Introduction
Breast cancer is a heterogeneous disease, presenting
with distinctive clinical features, behavior and responses to
therapy. An estimated 70–80% of breast cancer cases express
estrogen receptor (ER) (1–3). The therapeutic management of
ER-positive cancer includes the application of tamoxifen, which is
a selective ER modulator that competes with estrogen for binding
with ER. Theoretically, patients with ER-positive breast cancer
should benefit from treatment with tamoxifen and other endocrine
agents (such as aromatase inhibitors). However, a proportion of
breast cancer cases fail to initially respond, whereas certain
types of breast cancer develop acquired resistance to endocrine
therapy over time. Novel therapeutic approaches are required to
enhance the efficacy of endocrine therapy and overcome resistance
exhibited by breast cancer.
Mammalian target of rapamycin (mTOR), a
serine/threonine protein kinase and a downstream member of the
phosphoinositide 3-kinase (PI3K)/protein kinase B (Akt) signaling
pathway, is a crucial regulator of cell proliferation and
metabolism. A previous study identified that ER-positive cells
exposed to long-term culture in estrogen-deprived medium became
more dependent on the mTOR signaling pathway for proliferation and
exhibit greater sensitivity to inhibition of the mTOR signaling
pathway (4), which suggests that
inhibition of this signaling pathway using an mTOR inhibitor may
restore sensitivity to tamoxifen. Rapamycin and its analogs
represent promising candidates for the suppression of mTOR pathway
activity.
Bcl-2-associated athanogene 1 (BAG-1) is a
multifunctional protein that interacts with a wide range of
molecules and subsequently affects various cellular functions
(5–10). BAG-1 protects cells from apoptosis
otherwise induced by treatment with cytotoxic drugs, growth factor
withdrawal, radiation and stress (11–15).
BAG-1 is expressed in three main isoforms, which differ in their
translation initiation starting positions (16,17).
BAG-1 long (BAG-1L; p50) has been reported to enhance ER function
and to be associated with disease outcomes in patients with
ER-positive breast cancer receiving endocrine therapy (18,19).
BAG-1 mRNA has also been revealed to represent a prognostic
biomarker in Oncotype DX (20).
Although the significance of BAG-1 as a biomarker in ER-positive
breast cancer has been well-established, little is known about the
molecular mechanism underlying the association between BAG-1 and
the cellular response to tamoxifen. In the present study, BAG-1
expression was assessed in clinical breast cancer specimens. In
breast cancer cells, multiple approaches were used to investigate
the function of BAG-1 in endocrine therapy and to investigate the
underlying molecular mechanisms of the association of BAG-1 with
breast cancer endocrine resistance.
Materials and methods
Tumor tissues and immunohistochemistry
analysis
Human breast cancer tissue microarrays (TMAs) were
purchased from Shanghai Outdo Biotech Co., Ltd. (Shanghai, China;
cat. no. HBre-Duc159Sur-01). Informed consent was obtained from all
patients according to Shanghai Outdo Biotech Co., Ltd., and the
study protocol was approved by the Ethics Committee of Tianjin
Medical University Cancer Institute and Hospital (Tianjin, China).
The TMA slides included 119 cases of invasive ductal carcinoma of
stages I–III, and 40 cases of normal adjacent tissue. Slides (5-µm)
were deparaffinized and rehydrated through a graded series of
ethanol. Following antigen retrieval, slides were blocked for
endogenous peroxidase activity in 3% H2O2 and
were blocked in 5% normal goat serum blocking solution (OriGene
Technologies, Inc., Beijing, China) for 1 h at room temperature.
Tissue sections were incubated with rabbit anti-BAG-1 primary
antibodies (1:100; cat. no. ab32109; Abcam, Cambridge, UK)
overnight at 4°C. Slides were incubated with a biotinylated mouse
anti-hemagglutinin tag rabbit secondary antibody (1:1,000; cat. no.
TA183062; OriGene Technologies, Inc.) for 2 h at room temperature.
Each section was treated with horseradish peroxidase-conjugated
streptavidin (OriGene Technologies, Inc.) and subsequently stained
using 3,3′-diaminobenzidine (1:20) at room temperature. The nuclei
were counterstained with hematoxylin for 1 min at room
temperature.
Staining of BAG-1 in the nucleus and cytoplasm was
described in terms of intensity (0, absent; 1+, weak; 2+, moderate;
3+, intense) and the total percentage of cells exhibiting positive
staining (0–100%). Calculation of H-scores was on the basis of the
percentage of positively stained cells multiplied by the staining
intensity score, with H-scores ranging between 0 and 300 (18). An H-score of >100 was considered
to indicate a positive result. Evaluation was carried out by four
independent observers.
Cell culture and treatment
T47D breast cancer cells (American Type Culture
Collection, Manassas, VA, USA) were maintained in RPMI-1640 medium
(Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA) with 10%
fetal bovine serum (Gibco; Thermo Fisher Scientific, Inc.) and 100
U/100 µg penicillin/streptomycin (Gibco; Thermo Fisher Scientific,
Inc.) at 37°C in a humidified 5% CO2 incubator (Sanyo
Electric Co., Ltd., Osaka, Japan). 4-Hydroxytamoxifen (4-OH TAM),
an active metabolite of tamoxifen, and rapamycin were purchased
from Sigma-Aldrich; Merck KGaA (Darmstadt, Germany).
Cell transfection
BAG-1 short interfering RNA (siRNA; Guangzhou
RiboBio Co., Ltd., Guangzhou, China) sequences used for
transfection were as follows: 5′-CCACAAUAGAGCAGUUUAU-3′ (sense) and
3′-GGUGUUAUCUCGUCAAAUA-5′ (antisense). A scrambled siRNA (Guangzhou
RiboBio Co., Ltd.) was used in parallel. Overexpression of BAG-1L
was achieved by transfecting GV144-BAG-1 (Shanghai GeneChem Co.,
Ltd., Shanghai, China) into T47D cells and T47D cells transfected
with an empty vector GV144 (cytomegalovirus-enhanced green
fluorescent protein-multiple cloning site-simian virus 40-neomycin)
were used as a control. Transfection was performed using
Lipofectamine™ 3000 reagent (Thermo Fisher Scientific, Inc.),
according to the manufacturer's protocol. Briefly, between
2×105 and 3×105 cells/well were seeded into
6-well plates and incubated overnight in medium in the absence of
antibiotics. The following day, 2–4 µg plasmids were mixed with
Lipofectamine™ 3000, and the mixture was subsequently added to each
well. The cells were then incubated at 37°C in a CO2
incubator for 24 h. Following incubation, cells were prepared for
subsequent assays.
Western blot analysis
Cells were lysed on ice using
radioimmunoprecipitation assay mammalian cell lysis reagent
(Beijing Solarbio Science & Technology Co., Ltd., Beijing,
China) containing phenylmethylsulfonyl fluoride (Sigma; Merck
KGaA). Subsequently, cell lysates were centrifuged at 16,000 × g
for 20 min at 4°C, and the protein concentration was subsequently
determined using a Bicinchoninic Acid assay kit (Beijing Solarbio
Science & Technology Co., Ltd.). Following separation of
denatured proteins (20–50 µg) by SDS-PAGE (8–15% gels), proteins
were transferred onto a polyvinylidene difluoride membrane using a
transfer system (Bio-Rad Laboratories, Inc., Hercules, CA, USA) at
250 mA for 1–3 h, and the membrane was blocked in 5% non-fat milk
for 1 h at room temperature prior to incubation with the following
primary antibodies overnight at 4°C: Anti-BAG-1 (1:500; cat. no.
