Contents
Introduction
Brain-derived neurotrophic factor (BDNF)
Receptors of BDNF and signalling pathways
BDNF has neuroprotective effect in experimental
stroke models
Applications and challenges
Conclusion
Introduction
Hypoxia-ischemia (H/I) brain injury results in
various degrees of damage to the body, and the immature brain is
particularly fragile to oxygen deprivation, termed hypoxia-ischemia
brain damage (HIBD), which can be caused by extreme prematurity or
perinatal asphyxia. For adolescents or adults, similar pathological
changes are often caused by hypertension or aneurysm rupture,
termed ischemic stroke. These processes resulted in
hypoxic-ischemic encephalopathy (HIE).
Hypothermia and erythropoietin (EPO) have long been
known to be neuroprotective, based on the pathologic changes in
HIE. For instance, EPO may promote angiogenesis and reduce
apoptosis (1). Currently,
brain-derived neurotrophic factor (BDNF) is also considered to be a
potent modulator, beneficial to neuronal functions. In this review,
we first examined the background of BDNF, then focused on its
neuro-protective mechanisms against ischemic stress, and discussed
the potential application of BDNF in clinic
Brain-derived neurotrophic factor
Forms
Nerve growth factor (NGF), BDNF, neurotrophins (NT)
(NT-3, NT-4, NT 4/5 and NT-6) constitute the protein family of
mammalian NT (2,3). BDNF was discovered in 1982, originally
described as a small dimeric protein (4). There are two types of BDNF: pro- and
mature BDNF, present in the human body (5). Pro-BDNF is a 32-kDa precursor,
comprising 247 amino acids with N-glycosylated and glycosulphated
residues within the prodomain (6).
Following the initial generation, most of the pro-BDNF is then
packaged into vesicles in a regulated pathway and undergoes
N-terminal cleavage by extracellular proteases, such as plasmin,
metalloproteinase gene matrilysin (MMP7) (7,8),
tPA/plasmin cascade (8) and
extracellular matrix-metalloproteinases (9). In trans-Golgi network (TGN), the
‘pro-region’ is cleaved resulting in the formation of mature BDNF
(14 kDa), a biologically active form with C-terminal dimers
(10). This mature BDNF is then
released mainly by the neurons through constitutive secretion or in
an activity-dependent manner (6).
Another subtype of pro-BDNF, small amounts of a 28-kDa protein, was
identified by immunoprecipitation with BDNF antibodies. However, it
was not an obligatory intermediate in the formation of the mature
BDNF (11).
Proproteins must undergo a variety of
post-translational processes to yield biologically active peptides.
The two forms of extracellular BDNF, pro- and mature BDNF, act in
different ways. Mature BDNF is crucial in the protection of the
neonatal or developing brain from ischemia injury (12). In cultured hippocampal neurons, low-
and high-frequency neuronal activities increased pro-BDNF levels
(5). However, only high-frequency
activity induces tissue plasminogen activator secretion, resulting
in the conversion of pro- to mature BDNF (13). Additionally, the highest levels of
pro-BDNF are observed perinatally, then it declines with age,
although the proform remains detectable in adulthood. These data
partly provide the reason that brains of newborns and infants are
more fragile to ischemia stroke due to low-frequency neuronal
activities and the lack of an adequate amount of mature BDNF in the
central nervous system (CNS).
BDNF Val66Met is a common single-nucleotide
polymorphism (SNP) in the human BDNF gene resulting in a valine
(Val) to methionine substitution in the prodomain, termed Val66Met.
The BDNF Val66Met has shed light on psychiatric studies,
particularly in schizophrenia, anxiety-like behavior and depressive
symptoms (14), which belong to the
sequelae of H/I damage.
Synthesis and location
BDNF is broadly expressed in the developing and
adult mammalian brain (15),
synthesized in several areas of the hypothalamus, including the
paraventricular (PVN), ventromedial (VMN) and dorsomedial nuclei
(DMN), as well as the lateral hypothalamic area (LH). Accordingly,
BDNF mRNA and proteins are widespread in almost all the cortical
areas as well as other tissues, including the neural soma,
dendrites, fibres and amygdala (16).
Large amounts BDNF are believed to be stored or
secreted from non-neuron cells, when attacks occur, such as human
platelets (17). It was also found
to be present in the ependymal, microglial and endothelial cells of
cerebral arterioles and astrocytes, respectively (18). Peripherally, BDNF accumulates in the
vascular endothelium, neuromuscular synapse, muscle and liver
tissue (19), which is essential
for neuronal repair when stroke occurs.
BDNF gene
The rodent BDNF gene was initially described by Aid
et al (20) and comprises at
least eight distinct promoters, initiating the transcription of
multiple distinct mRNA transcripts, comprising four 5′-exons (I–IV)
linked to separate promoters, and one 3′-exon (V) that contains the
entire open reading frame for the BDNF protein. Pruunsild et
al (21) have identified new
splice variants in human and rodents, respectively, demonstrating
that at least 11 different BDNF transcripts can be generated from
the mammalian rodent BDNF gene by alternative splicing. The
activation of various BDNF promoters is region-specific and depends
on the type of stimulus (22). A
single BDNF protein is produced from several splice variants with
different 5′-UTRs (23,24).
Receptors of BDNF and signalling
pathways
If left uncleaved, pro-BDNF selectively activates
its high-affinity receptor, the p75 receptor, mainly inducing
pro-apoptotic signalling pathways (25). Mature BDNF binds with high
specificity to the tropomyosin-related kinase receptor type B
(TrkB) (26) and to the
low-affinity neurotrophin receptor p75, then exerts its actions via
interactions between these two trans-membrane receptors, separately
or in collectively, potentially leading to neuronal death or
survival.
TrkB
The Trk receptor tyrosine kinase family includes
TrkA and TrkC, which are receptors for NGF and NT3, respectively.
The family also includes TrkB, which mediates the effects of BDNF
and NT 4/5 (27).
BDNF exerts multiple biological actions through TrkB
receptors (13,28). Similar to BDNF, TrkB is widely
expressed in the adult brain, including the cortex, hippocampus,
multiple brain stem and spinal cord nuclei (29).
Several TrkB isoforms have been observed in the
mammalian CNS. The full-length TrkB isoform is a typical tyrosine
kinase wherein homodimerization during ligand binding causes
intracellular cross-tyrosine phosphorylations (30). In addition, truncated forms of TrkB
[T1 and T2 in rat; T1 and T-Src homology and collagen protein
(T-Shc) in humans] lacking the tyrosine kinase component of the
receptor, are found in neurons and glia. T1 is predominantly
expressed in the brain and may act as a dominant negative inhibitor
of BDNF signalling by forming heterodimers with full-length TrkB
(31).
BDNF binding to TrkB triggers autophosphorylation of
the tyrosine residue in its intracellular domain, leading to
ligand-induced dimerization in each receptor, which activates
several intracellular signalling pathways with various functions
(32). More specifically, when NT
binds to Trk receptors, three enzymes are considered to be the main
regulators: mitogen-activated protein kinase (MAPK),
phosphatidylinositol-3 kinase (PI3K) (33) and phospholipase C γ (PLCγ) (34).
Trk family members recruit and increase the
phosphorylation of PLCγ and the Src homology and collagen protein
(Shc). Binding of the adaptor proteins Shc and the growth factor
receptor substrates 2 (FRS-2) to Trk leads to the activation of the
PI3K/Akt and MAPK pathways (35).
