Introduction
Pancreatic cancer (PC) has one of the highest
mortality rates among all tumor-associated diseases (1). Less than one-fifth of patients with PC
survive the first year, with a 5-year survival rate <6%
(1,2). The majority of patients with PC are
diagnosed at a late stage and succumb due to the invasion and
migration of cancer cells (3,4).
Current treatment methods, including surgical resection, radiation
and chemotherapy do not significantly increase patient long-term
survival (5–8). However, advancements in molecular
biological techniques have created an opportunity for the
exploration of effective targeted therapies for the treatment of
PC.
Tetraspanins (TSPANs), also known as transmembrane 4
superfamily (TM4SF) proteins, is composed of a group of
heterogeneous adaptor proteins, which exist in the form of
TSPAN-enriched microdomains (9,10). As
its name indicates, TM4SF consists of four transmembrane domains
that interact with various cell surface signaling molecules,
including integrins (11,12). It has been reported that the TSPAN
superfamily affects the malignant properties of cancer cells,
including their proliferation, apoptosis, metastasis, infiltration
and cell-cell aggregation (13,14).
TSPAN1 has been identified as a member of the TSPAN family
(15) and previous studies have
revealed that TSPAN1 is highly expressed in gastric, colon, liver
and esophageal cancers (13,16,17).
TSPAN1 has also been demonstrated to be important in gastric and
colon cancer cell invasion and metastasis (18,19).
However, the role of TSPAN1 in PC, specifically in PC cell
migration and invasion, is yet to be fully elucidated.
In the present study, various methods including
immunohistochemistry (IHC), reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) and western blotting were
utilized to determine and assess the expression of TSPAN1 in human
PC tissue samples, respective adjacent normal pancreatic tissue
samples and in human pancreatic ductal adenocarcinoma (PDAC) cell
lines. Furthermore, RT-qPCR and western blotting were performed to
assess the expression of TSPAN1 following transfection with TSPAN1
small interfering RNAs (siRNAs) and pLNCX-TSPAN1-cDNA recombinant
plasmids in human PC cell lines. Subsequently, cell migration and
invasion were assessed via Transwell assays. The expression of
matrix metalloproteinase (MMP2) and MMP9 were also determined and
the molecular mechanism of TSPAN1 in human PC cell migration and
invasion was further examined by employing phospholipase Cγ (PLCγ)
siRNA.
Materials and methods
Tissues, cell lines and cell
culture
The following PC cell lines SW1990, BxPC3, Capan1
and PANC-1, 293 and the immortalized non-tumorigenic human normal
pancreatic epithelial cell line (HPDE) were purchased from Shanghai
Institute of Biochemistry and Cell Biology, Chinese Academy of
Sciences (Shanghai, China). Cells were grown in Dulbecco's modified
Eagle's medium (DMEM; Invitrogen; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA) supplemented with 10% fetal calf serum (FCS;
Invitrogen; Thermo Fisher Scientific, Inc.) in a humidified
incubator at 37°C under 5% CO2. Cells were passaged
until they reached 70–80% confluence using 0.25% (w/v) trypsin
solution in 0.02% (w/v) EDTA. A total of 20 pairs of human PC
tissue samples and their respective adjacent normal pancreatic
tissue samples were obtained from China Medical University
(Liaoning, China). Official written informed consent was obtained
from each patient. A total of 95 patients were recruited between
June 2015 and June 2016 (52 males and 43 females; age, 39–81
years). PC diagnosis was confirmed in all patients included in the
present study. Patients had not undergone radiation therapy or
chemotherapy. Tissue samples were collected during routine surgery
performed in the Gastrointestinal Surgery Department of Cancer
Hospital of China Medical University and the Department of General
Surgery, Shenjing Hospital of China Medical University. The present
study was approved by the Ethical Committee of China Medical
University.
IHC
The expression of TSPAN1 in tissue samples was
detected via IHC staining. Tissue was fixed with 10% formalin
(Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) at room temperature
for 18 h, paraffin-embedded using a tissue processor (Model,
ATP700; Histo-Line laboratories, Milan, Italy) and sectioned on a
microtome (Model, Leica RM 2165, Germany) to 4 µm thick sections.
Samples were collected on silane-coated slides for
immunohistochemical staining. The tissue sections were sequentially
deparaffinized and rehydrated. Antigens were then retrieved at 95°C
using 10 mM citrate buffer (pH 6.0; Merck KGaA). Freshly prepared
3% H2O2 was used to quench endogenous
peroxidase activity. Blocking was subsequently performed with 10%
normal serum (Invitrogen; Thermo Fisher Scientific, Inc.) at room
temperature for 1 h. The sections were then sequentially incubated
with anti-TSPAN1 primary antibodies (1:200; cat. no. NBP2-33867;
Bio-Techne, Minneapolis, MN, USA) at 37°C for 2 h. Tissues were
then incubated with biotinylated goat anti-rabbit IgG secondary
antibodies (1:1,000; cat. no. BAF008; R&D Systems, Inc.,
Minneapolis, MN, USA) at room temperature for 30 min and washed
with PBS. This was followed by incubation with horseradish
peroxidase (HRP)-conjugated streptavidin (1:2,000; cat. no. N100;
Thermo Fisher Scientific, Inc.) for 5 min at room temperature.
Immunostaining was developed using the 3,3-Diaminobenzidine
substrate (Gene Tech Co., Ltd., Hong Kong, China) for 10 min at
room temperature. Sections were then counterstained with
hematoxylin (Invitrogen; Thermo Fisher Scientific, Inc.) at room
temperature for 3 min. Sections that were incubated with PBS only
served as the negative control. Cells were examined under an
inverted-TS100 microscope (magnification, ×200; Nikon Corporation,
Tokyo, Japan) and the Aperio Scanscope XT system (Aperio, San
Diego, CA, USA) was used to analyze the stained tissues
sections.