3920; Cell Signaling Technology, Inc., Danvers, MA, USA), anti-ER
(1:1,000; cat. no. 8644; Cell Signaling Technology, Inc.),
anti-mTOR (1:1,000; cat. no. 2983; Cell Signaling Technology,
Inc.), anti-phospho (p-)mTOR (1:1,000; cat. no. 2971; Cell
Signaling Technology, Inc.), anti-Akt (1:1,000; cat. no. 9272; Cell
Signaling Technology, Inc.), anti-p-Akt (Ser473;
1:1,000; cat. no. 4060; Cell Signaling Technology, Inc.) and
anti-β-actin (1:10,000; cat. no. sc-47778; Santa Cruz
Biotechnology, Inc., Dallas, TX, USA). The membrane was washed
three times in TBST (Tris-buffered saline containing 0.05%
Tween-20) and incubated with horseradish peroxidase-conjugated goat
anti-mouse (1:3,000; cat. no. 7076; Cell Signaling Technology,
Inc.) or anti-rabbit secondary antibodies (1:3,000; cat. no. 7074;
Cell Signaling Technology, Inc.) for 2 h at room temperature.
Following washing with TBST, proteins were visualized using the
Enhanced Chemiluminescence detection reagent (EMD Millipore,
Billerica, MA, USA) and analyzed using a Gel Doc 1000 instrument
(Bio-Rad Laboratories, Inc.). All western blotting was performed
three times.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted using TRIzol®
reagent (Invitrogen; Thermo Fisher Scientific, Inc.), according to
the manufacturer's protocol. cDNA was synthesized from 0.5 µg total
RNA using a PrimeScript RT reagent kit (Takara Bio, Inc., Otsu,
Japan), according to the manufacturer's protocol. qPCR was carried
out in triplicate using a CFX96 real-time PCR system (Bio-Rad
Laboratories, Inc.) and SYBR Premix Ex Taq II (Takara Bio, Inc.).
The cycling conditions were 50°C for 2 min, 95°C for 10 min,
followed by 40 cycles of 95°C for 15 sec and 60°C for 30 sec.
β-actin was used as an internal control. Relative quantification of
BAG-1 and ER expression levels was performed using the
2−ΔΔCq method (21). The
sequences of the primers used were as follows: BAG-1,
5′-GTTCTTTGGATGGAGCCTGTG-3′ (forward) and
5′-TGCCTGCTTTACTCATTCTGGTG-3′ (reverse); ERα,
5′-TCAGGCACATGAGTAACAAAGG-3′ (forward) and
5′-AAGGAATGCGATGAAGTAGAGC-3′ (reverse); β-actin,
5′-TGACGTGGACATCCGCAAACG-3′ (forward) and
5′-CTGGAAGGTGGACAGCGAGG-3′ (reverse).
Cell cycle and apoptosis analysis
Cells were incubated in RPMI-1640 medium with 10%
fetal bovine serum with 10 µmol/l 4-OH TAM and/or 10 µmol/l
rapamycin. Cells were harvested after 48 h by trypsinization.
Cell cycle analysis was investigated using a cell
cycle detection kit (Nanjing KeyGen Biotech Co., Ltd., Nanjing,
China). Following harvesting of drug-treated cells, the cells were
collected and fixed using ice-cold 70% ethanol in PBS for 24 h at
4°C. Cells were treated with RNase A for 30 min, labeled with
propidium iodide for 30 min at 37°C in the dark and subsequently
analyzed via flow cytometry. Cell apoptosis assays were performed
using an Annexin V-allophycocyanin (APC)/7-aminoactinomycin D
(7-AAD) nuclear peridinin-chlorophyll protein complex (PerCP)
Apoptosis Detection kit (Nanjing KeyGen Biotech Co., Ltd.),
according to the manufacturer's protocol. Cells were resuspended in
binding buffer, incubated with Annexin V-APC followed by 7-AAD at
room temperature for 15 min in the dark and subsequently analyzed
by flow cytometry using a FACSCanto II flow cytometer (BD
Biosciences, San Jose, CA, USA). All experiments were performed in
triplicate.
Statistical analysis
All statistical analyses were performed using
GraphPad Prism (version 6.02; GraphPad Software, Inc., La Jolla,
CA, USA). A χ2 test was used to determine statistically
significant differences among clinicopathological features. Data
are presented as the mean ± standard deviation. Student's t-test or
one-way analysis of variance followed by Tukey's multiple
comparisons test was used to determine the differences between
groups. P<0.05 was considered to indicate a statistically
significant difference.
Results
BAG-1 is highly expressed in
ER-positive breast cancer
To assess the function of BAG-1 and its association
with ER status in human breast cancer, immunohistochemistry was
performed using TMAs including 119 human breast invasive ductal
carcinoma cases (Table I). The
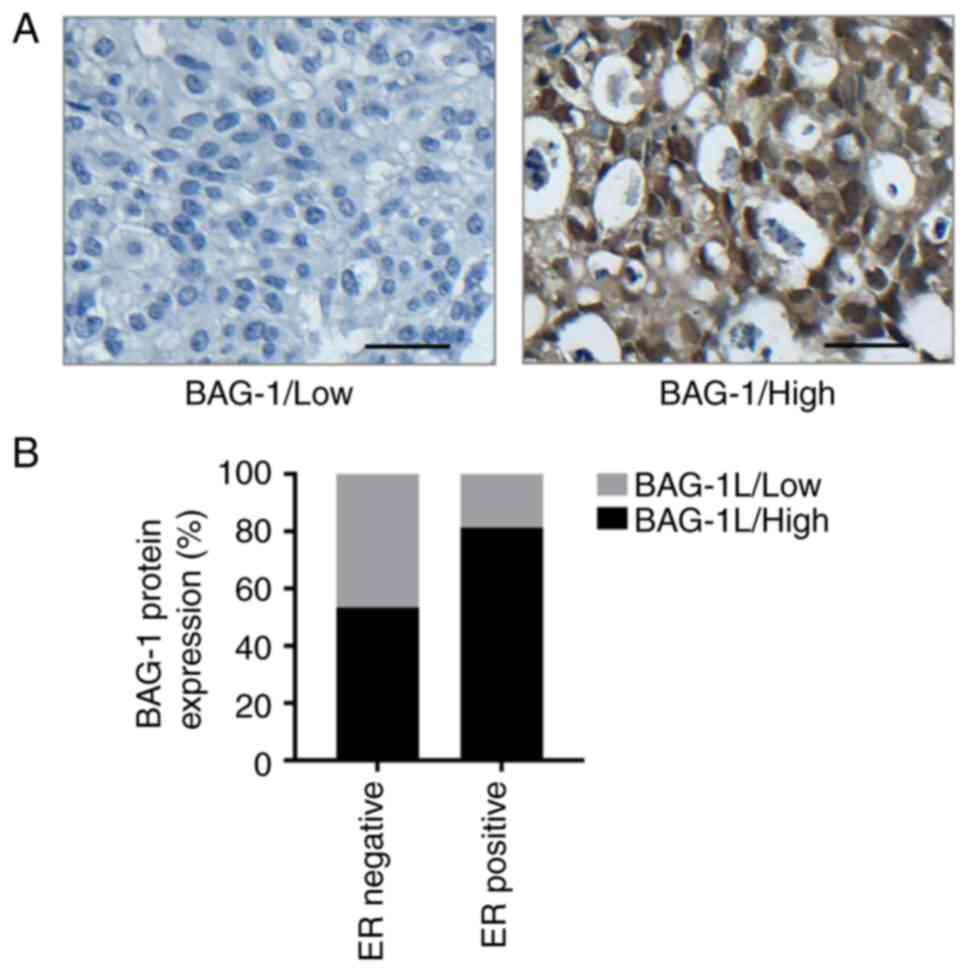
expression rate of BAG-1 protein was 95.8% (114/119). BAG-1 was
localized in either the nucleus or the cytoplasm, and the majority
of specimens exhibited positive nuclear and cytoplasmic staining
results (Fig. 1A). The association
of BAG-1 expression with the principal clinicopathological features
of patients with breast cancer included in the TMA are summarized
in Table I. Overexpression of BAG-1
was positively associated with ER positivity (P=0.001; Fig. 1B), progesterone receptor positivity
(P=0.003), tumor size (P=0.022) and tumor grade (P<0.001), but
negatively associated with human epidermal growth factor receptor-2
amplification status (P=0.004). To verify the association between
BAG-1 and ER expression, BAG-1L, which has been identified
previously (22) to interact with
numerous other nuclear hormone receptors was investigated. BAG-1L
expression levels were determined using western blotting and
RT-qPCR. The results indicated that BAG-1L was overexpressed in
T47D cells (Fig. 2A and B).
