAs
associated with EC (31). Then,
Chung et al studied miRNA expression in endometrioid EC
cells and normal cells of women from Hong Kong including
endometrium in the proliferative and secretory phases as well as
the postmenopausal atrophic endometrium. Thirty miRNAs were
significantly upregulated in EC cells (33). This finding indicated that the miRNA
distribution of cancer cells and normal cells is different.
Notably, four types of miRNAs, namely, miR-103, miR-106a, miR-107
and miR-210, were found to be deregulated in both studies,
highlighting their potential role in the pathogenesis of disease.
In those four deregulated miRNAs, miR-107 piqued our interest since
ERα was the predicted target of miR-107 using the miR target
prediction algorithm TargetScan (http://targetscan.org/).
Recent studies indicate that the role of miR-107 in
tumor development and progression is contradictory and differs in a
context-dependent manner. miR-107 may play the role of a tumor
suppressor gene by inducing cell cycle arrest in lung cancer and
glioma (35,36) whereas it may be an oncogene that
promotes tumor invasion and metastasis in breast and gastric cancer
(37,38). However, the role of miR-107 in the
progression and metastasis of EC remains unclear. In order to
confirm the relationship between the expression of miR-107 and the
progression and metastasis of EC, miR-107-5p expression with RT-PCR
in 71 EC patients was examined and it was revealed that miR-107-5p
was significantly upregulated in EC compared with normal
endometrium. The upregulation of miR-107 was associated with an
increased and more advanced histological grade and lymph node
metastasis. Functionally, our data revealed that miR-107 promoted
the proliferation and invasion of EC cells. Next, we found that
miR-107 was able to directly bind to the 3′UTR of ESR1 mRNA in EC
cells. Moreover, we observed that miR-107 could inhibit the
expression of ERα at the mRNA and protein levels in the cells,
indicating that miR-107 plays an important role in EC through
ERα.
Some limitations in the present study should be
addressed. Firstly, in a loss-of-function model, the function of
miR-107 was evaluated by miR-107 gene knockout. Gain-of-function
studies with overexpression of miR-107 in EC cell lines are
required to verify our findings. Secondly, in order to validate the
role of miR-107 in tumor development and progression, the mechanism
warrants further study. It has been reported by Zhou et al
that miR-107 activated the ATR/Chk1 pathway and suppressed cervical
cancer invasion by targeting MCL1 (39).
In conclusion, our data demonstrated that miR-107-5p
targeted ERα and that suppression of miR-107 decreased the
proliferation, migration and invasion of EC in vitro and
in vivo. It is thus proposed that miR-107 may serve as a
useful therapeutic strategy for EC.
Acknowledgements
Not applicable.
Funding
The present study was sponsored by the National
Natural Science Foundation of China (nos. 81402134 and 81502232),
the Shanghai PuJiang Program (no. 17PJD032) and the Project Young
Elite of the Shanghai Health System (no. 2017YQ063).
Availability of data and materials
The datasets used during the present study are
available from the corresponding author upon reasonable
request.
Authors' contributions
WB, SW and YZhu conceived and designed the study.
WB, YZhang, SL, QF, MQ, YW, YL, XJ and YeY performed the
experiments. SL, QF, MQ, YW, YL, XJ, YeY, ZS, WX and YoY analyzed
tha data. WB, YZhang, ZS, WX and YoY wrote the manuscript. WB,
YZhang, SW and YZhu reviewed and edited the manuscript. All authors
read and approved the manuscript and agree to be accountable for
all aspects of the research in ensuring that the accuracy or
integrity of any part of the work are appropriately investigated
and resolved.
Ethics approval and consent to
participate
The present study was approved by the Human
Investigation Ethical Committee of Shanghai General Hospital
Affiliated with Shanghai Jiao Tong University. The samples of EC
and normal endometrial tissues were collected after informed
consent was signed by the patients. All experimental protocols were
approved by the Ethics Committee of Animal Experiments of Shanghai
General Hospital Affiliated with Shanghai Jiaotong University
School of Medicine (Permit no. SYXK (hu) 2009-0086).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2018. CA Cancer J Clin. 68:7–30. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bokhman JV: Two pathogenetic types of
endometrial carcinoma. Gynecol Oncol. 15:10–17. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Salvesen HB, Haldorsen IS and Trovik J:
Markers for individualised therapy in endometrial carcinoma. Lancet
Oncol. 13:e353–e361. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Setiawan VW, Yang HP, Pike MC, McCann SE,
Yu H, Xiang YB, Wolk A, Wentzensen N, Weiss NS, Webb PM, et al:
Type I and II endometrial cancers: Have they different risk
factors? J Clin Oncol. 31:2607–2618. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Chuffa LG, Lupi-Júnior LA, Costa AB,
Amorim JP and Seiva FR: The role of sex hormones and steroid
receptors on female reproductive cancers. Steroids. 118:93–108.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Rosenfeld MG and Glass CK: Coregulator
codes of transcriptional regulation by nuclear receptors. J Biol
Chem. 276:36865–36868. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lu J, Getz G, Miska EA, Alvarez-Saavedra
E, Lamb J, Peck D, Sweet-Cordero A, Ebert BL, Mak RH, Ferrando AA,
et al: MicroRNA expression profiles classify human cancers. Nature.