Several small G proteins, including Rap-1 and the Cdc-42-Rac-Rho
family, also participate in this process (34). The role of the ERK and PI3-kinase
pathways in neonatal H/I brain injury in the presence of BDNF is
gradually becoming clear (36,37).
Docking of PLCγ to a separate site of TrkB leads to the production
of diacylglycerol, a transient activator of protein kinase C (PKC)
and inositol trisphosphate (IP3), and eventually mobilizes
intracellular calcium (38).
Trk receptor activation has variable downstream
specificity, and significant cross-talk is observed among the sites
of action in these three pathways.
p75
p75, also known as p75NTR (p75 neurotrophin
receptor), is a member of the tumor necrosis factor receptor (TNF)
superfamily. In adulthood its expression is restricted to basal
forebrain cholinergic neurons and is present in relatively few
cortical neurons (39). p75 is
mainly expressed during early neuronal development, whereas in
adults, p75 is re-expressed in various pathological conditions,
including epilepsy, axotomy and neurodegeneration (40).
The low-affinity receptor p75 binds to
pro-neurotrophin with high affinity, transmitting positive and
negative intracellular signals. It is particularly significant in
mediating pro-neurotrophin signalling and often induces inversed
biological effects on TrkB receptors (41). When compared to mature BDNF,
pro-BDNF promotes neuronal survival via TrkB, preferentially
activating p75 to mediate neuronal cell death, particularly
apoptosis (42). Therefore, the
amount of pro-BDNF is critical in neuronal cell death.
An analysis of the spatiotemporal profile of p75
expression after an ischemic lesion induced by cortical
devascularization (CD) demonstrated that p75 expression is
expressed in isolated neurons within the ischemic lesion site.
These p75+ neurons present morphological alterations and active
caspase-3 staining. Peak p75 expression has been shown to occur 3
days post-lesion in the penumbra. Therefore, the authors conclude
that p75 expression is localized in selected neurons in the
ischemic lesion and that these p75+ neurons are probably condemned
to apoptotic cell death (39).
Several signalling pathways have been implicated in
the actions of p75 in the absence of Trk receptors, including the
induction of nuclear factor-κ B (NF-κB) and c-Jun kinase activities
(43).
Chaperone proteins
Chaperone proteins, carboxypeptidase E (CPE)
(44) and sortilin (14), are two additional receptors of BDNF.
The binding of BDNF to the lipid-raft-associated sorting receptor
CPE in the TGN is necessary for sorting into secretory vesicles.
CPE is subsequently internalized and transported through the
endocytic recycling compartment back to the TGN (45). Most BDNF in this regulated secretory
pathway is transported to post-synaptic dendrites and spines
(46).
Sortilin interacts specifically with BDNF in a
region encompassing methionine substitution and co-localizes with
BDNF in secretory granules in neurons. Certain p75+ neurons are
also positive for sortilin (47).
CPE and sortilin have thus been identified as candidate proteins
that potentially regulate the intracellular localization of BDNF
within neurons (48). The ways BDNF
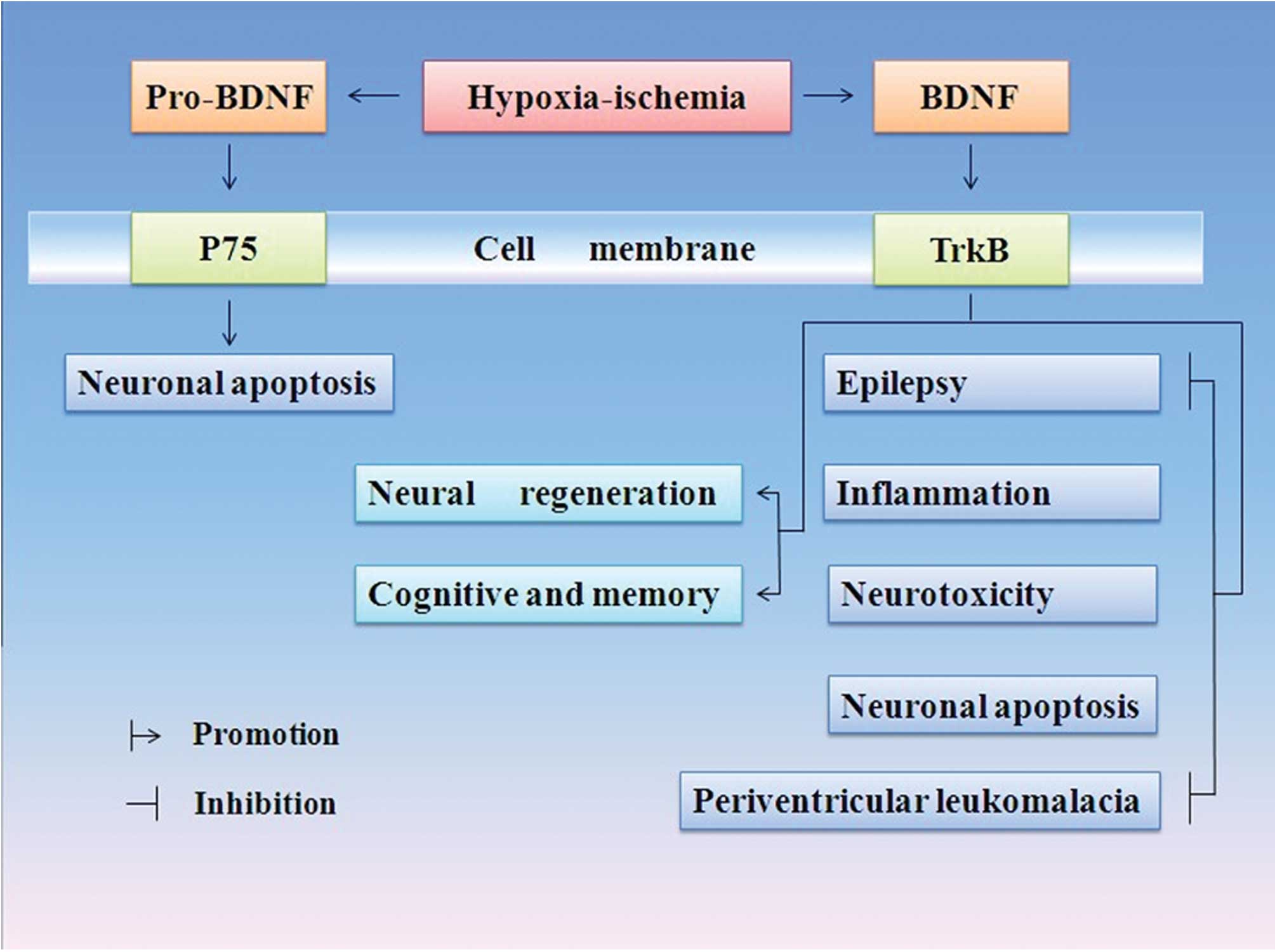
exhibits positive effects is shown in Fig. 1.
BDNF has neuroprotective effect in
experimental stroke models
The major neuropathology after H/I insult begins
during the acute insult and extends into the reperfusion phase
(49). The progress is short for
ischemia/reperfusion (I/R) injury. It may involve the following
pathophysiologic aspects: i) apoptosis; ii) free-radical generation
and activation of inflammatory mediators, e.g., acidosis (50); iii) excessive extracellular
glutamate excitotoxicity and intracellular accumulation of calcium
(51) and iv) depleted energy
reserves and loss of high-energy phosphate compounds. Thus, energy
deprivation along with increased levels of harmful factors, either
intracellular or extracellular, disrupts neuronal homeostasis.
