Introduction
Cardiac hypertrophy is an adaptive response to
stimulation to heart, eventually progressing to heart failure.
Features of cardiac hypertrophy include increased cardiomyocyte
size, protein synthesis, elevated fetal gene atrial natriuretic
peptide (ANP), brain natriuretic peptide (BNP), β-myosin heavy
chain (β-MHC) and fibronectin protein expression, and abnormal
sarcomeric organization. Cell signaling pathways involved in
cardiac hypertrophy include mitogen-activated protein kinase
(MAPK), adenosine monophosphate-activated protein kinase (AMPK),
transforming growth factor β (TGF-β)/smads, Ras/Rho, janus kinase
(JAK)/signal transducers and activators of transcription (STAT),
and calcinurin/nuclear factor of activated T-cells (NFAT) (1,2).
LKB1 is a signaling protein (3,4) that forms a
complex with sterile-20-related adaptor (STRAD) and mouse
protein-25 (MO25). STRAD and MO25 are required for the activity of
LKB1 (5,6). AMPK is a substrate of LKB1, and is
composed of AMPKα, AMPKβ and AMPKγ subunits. AMPKα is a catalytic
subunit, composed of AMPKα1 and AMPKα2. AMPKβ and AMPKγ are
regulatory subunits. AMPKα1 is widely expressed, whereas AMPKα2 is
primarily expressed in liver cells and skeletal and cardiac muscle
(7). The LKB1/AMPK pathway mediates
various biological functions, including proliferation, apoptosis,
autophagy and transcription (8,9).
Resistin is an adipocyte-secreted cytokine that is
linked to obesity, diabetes, insulin resistance, and cardiac
hypertrophy (10,11). Treatment of wild-type mice with
resistin causes glucose intolerance (12), and immunoneutralization of resistin in
obese mice decreases insulin sensitivity (10). Resistin can be regulated by a number of
cytokines, including endothelin (ET), insulin, insulin-like growth
factors (IGFs), and peroxisome proliferator-activated receptor
(PPAR). Furthermore, resistin has been reported to induce cardiac
hypertrophy through a number of signaling pathways, such as
extracellular signal-regulated kinases (ERK), AMPK/mammalian target
of rapamycin (mTOR), and the c-Jun NH (2)-terminal kinase (JNK)/insulin receptor
substrate 1 (IRS1) pathway (11,13). The
underlying molecular mechanisms by which resistin induces cardiac
hypertrophy are still not completely understood.
The aim of the present study was to investigate the
effects of resistin on LKB1/AMPK cell signaling and the induction
of cardiac hypertrophy in the H9c2 rat myoblast cell line.
Materials and methods
Reagents
Recombinant human resistin was purchased from
PeproTech, Inc. (Rocky Hill, NJ, USA). Metformin was ordered from
Sigma-Aldrich (St. Louis, MO, USA). The H9c2 rat cardiomyoblast
cells were obtained from the American Type Culture Collection
(Manassas, VA, USA). Fetal calf serum (FCS) was purchased from
Zhejiang Tianhang Biological Technology (Zhejiang, China).
Antibodies raised against phospho-LKB1, LKB, phospho-AMPK and AMPK
were purchased from Cell Signaling Technology, Inc. (Danvers, MA,
USA). The UNIQ-10 column TRIzol® kit was ordered from Sangon
Biotech Co., Ltd. (Shanghai, China). PrimeScript®RT Master Mix
Perfect Real Time and SYBR® Premix Ex Taq™ II were obtained from
Takara (Tokyo, Japan).
H9c2 cell culture
H9c2 rat cardiomyoblast cells were cultured in DMEM
containing 10% FCS, 1% penicillin and 1% streptomycin at a
temperature of 37°C with a 5% CO2 atmosphere. Once cells
had reached 70–80% confluence, they were passaged according to a
1:2 or 1:3 proportion. The medium was changed every 2 days. Cells
were seeded at a density of 1×105 into a 35 mm tissue
culture dish. Cells were cultured in serum-free medium overnight
and treated with resistin at a concentration of 50 ng/ml for the
indicated time.
Determination of cell surface
area
In total, 8×104 cells were seeded in a
35-mm dish. Cells were cultured with serum-free DMEM overnight and
treated with resistin for 48 h. The cell surface area was
determined via quantification of the total surface (NIH ImageJ
version 1.49 software, Bethesda, MD, USA). Five observation fields
were selected at random and 10 cells in each observation field were
selected for measurement of cell surface area (14).