Expression of ER mRNA was increased almost 7-fold, and enhanced
expression of ER protein was also observed in
T47D/BAG-1L+ cells at 48 h post-transfection using
western blot analysis (Fig. 2C and
D). In addition, the expression levels of ER protein and mRNA
following knockdown of BAG-1 were decreased (Fig. 2C and D).
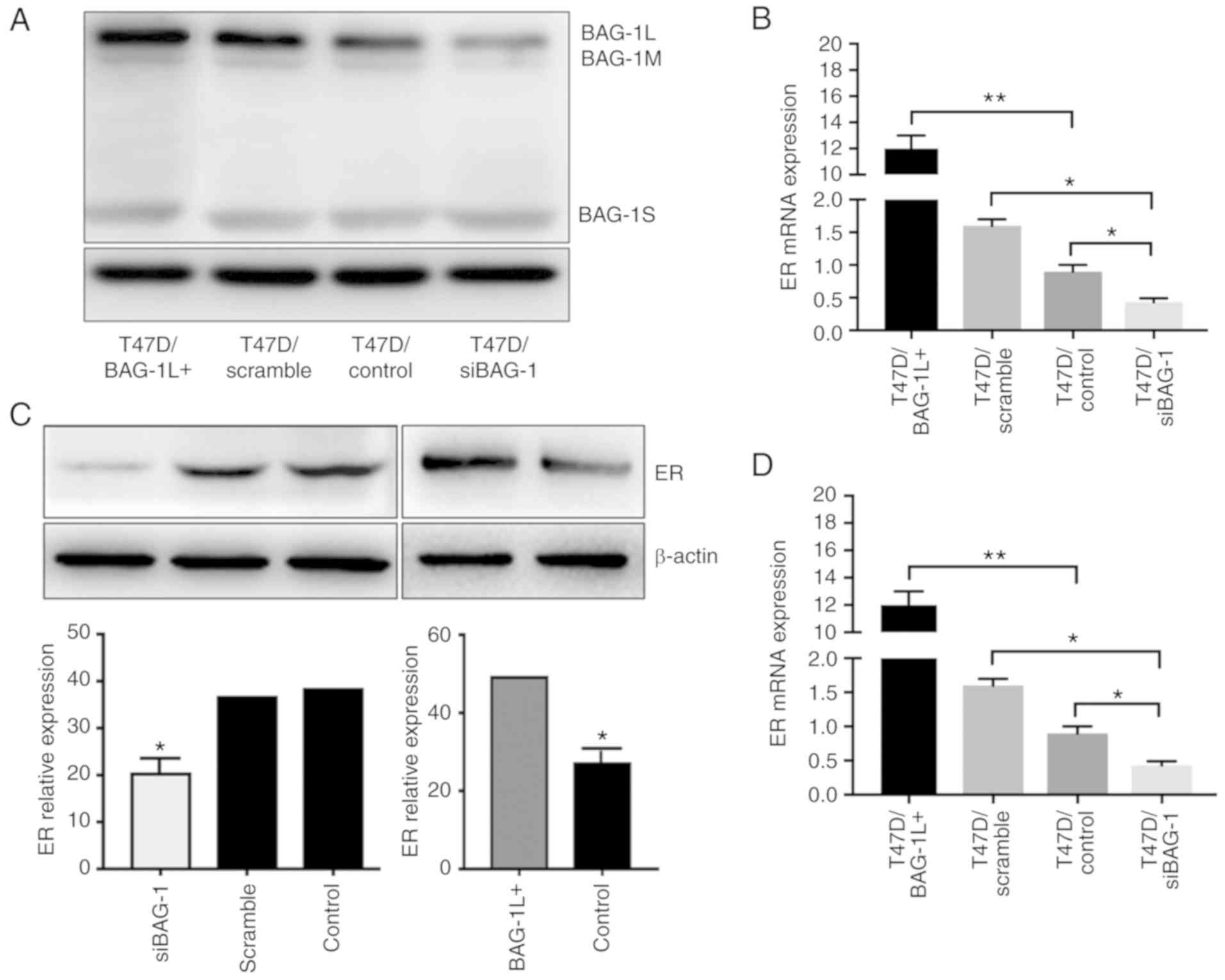 | Figure 2.BAG-1 expression in T47D cells is
positively associated with ER expression. (A) Representative
western blot analysis of T47D cells overexpressing BAG-1L
(T47D/BAG-1L+), and T47D cells transfected with vector
controls (T47D/control) or with scrambled siRNA (T47D/scramble) and
siRNA (T47D/siBAG-1). β-actin was used as a loading control. (B)
BAG-1 mRNA expression in T47D/BAG-1L+, T47D/control and
T47D/siBAG-1 and T47D/scramble siRNA cells. (C) Representative
western blot analyses of ER proteins in T47D/BAG-1L+,
T47D/control, T47D/siBAG-1 cells at 48 h post-transfection. β-actin
was used as a loading control. (D) mRNA expression of ER in
T47D/BAG-1L+, T47D/control, T47D/siBAG-1 and
T47D/scramble siRNA cells. BAG-1, Bcl-2-associated athanogene 1;
BAG-1L, BAG-1 long; si/siRNA, short interfering RNA; ER, estrogen
receptor; BAG-1M, BAG-1 medium; BAG-1S, BAG-1 short. *P<0.05,
**P<0.001 (one-way analysis of variance followed by Tukey's
multiple comparisons test). |
 | Table I.Association of BAG-1 expression with
clinicopathological features of patients with breast cancer
included in the tissue microarray. |
Table I.
Association of BAG-1 expression with
clinicopathological features of patients with breast cancer
included in the tissue microarray.
|
|
| BAG-1
expression |
|
|---|
|
|
|
|
|
|---|
| Characteristic | Total
no.b | Low | High | P-value
(χ2) |
|---|
| Age, years |
|
|
| 0.820 |
|
<50 | 54 | 15 | 39 |
|
|
≥50 | 64 | 19 | 45 |
|
| Tumor size, cm |
|
|
| 0.022 |
| ≤5 | 96 | 28 | 78 |
|
|
>5 | 12 | 7 | 5 |
|
| Tumor grade |
|
|
| <0.001 |
| I | 40 | 21 | 19 |
|
|
II+III | 79 | 14 | 65 |
|
| Nodal status |
|
|
| 0.458 |
|
Negative | 43 | 15 | 28 |
|
|
Positive | 64 | 18 | 46 |
|
| ERa |
|
|
| 0.001 |
|
Negative | 43 | 20 | 23 |
|
|
Positive | 75 | 14 | 61 |
|
| PRa |
|
|
| 0.003 |
|
Negative | 51 | 22 | 29 |
|
|
Positive | 66 | 12 | 54 |
|
| HER-2a |
|
|
| 0.002 |
|
Negative | 89 | 20 | 69 |
|
|
Positive | 30 | 15 | 15 |
|
Modulation of BAG-1 expression affects
the proliferation of T47D cells treated with tamoxifen
Tamoxifen has been identified to significantly
improve survival rates of patients with breast cancer. However,
patients treated with tamoxifen remain at risk of cancer recurrence
and mortality owing to drug resistance. To investigate the
potential association between BAG-1 and the response to tamoxifen
exhibited by ER-positive cells, cells were treated with 4-OH TAM at
a concentration of 10 µmol/l for 48 h, and cell cycle and apoptosis
analyses were performed to determine cell proliferation. BAG-1
silencing led to a decreased apoptosis rate (41.70±1.93 vs.