RT-qPCR
An RNeasy Mini kit (Qiagen GmbH, Hilden, Germany)
was used to extract RNA from cells (HPDE, SW1990, BxPC3, Capan1,
PANC-1, 293 cells, the transfected SW1990 and the transfected BxPC3
cells) following the manufacturer's protocol. Reverse transcription
(RT) was performed as follows: A mixture of 2 µg total RNA and 1 µl
Oligo (dT) was heated to 70°C for 5 min, rapidly cooled on ice and
mixed with a reaction mixture containing 5 µl M-MLV RT 5X Buffer
(cat. no. M531A; Promega Corporation, Madison, WI, USA), 0.5 µl
deoxyribonucleotide triphosphate (25 µM; cat. no. U1240; Promega
Corporation), 0.7 µl RNAse inhibitor (2500 U; RNasin®;
cat. no. N211A; Promega Corporation) and 1 µl M-MLV reverse
transcriptase (10,000 U; cat. no. M170A; Promega Corporation). The
reaction volume was made up to 25 µl with RNA-free water, and then
incubated at 42°C for 1 h and then 70°C for 10 min. Real-time PCR
was performed using a total reaction volume of 20 µl, which
included 2X Mix SYBR-Green I (10 µl; Promega Corporation), primer
(0.25 µl, 10 pmol/l), template DNA (1 µl) and sterile water.
RT-qPCR was performed using the Applied Biosystems 7900HT thermal
cycler instrument (Applied Biosystems; Thermo Fisher Scientific,
Inc.). The following thermocycling conditions were used: Initial
denaturation at 95°C for 2 min, followed by 50 cycles of 95°C for
15 sec and then 60°C for 45 sec. Target gene expression were
assessed by calculating fold change using the formula: Fold change
= 2 - [Cq (control) gene X-Cq (control) actin] [Cq (activated) gene
X-Cq (activated) actin] (20). The
following primer sequences were utilized: Human TSPAN1 forward,
5′-CGTTGTGGTCTTTGCTCTTG-3′ and reverse, 5′-TTCTTGATGGCAGGCACTAC-3′;
Human MMP2 forward, 5′-CTCGGTAGGGACATGCTAAGTAGAG-3′ and reverse,
5′-CCTCTGGAGGTTCGACGTGA-3′; Human MMP9 forward
5′-TCATGAGGAAGAGCTCTGAGT-3′ and reverse
5′-TCATGAGGAAGAGCTCTGAGT-3′; and Human GAPDH forward,
5′-GAAGGTGAAGGTCGGAGTC-3′ and reverse, 5′-GAAGATGGTGATGGGATTTC-3′.
The relative mRNA expression of TSPAN1, PLCγ and MMPs were
normalized to the respective endogenous GAPDH mRNA expression.
Western blot analysis
Radioimmunoprecipitation assay buffer and a protease
inhibitor were used to lyse cells and collect protein
(Sigma-Aldrich; Merck KGaA) (21).
A BCA assay (Thermo Fisher Scientific, Inc.) was used to quantify
protein concentration. A total of 50 µg of protein from each sample
was denatured, resolved using 10% SDS-PAGE gradient gel (EMD
Millipore, Billerica, MA, USA) and electro-blotted to a
polyvinylidene difluoride (PVDF) membrane (EMD Millipore).
Membranes were then incubated with 5% non-fat milk for 1 h at room
temperature and stained with primary antibodies against TSPAN1
(1:300; cat. no. NBP2-33867; Bio-Techne) MMP2 (1:500; cat. no.
4022; Cell Signaling Technology, Inc., Danvers, MA, USA), PLCγ1
(1:500; cat. no. 14008; Cell Signaling Technology, Inc.),
phosphorylated PLCγ1 (1:500; cat. no. 5690; Cell Signaling
Technology, Inc.) and anti-β-actin (1:1,000; cat. no. NB600-503;
Novus Biologicals, LLC, Littleton, CO, USA) at 4°C overnight. The
PVDF membrane was washed with TBST three times and incubated with
HRP-conjugated secondary antibodies (HRP-linked anti-rabbit IgG
antibodies; 1:3,000; cat. no. NB710-57836; Novus Biologicals, LLC)
for 90 min at room temperature. Enhanced chemiluminescence (Pierce;
Thermo Fisher Scientific, Inc.) was used to develop protein
signals. A scanning densitometer along with Quantity One Software
(version 4.4.1; Bio-Rad Laboratories, Inc., Hercules, CA, USA) was
used to quantify the optical density of each protein band. Protein
band density was normalized to the corresponding β-actin
density.
Cytosolic protein was isolated from
particulate-conjugated protein using the digitonin separation
method from the transfected SW1990 and BxPC3 cells (22). Cells were collected and re-suspended
in 1 ml saline solution (1 mM EDTA, 150 mM NaCl, 1 mM PMSF, 2 mM
EGTA, 1 µg/ml aprotinin, 10 µg/ml leupeptin, and 100 µg/ml
digitonin) with occasional shaking for 10 min. Cells were then
centrifuged at 13,000 × g for 5 min at 4°C and the resulting
supernatant contained the cytosolic proteins. The cell pellet was
dissolved in 1 ml lysis buffer (pH 7.4, 1 mM EDTA, 10 mM PBS, 1%
Triton X-100, 2 mM EGTA, 1 mM PMSF, 0.1% SDS, 1 µg/ml aprotinin and
10 µg/ml leupeptin) and the membrane protein was obtained
(particulate-conjugated proteins). A total of 80 µg of protein was
separated via SDS-PAGE and transferred to a PVDF membrane as
aforementioned. The expression of PLCγ1 and phosphorylated PLCγ1
were detected using western blotting as aforementioned.
Knockdown of TSPAN1 in SW1990 and
BxPC3 cells by RNAi
The downregulation of TSPAN1 in SW1990 and BxPC3
cells was achieved using specific siRNAs targeting TSPAN1 (TSPAN1
siRNA; cat. no. 4392420; Thermo Fisher Scientific, Inc.). Cells
transfected with scramble siRNA (cat. no. 4390846; Thermo Fisher
Scientific, Inc.) were used as the control. The TSPAN1 siRNA group
sequences were: 5′-GGCUCACGACCAAAAAGUAtt-3′ (sense) and
5′-UACUUUUUGGUCGUGAGCCtt-3′ (antisense). The scramble siRNA group
sequences were: 5′-UUCUCCGAACGUGUCACGUdTdT-3′ (sense),
5′-ACGUGACACGUUCGGAGAAdTdT-3′ (antisense). The transfection of
siRNAs into SW1990 and BxPC3 cells was performed following the
manufacturer's protocol. SW1990 and BxPC3 cells were re-suspended
at a density of 1×105 cells/ml in DMEM medium. The RNAi
MAX Lipofectamine transfection agent (Ambion; Thermo Fisher
Scientific, Inc.), siRNA (20 nM) and DMEM medium were mixed to
prepare transfection complexes. SW1990 and BxPC3 cells were then
mixed with the transfection complexes, seeded in 6-well
(2×105 cells/well) plates (Nalge Nunc International,
Rochester, NY, USA) and incubated for 24 h at 37°C. RNA analysis
was conducted on cells 48 h post-transfection, while protein
analysis was performed 72 h post-transfection. The silencing
efficiency of TSPAN1 siRNA was confirmed using RT-qPCR and western
blotting. The transfection method of PLCγ siRNA (cat. no. 1299001;
Thermo Fisher Scientific, Inc.) into 293, SW1990 and BxPC3 cells,
along with the determination of silencing efficacy were the same as
aforementioned.