Protein synthesis measurement
In total, 1×105 cells were seeded in a
35-mm dish. Cells were cultured with serum-free DMEM overnight and
treated with resistin for 48 h. Cells were digested with trypsin
and counted under a microscope. The cells were then collected and
lysed in 100 µl of RIPA buffer. Protein concentrations were
measured using the Bradford protein assay kit (Bio-Rad
Laboratories, Inc., Hercules, CA, USA). Cell protein synthesis was
determined by dividing the total amount of protein by the cell
number (15). The trypan blue
exclusion test of cell viability was used to determine the number
of viable cells present in a cell suspension.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Cells were collected and RNA was extracted using the
UNIQ-10 column TRIzol kit (Shanghai Sangon Biotech) and treated
with DNase. A total of 1 µg of RNA was reverse transcribed to cDNA
using the PrimeScript®RT Master Mix Perfect Real Time Kit (Takara),
according to the manufacturer's instructions. PCR amplification was
conducted with SYBR® Premix Ex Taq™ II kit (Takara) using the
Applied Biosystems® 7500 Fast Real-Time PCR System (Thermo Fisher
Scientific, Inc., Waltham, MA, USA). The PCR reaction conditions
were: 95°C for 30 sec, then 40 cycles of 95°C for 5 sec followed by
60°C for 31 sec. 18s was used as a reference gene. The ΔΔCq method
was used for relative quantification. The BNP, β-MHC and 18s
primers were designed and synthesized by Sangon Biotech Co., Ltd.
The nucleotide sequences of the primers were as follows: BNP
forward: 5′-GGAGCATTGAGTTGGCTCTC-3′, and reverse:
5′-CCAGCTCTCCGAAGTGTTTC-3′; β-MHC forward:
5′-CACCCGCGAGTACAACCTTC-3′, and reverse:
5′-CCCATACCCACCATCACACC-3′; 18s forward: 5′-CACCCGCGAGTACAACCTTC-3′
and reverse: 5′-CCCATACCCACCATCACACC-3′.
Western blot analysis
Once the cells has reached 80–90% confluence, they
were washed twice with phosphate-buffered saline, digested with
0.05% trypsin (Beyotime Biotechnology, Beijing, China) for 1 min
and centrifuged at 1,000 × g for 5 min. Cells were mixed with 100
µl of lysis buffer and incubated on ice for 20 min. Lysates were
centrifuged at 1,000 × g and protein concentration was measured
using the bicinchoninic acid assay. 5X Laemmli's buffer was added
to samples, which were then heated at 95°C for 5 min and then
separated by 10% sodium dodecyl sulfate-polyacrylamide gel
electrophoresis. The proteins were transferred onto polyvinylidene
fluoride membranes (EMD Millipore, Billerica, MA, USA). The
membranes were blocked with TBST buffer (20 mM Tris-HCl, 150 mM
NaCl and 0.1% Tween-20) containing 5% non-fat milk for 1 h at room
temperature. The membranes were incubated in TBST buffer containing
5% non-fat milk with the primary antibodies p-LKB1 (cat. no. 3482S;
monoclonal rabbit anti-rat; 1:1,000; Cell Signaling Technology,
Inc.), LKB1 (cat. no. 3047S, monoclonal rabbit anti-rat; 1:1,000;
Cell Signaling Technology, Inc.), p-AMPK (cat. no. 2531S;
polyclonal rabbit anti-rat; 1:1,000; Cell Signaling Technology,
Inc.), AMPK (cat. no. 5831S; monoclonal rabbit anti-rat; 1:1,000,
Cell Signaling Technology, Inc.), β-actin (cat. no. 4967S;
polyclonal rabbit anti-rat; 1:1,000; Cell Signaling Technology,
Inc.) at 4°C overnight. Following primary antibody incubation, the
membranes were incubated with anti-rabbit secondary antibodies
(cat. no. 111-035-003; polyclonal goat-anti rabbit; 1:10,000;
Jackson ImmunoResearch, Inc., West Grove, PA, USA) conjugated to
horseradish peroxidase at room temperature for 1 h. The protein
bands were visualised using an enhanced chemiluminescence kit
(ComWin Biotech, Beijing, China) and FluorChem™Q Quantitative
Western Blot Imaging System (Bio-Techne, Minneapolis, MN, USA). The
band intensities were measured with ImageJ software, and the ratio
of phosphorylated protein antibodies over corresponding total
protein antibodies was calculated.
Statistical analysis
All experiments data were expressed as means ± SD
and performed at least three times. The Student's t-test was used
for statistical analysis and the differences were considered
statistically significant if P<0.05.