55.03±2.39%; P=0.012; Fig. 3A and
B) when compared with control cells following treatment with 10
µmol/l 4-OH TAM. In contrast, T47D cells overexpressing BAG-1L
exhibited significantly increased levels of 4-OH TAM-induced
apoptosis at a concentration of 10 µmol/l (65.10±2.35 vs.
55.03±2.39%; P=0.039; Fig. 3A and
B). Furthermore, a decreased proportion of T47D/siBAG-1 cells
was blocked in G1 phase compared with that of control
cells (51.27±2.67 vs. 62.15±2.46%; P=0.007; Fig. 3C and D) following treatment with 10
µmol/l 4-OH TAM. In addition, cells overexpressing BAG-1L exhibited
a slightly increased proportion of cells in G1 phase
compared with that of control cells (66.72±2.63 vs. 62.15±2.46%;
P=0.093; Fig. 3C and D). Taken
together, these results indicated that BAG-1L overexpression may
enhance the sensitivity of ER-positive cells to treatment with
tamoxifen, and silencing of BAG-1 may attenuate the
proliferation-inhibitory effect of tamoxifen in ER-positive
cells.
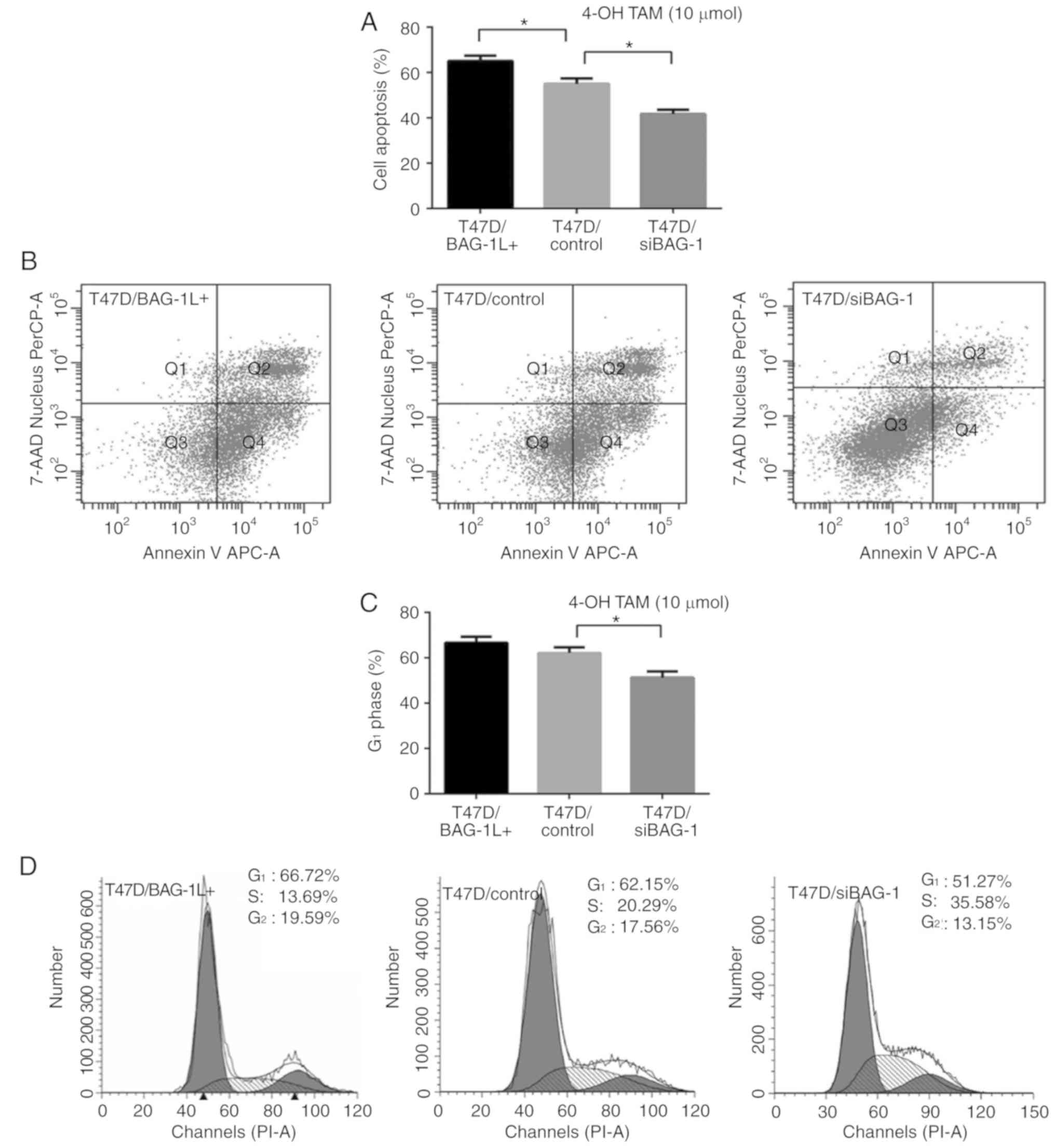 | Figure 3.Modulation of BAG-1 expression
affects the proliferation of ER-positive cells treated with
tamoxifen. (A) T47D/BAG-1L+, T47D/control and
T47D/siBAG-1 cells were cultured in 4-OH TAM (10 µmol/l) for 48 h.
Cell apoptosis was determined using flow cytometry. (B)
Representative flow cytometric plots of Annexin V assays using
three groups of cells treated with 4-OH TAM (10 µmol/l) for 48 h.
(C) Quantification of G1-phase cells in
T47D/BAG-1L+, T47D/control and T47D/siBAG-1 cells after
48 h of treatment with 4-OH TAM (10 µmol/l). (D) Representative
cell cycle histograms of T47D/BAG-1L+, T47D/control and
T47D/siBAG-1 cells after 48 h of treatment with 4-OH TAM (10
µmol/l). *P<0.05 (one-way ANOVA with Tukey's multiple
comparisons test; n=3). BAG-1, Bcl-2-associated athanogene 1;
BAG-1L, BAG-1 long; 4-OH TAM, 4-hydroxytamoxifen; si, short
interfering RNA; 7-AAD, 7-aminoactinomycin D; PerCP,
peridinin-chlorophyll protein complex; APC, allophycocyanin; PI,
propidium iodide. |
mTOR inhibitor restores tamoxifen
sensitivity in ER-positive breast cancer cells
The mTOR protein is a critical component in the
PI3K/Akt/mTOR signaling pathway, which is a key intracellular
signaling system involving in multiple cellular processes,
including proliferation and survival. Hyperactivation of this
pathway is associated with cell resistance to endocrine therapy
(23). The mTOR inhibitor rapamycin
was reported to be able to reverse endocrine therapy resistance
exhibited by breast cancer cells (24,25).
To demonstrate whether the inhibition of mTOR signaling was able to
restore cell sensitivity to treatment with tamoxifen,
T47D/BAG-1L+, T47D/control and T47D/siBAG-1 cells were
treated with either 4-OH TAM or rapamycin alone, or 4-OH TAM in
combination with rapamycin at a concentration based on a
predetermined half-maximal inhibitory concentration. The results
suggested that T47D/siBAG-1 cells were more susceptible to
treatment with rapamycin alone compared with the two other groups
of cells (T47D/siBAG-1 vs. T47D/BAG-1L+, 66.87±2.27 vs.
50.7±0.59%; P=0.003; T47D/siBAG-1 vs. T47D/control, 66.87±2.27 vs.
57.07±2.46%; P=0.037; T47D/BAG-1L+ vs. T47D/control,
50.7±0.59 vs. 57.07±2.46%; P=0.173; Fig. 4A). T47D/siBAG-1 cells exposed to
combination treatment exhibited a greater increase in
susceptibility to rapamycin treatment compared with the two other
groups of cells (T47D/siBAG-1 vs. T47D/control vs.