Overexpression of TSPAN1 in SW1990 and
BxPC3 cells
A TSPAN1 cDNA clone plasmid was purchased from
Beijing Zeping Bioscience & Technology Co., Ltd. (Beijing,
China; cat. no. HG 13073-UT). Recombinant plasmids
(pLNCX-TSPAN1-cDNA) were constructed and sequenced by Bio Sune
Biotechnologies (Shanghai, China). A total of 10 µg of
pLNCX-TSPAN1-cDNA recombinant plasmid was transfected into 293,
SW1990 and BxPC3 cells via the calcium phosphate precipitation
method (23). SW1990, 293 and BxPC3
cells were grown on a 35 mm cell culture dish until they reached
70–80% confluence. The DNA-calcium phosphate precipitation
transfection complexes were prepared by mixing plasmid DNA and 2 M
CaCl2 (Sigma-Aldrich; Merck KGaA) in HEPES buffered
saline (included 50 mM HEPES, 280 mM NaCl, 1.5 mM
Na2HPO4; pH 7.1; Sigma-Aldrich; Merck KGaA).
The transfection complexes were subsequently added into the cells
drop by drop and incubated for 8 h at 37°C. Fresh DMEM medium was
then added for cell culture at 37°C. The pLNCX plasmid was
transfected into cells to serve as the control. At 48 h
post-transfection, 0.25% (w/v) trypsin solution in 0.02% (w/v) EDTA
was used to harvest cells, which were then cultured in DMEM medium
with G418 (500 mg/l; Thermo Fisher Scientific, Inc.) to screen
positive transfection clones. Following 2–3 weeks, positive clones
were cultured in DMEM supplemented with 20% (v/v) heat-inactivated
calf serum (Invitrogen; Thermo Fisher Scientific, Inc.) and G418
(200 µg/ml) at 37°C in 5% CO2.
Assessment of cell migration and cell
invasion in vitro by Transwell assay
Cell migration was measured using the BD Falcon HTS
multi-well insert system (pore size, 8.0 µm, BD Biosciences, San
Jose, CA, USA) (24). DMEM medium
supplemented with 10% FCS at a volume of 500 µl was added to the
lower chamber of the wells. Transfected SW1990 and BxPC3 cells
suspended in serum-free DMEM medium were added into the insert at a
density of 1.5×105 cells/ml. Non-migrated cells (the
cells remaining in the insert) were removed following incubation
for 6 h at 37°C by gently scraping with cotton swabs. Cells at the
bottom of the insert, which represent the migrated cells, were
fixed with 4% formaldehyde for 20 min at room temperature. Fixed
cells were then stained with 0.1% crystal violet for 30 min at room
temperature. Excess stain was removed by washing the inserts with
Milli-Q water (Merck KGaA). Filters were then dried overnight. The
number of migrated cells was estimated by counting cells in five
random fields and images were obtained using light microscopy and
captured using a camera (magnification, ×100; Nikon Digital Camera
Dxm 1200F; Nikon Corporation).
The cell invasion assay was performed using BioCoat
Matrigel invasion chambers (BD Biosciences) (25). A total of 50 µg of reduced serum
Matrigel (BD Biosciences) was added to the upper chamber of the
inserts to coat the Transwell membrane. Transfected SW1990 and
BxPC3 cellsin serum-free DMEM (1×106) were added into
the upper chambers. DMEM supplemented with 10% FCS was added to the
lower chamber and served as a chemoattractant. Following 48 h of
incubation at 37°C, the membrane facing the lower chamber (where
invaded cells were present) was gently removed and mounted on a
glass slide. The subsequent steps for fixation, staining and
measuring the level of cell invasion were as aforementioned for the
migration assay.
Statistical analysis
The results were expressed as the mean ± standard
deviation of at least three independent experiments performed in
triplicate. SPSS 17.0 software (SPSS, Inc., Chicago, IL, USA) was
used to determine significant differences. For comparisons between
two groups, a two-tailed Student's t-test was performed. One-way
analysis of variance followed by a Tukey's post hoc test was used
to assess significant differences among multiple groups. P<0.05
was considered to indicate a statistically significant
difference.