Results
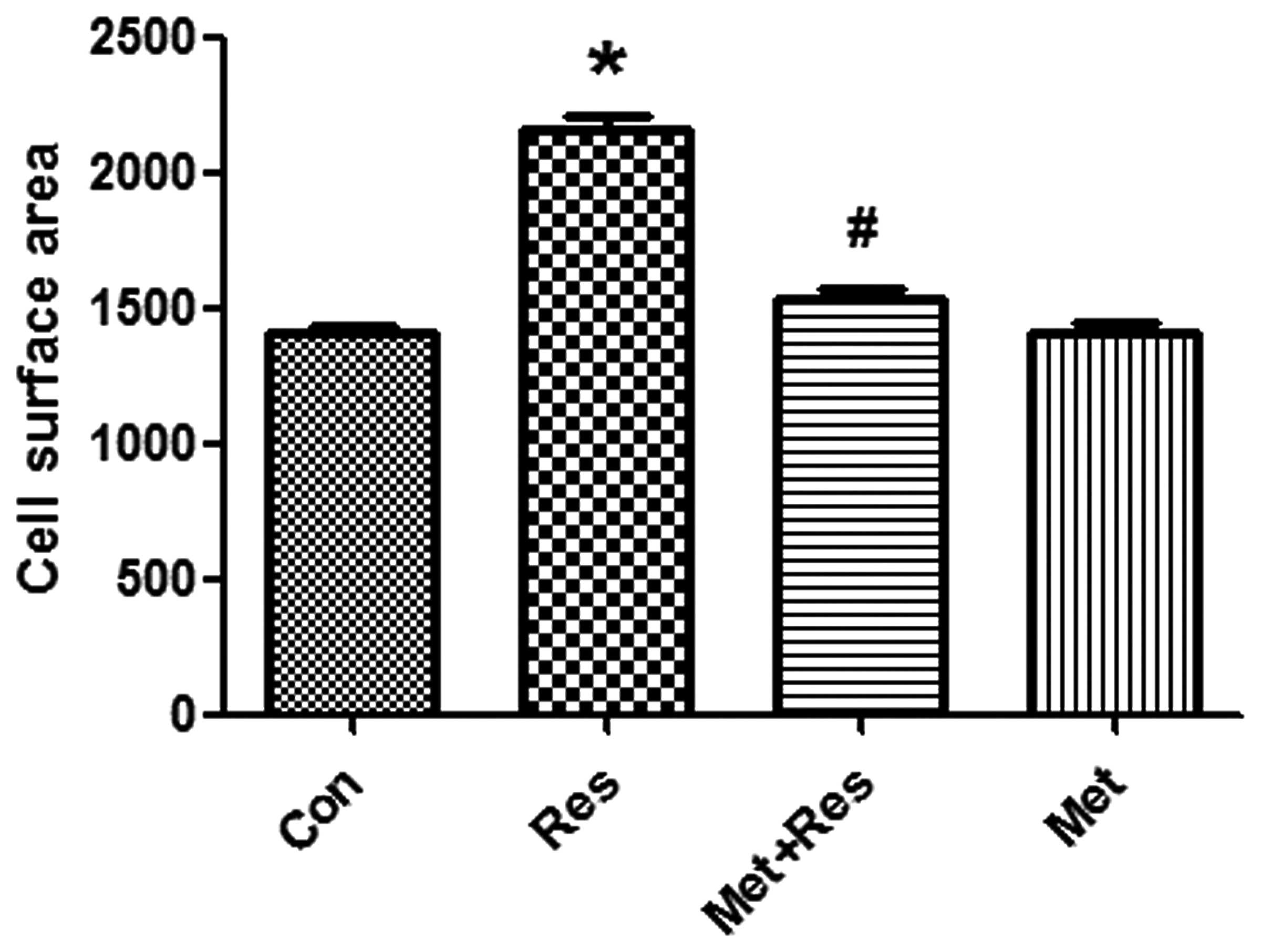
Resistin treatment increases H9c2 cell
size, which is reversed by metformin
Resistin treatment was used to induce cardiomyocyte
hypertrophy. H9c2 cells were treated with resistin at 50 mg/ml for
48 h. Resistin increased cell surface area significantly compared
to the control group (P<0.01; Fig.
1). Pre-treatment of cardiomyocytes with metformin led to a
significantly decreased resistin-induced cell surface area
(P<0.01, Fig. 1).