T47D/BAG-1L+, 71.23±2.23 vs. 58.8±1.89 vs. 66.93±1.27%;
P=0.008; Fig. 4A). Furthermore,
rapamycin markedly enhanced 4-OH TAM-induced apoptosis in
T47D/siBAG-1 cells (41.70±1.93 vs. 71.23±2.23%; P<0.001;
Fig. 4B, whereas levels of
apoptotic cell death were slightly increased in
T47D/BAG-1L+ cells (65.1±2.35 vs. 66.93±1.27%; P=0.531;
Fig. 4C and T47D/control cells
(55.03±2.39 vs. 58.8±1.89%; P=0.284; Fig. 4B). Representative results of cell
apoptosis 48 h after treatment are presented in Fig. 4C.
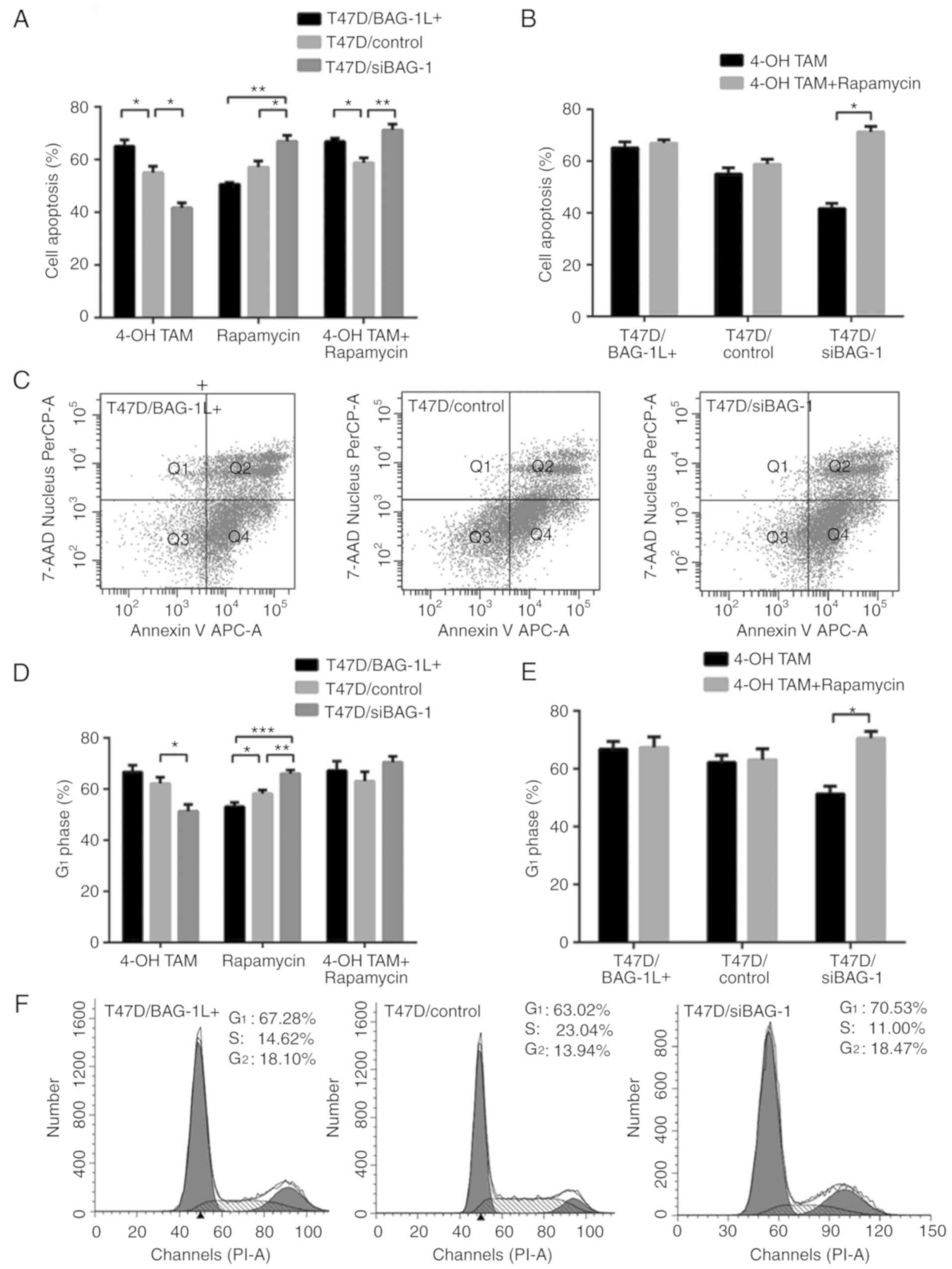 | Figure 4.Rapamycin restores cell sensitivity
to 4-OH TAM. (A) T47D/BAG-1L+, T47D/control and
T47D/siBAG-1 cells were cultured with either 4-OH TAM (10 µmol/l)
or rapamycin (10 µmol/l) alone, or with 4-OH TAM (10 µmol/l) in
combination with rapamycin (10 µmol/l), for 48 h. Cell apoptosis
was determined using flow cytometry. *P<0.05, **P<0.01
(one-way ANOVA with Tukey's multiple comparisons test; n=3). (B)
T47D/BAG-1L+, T47D/control and T47D/siBAG-1 cells were
cultured with either 4-OH TAM (10 µmol/l) alone, or with 4-OH TAM
(10 µmol/l) in combination with rapamycin (10 µmol/l), for 48 h.
Cell apoptosis was determined using flow cytometry. *P<0.05
(Student's t-test; n=3). (C) Representative flow cytometric plots
of annexin assays using T47D cells treated with a combination of
4-OH TAM (10 µmol/l) and rapamycin (10 µmol/l) for 48 h. (D)
Quantification of G1-phase cells in
T47D/BAG-1L+, T47D/control and T47D/siBAG-1 cells after
48 h of treatment with either 4-OH TAM (10 µmol/l) or rapamycin (10
µmol/l) alone, or with 4-OH TAM (10 µmol/l) in combination with
rapamycin (10 µmol/l). *P<0.05, **P<0.01, ***P<0.001
(one-way ANOVA with Tukey's multiple comparisons test; n=3). (E)
Quantification of G1-phase cells in
T47D/BAG-1L+, T47D/control and T47D/siBAG-1 cells after
48 h of treatment with 4-OH TAM (10 µmol/l) and rapamycin (10
µmol/l) in combination. *P<0.05 (Student's t-test; n=3). (F)
Representative cell cycle histograms of T47D/BAG-1L+,
T47D/control and T47D/siBAG-1 cells after 48 h of treatment with
4-OH TAM (10 µmol/l) and rapamycin (10 µmol/l) in combination. 4-OH
TAM, 4-hydroxytamoxifen; BAG-1, Bcl-2-associated athanogene 1;
BAG-1L, BAG-1 long; si, short interfering RNA; ANOVA, analysis of
variance; 7-AAD, 7-aminoactinomycin D; PerCP, peridinin-chlorophyll
protein complex; APC, allophycocyanin; PI, propidium iodide. |
Analysis of the cell cycle was performed to assess
the inhibition of proliferation mediated by treatment with 4-OH TAM
and rapamycin either separately or in combination. The proportion
of cells in G1 phase was increased following treatment
with a combination of 4-OH TAM and rapamycin compared with cells
treated with 4-OH TAM alone. T47D/siBAG-1 cells exhibited a more
marked increase in G1 arrest (70.53±2.30 vs.
51.27±2.67%; P<0.001; Fig. 4D),
whereas the proportion of cells in G1 phase in the
control group (63.02±3.80 vs. 62.15±2.46%; P=0.755; Fig. 4D) and the T47D/BAG-1L+
group (67.28±3.80 vs. 66.72±2.63%; P=0.840; Fig. 4D) was only slightly increased
compared with that of T47D/siBAG-1 group. The results indicated
that a greater proportion of T47D/siBAG-1 cells were arrested in
G1 phase compared with the other two groups 48 h after
combination treatment; however, the differences were not
statistically significant (T47D/siBAG-1 vs. T47D/control vs.