Results
Expression of TSPAN1 in clinical PC
tissue samples and cell lines
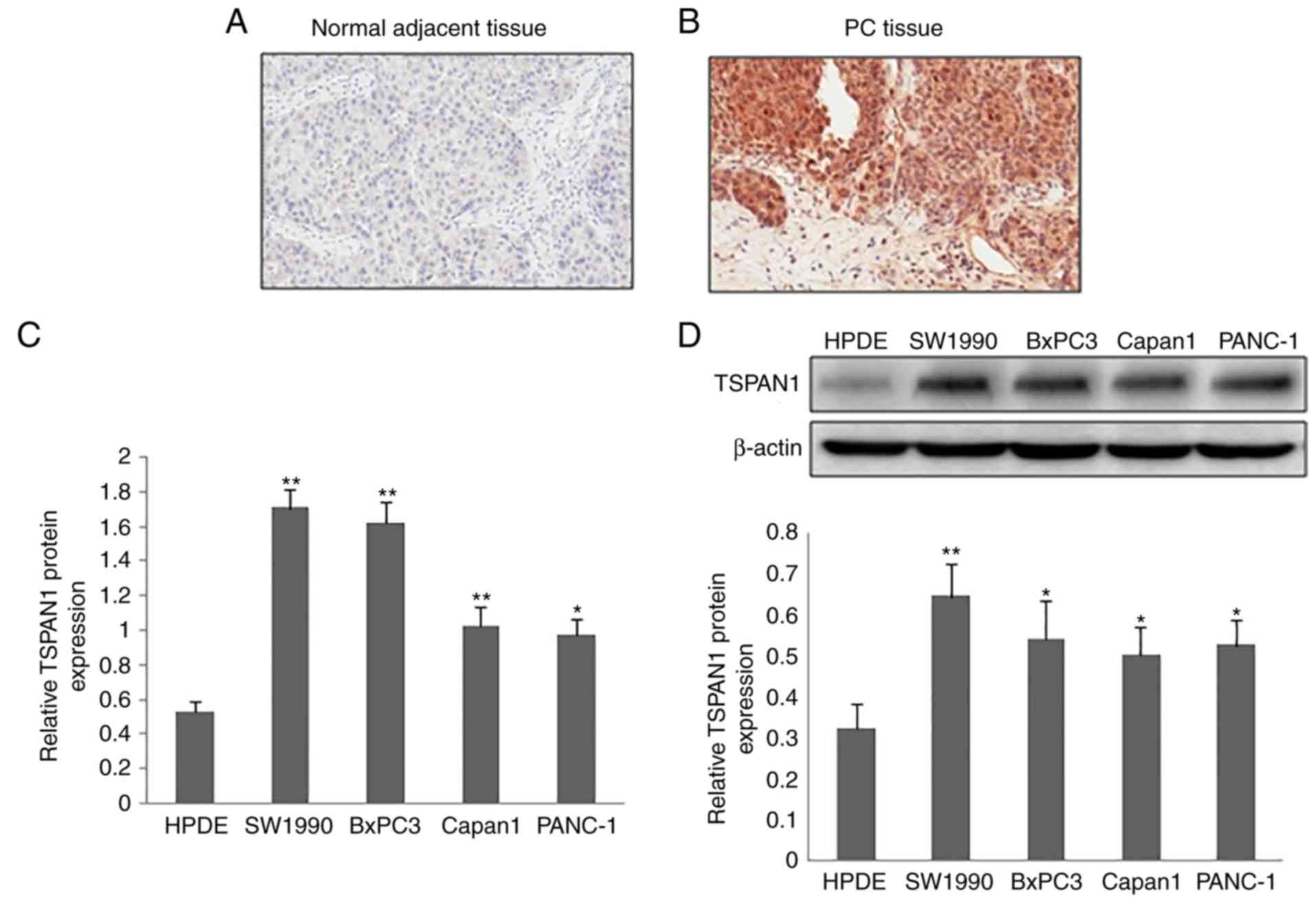
The protein expression of TSPAN1 in clinical tissue
samples of PC was detected via IHC staining. Compared with normal
adjacent tissue samples, a markedly increased TSPAN1expression was
observed in the PC tissue samples (Fig.
1A and B). TSPAN1 expression was predominantly observed in the
cytoplasm of the cells. The mRNA expression of TSPAN1, as
determined by RT-qPCR, was significantly increased in PC cell lines
(SW1990, BxPC3, Capan1 and PANC-1) compared with the normal
pancreatic cell line (HPDE; Fig.
1C). Similarly, TSPAN1 protein expression in the PC cell lines
(SW1990, BxPC3, Capan1 and PANC-1) was higher when compared with
the normal pancreatic cell line (HPDE; Fig. 1D).
Knockdown of TSPAN1 expression via
TSPAN1 siRNA transfection
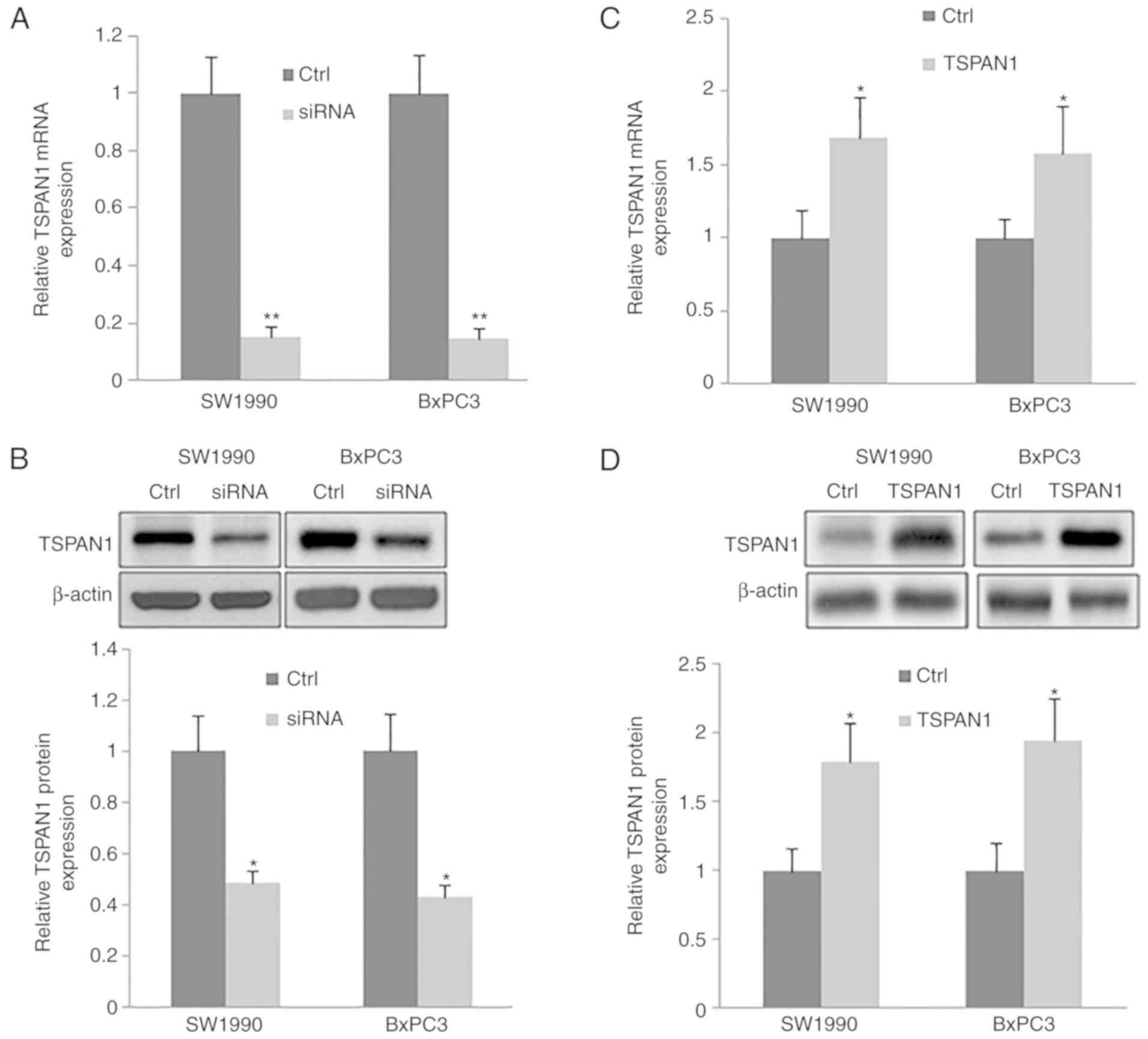
TSPAN1-siRNAs were transfected into human PC cell
lines (SW1990 and BxPC3). The silencing efficiency of TSPAN1-siRNAs
at the gene and protein level was confirmed by RT-qPCR and western
blotting, respectively. TSPAN1 mRNA was significantly decreased in
TSPAN1-siRNA transfected cells when compared with the control cells
(those transfected with scrambled siRNA; Fig. 2A). Consistently, a significant
downregulation of TSPAN1 protein was
observedinTSPAN1-siRNA-transfected cells compared with the control
cells (Fig. 2B).