Consequently, clinical manifestations, such as periventricular
leukomalacia, epilepsy, cognition and memory deficiency were also
present.
In the subsequent sections, the mechanism of the
action of BDNF in multiple protective roles against ischemic brain
injury is examined.
Anti-apoptosis
Evidence showed that BDNF was beneficial for the
survival of neurons through anti-apoptotic effect. Infected with
adeno-associated viral vector inserted with BDNF gene (AAV-BDNF),
neurite cells may be able to produce BDNF function to promote its
own outgrowth and protect neurons from serum deprivation-induced
apoptosis (52). Besides, cultured
rat hippocampal neurons were injured by amyloid β and then infected
with AAV-BDNF to examine the neuroprotective effects of BDNF. The
results showed that the Ca2+ balance was maintained in
the AAV-BDNF treatment group, and that BDNF may reduce neuron
apoptosis by increasing the expression of the Bcl-2 anti-apoptosis
protein and inhibiting intracellular calcium overload (53).
In vitro, 2.1 μg/day BDNF were
delivered continuously via intraventricular infusion pumps. The
mean infarct volume after venous occlusion was significantly
smaller in BDNF-treated rats at 2 days (1.49±1.44 vs. 3.66±1.51%),
and fewer TUNEL-positive apoptotic cells were detected 2 days later
(17.0±15.1 vs. 39.0±19.6) compared to the controls (54). Similarly, in global ischemia induced
by a four-vessel occlusion in rat, 0.06 mg/h BDNF diluted in
artificial cerebral spinal fluid was administered by an osmotic
minipump, which was implanted after reperfusion. Data showed that
the pyramidal cell count was 439.6±18.5 in the BDNF group, and
18.3±10.6 in the ischemia group at day 7 (55). When 5 mg human BDNF was injected
intravitreally after H/R, the number of TUNEL- and
caspase-2-positive cells in the BDNF-treated group vs. the control
was 545.2±29.7 vs. 22370.3±122.5 cells/mm2 and
124.4±35.4 vs. 244.6±15.7 cells/mm2 at 6 h after
reperfusion (56). Additionally, in
a postnatal day 7 rat model, H/I injury to the developing brain is
a strong apoptotic stimulus leading to caspase-3 activation,
although BDNF can block this process in vivo (28).
Anti-inflammation
Inflammation responds to cerebral ischemia rapidly,
activates the local inflammatory cells (mostly microglia),
producing relevant mediators and translocation of intercellular
nuclear factors.
Cytokines and chemokines that trigger leukocyte
infiltration or glial activation and proliferation, are released
following ischemia, and might either be beneficial or detrimental.
A possible contributor to this dichotomy of responses depends on
the degree to which proximal neurons are injured. Twenty four hours
after hypoxia-exposure of the neuronal cultures, the classic
microglial proinflammatory mediators, including inducible nitric
oxide synthase (iNOS), TNF and interleukin-1-β (IL-1β), are
upregulated only in response to mild neuronal injuries, while the
trophic microglial effector BDNF is upregulated in response to the
degrees of neuronal ischemia injuries (57,58).
In the inflammatory process, BDNF may promote
microglial proliferation and phagocytic activity in vitro
(59) and increase the number of
phagocytotic microglia and activated microglia that, in turn,
secretes BDNF (60). When
conditioned media from injured neurons [neuron-conditioned media;
(NCM)] were added to microglial cultures following H/I, BDNF
released from microglia was upregulated, suggesting that BDNF might
contribute to the anti-inflammatory activity induced by microglia
(61). TNF-α has been proven to
exacerbate cerebral injury of ischemia (62), while interleukin (IL-10), an
anti-inflammatory cytokine, has a neuroprotective role in ischemic
stroke (63). Jiang et al
(60) reconfirmed that intranasal
administration of BDNF in H/I rats can suppress TNF-α and its mRNA
expression, while increasing IL-10 and its mRNA expression. Peng
et al (64) treated neural
stem cells (NSCs) with BDNF siRNA, and found that imipramine (IM)
increases the neuroprotective effects, suppresses the inflammatory
process in NSCs via the modulation of the MAPK pathway and Bcl-2
cascades, indirectly evaluating the anti-inflammatory effect of
BDNF.
Anti-neurotoxicity
Depletion of glucose and oxygen supply causes a
primary energy failure and initiates a cascade of biochemical
events leading to cell dysfunction. A consequent reperfusion injury
often deteriorates the brain metabolism by increasing the oxidative
stress damage.
Neurotoxicant trimethyltin (TMT) induces a
significant reduction of cell survival, neuronal differentiation
and concomitant earlier activation of cleaved caspase-3. However,
overexpression of BDNF firmly protects differentiated NSC against
TMT-induced neurotoxicity through the PI3K/Akt and MAPK-signaling
pathways (65). Addition of 100 mM
ethanol to a human neuronal cell model, SH-SY5Y cells, showed the
secreted amount of BDNF and the cyclic AMP-responsive
element-binding protein (CREB) activity to be significantly reduced
by ethanol. Additionally, exogenous BDNF has a protective effect
against ethanol-induced damage in primary culture neurons from rat
hippocampi (66).
BDNF resists glutamate cytotoxicity depending on its
concentration (67). Glutamate is
the major excitatory neurotransmitter in the brain, always binding
its receptor glutamate receptors N-methyl-D-aspartate receptor
(NMDAR) under pathological circumstances, causing ascendency of
cytosolic calcium (68). Following
H/I, the concentrations of glutamate and ATP are increased
(61), and excitatory amino acids
(EAA) are secreted, allowing glutamate to accumulate to excitotoxic
levels. BDNF inhibited neurotoxicity induced by glutamate and NO
donors in cultured cortical neurons, especially dopamine neurons
(69).
Furthermore, BDNF mRNA accumulates in distal
dendrites to activate NMDAR and TrkB receptor (70), the former might have pro-apoptotic
excitotoxic activity. By contrast, signalling via TrkB has been
largely considered to protect neurons antagonizing the
NMDAR-mediated excitotoxic cell death. The cross-talk and feedback
loops between BDNF and the NMDAR signalling was reviewed by
Georgiev et al (71).
Promotion of neural regeneration
Neurogenesis involves cell proliferation, migration
and differentiation (46). To
facilitate regeneration among central and peripheral neurons after
H/I, the enrichment of BDNF around the injured region is essential.
Zhu et al (72) evaluated
functional recovery following the transplantation of BDNF-modified
neural stem cells (NSCs) in a rat model of cerebral ischemia damage
induced by temporary middle cerebral artery occlusion (tMCAO).
Their findings showed that BDNF protein expression in rat embryonic
NSCs transfected with the human BDNF gene (BDNF-NSCs) was
upregulated, while neurite outgrowth in ganglion neurons were
simulated, suggesting that BDNF increased neurogenesis in vitro.
In vivo, BDNF promoted recovery of temporary middle cerebral
artery occlusion. Zhu et al also assessed the neurological
function deficiency for 12 weeks using the neurological severity
score (NSS). NSS was significantly lower in the
BDNF-NSC-transfected transplant group compared to the control
groups for the 10-week time period (72).
BDNF may allow sustained regenerative signalling at
synaptic sites (35). It induces
structural instability in dendrites and spines restricted to
particular portions of a dendritic arbor, and may help translate
activity patterns into specific morphological changes (73). Furthermore, BDNF may increase the
expression of markers for axonal sprouting and synaptogenesis, such
as MAP1/2 or synaptophysin. Post-ischemic intravenous BDNF
treatment improves functional motor recovery after thrombotic
stroke (74).