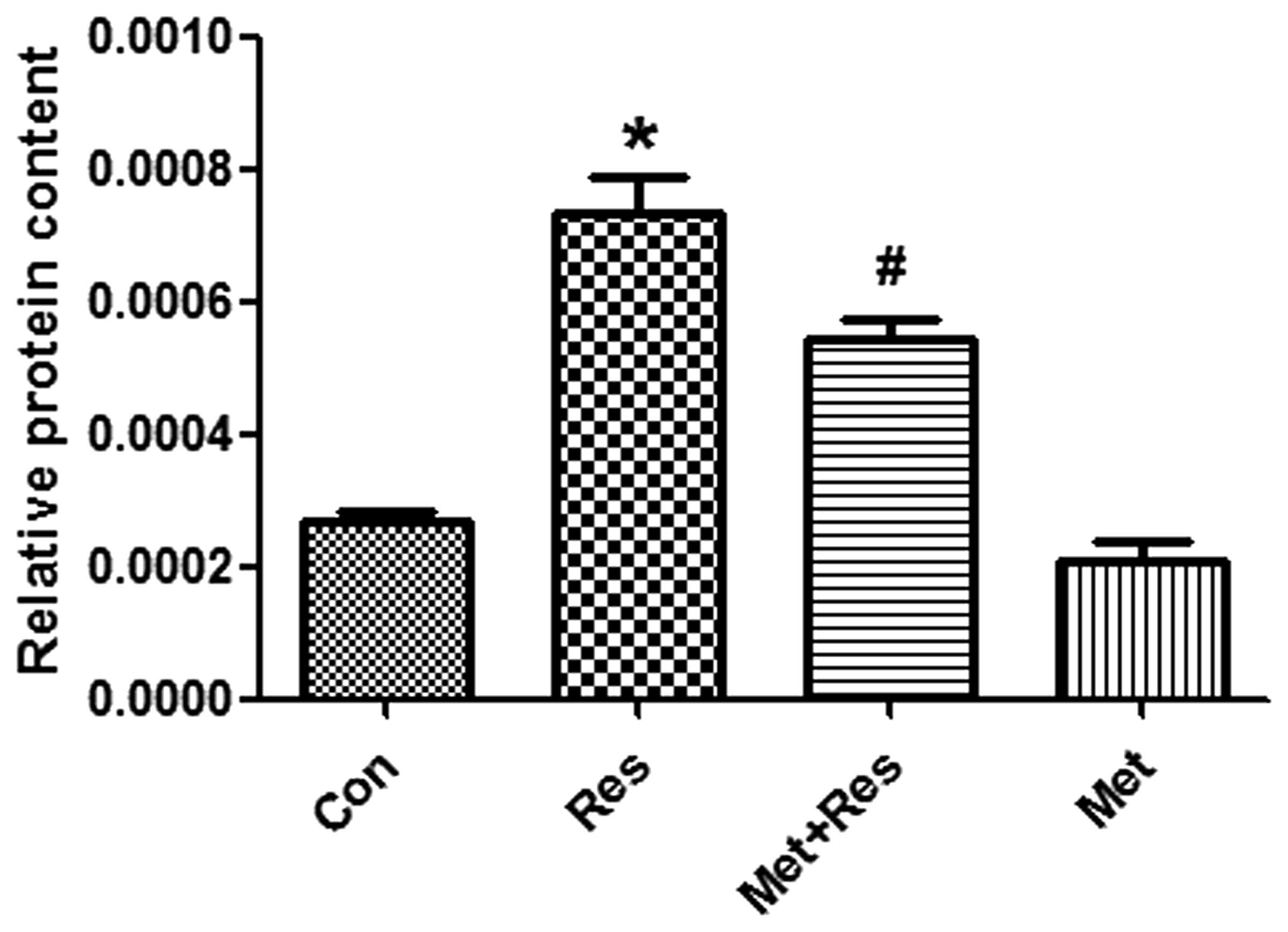
Resistin increases the synthesis
cardiomyocyte protein synthesis and opposes the effects of
metformin treatment
To investigate whether resistin treatment increases
protein synthesis in H9c2 cells and that this increase is inhibited
by metformin, cultured cardiomyocytes were exposed to resistin in
the presence and/or absence of metformin for 48 h. The results
demonstrate that resistin significantly increased protein synthesis
in cardiomyocyotes (P<0.05; Fig.
2). Furthermore, metformin treatment significantly decreased
the synthesis of proteins that were increased by resistin
(P<0.05; Fig. 2).