Overexpression of TSPAN1 in SW1990 and
BxPC3 cells
The expression of TSPAN1 in PC cell lines (SW1990
and BxPC3) transfected with pLNCX-TSPAN1-cDNA recombinant plasmid
and pLNCX plasmid was determined via RT-qPCR and western blotting
(Fig. 2C and D). Cells transfected
with pLNCX were used as the control. The mRNA and protein
expression of TSPAN1 was significantly upregulated in cells
transfected with pLNCX-TSPAN1-cDNA compared with pLNCX-transfected
cells. The results indicate that the TSPAN1-cDNA recombinant
plasmid was successfully transfected into SW1990 and BxPC3
cells.
TSPAN1 inhibits SW1990 and BxPC3 cell
migration and invasion, and MMP2/9 expression
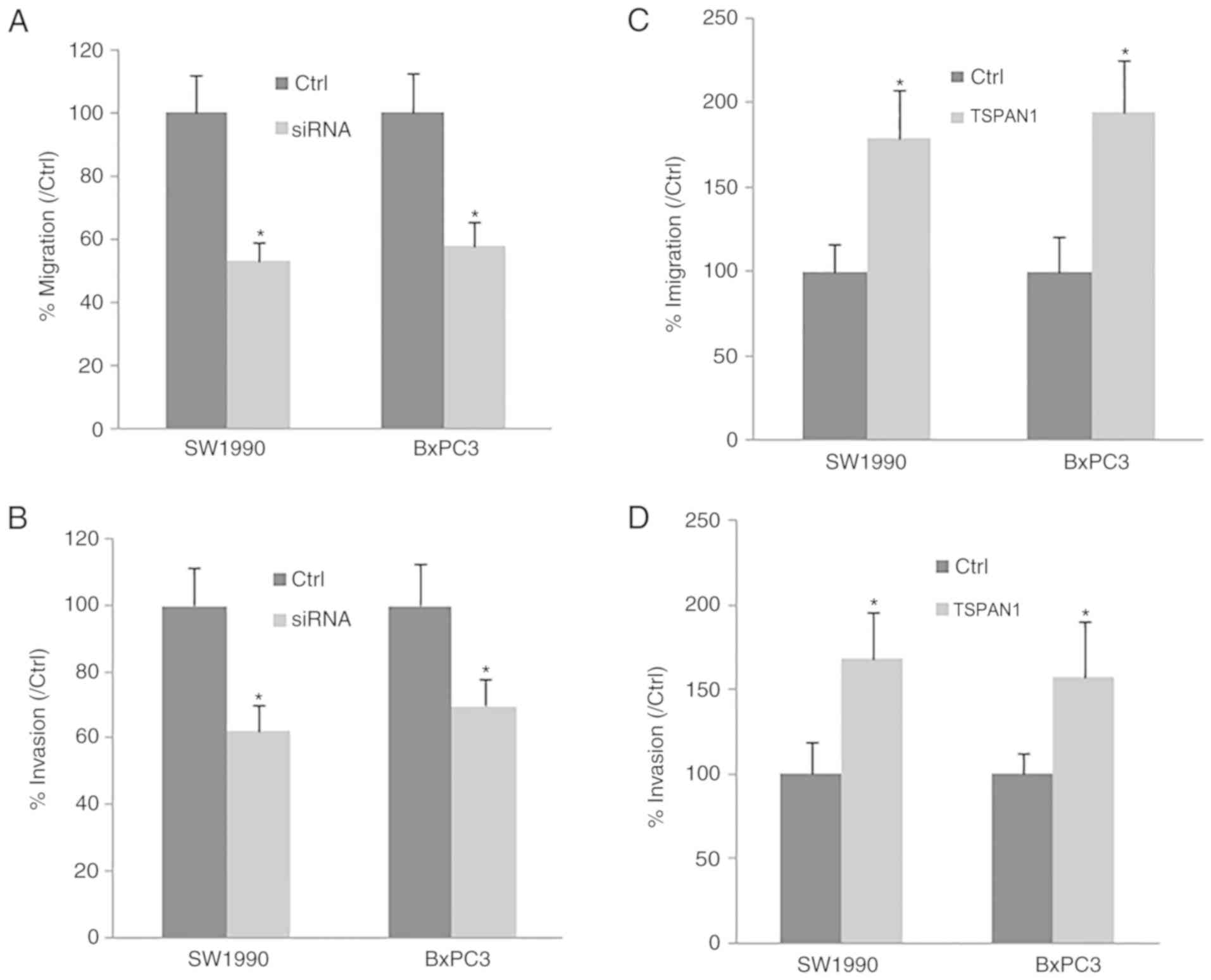
Cell migration and invasion were determined in PC
cell lines (SW1990 and BxPC3). Following TSPAN1-siRNA transfection,
the downregulation of TSPAN1 significantly decreased cell migration
and invasion compared with the control cells transfected with
scrambled siRNA (Fig. 3A and B).
Consistent with this, TSPAN1 ectopic overexpression in SW1990 and
BxPC3 cells markedly increased cell migration and invasion in these
cell lines (Fig. 3C and D). The
results indicate that TSPAN1 knockdown suppresses PC cell migration
and invasion.