T47D/BAG-1L+, 70.533±2.30 vs. 63.02±3.80 vs.
66.72±2.63%; P=0.086; Fig. 4E).
Overall, these results indicated that rapamycin could restore
sensitivity to 4-OH TAM in ER-positive breast cancer cells,
particularly in cells exhibiting low levels of BAG-1 expression.
Representative results of the cell cycle distribution 48 h after
combination treatment of 4-OH TAM and rapamycin are presented in
Fig. 4F.
Additionally, the protein levels of total Akt,
p-Akt, total mTOR and p-mTOR in cells overexpressing BAG-1 and in
BAG-1-knockdown cells were investigated. The results of western
blot analysis revealed a noticeable increase in p-Akt and p-mTOR
levels in T47D cells following the silencing of BAG-1 expression,
whereas upregulation of BAG-1L was revealed to decrease levels of
p-Akt and p-mTOR (Fig. 5A and B).
This suggests that Akt/mTOR signaling was activated in
BAG-1-silenced cells.
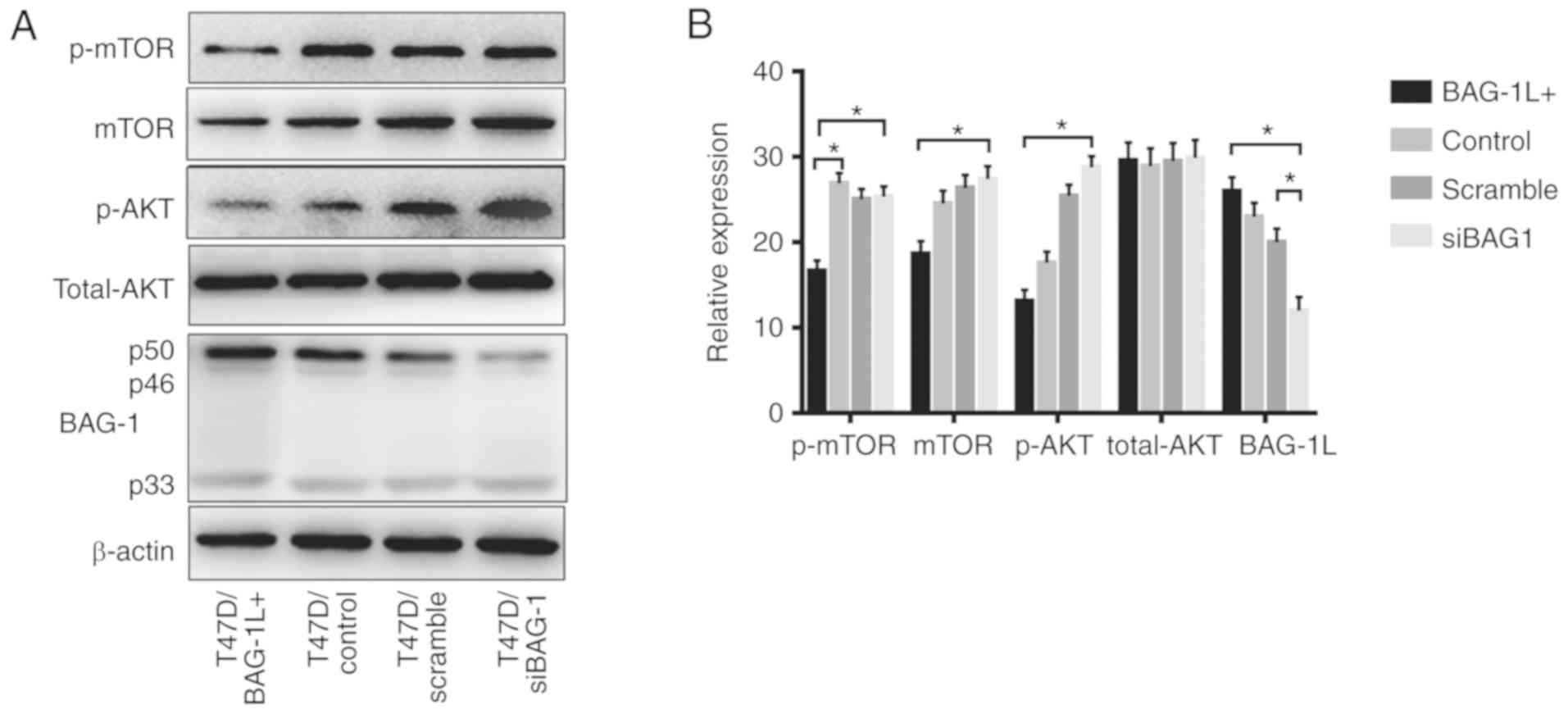 | Figure 5.Western blot analysis of protein
levels of p-mTOR, mTOR, p-Akt, Akt, BAG-1 and β-actin in
T47D/BAG-1L+, T47D/control, T47D/siBAG-1 and
T47D/scramble siBAG-1 cells. (A) Representative western blot
images. (B) Quantitative analysis. *P<0.05 one-way ANOVA with
Tukey's multiple comparisons test; n=3. p-, phospho-; mTOR,
mammalian target of rapamycin; Akt, protein kinase B; BAG-1,
Bcl2-associated athanogene 1; BAG-1L, BAG-1 long; si, short
interfering RNA. |
Discussion
ER is expressed in the majority of patients with
breast cancer. The functions of estrogen and anti-estrogen are
associated with ER expression. Previous studies have yielded
inconsistent results regarding the association between BAG-1
expression and ER status in breast cancer. The first study
analyzing this association was published in 1999 and exhibited
increased levels of positive BAG-1 nuclear or cytoplasmic staining
in the majority of ER-positive breast cancer tumors compared with
ER-negative tumors (26); however,
no statistical analysis was performed owing to the limited sample
size. A further cohort study of 122 female patients with stage I to
II breast cancer failed to identify an association between BAG-1
expression and ER positivity (27).
In a more homogeneous and larger cohort study including 138 breast
cancer cases, BAG-1 expression was identified to be moderately
correlated with ER (19). A further
two studies identified the positive correlation between BAG-1 and
ER expression (18,28). The association between BAG-1 protein
and ER expression has also been investigated in endometrial cancer,
which is another estrogen-dependent cancer. No significant
difference between the ER-negative and ER-positive groups was
identified in the proportion of samples exhibiting positive BAG-1
expression (29). Explanations for
these inconsistent results may include differences in the
antibodies used, differing immunohistochemistry techniques and
scoring methods, and a different proportion of patients with
ER-positive breast cancer in the clinical cohorts. In the present
study, total BAG-1 protein expression was investigated via
immunohistochemistry using 119 breast cancer cases. It was
identified that high BAG-1 expression was associated with
ER-positive breast cancer. In addition, in vitro experiments
were performed, which revealed that knockdown of BAG-1 subsequently
downregulated ER expression in ER-positive breast cancer cells. In
contrast, BAG-1 overexpression was demonstrated to upregulate ER
expression. However, subcellular localization of associated
isoforms was not investigated in the present study. Further studies
are required to determine the respective contribution of specific
isoforms to the association between BAG-1 and ER expression. In
conclusion, this association between BAG-1 and ER expression
suggested that BAG-1 may have a function in the response to
endocrine therapy in ER-positive breast cancer. The PI3K/Akt/mTOR
signaling pathway is able to promote cell proliferation in the
absence of estrogen via activation of estrogen-independent ER
transcription activity (23). Thus,
hyperactivation of this pathway may contribute to endocrine
resistance by promoting hormone-independent proliferation.
Previous clinical studies have revealed that
enhanced BAG-1 protein expression in breast cancer specimens is an
independent factor that may predict a favorable outcome in
ER-positive patients receiving endocrine therapy (18,19).