Angiogenesis, another contribution of BDNF should be
mentioned. Injection of BDNF fused with a collagen-binding domain
(CBD-BDNF) into the lateral ventricle of MCAO rats, promoted neural
regeneration and angiogenesis. Induction of neural differentiation
of adipose-derived mesenchymal stem cells (ASCs) led to nerve
repair and growth also via BDNF production. Nerve fiber length in
ASCs matrigel implants was 1.3-fold greater compared to the control
(75).
Protection against periventricular
leukomalacia (PVL)
In premature infants, the H/I damage to the cerebral
white matter usually involves PVL. Selected neuronal circuits as
well as immature periventricular oligodendroglia, may die from the
excitotoxicity, leading to chronic neurologic disability with
cerebral palsy (76). In a previous
study, Husson et al (77)
injected rats with ibotenate generating white matter cysts
resembling those detected in PVL. Those authors found that such
white matter cysts in cortical and white matter lesions are reduced
by BDNF. However, the exact effect was dependent upon the type of
activated glutamate receptors, lesion localization and the
developmental stage (Table I).
 | Table IExperiments on the roles of
brain-derived neurotrophic factor (BDNF) in hypoxic-ischemic (H/I)
injury. |
Table I
Experiments on the roles of
brain-derived neurotrophic factor (BDNF) in hypoxic-ischemic (H/I)
injury.
| Experimental
model | Treatment for
injury | Intervention | BDNF activity for
neuroprotection | Refs. |
|---|
| BDNF protects the
brain from hypoxic-ischemic injury |
|
| Cerebellar granule
neurons culture | H/I
Low K+ (5 mM) | BDNF 100 ng/ml | BDNF has a direct
effect on mature cerebellar granule neurons and can protect these
neurons from apoptosis in low K+ | (84) |
| Cerebellar granule
neurons culture | With K+
(5 mM) | BDNF 1–100
ng/ml | BDNF protects from
K+/serum deprivation-induced apoptotic death of
cerebellar granule neurons | (33) |
| Cerebellar granule
neurons culture | H/I
Low K+ (5 mM) | BDNF 100 ng/ml | BDNF in the
anti-apoptotic effect of NMDA in cerebellar granule neurons | (85) |
Newborn
SD rats | Left common carotid
artery was permanently ligated | BDNF ICV injection
5 μg/animal | BDNF treatment
virtually eliminated the increase in caspase-3-like activity
induced by HI BDNF to block H/I-induced caspase-3 activation and
tissue loss | (86) |
Newborn
SD rats | Left common carotid
artery was permanently ligated, 8% oxygen flowed for 2.5 h | BDNF ICV injection
5 μg/animal | BDNF protects the
neonatal brain from H/I injury in vivo via the ERK pathway,
BDNF provides a promising solution to hypoxic injury due to its
survival-promoting effects | (37) |
| Cultured neurons
from embryonic SD rats | Hypoxia 85%
N2, 5% CO2, 10% H2 | BDNF 100 ng/ml 24 h
prior to hypoxia/immediately | BDNF is highly
involved in preventing cortical neurons from hypoxia-induced
neurotoxicity | (87) |
| Cultured neurons
from embryonic SD rats | Hypoxia 85%
N2, 5% CO2, 10% H2 | BDNF 50 ng/ml 30
min before hypoxia | The activation of
ERK- and AKT-signalling pathway-mediated BDNF neuroprotective
function against hypoxic-induced neurotoxicity | (37) |
| Cultured neurons
from embryonic SD rats | Hypoxia 85%
N2, 5% CO2, 10% H2 | BDNF 25, 50, 100
ng/ml 24 h before hypoxia/immediately | Extrinsic BDNF has
a neuroprotective effect against hypoxic-induced neurotoxicity | (88) |
| Cultured neurons
from embryonic SD rats | Hypoxia 85%
N2, 5% CO2, 10% H2 | BDNF 100 ng/ml 24 h
before hypoxia/immediately | The Ras-MAPK
approach may be the major signal transferring way of BDNF in
protecting the cortical neurons from H/I-induced neurotoxicity | (89) |
| Pregnant rats | Artery clamp | BDNF 2 μg
was injected to caudal veins | BDNF demonstrates
neuroprotective effects on rat embryo brain cells suffering from
intrauterine H/I injury via the ERK signalling pathway | (90) |
| Adult female SD
rats | MCAO 10 min of
four-vessel occlusion | HBO | HBO preconditioning
may be neuroprotective by reducing early apoptosis and inhibition
of the conversion of early to late apoptosis, possibly through an
increased brain BDNF level | (91) |
| Gerbil | Transient cerebral
ischemia | Extract from
TCE | Repeated supplement
of TCE-protected neurons from ischemic damage induced by transient
cerebral ischemia by maintaining BDNF levels | (92) |
| Cultured neurons
from embryonic SD rats | Transient cerebral
ischemia in the hippocampal CA1 pyramidal cells | Escitalopram 30
mg/kg | Pre- and
post-treatments with escitalopram protects against ischemia-induced
neuronal death in the CA1 induced by transient cerebral ischemic
damage by the increase of BDNF | (93) |
| Cultured neurons
from embryonic SD rats | Hypoxia 85%
N2, 5% CO2, 10% H2 | BDNF 50 ng/ml | BDNF has
neuroprotective effect on embryonic rat cortical neurons against
hypoxia via CREB phosphorylation | (31) |
| Cultured neurons
from embryonic SD rats | Hypoxia 85%
N2,5% CO2, 10% H2 | BDNF 50 ng/ml | Hypoxia rapidly
blocking ERKl/2 signalling pathway is involved in the protective
effect of BDNF against hypoxic injury on in vitro-cultured
neurons | (94) |
| Rat | Global
ischemia | 0.06 mg/h BDNF
intracerebroventricularly | BDNF inhibited the
neuronal degeneration in the hippocampal regions | (55) |
| Rats | Cerebral venous
ischemia | BDNF 2.1
μg/day | The mean infarct
volume after venous occlusion was smaller, and fewer TUNEL-positive
apoptotic cells were detected in BDNF-treated rats | (54) |
| Adult male SD
rats | MCAO | LBD-BDNF 0.2 nmol
injected into the right brain | LBD-BDNF-reduced
infarct volume is associated with a parallel improvement in
neurological functional outcome and neurogenesis in the dentate
gyrus of the hippocampi | (86) |
| Rats | Temporary occlusion
of the MCA (120 min) | Intranasal
BDNF | BDNF protects brain
from ischemic insult via modulating local inflammation in rats | (95) |
| Adult female SD
rats | Ischemia (60
min) | Human BDNF 5
μg | The number of
TUNEL- and caspase-2-positive cells was lower in the BDNF-treated
group at 6 h, after reperfusion | (56) |
| Cortical neurons
from rat embryos | H/I | 1, 10, 50, 100
ng/ml | BDNF demonstrated
protection against apoptotic cell death | (28) |
| Rats | MCAO | CBD-BDNF 10
μl/Nat-BDNF 10 μl/lateral ventricle | CBD-BDNF promoted
neural regeneration and angiogenesis, reduced cell loss, decreased
apoptosis and improved functional recovery | (96) |
|
| BDNF inhibition
protects the brain from hypoxic-ischemic injury |
|
| Rat cortical cell
cultures | | BDNF 10, 30, 100
ng/ml | BDNF-induced
neuronal necrosis was accompanied by reactive oxygen species
production | (97) |
| Mixed cortical cell
cultures | Serum-free
EMEM | BDNF 100 ng/ml | The role of NADPH
oxidase in oxidative neuronal death induced in cortical cultures by
BDNF | (98) |
Anti-epilepsy
Various studies have demonstrated that BDNF
contributes to epileptogenesis (78). For example, mesio-temporal lobe
epilepsy (MTLE) was significantly aggravated in mice with increased
TrkB signals, but delayed in mutant mice with reduced TrkB signals.