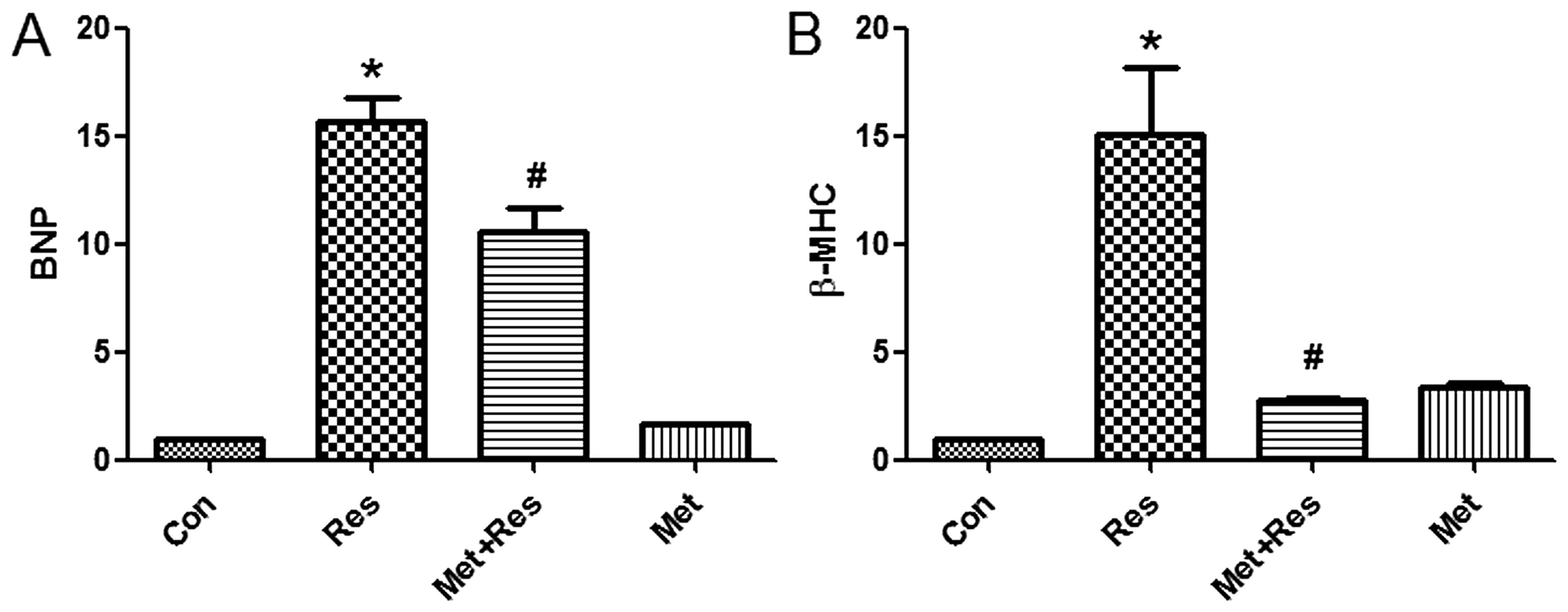
Resistin treatment elevates the mRNA
expression levels of BNP and β-MHC
As BNP and β-MHC are markers of cardiomyocyte
hypertrophy, the effects of resistin treatment on the expression of
BNP and β-MHC mRNA was investigated in H9c2 cells. The results
demonstrate that resistin treatment increased the expression of BNP
and β-MHC mRNA. Additionally, the results identified that metformin
treatment suppressed resistin-induced increase of BNP and β-MHC
mRNA expression (Fig. 3).
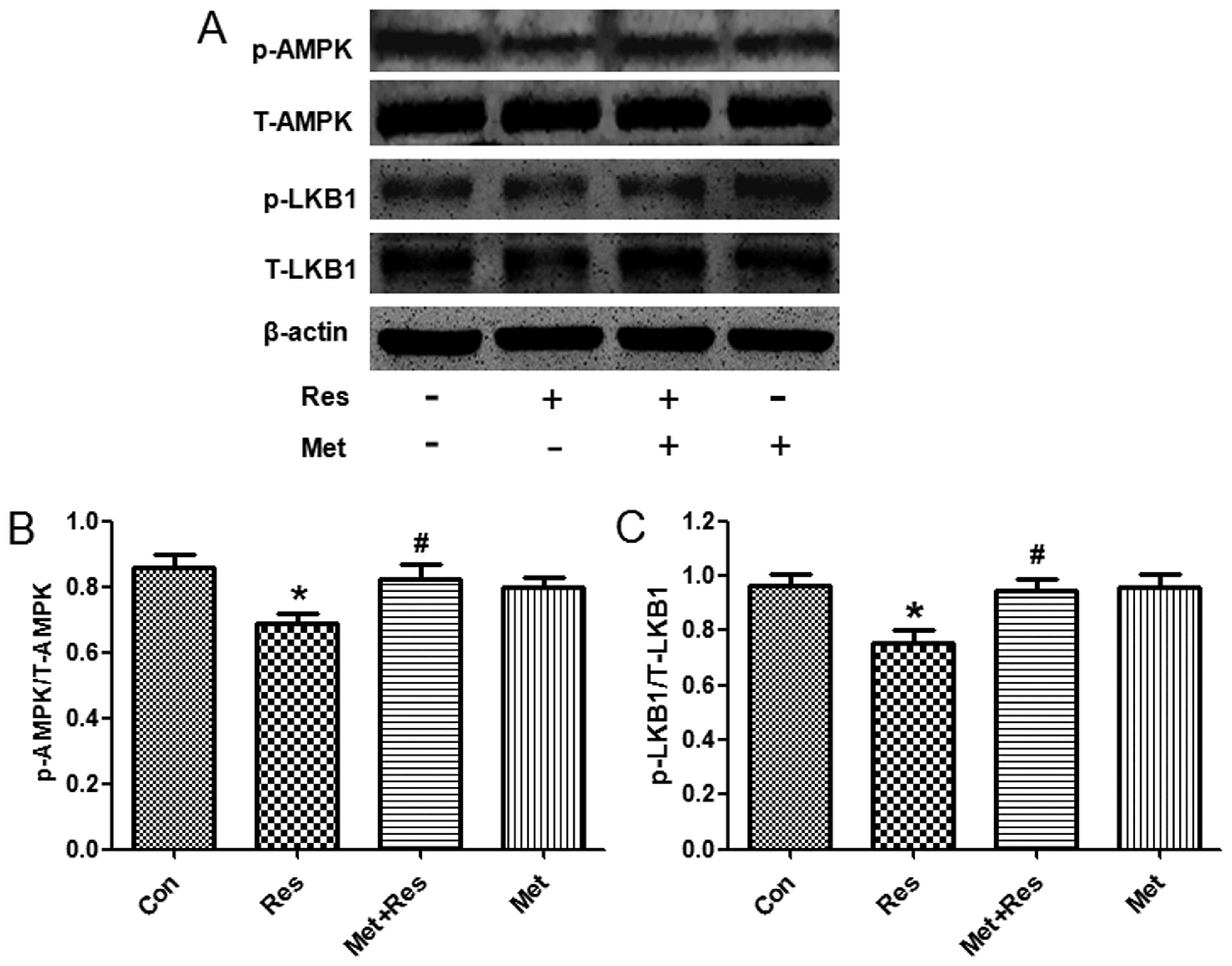
Resistin treatment reduces the
phosphorylation of LKB1 and AMPK
To further investigate the underlying molecular
mechanism by which resistin induces cardiac hypertrophy, we used
western blot analysis to evaluate the phosphorylation status of
LKB1 and AMPK following resistin treatment. Treatment of resistin
decreased phosphorylation of LKB1 and AMPK, whereas total LKB1 and
AMPK protein expression was unchanged. Since metformin is an
activator of LKB1 and AMPK (16,17),
treatment of metformin increased expression of phosphorylated LKB1
and AMPK that was decreased by resistin (Fig. 4).
Discussion
Increasing evidence indicates that resistin induces
cardiac hypertrophy. However, the underlying mechanisms by which
resistin induces cardiac hypertrophy remain largely unknown. In the
present study, resistin treatment increased cell size, protein
synthesis and hypertrophic marker BNP and β-MHC mRNA expression,
suggesting that resistin can induce cardiomyocyte hypertrophy.
Resistin-induced cardiac hypertrophy may be mediated by LKB1/AMPK
pathway.
Resistin is an adipocyte-secreted cytokine, released
from fat cells in rodents (10).