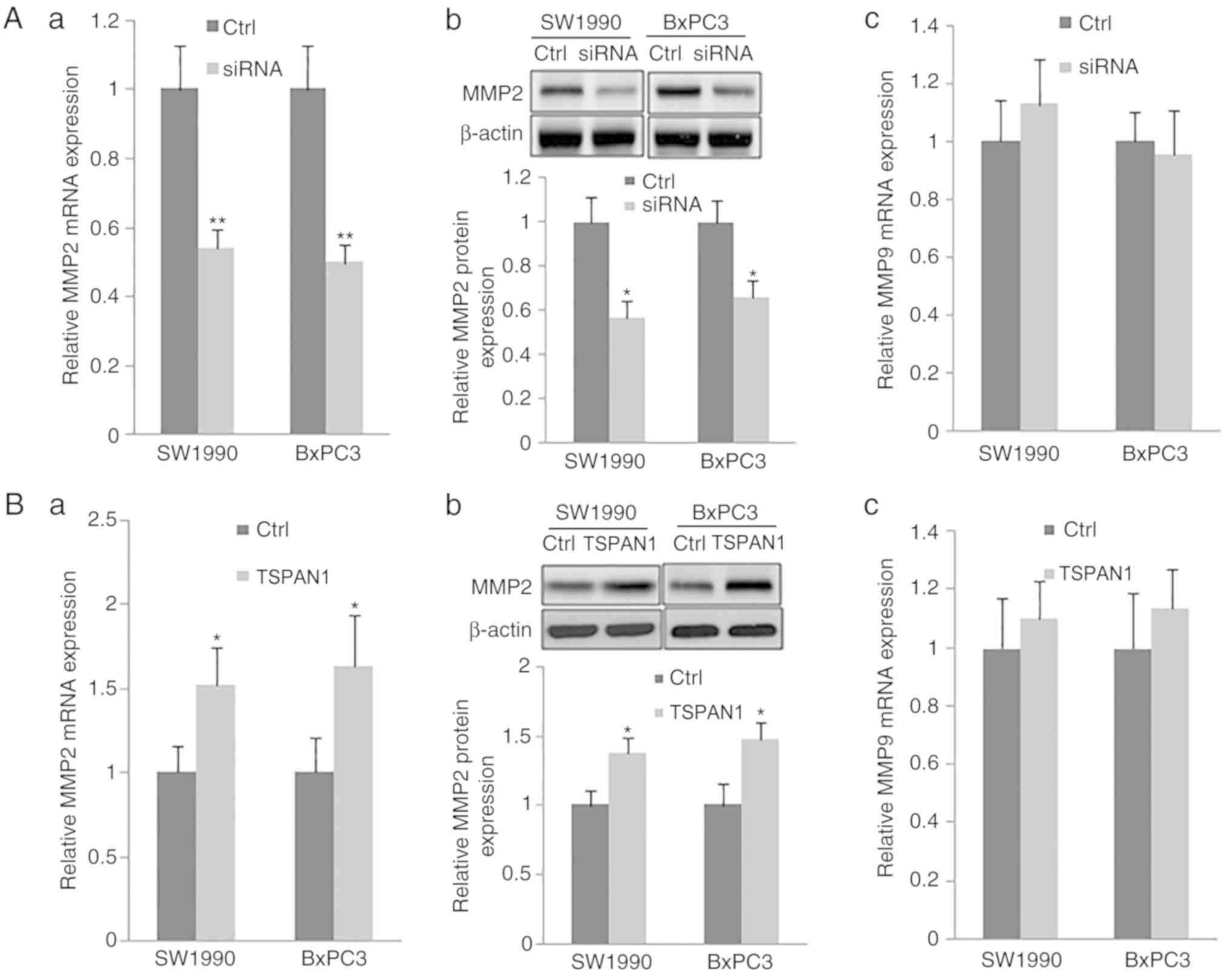
Additionally, the expression of MMP2 at the mRNA and
protein levels, as assessed via RT-qPCR and western blotting,
respectively, were significantly decreased in TSPAN1
siRNA-transfected SW1990 and BxPC3 cells in comparison with
scrambled siRNA transfected cells (Control). TSPAN1 ectopic
overexpression in the same cell lines increased the expression of
MMP2 at the mRNA and protein level (Fig. 4Ba and b). However, the mRNA levels
of MMP9 were not changed following transfection with TSPAN1 siRNA
or pLNCX-TSPAN1-cDNA in SW1990 and BxPC3 cells (Fig. 4Ac and Bc). These results indicate
that TSPAN1 knockdown suppresses the expression of MMP2, but not
that of MMP9.
Overexpression of TSPAN1 in PC cell
lines promotes cell migration, invasion and MMP2 expression via
PLCγ
The present study assessed the effects of PLCγ siRNA
on PC cell migration and invasion and MMP2 expression following
LNCX-TSPAN1-cDNA transfection. The expression of PLCγ was
significantly downregulated after PLCγ siRNA was transfected into
293 cells, as determined by RT-qPCR and western blot analysis
(Fig. 4C). Furthermore, the
upregulated MMP2 expression, induced by the overexpression of
TSPAN1 in the SW1990 and BxPC3 PC cell lines, was significantly
reduced following the transfection of PLCγ siRNA (Fig. 4D and E). Thus, the results indicate
that the overexpression of TSPAN1 promotes the expression of MMP2
and may be dependent on PLCγ. In addition, the increased cell
migration and invasion induced by TSPAN1 overexpression was
significantly attenuated following transfection with PLCγ siRNA
(Fig. 5A and B). Therefore, the
increased cell migration and invasion of TSPAN1-overexpressing
cells and the upregulation of MMP2 expression may be dependent on
PLCγ.
To confirm this hypothesis, the effect of TSPAN1
siRNA transfection on the activation of PLCγ was assessed. The
proteins in the cytoplasm were isolated from particulate-conjugated
proteins. The content of PLCγ1 and phosphorylated PLCγ1 were
detected via western blotting. The results presented in Fig. 5C and D revealed that TSPAN1 siRNA
transfection decreases the content of PLCγ1 in the cytoplasm and
decreases the content of PLCγ1 and phosphorylated PLCγ1 on the
plasma membrane.
The results demonstrated that TSPAN1 siRNA
transfection inhibits the translocation and phosphorylation of
PLCγ1, and thus inhibits the activation of PLCγ1. Thus, the
aforementioned results indicate that TSPAN1 may affect PC cell
migration and invasion as well as MMP2 expression via the
activation of PLCγ.
Discussion
PC is the most common malignancy of the pancreas
(1–3). PC arising from the ductal epithelium
of the pancreas, otherwise known as PDAC, is the most frequently
diagnosed type of PC, accounting for ~80–90% of all cases (1–3). PC
exhibits high rates of metastasis and mortality. As such, the
5-year survival rate following PC diagnosis is poor, often at
<6% (1,2). Therefore, effective therapeutic
strategies are urgently required for PC.