Paradoxically, with respect to temporal lobe epilepsy (TLE),
previous studies have demonstrated that BDNF-induced Trk activation
may lead to neuropeptide Y (NPY) upregulation, while NPY-knockout
animals are more susceptible to seizures. Therefore,
intrahippocampal infusion of BDNF potentially attenuates (or
retards) the development of epilepsy (79).
Contributions to cognitive functions and
memory acquisition
Cerebral ischemia may lead to a progressive global
cognitive deterioration. The involvement of BDNF in cognitive
functions, particularly in memory acquisition and consolidation is
highly attractive.
BDNF is essential for NSC-induced cognitive rescue,
which has been observed in aged 3x transgenic Alzheimer’s disease
(Tg-AD) mice with spatial learning and memory deficiencies.
Gain-of-function studies demonstrated that recombinant BDNF mimics
might have beneficial effects on NSC transplantation, while
loss-of-function studies showed that mice depleted of NSC-derived
BDNF failed to improve cognition or restore hippocampal synaptic
density (80). To investigate the
effect of BDNF on hippocampal cognitive functions after global
cerebral ischemia in rats, BDNF was administered continuously over
14 days via an osmotic mini-pump, intracerebroventricularly after
four-vessel occlusion. Cognitive impairment was also assessed
repeatedly using a passive avoidance test. In ischemic animals
treated with BDNF, the working and reference memory ratios 15 days
after ischemia were lower in the ischemic rats. These data indicate
a protective effect of BDNF for synaptic transmission and cognitive
functions after transient forebrain ischemia (81).
Furthermore, voluntary exercise upregulates BDNF
within the hippocampus, inducing improvements in cognitive
performance after traumatic brain injury in rats (82).
With regard to the function of memory, BDNF has been
known to induce memory persistence, and convert a non-lasting
long-term memory (LTM) trace into a persistent one. When BDNF gene
expression in the hippocampus was inhibited, a deficiency of memory
formation was observed (83).
Applications and challenges
While designing treatment strategies aimed at
improving stroke recovery, greater attention should be paid to
non-neuronal cells which are able to produce substantial amounts of
BDNF after ischemic stroke. Evidence has shown that ischemic stroke
in rats results in increased BDNF staining in neurons and ependymal
cells in the non-lesioned hemisphere. Similarly, in the lesioned
hemisphere, microglial and endothelial cells of cerebral arterioles
and astrocytes also exhibit robust BDNF staining (18).
Transposition of BDNF to the target injury regions
is a challenge in clinical applications, while a short half-life
and a low rate of transport through the blood-brain barrier (BBB)
is hampered. Such problems may be solved in various ways.
First, by fusing a laminin-binding domain (LBD) to
BDNF a laminin-binding BDNF (LBD-BDNF) form is constructed, since
laminin is a rich extracellular matrix in the CNS, and is highly
expressed in ischemic regions. LBD-BDNF is associated with a
parallel improvement in neurological functional outcomes, and
effectively attenuates neural degeneration after permanent middle
cerebral artery occlusion in rats (86). Similarly, injection of CBD-BDNF in
the ventricle remained stable for much longer compared to mature
BDNF, while the CBD-BDNF concentrated at the infarcted hemisphere
and exerted a more enduring therapeutic effect (96).
Second, the problem may also be resolved by
improving BDNF delivery into the target region. Studies have
suggested the intranasal access to potentially be effective when
delivering BDNF to the target region. Elevated concentration of
BDNF in brain tissues following intranasal delivery can reach 4
ng/g, as opposed to only 0.2 ng/g in the controls (60).
Third, BDNF mimetics may also be used to overcome
the therapeutic challenges. Based on a loop domain of BDNF that
binds to a key receptor of TrkB, pharmacophores were generated.
Four candidate molecules designated as LM22A1-LM22A4 were selected.
In mouse hippocampal neuronal cultures, these compounds promoted
cell survival with an efficacy comparable to that of BDNF. Of note,
unlike BDNF, LM22A4 did not bind to the receptor p75, which is
considered to mediate the pain-promoting effects of BDNF.
Furthermore, LM22A4 was considered suitable for intranasal
administration to mice. Once-daily dosing of this compound for 7
days in in vivo experiments, not only increased the
activation of TrkB in the hippocampus and striatum, but also
significantly improved the impairment in motor learning, following
traumatic brain injury. Such mimetics provide a promising new
approach to the application of BDNF in the treatment of H/I injury
(99).
Conclusion
During the last decade, the neuroprotective effects
of BDNF, its underlying mechanisms and signal transductions have
been investigated. Evidence from in vitro studies as well as
animal models have demonstrated that BDNF is a potential novel
candidate of defence against ischemia brain injury. However, since
the signalling pathway is complicated and bidirectional,
application of BDNF in neuroprotection in humans remains to be
elucidated. Therefore, additional studies focusing on BDNF, its
mechanisms or application, need to be conducted in the future.
Abbreviations:
|
BDNF
|
brain-derived neurotrophic factor;
|
|
HI
|
hypoxia-ischemia;
|
|
HIBD
|
hypoxia-ischemia brain damage;
|
|
HIE
|
hypoxic-ischemic encephalopathy;
|
|
EPO
|
erythropoietin;
|
|
NGF
|
nerve growth factor;
|
|
NT
|
neurotrophins;
|
|
MMP7
|
metalloproteinase gene matrilysin;
|
|
TGN
|
trans-Golgi network;
|
|
PVN
|
paraventricular nucleus;
|
|
VMN
|
ventromedial nucleus;
|
|
DMN
|
dorsomedial nucleus;
|
|
LH
|
lateral hypothalamic area;
|
|
SNP
|
single-nucleotide polymorphism;
|
|
Val
|
valine;
|
|
Met
|
methionine;
|
|
TrkA
|
kinase receptor type A;
|
|
TrkB
|
kinase receptor type B;
|
|
TrkC
|
kinase receptor type C;
|
|
CNS
|
central nerve system;
|
|
MAPK
|
mitogen-activated protein kinase;
|
|
PI-3K
|
phosphatidylinositol-3 kinase;
|
|
PLCγ
|
phospholipase C γ;
|
|
c-Jun
|
c-Jun kinase;
|
|
NF-κB
|
nuclear factor κ-light-chain-enhancer
of activated B cells;
|
|
PKC
|
protein kinase C;
|
|
IP3
|
inositol trisphosphate;
|
|
TNF
|
tumor necrosis factor receptor;
|
|
CD
|
cortical devascularization;
|
|
CPE
|
carboxypeptidase E;
|
|
mTOR
|
mammalian target of rapamycin;
|
|
ERK
|
extracellular signal-regulated
kinases;
|
|
CREB
|
cAMP response element-binding
protein;
|
|
NCM
|
neuron-conditioned media;
|
|
NSCs
|
neural stem cells;
|
|
IM
|
imipramine;
|
|
EAA
|
excitatory amino acids;
|
|
NMDAR
|
N-methyl-D-aspartate receptor;
|
|
PVL
|
periventricular leukomalacia;
|
|
MTLE
|
mesio-temporal lobe epilepsy;
|
|
TLE
|
temporal lobe epilepsy;
|
|
NPY
|
neuropeptide Y;
|
|
LTM
|
non-lasting long-term memory;
|
|
BBB
|
blood-brain barrier;
|
|
LBD-BDNF
|
laminin-binding BDNF;
|
|
NADPH
|
nicotinamide adenine dinucleotide
phosphate;
|
|
EMEM
|
Eagle’s minimal essential medium
|
Acknowledgements
This study was supported by the
National Natural Science Foundation of China (no. 30973215) and the
Program for Changjiang Scholars and Innovative Research Team in
University (No.IRT 0935).