However, in humans, resistin is secreted from monocytes and
macrophages (18–21). The function of resistin is related to
obesity, diabetes and insulin resistance (10). Furthermore, treatment of resistin
impairs glucose tolerance and insulin action, whereas loss of
resistin function improves insulin resistance (22–24).
Overexpression of resistin induces cardiac hypertrophy in neonatal
rat cardiomyocytes through the activation of the oxidative stress
(25), IRS1/MAPK (11), AMPK/mTOR/p70S6K and Apoptosis
signal-regulating kinase 1 (ASK1)/JNK/IRS1 signaling pathways
(13).
Liver kinase 1 is a tumor suppressor gene widely
expressed in all tissues, and is involved in cell polarity, cell
motility, protein translation, energy metabolism and various signal
transduction pathways (26–29). LKB1 can phosphorylate downstream AMPK
at threonine 172 (30–32). Metformin, phenformin and AICAR are
activators of AMPK (4,33–35). The
activation of AMPK has antiproliferative activity. The mTOR pathway
is one of the downstream targets of AMPK, which is negatively
regulated by LKB1/AMPK signaling. Inhibition of mTOR activity leads
to inhibition of protein synthesis and proliferation (36). Furthermore, the LKB1-AMPK pathway plays
an important role in cardiac hypertrophy development. It has been
shown previously that 4-hydroxy-trans-2-nonenal (HNE) and miR-451
promote cardiac hypertrophy through suppression of the LKB1/AMPK
pathway (37,38). By contrast, resveratrol or NAD prevents
cardiac hypertrophy through enhancing the LKB1/AMPK signal
transduction pathway (31,39,40). The
current study demonstrates that resistin decreases phosphorylation
of LKB1, and subsequently decreases phosphorylation of AMPK.