435:834–838. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Banno K, Yanokura M, Kisu I, Yamagami W,
Susumu N and Aoki D: MicroRNAs in endometrial cancer. Int J Clin
Oncol. 18:186–192. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Bao W, Wang HH, Tian FJ, He XY, Qiu MT,
Wang JY, Zhang HJ, Wang LH and Wan XP: A TrkB-STAT3-miR-204-5p
regulatory circuitry controls proliferation and invasion of
endometrial carcinoma cells. Mol Cancer. 12:1552013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Chen L, Li ZY, Xu SY, Zhang XJ, Zhang Y,
Luo K and Li WP: Upregulation of miR-107 inhibits glioma
angiogenesis and VEGF expression. Cell Mol Neurobiol. 36:113–120.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lu C, Xie Z and Peng Q: MiRNA-107 enhances
chemosensitivity to paclitaxel by targeting antiapoptotic factor
Bcl-w in non small cell lung cancer. Am J Cancer Res. 7:1863–1873.
2017.PubMed/NCBI
|
|
13
|
Creasman W: Revised FIGO staging for
carcinoma of the endometrium. Int J Gynaecol Obstet. 105:1092009.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2ΔΔCT method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Bao W, Qiu H, Yang T, Luo X, Zhang H and
Wan X: Upregulation of TrkB promotes epithelial-mesenchymal
transition and anoikis resistance in endometrial carcinoma. PLoS
One. 8:e706162013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Chen J, Zhou X, Xiao Q, Wang T, Shao G, Li
Y and Zhang Z: MiR-107 suppresses cell proliferation and tube
formation of Ewing sarcoma cells partly by targeting HIF-1β. Hum
Cell. 31:42–49. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wang Y, Chen F, Zhao M, Yang Z, Zhang S,
Ye L, Gao H and Zhang X: MiR-107 suppresses proliferation of
hepatoma cells through targeting HMGA2 mRNA 3′UTR. Biochem Biophys
Res Commun. 480:455–460. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Mercatelli N, Coppola V, Bonci D, Miele F,
Costantini A, Guadagnoli M, Bonanno E, Muto G, Frajese GV, De Maria
R, et al: The inhibition of the highly expressed miR-221 and
miR-222 impairs the growth of prostate carcinoma xenografts in
mice. PLoS One. 3:e40292008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yeramian A, Moreno-Bueno G, Dolcet X,
Catasus L, Abal M, Colas E, Reventos J, Palacios J, Prat J and
Matias-Guiu X: Endometrial carcinoma: Molecular alterations
involved in tumor development and progression. Oncogene.
32:403–413. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Schouten LJ, Goldbohm RA and van den
Brandt PA: Anthropometry, physical activity, and endometrial cancer
risk: Results from the Netherlands cohort study. Int J Gynecol
Cancer. 16 Suppl 2:S4922006. View Article : Google Scholar
|
|
21
|
Dedes KJ, Wetterskog D, Ashworth A, Kaye
SB and Reis-Filho JS: Emerging therapeutic targets in endometrial
cancer. Nat Rev Clin Oncol. 8:261–271. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Chen WY: Exogenous and endogenous hormones
and breast cancer. Best Pract Res Clin Endocrinol Metab.