References
|
1
|
Xiong T, Qu Y, Mu D and Ferriero D:
Erythropoietin for neonatal brain injury: opportunity and
challenge. Int J Dev Neurosci. 29:583–591. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Levi-Montalcini R: The nerve growth factor
35 years later. Science. 237:1154–1162. 1987.PubMed/NCBI
|
|
3
|
Gotz R, Koster R, Winkler C, et al:
Neurotrophin-6 is a new member of the nerve growth factor family.
Nature. 372:266–269. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lessmann V and Brigadski T: Mechanisms,
locations, and kinetics of synaptic BDNF secretion: an update.
Neurosci Res. 65:11–22. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yang J, Siao CJ, Nagappan G, et al:
Neuronal release of proBDNF. Nat Neurosci. 12:113–115. 2009.
View Article : Google Scholar
|
|
6
|
Mowla SJ, Farhadi HF, Pareek S, et al:
Biosynthesis and post-translational processing of the precursor to
brain-derived neurotrophic factor. J Biol Chem. 276:12660–12666.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lu B: Pro-region of neurotrophins: role in
synaptic modulation. Neuron. 39:735–738. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Pang PT, Teng HK, Zaitsev E, et al:
Cleavage of proBDNF by tPA/plasmin is essential for long-term
hippocampal plasticity. Science. 306:487–491. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lee R, Kermani P, Teng KK and Hempstead
BL: Regulation of cell survival by secreted proneurotrophins.
Science. 294:1945–1948. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Seidah NG, Benjannet S, Pareek S, Chretien
M and Murphy RA: Cellular processing of the neurotrophin precursors
of NT3 and BDNF by the mammalian proprotein convertases. FEBS Lett.
379:247–250. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Goodman LJ, Valverde J, Lim F, et al:
Regulated release and polarized localization of brain-derived
neurotrophic factor in hippocampal neurons. Mol Cell Neurosci.
7:222–238. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Thomas K and Davies A: Neurotrophins: a
ticket to ride for BDNF. Curr Biol. 15:R262–R264. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Boutilier J, Ceni C, Pagdala PC, Forgie A,
Neet KE and Barker PA: Proneurotrophins require endocytosis and
intracellular proteolysis to induce TrkA activation. J Biol Chem.
283:12709–12716. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Chen ZY, Bath K, McEwen B, Hempstead B and
Lee F: Impact of genetic variant BDNF (Val66Met) on brain structure
and function. Novartis Found Symp. 289:180–195. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yu H and Chen ZY: The role of BDNF in
depression on the basis of its location in the neural circuitry.
Acta Pharmacol Sin. 32:3–11. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Tapia-Arancibia L, Rage F, Givalois L and
Arancibia S: Physiology of BDNF: focus on hypothalamic function.
Front Neuroendocrinol. 25:77–107. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yamamoto H and Gurney ME: Human platelets
contain brain-derived neurotrophic factor. J Neurosci.
10:3469–3478. 1990.PubMed/NCBI
|
|
18
|
Bejot Y, Prigent-Tessier A, Cachia C, et
al: Time-dependent contribution of non neuronal cells to BDNF
production after ischemic stroke in rats. Neurochem Int.
58:102–111. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Cassiman D, Denef C, Desmet VJ and Roskams
T: Human and rat hepatic stellate cells express neurotrophins and
neurotrophin-receptors. Hepatology. 33:148–158. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Aid T, Kazantseva A, Piirsoo M, Palm K and
Timmusk T: Mouse and rat BDNF gene structure and expression
revisited. J Neurosci Res. 85:525–535. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Pruunsild P, Kazantseva A, Aid T, Palm K
and Timmusk T: Dissecting the human BDNF locus: bidirectional
transcription, complex splicing, and multiple promoters. Genomics.
90:397–406. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Givalois L, Marmigere F, Rage F, Ixart G,
Arancibia S and Tapia-Arancibia L: Immobilization stress rapidly
and differentially modulates BDNF and TrkB mRNA expression in the
pituitary gland of adult male rats. Neuroendocrinology. 74:148–159.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Tongiorgi E and Baj G: Functions and
mechanisms of BDNF mRNA trafficking. Novartis Found Symp.
289:136–195. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Greenberg ME, Xu B, Lu B and Hempstead BL:
New insights in the biology of BDNF synthesis and release:
implications in CNS function. J Neurosci. 29:12764–12767. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Teng HK, Teng KK, Lee R, et al: ProBDNF
induces neuronal apoptosis via activation of a receptor complex of
p75NTRand sortilin. J Neurosci. 25:5455–5463. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Massa SM, Yang T, Xie Y, et al: Small
molecule BDNF mimetics activate TrkB signaling and prevent neuronal
degeneration in rodents. J Clin Invest. 120:1774–1785. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Numakawa T, Suzuki S, Kumamaru E, Adachi
N, Richards M and Kunugi H: BDNF function and intracellular
signaling in neurons. Histol Histopathol. 25:237–258.
2010.PubMed/NCBI
|
|
28
|
Han BH, D’Costa A, Back SA, et al: BDNF
blocks caspase-3 activation in neonatal hypoxia-ischemia. Neurobiol
Dis. 7:38–53. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Chen AI, Nguyen CN, Copenhagen DR, et al:
TrkB (tropomyosin-related kinase B) controls the assembly and
maintenance of GABAergic synapses in the cerebellar cortex. J
Neurosci. 31:2769–2780. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wong J, Woon HG and Weickert CS: Full
length TrkB potentiates estrogen receptor alpha mediated
transcription suggesting convergence of susceptibility pathways in
schizophrenia. Mol Cell Neurosci. 46:67–78. 2011. View Article : Google Scholar
|
|
31
|
Luberg K, Wong J, Weickert CS and Timmusk
T: Human TrkB gene: novel alternative transcripts, protein isoforms
and expression pattern in the prefrontal cerebral cortex during
postnatal development. J Neurochem. 113:952–964. 2010. View Article : Google Scholar
|
|
32
|
Zampieri N and Chao MV: Mechanisms of
neurotrophin receptor signalling. Biochem Soc Trans. 34:607–611.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Skaper SD, Floreani M, Negro A, Facci L
and Giusti P: Neurotrophins rescue cerebellar granule neurons from
oxidative stress-mediated apoptotic death: selective involvement of
phosphatidylinositol 3-kinase and themitogen-activated protein
kinase pathway. J Neurochem. 70:1859–1868. 1998. View Article : Google Scholar
|
|
34
|
Huang EJ and Reichardt LF: Trk receptors:
roles in neuronal signal transduction. Annu Rev Biochem.