Whereas metformin, an activator of LKB 1 and AMPK, increased
phosphorylation of LKB1 that is decreased by resistin. Similarly,
metformin increased expression of phosphorylated AMPK, is
suppressed by resistin. These results suggest that resistin
promotes cardiomyocyte hypertrophy via the LKB1/AMPK pathway.
In conclusion, resistin treatment elevates BNP and
β-MHC mRNA expression, cell surface area and protein synthesis, and
decreases the levels of LKB1 and AMPK phosphorylation, suggesting
that resistin induces cardiac hypertrophy through inactivation of
LKB1/AMPK signaling pathway. These results suggest that prevention
of resistin may be useful in the treatment of cardiac
hypertrophy.
References
|
1
|
Frey N, Katus HA, Olson EN and Hill JA:
Hypertrophy of the heart: A new therapeutic target? Circulation.
109:1580–1589. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Carreño JE, Apablaza F, Ocaranza MP and
Jalil JE: Cardiac hypertrophy: Molecular and cellular events. Rev
Esp Cardiol. 59:473–486. 2006.(In Spanish). View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Alessi DR, Sakamoto K and Bayascas JR:
LKB1-dependent signaling pathways. Annu Rev Biochem. 75:137–163.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Shaw RJ, Kosmatka M, Bardeesy N, Hurley
RL, Witters LA, DePinho RA and Cantley LC: The tumor suppressor
LKB1 kinase directly activates AMP-activated kinase and regulates
apoptosis in response to energy stress. Proc Natl Acad Sci USA.
101:3329–3335. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hawley SA, Boudeau J, Reid JL, Mustard KJ,
Udd L, Mäkelä TP, Alessi DR and Hardie DG: Complexes between the
LKB1 tumor suppressor, STRAD alpha/beta and MO25 alpha/beta are
upstream kinases in the AMP-activated protein kinase cascade. J
Biol. 2:282003. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Baas AF, Boudeau J, Sapkota GP, Smit L,
Medema R, Morrice NA, Alessi DR and Clevers HC: Activation of the
tumour suppressor kinase LKB1 by the STE20-like pseudokinase STRAD.
EMBO J. 22:3062–3072. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Stapleton D, Mitchelhill KI, Gao G, Widmer
J, Michell BJ, Teh T, House CM, Fernandez CS, Cox T, Witters LA and
Kemp BE: Mammalian AMP-activated protein kinase subfamily. J Biol
Chem. 271:611–614. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Alexander A and Walker CL: The role of
LKB1 and AMPK in cellular responses to stress and damage. FEBS
Lett. 585:952–957. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zaha VG and Young LH: AMP-activated
protein kinase regulation and biological actions in the heart. Circ
Res. 111:800–814. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Steppan CM, Bailey ST, Bhat S, Brown EJ,
Banerjee RR, Wright CM, Patel HR, Ahima RS and Lazar MA: The
hormone resistin links obesity to diabetes. Nature. 409:307–312.
2001. View
Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kim M, Oh JK, Sakata S, Liang I, Park W,
Hajjar RJ and Lebeche D: Role of resistin in cardiac contractility
and hypertrophy. J Mol Cell Cardiol. 45:270–280. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Graveleau C, Zaha VG, Mohajer A, Banerjee
RR, Dudley-Rucker N, Steppan CM, Rajala MW, Scherer PE, Ahima RS,
Lazar MA and Abel ED: Mouse and human resistins impair glucose
transport in primary mouse cardiomyocytes and oligomerization is
required for this biological action. J Biol Chem. 280:31679–31685.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kang S, Chemaly ER, Hajjar RJ and Lebeche
D: Resistin promotes cardiac hypertrophy via the AMP-activated
protein kinase/mammalian target of rapamycin (AMPK/mTOR) and c-Jun
N-terminal kinase/insulin receptor substrate 1 (JNK/IRS1) pathways.
J Biol Chem. 286:18465–18473. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Sun B, Huo R, Sheng Y, Li Y, Xie X, Chen
C, Liu HB, Li N, Li CB, Guo WT, et al: Bone morphogenetic protein-4
mediates cardiac hypertrophy, apoptosis and fibrosis in
experimentally pathological cardiac hypertrophy. Hypertension.