BAG-1 was included as a favorable predictive biomarker in the
Oncotype DX test. In accordance with these published results, it
was identified in the present study that increased BAG-1 expression
levels were associated with enhanced responses to tamoxifen in
breast cancer cells, whereas knockdown of BAG-1 expression led to
attenuated inhibition of cell proliferation. Furthermore, enhanced
levels of p-Akt and p-mTOR were observed in cells following
silencing of BAG-1 expression. On the basis of the results of the
present and previous studies (4,30)
suggesting that activation of the mTOR signaling pathway promotes
anti-estrogen resistance, we hypothesized that BAG-1 may represent
an inhibitor of the PI3K/Akt/mTOR signaling pathway and may be
associated with tamoxifen resistance. Knockdown of BAG-1 expression
activated this pathway, resulting in decreased sensitivity to
tamoxifen. Furthermore, an association between BAG-1 overexpression
and resistance to tamoxifen in MCF-7 cells was observed (Lu et
al, unpublished data). Such differing results may be due to the
following factors: Different cell lines with distinct biological
characteristics used for investigation, complex crosstalk in
cellular signaling pathways and different concentrations of
tamoxifen used to treat cells.
The PI3K/Akt/mTOR signaling pathway is frequently
activated in breast cancer (31,32).
Activation of this pathway has been associated with resistance to
endocrine therapy in patients with ER-positive breast cancer
(33–35). Pre-clinical studies have revealed
that the mTOR inhibitor rapamycin and its analogs may reverse
endocrine resistance in ER-positive breast cancer cells with
activated PI3K/Akt/mTOR signaling pathways (30,36).
Clinical trials have demonstrated that combined treatment of
everolimus and tamoxifen, or a combinatory treatment of everolimus
and exemestane, are more effective than the use of either endocrine
agent alone in patients who have become resistant to endocrine
therapy and their disease state has progressed. The BOLERO-2 trial
was a phase III randomized trial that compared the combinatory
treatment of everolimus (an mTOR inhibitor) and exemestane with
treatment using a placebo and exemestane in post-menopausal women
with HR-positive advanced breast cancer. Improved progression-free
survival was observed in patients treated with a combination of
everolimus and exemestane (10.6 vs. 4.1 months; P<0.0001)
(37). The TAMRAD trail evaluated
the efficacy of treatment with tamoxifen alone compared with
combinatory treatment with tamoxifen and everolimus in patients
with aromatase inhibitor-resistant metastatic breast cancer. This
study revealed that combinatory treatment with tamoxifen and
everolimus significantly improved the clinical benefit rate (61 vs.
42%; P=0.045) and progression rates (8.6 vs. 4.5 months; P=0.002)
(38). Considering the possible
involvement of BAG-1 in the mTOR pathway, the inhibitory effect on
cells was assessed following silencing of BAG-1 expression
post-treatment with tamoxifen alone as well as post-combinatory
treatment with tamoxifen and rapamycin. The results of the present
study indicated that rapamycin is able to restore the sensitivity
of ER-positive breast cancer cells to treatment with 4-OH TAM.
Furthermore, cells exhibiting decreased BAG-1 expression appeared
to have a greater sensitivity to rapamycin, and combinatory
treatment involving the targeting of ER with tamoxifen and the
targeting of mTOR with rapamycin significantly potentiated
inhibition of cell proliferation following the silencing of BAG-1
expression. These results may be partly due to hyperactivation of
the PI3K/Akt/mTOR signaling pathway induced by the downregulation
of the BAG-1 pathway inhibitor. A previous study has revealed that
overactivity of the PI3K/Akt/mTOR signaling pathway may result in
ER-positive cells exhibiting a greater sensitivity to rapamycin and
its analogs (27). In further work,
animal experiments are required, using animals with high expression
of target genes and gene knockout animals, or using in vivo
injection blockers to further verify the results of cell
experiments to explore the function of BAG-1 in tamoxifen
resistance and its potential underlying molecular mechanism.
In conclusion, the results of the present study
revealed that loss of BAG-1 expression may activate the
PI3K/Akt/mTOR signaling pathway and protect ER-positive breast
cancer cells from tamoxifen-induced inhibition of proliferation.
ER-positive breast cancer cells with low BAG-1 expression appeared
to exhibit a greater sensitivity to treatment with the mTOR
inhibitor rapamycin. Furthermore, the results of the present study
suggested that combinatory treatment targeting ER with tamoxifen
and targeting mTOR with rapamycin may significantly potentiate the
proliferation inhibitory effect in cells following the silencing of
BAG-1.
Acknowledgements
Not applicable.
Funding
This research did not receive any specific grant
from funding agencies in the public, commercial or not-for-profit
sectors.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
HL made substantial contributions to conception and
design. SL contributed to acquisition of data, and analysis and
interpretation of data. YD drafted and revised the manuscript. XF
performed the histological examination of the breast cancer. FC and
YM contributed to critical revision of the manuscript. All authors
approved the paper and HL gave the final approval of the version to
be published.
Ethics approval and consent to
participate
Human breast cancer tissue microarrays (TMAs) were
purchased from Shanghai Outdo Biotech Co., Ltd. (Shanghai, China;
cat. no. HBre-Duc159Sur-01). Informed consent was obtained from all
patients according to Shanghai Outdo Biotech Co., Ltd., and the
study protocol was approved by the Ethics Committee of Tianjin
Medical University Cancer Institute and Hospital. All procedures
performed in studies involving human participants were in
accordance with the ethical standards of the institutional and/or
national research committee, and with The Declaration of Helsinki
(1964) and its later amendments, or comparable ethical
standards.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Setiawan VW, Monroe KR, Wilkens LR,
Kolonel LN, Pike MC and Henderson BE: Breast cancer risk factors
defined by estrogen and progesterone receptor status: The
multiethnic cohort study. Am J Epidemiol. 169:1251–1259. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Li CI, Daling JR and Malone KE: Incidence
of invasive breast cancer by hormone receptor status from 1992 to
1998. J Clin Oncol. 21:28–34. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Systemic treatment of early breast cancer
by hormonal, cytotoxic, or immune therapy. 133 randomised trials
involving 31,000 recurrences and 24,000 deaths among 75,000 women.
Early Breast Cancer Trialists' Collaborative Group. Lancet.
339:71–85. 1992.PubMed/NCBI
|
|
4
|
Yue W, Fan P, Wang J, Li Y and Santen RJ:
Mechanisms of acquired resistance to endocrine therapy in
hormone-dependent breast cancer cells. J Steroid Biochem Mol Biol.
106:102–110. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Takayama S, Sato T, Krajewski S, Kochel K,
Irie S, Millan JA and Reed JC: Cloning and functional analysis of
BAG-1: A novel Bcl-2-binding protein with anti-cell death activity.
Cell. 80:279–284. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wang HG, Takayama S, Rapp UR and Reed JC:
Bcl-2 interacting protein, BAG-1, binds to and activates the kinase
Raf-1. Proc Natl Acad Sci USA. 93:7063–7068. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Takayama S, Bimston DN, Matsuzawa S,
Freeman BC, Aime-Sempe C, Xie Z, Morimoto RI and Reed JC: BAG-1
modulates the chaperone activity of Hsp70/Hsc70. EMBO J.
16:4887–4896. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Devireddy LR, Kumar KU, Pater MM and Pater
A: BAG-1, a novel Bcl-2-interacting protein, activates expression
of human JC virus. J Gen Virol. 81:351–357. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhou R, Gray NA, Yuan P, Li X, Chen J,
Chen G, Damschroder-Williams P, Du J, Zhang L and Manji HK: The
anti-apoptotic, glucocorticoid receptor cochaperone protein BAG-1
is a long-term target for the actions of mood stabilizers. J
Neurosci. 25:4493–4502. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hinitt CA, Wood J, Lee SS, Williams AC,
Howarth JL, Glover CP, Uney JB and Hague A: BAG-1 enhances
cell-cell adhesion, reduces proliferation and induces
chaperone-independent suppression of hepatocyte growth
factor-induced epidermal keratinocyte migration. Exp Cell Res.