72:609–642. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Lessmann V, Gottmann K and Malcangio M:
Neurotrophin secretion: current facts and future prospects. Prog
Neurobiol. 69:341–374. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Han BH and Holtzman DM: BDNF protects the
neonatal brain from hypoxic-ischemic injury in vivo via the ERK
pathway. J Neurosci. 20:5775–5781. 2000.PubMed/NCBI
|
|
37
|
Sun X, Zhou H, Luo X, et al:
Neuroprotection of brain-derived neurotrophic factor against
hypoxic injury in vitro requires activation of extracellular
signal-regulated kinase and phosphatidylinositol 3-kinase. Int J
Dev Neurosci. 26:363–370. 2008. View Article : Google Scholar
|
|
38
|
Amaral MD and Pozzo-Miller L: BDNF induces
calcium elevations associated with IBDNF, a nonselective cationic
current mediated by TRPC channels. J Neurophysiol. 98:2476–2482.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Angelo MF, Aviles-Reyes RX, Villarreal A,
Barker P, Reines AG and Ramos AJ: p75 NTR expression is induced in
isolated neurons of the penumbra after ischemia by cortical
devascularization. J Neurosci Res. 87:1892–1903. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Dechant G and Barde YA: The neurotrophin
receptor p75(NTR): novel functions and implications for diseases of
the nervous system. Nat Neurosci. 5:1131–1136. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Ho R, Minturn JE, Simpson AM, et al: The
effect of P75 on Trk receptors in neuroblastomas. Cancer Lett.
305:76–85. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Kenchappa RS, Tep C, Korade Z, et al: p75
neurotrophin receptor-mediated apoptosis in sympathetic neurons
involves abiphasic activation of JNK and up-regulation of tumor
necrosisfactor-alpha-converting enzyme/ADAM17. J Biol Chem.
285:20358–20368. 2010. View Article : Google Scholar
|
|
43
|
Schecterson LC and Bothwell M:
Neurotrophin receptors: old friends with new partners. Dev
Neurobiol. 70:332–338. 2010.PubMed/NCBI
|
|
44
|
Lou H, Kim SK, Zaitsev E, Snell CR, Lu B
and Loh YP: Sorting and activity-dependent secretion of BDNF
require interaction of aspecific motif with the sorting receptor
carboxypeptidase e. Neuron. 45:245–255. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Lou H, Park JJ, Cawley NX, et al:
Carboxypeptidase E cytoplasmic tail mediates localization of
synaptic vesicles to the pre-active zone in hypothalamic
pre-synaptic terminals. J Neurochem. 114:886–896. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Kuczewski N, Porcher C, Lessmann V, Medina
I and Gaiarsa JL: Activity-dependent dendritic release of BDNF and
biological consequences. Mol Neurobiol. 39:37–49. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Pallesen LT and Vaegter CB: Sortilin and
SorLA regulate neuronal sorting of trophic and dementia-linked
proteins. Mol Neurobiol. 45:379–387. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Evans SF, Irmady K, Ostrow K, et al:
Neuronal brain-derived neurotrophic factor is synthesized in
excess, with levels regulated by sortilin-mediated trafficking and
lysosomal degradation. J Biol Chem. 286:29556–29567. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Shah PS, Ohlsson A and Perlman M:
Hypothermia to treat neonatal hypoxic ischemic encephalopathy:
systematic review. Arch Pediatr Adolesc Med. 161:951–958. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Chalak LF, Rollins N, Morriss MC, Brion
LP, Heyne R and Sanchez PJ: Perinatal acidosis and hypoxic-ischemic
encephalopathy in preterm infants of 33 to 35 weeks’ gestation. J
Pediatr. 160:388–394. 2012.PubMed/NCBI
|
|
51
|
Szydlowska K and Tymianski M: Calcium,
ischemia and excitotoxicity. Cell Calcium. 47:122–129. 2010.
View Article : Google Scholar
|
|
52
|
Zhang J, Yu Z, Yu Z, et al: rAAV-mediated
delivery of brain-derived neurotrophic factor promotes neurite
outgrowth and protects neurodegeneration in focal ischemic model.
Int J Clin Exp Pathol. 4:496–504. 2011.
|
|
53
|
Liu Z, Ma D, Feng G, Ma Y and Hu H:
Recombinant AAV-mediated expression of human BDNF protects neurons
against cell apoptosis in Abeta-induced neuronal damage model. J
Huazhong Univ Sci Technolog Med Sci. 27:233–236. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Takeshima Y, Nakamura M, Miyake H, et al:
Neuroprotection with intraventricular brain-derived neurotrophic
factor in rat venous occlusion model. Neurosurgery. 68:1334–1341.
2011.PubMed/NCBI
|
|
55
|
Kiprianova I, Freiman TM, Desiderato S, et
al: Brain-derived neurotrophic factor prevents neuronal death and
glial activation after global ischemia in the rat. J Neurosci Res.
56:21–27. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Kurokawa T, Katai N, Shibuki H, et al:
BDNF diminishes caspase-2 but not c-Jun immunoreactivity of neurons
in retinal-ganglion cell layer after transient ischemia. Invest
Ophthalmol Vis Sci. 40:3006–3011. 1999.PubMed/NCBI
|
|
57
|
Neumann J, Gunzer M, Gutzeit HO, Ullrich
O, Reymann KG and Dinkel K: Microglia provide neuroprotection after
ischemia. FASEB J. 20:714–716. 2006.PubMed/NCBI
|
|
58
|
Lai AY and Todd KG: Differential
regulation of trophic and proinflammatory microglial effectors is
dependent on severity of neuronal injury. Glia. 56:259–270. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Zhang J, Geula C, Lu C, Koziel H, Hatcher
LM and Roisen FJ: Neurotrophins regulate proliferation and survival
of two microglial cell lines in vitro. Exp Neurol. 183:469–481.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Jiang Y, Wei N, Lu T, Zhu J, Xu G and Liu
X: Intranasal brain-derived neurotrophic factor protects brain from
ischemic insult via modulating local inflammation in rats.
Neuroscience. 172:398–405. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Nakajima K and Kohsaka S: Microglia:
neuroprotective and neurotrophic cells in the central nervous
system. Curr Drug Targets Cardiovasc Haematol Disord. 4:65–84.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
McCoy MK and Tansey MG: TNF signaling
inhibition in the CNS: implications for normal brain function and
neurodegenerative disease. J Neuroinflammation. 5:452008.
View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Frenkel D, Pachori AS, Zhang L, et al:
Nasal vaccination with troponin reduces troponin specific T-cell
responses and improves heart function in myocardial
ischemia-reperfusion injury. Int Immunol. 21:817–829. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Peng CH, Chiou SH, Chen SJ, et al:
Neuroprotection by Imipramine against lipopolysaccharide-induced
apoptosis in hippocampus-derived neural stem cells mediated by
activation of BDNF and the MAPK pathway. Eur Neuropsychopharmacol.