22:573–585. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chuffa LG, Seiva FR, Fávaro WJ, Amorim JP,
Teixeira GR, Mendes LO, Fioruci-Fontanelli BA, Pinheiro PF,
Martinez M and Martinez FE: Melatonin and ethanol intake exert
opposite effects on circulating estradiol and progesterone and
differentially regulate sex steroid receptors in the ovaries,
oviducts, and uteri of adult rats. Reprod Toxicol. 39:40–49. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lebeau A, Grob T, Holst F,
Seyedi-Fazlollahi N, Moch H, Terracciano L, Turzynski A, Choschzick
M, Sauter G and Simon R: Oestrogen receptor gene (ESR1)
amplification is frequent in endometrial carcinoma and its
precursor lesions. J Pathol. 216:151–157. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Jongen V, Briët J, de Jong R, ten Hoor K,
Boezen M, van der Zee A, Nijman H and Hollema H: Expression of
estrogen receptor-alpha and -beta and progesterone receptor-A and
-B in a large cohort of patients with endometrioid endometrial
cancer. Gynecol Oncol. 112:537–542. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Rahman MT, Nakayama K, Rahman M, Ishikawa
M, Katagiri H, Katagiri A, Ishibashi T, Sato E, Iida K, Ishikawa N,
et al: ESR1 gene amplification in endometrial carcinomas: A
clinicopathological analysis. Anticancer Res. 33:3775–3781.
2013.PubMed/NCBI
|
|
27
|
Ohshiro K, Mudvari P, Meng QC, Rayala SK,
Sahin AA, Fuqua SA and Kumar R: Identification of a novel estrogen
receptor-alpha variant and its upstream splicing regulator. Mol
Endocrinol. 24:914–922. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Giacinti L, Claudio PP, Lopez M and
Giordano A: Epigenetic information and estrogen receptor alpha
expression in breast cancer. Oncologist. 11:1–8. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Pelekanou V, Kampa M, Gallo D, Notas G,
Troullinaki M, Duvillier H, Jacquot Y, Stathopoulos EN, Castanas E
and Leclercq G: The estrogen receptor alpha-derived peptide ERα17p
(P295-T311) exerts pro-apoptotic actions in
breast cancer cells in vitro and in vivo, independently from their
ERα status. Mol Oncol. 5:36–47. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Sharma S, Kelly TK and Jones PA:
Epigenetics in cancer. Carcinogenesis. 31:27–36. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Boren T, Xiong Y, Hakam A, Wenham R, Apte
S, Wei Z, Kamath S, Chen DT, Dressman H and Lancaster JM: MicroRNAs
and their target messenger RNAs associated with endometrial
carcinogenesis. Gynecol Oncol. 110:206–215. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wu W, Lin Z, Zhuang Z and Liang X:
Expression profile of mammalian microRNAs in endometrioid
adenocarcinoma. Eur J Cancer Prev. 18:50–55. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Chung TK, Cheung TH, Huen NY, Wong KW, Lo
KW, Yim SF, Siu NS, Wong YM, Tsang PT, Pang MW, et al: Dysregulated
microRNAs and their predicted targets associated with endometrioid
endometrial adenocarcinoma in Hong Kong women. Int J Cancer.
124:1358–1365. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Hiroki E, Akahira J, Suzuki F, Nagase S,
Ito K, Suzuki T, Sasano H and Yaegashi N: Changes in microRNA
expression levels correlate with clinicopathological features and
prognoses in endometrial serous adenocarcinomas. Cancer Sci.
101:241–249. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Chen L, Zhang R, Li P, Liu Y, Qin K, Fa
ZQ, Liu YJ, Ke YQ and Jiang XD: P53-induced microRNA-107 inhibits
proliferation of glioma cells and down-regulates the expression of
CDK6 and Notch-2. Neurosci Lett. 534:327–332. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Takahashi Y, Forrest AR, Maeno E,
Hashimoto T, Daub CO and Yasuda J: MiR-107 and MiR-185 can induce
cell cycle arrest in human non small cell lung cancer cell lines.
PLoS One. 4:e66772009. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Li X, Zhang Y, Shi Y, Dong G, Liang J, Han
Y, Wang X, Zhao Q, Ding J, Wu K, et al: MicroRNA-107, an oncogene
microRNA that regulates tumour invasion and metastasis by targeting
DICER1 in gastric cancer. J Cell Mol Med. 15:1887–1895. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Martello G, Rosato A, Ferrari F, Manfrin
A, Cordenonsi M, Dupont S, Enzo E, Guzzardo V, Rondina M, Spruce T,
et al: A MicroRNA targeting dicer for metastasis control. Cell.
141:1195–1207. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Zhou C, Li G, Zhou J, Han N, Liu Z and Yin
J: miR-107 activates ATR/Chk1 pathway and suppress cervical cancer
invasion by targeting MCL1. PLoS One. 9:e1118602014. View Article : Google Scholar : PubMed/NCBI
|