TSPANs are a membrane protein superfamily that has
four transmembrane domains (9).
This four-transmembrane structure constructs
intracellular-membrane-extracellular signaling pathways and
facilitates the binding and interactions of cell surface signaling
molecules, including integrin and cadherin (10–12).
It has been demonstrated that the TSPAN superfamily is implicated
in essential cellular activities, including cell survival, growth,
proliferation, differentiation, migration, signal transduction and
motility (9–14). TSPAN1 is a novel member of the
TSPANs family (15). Previous
studies have reported an overexpression of TSPAN1 in various types
of cancers, including liver, colon and gastric cancer (16–19).
In the present study, the results indicated that TSPAN1 was highly
expressed in human PC tissue samples compared with adjacent normal
pancreatic tissue samples. This observation was consistent in cell
lines, where the expression of TSPAN1 was significantly higher in
human PC cells (SW1990, BxPC3, Capan1 and PANC-1) when compared
with that observed in HPDE, a normal pancreatic cell line.
To date, the function of TSPAN1 in PC is still
undetermined. Previous reports have revealed that TSPAN1is
positively correlated with clinical stage and negatively correlated
with the survival rate of patients with cancer (25,26).
In colon cancer and skin carcinoma cells, TSPAN1 has been revealed
to inhibit cell survival, proliferation, migration and invasion
(19,27). Furthermore, TSPAN1 has been
demonstrated to promote cell migration and invasion in human
cervical tumor cells (28). The
present study assessed the role of TSPAN1 in the progression of PC.
The results demonstrated that the ectopic expression of TSPAN1
promoted PC cell migration and invasion. Consistent with the
results of previous studies, the silencing of TSPAN1 suppressed PC
cell migration and invasion (12,27,29).
These results indicate that TSPAN1 may be an oncogenic gene.
A complex, continuous multi-step process is involved
in the invasion and metastasis of PC cells. Among these, the
proteolytic degradation of the extracellular matrix (ECM) is an
essential event (30).
Membrane-type metalloproteinase directly or indirectly degrades
various components of the ECM including collagens, fibronectin,
laminins, HS proteoglycans and vitronectin (31,32).
MMP2 and MMP9, which are members of the MMP protein family, cleave
collagen and other types of ECM (33,34).
Previous studies have revealed a close association between MMP2 and
MMP9, and PC invasion and metastasis, which is further supported by
the high expression of MMP2 and MMP9 demonstrated in PC tissue
samples of the present study (35,36).
The present results also revealed that the knockdown of TSPAN1
significantly downregulated the expression of MMP2, but did not
affect MMP9 mRNA expression. In addition, MMP2 expression in PC
cells was markedly increased following TSPAN1 ectopic
overexpression. Therefore, the downregulation of MMP2 expression
maybe one way in which the knockdown of TSPAN1 inhibits PC cell
migration and invasion. Furthermore, the present results indicated
that PLCγ siRNA significantly reduced cell migration and invasion,
and the expression of MMP2 induced by TSPAN1 overexpression,
indicating that TSPAN1 siRNA inhibits PC cell migration and
invasion and that MMP2 expression may be dependent on PLCγ. To
confirm this hypothesis, the effect of TSPAN1 siRNA transfection on
the activation of PLCγ was assessed. The results demonstrated that
TSPAN1 siRNA transfection inhibited the translocation and
phosphorylation of PLCγ1, and thus inhibited the activation of
PLCγ1. In conclusion, the results of the present study indicate
that the knockdown of TSPAN1 suppresses PC cell migration and cell
invasion by downregulating MMP2 induced by PLCγ. The results
indicate that TSPAN1 may serve as an important biomarker for the
diagnosis and prognosis of PC. Therefore, TSPAN1-targeted siRNA may
be a promising therapy for PC and other types of cancer with
increased TSPAN1 expression. However, future studies should assess
the feasibility of this therapy in animal models to confirm these
results.
Acknowledgements
Not applicable.
Funding
The present study was funded by the Liaoning
Doctoral Scientific Research Initial Foundation (grant no.
201601417), the Project of Shenyang Municipal Science and
Technology Bureau (grant no. 18-014-4-75), the National Natural
Science Foundation (grant no. 81672427) and the National Natural
Science Foundation of China (grant no. 30973501).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Author's contributions
XZ, GS and XT conceived and designed the present
study. XZ, GS, FG and PL performed the experiments. FG, PL and HW
analyzed the data and wrote the manuscript. GS and XT wrote the
manuscript. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
The present study was performed after obtaining
approval from China Medical University. Samples were taken from
China Medical University with official written ethical consent from
the patients.
Patient consent for publication
All patients provided their written informed consent
for Publication and agreed to the publication of their associated
data and any accompanying images as appropriate.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Vincent A, Herman J, Schuick R, Hruban RH
and Goggins M: Pancreatic cancer. Lancet. 378:607–620. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel R, Ma J, Zou Z and Jemal A: Cancer
statistics, 2014. CA Cancer J Clin. 64:9–29. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Bosetti C, Bertuccio P, Negri E, La
Vecchia C, Zeegers MP and Boffetta P: Pancreatic cancer: Overview
of descriptive epidemiology. Mol Carcinog. 51:3–13. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Loos M, Kleeff J, Friess H and Büchler MW:
Surgical treatment of pancreatic cancer. Ann N Y Acad Sci.
1138:169–180. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Mitchem JB, Hamilton N, Gao F, Hawkins WG,
Linehan DC and Strasberg SM: Long-term results of resection of
adenocarcinoma of the body and tail of the pancreas using radical
antegrade modular pancreatosplenectomy procedure. J Am Coll Surg.