18:128–140. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Casalbore P, Barone I, Felsani A, et al:
Neural stem cells modified to express BDNF antagonize
trimethyltin-induced neurotoxicity through PI3K/Akt and MAP kinase
pathways. J Cell Physiol. 224:710–721. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Sakai R, Ukai W, Sohma H, et al:
Attenuation of brain derived neurotrophic factor (BDNF) by ethanol
and cytoprotective effect of exogenous BDNF against ethanol damage
in neuronal cells. J Neural Transm. 112:1005–1013. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Kume T, Kouchiyama H, Kaneko S, et al:
BDNF prevents NO mediated glutamate cytotoxicity in cultured
cortical neurons. Brain Res. 756:200–204. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Endres M, Dirnagl U and Moskowitz MA: The
ischemic cascade and mediators of ischemic injury. Handb Clin
Neurol. 92:31–41. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Akaike A, Katsuki H, Kume T and Maeda T:
Reactive oxygen species in NMDA receptor-mediated glutamate
neurotoxicity. Parkinsonism Relat Disord. 5:203–207. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Crozier RA, Bi C, Han YR and Plummer MR:
BDNF modulation of NMDA receptors is activity-dependent. J
Neurophysiol. 100:3264–3274. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Georgiev DD, Taniura H, Kambe Y and Yoneda
Y: Crosstalk between brain-derived neurotrophic factor and
N-methyl-D-aspartate receptor signaling in neurons. Biomed Rev.
19:17–27. 2008. View Article : Google Scholar
|
|
72
|
Zhu JM, Zhao YY, Chen SD, Zhang WH, Lou L
and Jin X: Functional recovery after transplantation of neural stem
cells modified by brain-derived neurotrophic factor in rats with
cerebral ischaemia. J Int Med Res. 39:488–498. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Horch HW, Kruttgen A, Portbury SD and Katz
LC: Destabilization of cortical dendrites and spines by BDNF.
Neuron. 23:353–364. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Schabitz WR, Berger C, Kollmar R, et al:
Effect of brain-derived neurotrophic factor treatment and forced
arm use on functional motor recovery after small cortical ischemia.
Stroke. 35:992–997. 2004. View Article : Google Scholar
|
|
75
|
Lopatina T, Kalinina N, Karagyaur M, et
al: Adipose-derived stem cells stimulate regeneration of peripheral
nerves: BDNF secreted by these cells promotes nerve healing and
axon growth de novo. PLoS One. 6:e178992011. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Johnston MV, Trescher WH, Ishida A and
Nakajima W: Neurobiology of hypoxic-ischemic injury in the
developing brain. Pediatr Res. 49:735–741. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Husson I, Rangon CM, Lelievre V, et al:
BDNF-induced white matter neuroprotection and stage-dependent
neuronal survival following a neonatal excitotoxic challenge. Cereb
Cortex. 15:250–261. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Binder DK: The role of BDNF in epilepsy
and other diseases of the mature nervous system. Adv Exp Med Biol.
548:34–56. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Binder DK, Croll SD, Gall CM and Scharfman
HE: BDNF and epilepsy: too much of a good thing. Trends Neurosci.
24:47–53. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Blurton-Jones M, Kitazawa M,
Martinez-Coria H, et al: Neural stem cells improve cognition via
BDNF in a transgenic model of Alzheimer disease. Proc Natl Acad Sci
USA. 106:13594–13599. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Kiprianova I, Sandkuhler J, Schwab S,
Hoyer S and Spranger M: Brain-derived neurotrophic factor improves
long-term potentiation and cognitive functions after transient
forebrain ischemia in the rat. Exp Neurol. 159:511–519. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Griesbach GS, Hovda DA and Gomez-Pinilla
F: Exercise-induced improvement in cognitive performance after
traumatic brain injury in rats is dependent on BDNF activation.
Brain Res. 1288:105–115. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Bekinschtein P, Cammarota M, Katche C, et
al: BDNF is essential to promote persistence of long-term memory
storage. Proc Natl Acad Sci USA. 105:2711–2716. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Kubo T, Nonomura T, Enokido Y and Hatanaka
H: Brain-derived neurotrophic factor (BDNF) can prevent apoptosis
of rat cerebellar granule neurons in culture. Brain Res Dev Brain
Res. 85:249–258. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Bhave SV, Ghoda L and Hoffman PL:
Brain-derived neurotrophic factor mediates the anti-apoptotic
effect of NMDA in cerebellar granule neurons: signal transduction
cascades and site of ethanol action. J Neurosci. 19:3277–3286.
1999.
|
|
86
|
Han Q, Li B, Feng H, et al: The promotion
of cerebral ischemia recovery in rats by laminin-binding BDNF.
Biomaterials. 32:5077–5085. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Zhou H, Mao M, Liu W, Li S and Wang H:
Expression of BDNF Receptor TrkBmRNA in hypoxia-induced fetal
cortical neurons. J West China Univ Med Sci. 33:573–576. 2002.
|
|
88
|
Luo XL, Mao M, Zhou H, Sun XM and Li SF:
Neuroprotective effect of BDNF on hypoxia for embryonic rat
cortical neurons in vitro. Sichuan Da Xue Xue Bao Yi Xue Ban.
37:373–377. 2006.(In Chinese).
|
|
89
|
Meng M, Zhiling W, Hui Z, Shengfu L, Dan Y
and Jiping H: Cellular levels of TrkB and MAPK in the
neuroprotective role of BDNF for embryonic rat cortical neurons
against hypoxia in vitro. Int J Dev Neurosci. 23:515–521. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Meng M, Dan Y, Jie Z, Hui Z and Zhiling W:
Effect of brain-derived neurotrophic factor on apoptosis of embryo
brain suffered from intrauterine hypoxic-ischemic injury. J Appl
Clin Pediatr. 19:1062–1064. 2004.
|
|
91
|
Ostrowski RP, Graupner G, Titova E, et al:
The hyperbaric oxygen preconditioning-induced brain protection is
mediated by a reduction of early apoptosis after transient global
cerebral ischemia. Neurobiol Dis. 29:1–13. 2008. View Article : Google Scholar
|
|
92
|
Park JH, Joo HS, Yoo KY, et al: Extract
from Terminalia chebula seeds protect against experimental
ischemic neuronal damage via maintaining SODs and BDNF levels.
Neurochem Res. 36:2043–2050. 2011.
|
|
93
|
Lee CH, Park JH, Yoo KY, et al: Pre- and
post-treatments with escitalopram protect against experimental
ischemic neuronal damage via regulation of BDNF expression and
oxidative stress. Exp Neurol. 229:450–459. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Liu W, Sun X, Zhou H, Luo X and Mao M:
Correlation of a protective effect of brain-derived neurotrophic
factor on hypoxic neurons to extracellular signal-regulated kinase.
J Clin Rehabil Tissue Eng Res. 12:3884–3888. 2008.
|
|
95
|
Alcala-Barraza SR, Lee MS, Hanson LR,
McDonald AA, Frey WH II and McLoon LK: Intranasal delivery of
neurotrophic factors BDNF, CNTF, EPO, and NT-4 to the CNS. J Drug
Target. 18:179–190. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Guan J, Tong W, Ding W, et al: Neuronal
regeneration and protection by collagen-binding BDNF in the rat
middle cerebral artery occlusion model. Biomaterials. 33:1386–1395.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Kim SH, Won SJ, Sohn S, et al:
Brain-derived neurotrophic factor can act as a pronecrotic factor
through transcriptional and translational activation of NADPH
oxidase. J Cell Biol. 159:821–831. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Hwang JJ, Choi SY and Koh JY: The role of
NADPH oxidase, neuronal nitric oxide synthase and poly(ADP ribose)
polymerase in oxidative neuronal death induced in cortical cultures
by brain-derived neurotrophic factor and neurotrophin-4/5. J
Neurochem. 82:894–902. 2002. View Article : Google Scholar
|
|
99
|
Kingwell K: Neurodegenerative disease:
BDNF copycats. Nat Rev Drug Discov. 9:4332010. View Article : Google Scholar : PubMed/NCBI
|