316:2042–2060. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Clemo NK, Collard TJ, Southern SL, Edwards
KD, Moorghen M, Packham G, Hague A, Paraskeva C and Williams AC:
BAG-1 is up-regulated in colorectal tumour progression and promotes
colorectal tumour cell survival through increased NF-kappaB
activity. Carcinogenesis. 29:849–857. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wang YD, Ha MW, Cheng J, Zhang WL, Cong X,
Tong CY and Sun J: The role of expression and polymorphism of the
BAG-1 gene in response to platinum-based chemotherapeutics in
NSCLC. Oncol Rep. 27:979–986. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ni W, Chen B, Zhou G, Lu C, Xiao M, Guan
C, Zhang Y, He S, Shen A and Ni R: Overexpressed nuclear BAG-1 in
human hepatocellular carcinoma is associated with poor prognosis
and resistance to doxorubicin. J Cell Biochem. 114:2120–2130. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ozfiliz P, Kizilboga T, Demir S, Alkurt G,
Palavan-Unsal N, Arisan ED and Dinler-Doganay G: Bag-1 promotes
cell survival through c-Myc-mediated ODC upregulation that is not
preferred under apoptotic stimuli in MCF-7 cells. Cell Biochem
Funct. 33:293–307. 2015. View
Article : Google Scholar : PubMed/NCBI
|
|
15
|
Papadakis E, Robson N, Yeomans A, Bailey
S, Laversin S, Beers S, Sayan AE, Ashton-Key M, Schwaiger S,
Stuppner H, et al: A combination of trastuzumab and BAG-1
inhibition synergistically targets HER2 positive breast cancer
cells. Oncotarget. 7:18851–18864. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Packham G, Brimmell M and Cleveland JL:
Mammalian cells express two differently localized Bag-1 isoforms
generated by alternative translation initiation. Biochem J.
328:807–813. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Takayama S, Krajewski S, Krajewska M,
Kitada S, Zapata JM, Kochel K, Knee D, Scudiero D, Tudor G, Miller
GJ, et al: Expression and location of Hsp70/Hsc-binding
anti-apoptotic protein BAG-1 and its variants in normal tissues and
tumor cell lines. Cancer Res. 58:3116–3131. 1998.PubMed/NCBI
|
|
18
|
Millar EK, Anderson LR, McNeil CM, O'Toole
SA, Pinese M, Crea P, Morey AL, Biankin AV, Henshall SM, Musgrove
EA, et al: BAG-1 predicts patient outcome and tamoxifen
responsiveness in ER-positive invasive ductal carcinoma of the
breast. Br J Cancer. 100:123–133. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Cutress RI, Townsend PA, Sharp A, Maison
A, Wood L, Lee R, Brimmell M, Mullee MA, Johnson PW, Royle GT, et
al: The nuclear BAG-1 isoform, BAG-1L, enhances oestrogen-dependent
transcription. Oncogene. 22:4973–4982. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Paik S, Shak S, Tang G, Kim C, Baker J,
Cronin M, Baehner FL, Walker MG, Watson D, Park T, et al: A
multigene assay to predict recurrence of tamoxifen-treated,
node-negative breast cancer. N Engl J Med. 351:2817–2826. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2ΔΔCT method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Chen JM, Dando PM, Stevens RA, Fortunato M
and Barrett AJ: Cloning and expression of mouse legumain, a
lysosomal endopeptidase. Biochem J. 335:111–117. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ciruelos Gil EM: Targeting the
PI3K/AKT/mTOR pathway in estrogen receptor-positive breast cancer.
Cancer Treat Rev. 40:862–871. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ghayad SE, Bieche I, Vendrell JA, Keime C,
Lidereau R, Dumontet C and Cohen PA: mTOR inhibition reverses
acquired endocrine therapy resistance of breast cancer cells at the
cell proliferation and gene-expression levels. Cancer Sci.
99:1992–2003. 2008.PubMed/NCBI
|
|
25
|
Chang SB, Miron P, Miron A and Iglehart
JD: Rapamycin inhibits proliferation of estrogen-receptor-positive
breast cancer cells. J Surg Res. 138:37–44. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Brimmell M, Burns JS, Munson P, McDonald
L, O'Hare MJ, Lakhani SR and Packham G: High level expression of
differentially localized BAG-1 isoforms in some oestrogen
receptor-positive human breast cancers. Br J Cancer. 81:1042–1051.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Turner BC, Krajewski S, Krajewska M,
Takayama S, Gumbs AA, Carter D, Rebbeck TR, Haffty BG and Reed JC:
BAG-1: A novel biomarker predicting long-term survival in
early-stage breast cancer. J Clin Oncol. 19:992–1000. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Naderi A, Teschendorff AE, Barbosa-Morais
NL, Pinder SE, Green AR, Powe DG, Robertson JF, Aparicio S, Ellis
IO, Brenton JD, et al: A gene-expression signature to predict
survival in breast cancer across independent data sets. Oncogene.
26:1507–1516. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Moriyama T, Littell RD, Debernardo R,
Oliva E, Lynch MP, Rueda BR and Duska LR: BAG-1 expression in
normal and neoplastic endometrium. Gynecol Oncol. 94:289–295. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Beeram M, Tan QT, Tekmal RR, Russell D,
Middleton A and DeGraffenried LA: Akt-induced endocrine therapy
resistance is reversed by inhibition of mTOR signaling. Ann Oncol.
18:1323–1328. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
McCubrey JA, Davis NM, Abrams SL, Montalto
G, Cervello M, Libra M, Nicoletti F, D'Assoro AB, Cocco L, Martelli
AM and Steelman LS: Targeting breast cancer initiating cells:
Advances in breast cancer research and therapy. Adv Biol Regul.
56:81–107. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Gonzalez-Angulo AM, Ferrer-Lozano J,
Stemke-Hale K, Sahin A, Liu S, Barrera JA, Burgues O, Lluch AM,
Chen H, Hortobagyi GN, et al: PI3K pathway mutations and PTEN
levels in primary and metastatic breast cancer. Mol Cancer Ther.
10:1093–1101. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Pérez-Tenorio G and Stål O; Southeast
Sweden Breast Cancer Group, : Activation of AKT/PKB in breast
cancer predicts a worse outcome among endocrine treated patients.
Br J Cancer. 86:540–545. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Tokunaga E, Kimura Y, Oki E, Ueda N,
Futatsugi M, Mashino K, Yamamoto M, Ikebe M, Kakeji Y, Baba H and
Maehara Y: Akt is frequently activated in HER2/neu-positive breast
cancers and associated with poor prognosis among hormone-treated
patients. Int J Cancer. 118:284–289. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Campbell RA, Bhat-Nakshatri P, Patel NM,
Constantinidou D, Ali S and Nakshatri H: Phosphatidylinositol
3-kinase/AKT-mediated activation of estrogen receptor alpha: A new
model for anti-estrogen resistance. J Biol Chem. 276:9817–9824.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
deGraffenried LA, Friedrichs WE, Russell
DH, Donzis EJ, Middleton AK, Silva JM, Roth RA and Hidalgo M:
Inhibition of mTOR activity restores tamoxifen response in breast
cancer cells with aberrant Akt Activity. Clin Cancer Res.
10:8059–8067. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Baselga J, Campone M, Piccart M, Burris HA
III, Rugo HS, Sahmoud T, Noguchi S, Gnant M, Pritchard KI, Lebrun
F, et al: Everolimus in postmenopausal hormone-receptor-positive
advanced breast cancer. N Engl J Med. 366:520–529. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Bachelot T, Bourgier C, Cropet C,
Ray-Coquard I, Ferrero JM, Freyer G, Abadie-Lacourtoisie S, Eymard
JC, Debled M, Spaëth D, et al: Randomized phase II trial of
everolimus in combination with tamoxifen in patients with hormone
receptor-positive, human epidermal growth factor receptor
2-negative metastatic breast cancer with prior exposure to
aromatase inhibitors: A GINECO study. J Clin Oncol. 30:2718–2724.
2012. View Article : Google Scholar : PubMed/NCBI
|