214:46–52. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Nakai Y, Isayama H, Sasaki T, Sasahira N,
Tsujino T, Toda N, Kogure H, Matsubara S, Ito Y, Togawa O, et al: A
multicentre randomised phase II trial of gemcitabine alone vs
gemcitabine and S-1 combination therapy in advanced pancreatic
cancer: GEMSAP study. Br J Cancer. 106:1934–1939. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Long Y, Sun Q, Wu J, Wang Y and Jiao S:
Allogeneic cell-based immunotherapy combined with chemotherapy and
targeted therapy in advanced pancreatic cancer with metastases: A
case report. Oncol Lett. 7:1594–1598. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hall WA, Colbert LE, Nickleach D,
Switchenko J, Liu Y, Gillespie T, Lipscomb J, Hardy C, Kooby DA,
Prabhu RS, et al: The influence of radiation therapy dose
escalation on overall survival in unresectable pancreatic
adenocarcinoma. J Gastrointest Oncol. 5:77–85. 2014.PubMed/NCBI
|
|
9
|
Hemler ME: Tetraspanin functions and
associated microdomains. Nat Rev Mol Cell Biol. 6:801–811. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yáñez-Mó M, Barreiro O, Gordon-Alonso M,
Sala-Valdés M and Sánchez-Madrid F: Tetraspanin-enriched
microdomains: A functional unit in cell plasma membranes. Trends
Cell Biol. 19:434–446. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Stipp CS, Kolesnikova TV and Hemler ME:
Functional domains in tetraspanins proteins. Trends Biochem Sci.
28:106–122. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Liu L, He B, Liu WM, Zhou D, Cox JV and
Zhang XA: Tetraspanin CD151 promotes cell migration by regulating
integrin trafficking. J Biol Chem. 282:31631–31642. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Richardson MM, Jennings LK and Zhang XA:
Tetraspanins and tumor progression. Clin Exp Metastasis.
28:261–270. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Pedro AL: Functional implications of
tetraspanin proteins in cancer biology. Cancer Science.
98:1666–1677. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Garcia-España A, Chung PJ, Sarkar IN,
Stiner E, Sun TT and Desalle R: Appearance of new tetraspanin genes
during vertebrate evolution. Genomics. 91:326–334. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Guo Q, Xia B, Zhang F, Richardson MM, Li
M, Zhang JS, Chen F and Zhang XA: Tetraspanin CO-029 inhibits
colorectal cancer cell movement by deregulating cell-matrix and
cell-cell adhesions. PLoS One. 7:e384642012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ke AW, Shi GM, Zhou J, Wu FZ, Ding ZB, Hu
MY, Xu Y, Song ZJ, Wang ZJ, Wu JC, et al: Role of overexpression of
CD151 and/or c-Met in predicting prognosis of hepatocellular
carcinoma. Hepatology. 49:491–503. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Duan H, Qu L and Shou C: Activation of
EGFR-PI3K-AKT signaling is required for Mycoplasma
hyorhinis-promoted gastric cancer cell migration. Cancer Cell Int.
14:1352014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Chen L, Yuan D, Zhao R, Li H and Zhu J:
Suppression of TSPAN1 by RNA interference inhibits proliferation
and invasion of colon cancer cells in vitro. Tumori. 96:744–750.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2−ΔΔCT method. Methods. 25:402–408. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Albrethsen J, Bøgebo R, Gammeltoft S,
Olsen J, Winther B and Raskov H: Upregulated expression of human
neutrophil peptides 1, 2 and 3 (HNP 1–3) in colon cancer serum and
tumours: A biomarker study. BMC Cancer. 5:82005. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Cordeiro C and Freire AP: Digitonin
permeabilization of Saccharomyces cerevisiae cells for in
situ enzyme assay. Anal Biochem. 229:145–148. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chen CA and Okayama H: Calcium
phosphate-mediated gene transfer: A highly efficient transfection
system for stably transforming cells with plasmid DNA.
Biotechniques. 6:632–638. 1988.PubMed/NCBI
|
|
24
|
Takahashi M, Sugiura T, Abe M, Ishii K and
Shirasuna K: Regulation of c-Met signaling by the tetraspanin
KAI-1/CD82 affects cancer cell migration. Int J Cancer.
121:1919–1929. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Fei Y, Wang J, Liu W, Zuo H, Qin J, Wang
D, Zeng H and Liu Z: CD151 promotes cancer cell metastasis via
integrins α3β1 and α6β1 in vitro. Mol Med Rep. 6:1226–1230.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Haeuw JF, Goetsch L, Bailly C and Corvaia
N: Tetraspanin CD151 as a target for antibody-based cancer
immunotherapy. Biochem Soc Trans. 39:553–558. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Chen L, Zhu Y, Li H, Wang GL, Wu YY, Lu
YX, Qin J, Tuo J, Wang JL and Zhu J: Knockdown of TSPAN1 by RNA
silencing and antisense technique inhibits proliferation and
infiltration of human skin squamous carcinoma cells. Tumori.
96:289–295. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Hölters S, Anacker J, Jansen L,
Beer-Grondke K, Dürst M and Rubio I: Tetraspanin 1 promotes
invasiveness of cervical cancer cells. Int J Oncol. 43:503–512.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Lu Z, Luo T, Nie M, Pang T, Zhang X, Shen
X, Ma L, Bi J, Wei G, Fang G and Xue X: TSPAN1 functions as an
oncogene in gastric cancer and is downregulated by miR-573. FEBS
Lett. 589:1988–1994. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ellenrieder V, Adler G and Gress TM:
Invasion and metastasis in pancreatic cancer. Ann Oncol. 10 (Suppl
4):S46–S50. 1999. View Article : Google Scholar
|
|
31
|
Nagase H and Woessner JF Jr: Matrix
metalloproteinases. J Biol Chem. 274:21491–21494. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Curran S and Murray GI: Matrix
metalloproteinases in tumour invasion and metastasis. J Pathol.
189:300–308. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Bramhall SR, Neoptolemos JP, Stamp GW and
Lemoine NR: Imbalance of expression of matrix metalloproteinases
(MMPs) and tissue inhibitors of the matrix metalloproteinases
(TIMPs) in human pancreatic carcinoma. J Pathol. 182:347–355. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Khokha R and Denhardt DT: Matrix
metalloproteinases and tissue inhibitor of metalloproteinases: A
review of their role in tumorigenesis and tissue invasion. Invasion
Metastasis. 9:391–405. 1989.PubMed/NCBI
|
|
35
|
Koshiba T, Hosotani R, Wada M, Fujimoto K,
Lee JU, Doi R, Arii S and Imamura M: Detection of matrix
metalloproteinase activity in human pancreatic cancer. Surg Today.
27:302–304. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Nagakawa Y, Aoki T, Kasuya K, Tsuchida A
and Koyanagi Y: Histologic features of venous invasion, expression
of vascular endothelial growth factor and matrix
metalloproteinase-2 and matrix metalloproteinase-9, and the
relation with liver metastasis in pancreatic cancer. Pancreas.
24:169–178. 2002. View Article : Google Scholar : PubMed/NCBI
|