61:352–360. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Merten KE, Jiang Y and Kang YJ: Zinc
inhibits doxorubicin-activated calcineurin signal transduction
pathway in H9c2 embryonic rat cardiac cells. Exp Biol Med
(Maywood). 232:682–689. 2007.PubMed/NCBI
|
|
16
|
Shaw RJ, Lamia KA, Vasquez D, Koo SH,
Bardeesy N, Depinho RA, Montminy M and Cantley LC: The kinase LKB1
mediates glucose homeostasis in liver and therapeutic effects of
metformin. Science. 310:1642–1646. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhou G, Myers R, Li Y, Chen Y, Shen X,
Fenyk-Melody J, Wu M, Ventre J, Doebber T, Fujii N, et al: Role of
AMP-activated protein kinase in mechanism of metformin action. J
Clin Invest. 108:1167–1174. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
18
|
Patel L, Buckels AC, Kinghorn IJ, Murdock
PR, Holbrook JD, Plumpton C, Macphee CH and Smith SA: Resistin is
expressed in human macrophages and directly regulated by PPAR gamma
activators. Biochem Biophys Res Commun. 300:472–476. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lu SC, Shieh WY, Chen CY, Hsu SC and Chen
HL: Lipopolysaccharide increases resistin gene expression in vivo
and in vitro. FEBS Lett. 530:158–162. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kaser S, Kaser A, Sandhofer A, Ebenbichler
CF, Tilg H and Patsch JR: Resistin messenger-RNA expression is
increased by proinflammatory cytokines in vitro. Biochem Biophys
Res Commun. 309:286–290. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lehrke M, Reilly MP, Millington SC, Iqbal
N, Rader DJ and Lazar MA: An inflammatory cascade leading to
hyperresistinemia in humans. PLoS Med. 1:e452004. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Banerjee RR, Rangwala SM, Shapiro JS, Rich
AS, Rhoades B, Qi Y, Wang J, Rajala MW, Pocai A, Scherer PE, et al:
Regulation of fasted blood glucose by resistin. Science.
303:1195–1198. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Muse ED, Obici S, Bhanot S, Monia BP,
McKay RA, Rajala MW, Scherer PE and Rossetti L: Role of resistin in
diet-induced hepatic insulin resistance. J Clin Invest.
114:232–239. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Rajala MW, Obici S, Scherer PE and
Rossetti L: Adipose-derived resistin and gut-derived resistin-like
molecule-beta selectively impair insulin action on glucose
production. J Clin Invest. 111:225–230. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chemaly ER, Hadri L, Zhang S, Kim M,
Kohlbrenner E, Sheng J, Liang L, Chen J, K-Raman P, Hajjar RJ and
Lebeche D: Long-term in vivo resistin overexpression induces
myocardial dysfunction and remodeling in rats. J Mol Cell Cardiol.
51:144–155. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Sanchez-Cespedes M, Parrella P, Esteller
M, Nomoto S, Trink B, Engles JM, Westra WH, Herman JG and Sidransky
D: Inactivation of LKB1/STK11 is a common event in adenocarcinomas
of the lung. Cancer Res. 62:3659–3662. 2002.PubMed/NCBI
|
|
27
|
Guldberg P, Straten Thor P, Ahrenkiel V,
Seremet T, Kirkin AF and Zeuthen J: Somatic mutation of the
Peutz-Jeghers syndrome gene, LKB1/STK11, in malignant melanoma.
Oncogene. 18:1777–1780. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
McCabe MT, Powell DR, Zhou W and Vertino
PM: Homozygous deletion of the STK11/LKB1 locus and the generation
of novel fusion transcripts in cervical cancer cells. Cancer Genet
Cytogenet. 197:130–141. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zheng B, Jeong JH, Asara JM, Yuan YY,
Granter SR, Chin L and Cantley LC: Oncogenic B-RAF negatively
regulates the tumor suppressor LKB1 to promote melanoma cell
proliferation. Mol Cell. 33:237–247. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Oakhill JS, Chen ZP, Scott JW, Steel R,
Castelli LA, Ling N, Macaulay SL and Kemp BE: β-Subunit
myristoylation is the gatekeeper for initiating metabolic stress
sensing by AMP-activated protein kinase (AMPK). Proc Natl Acad Sci
USA. 107:19237–19241. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Dolinsky VW, Chan AY, Frayne Robillard I,
Light PE, Des Rosiers C and Dyck JR: Resveratrol prevents the
prohypertrophic effects of oxidative stress on LKB1. Circulation.
119:1643–1652. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Calamaras TD, Lee C, Lan F, Ido Y, Siwik
DA and Colucci WS: Post-translational modification of
serine/threonine kinase LKB1 via adduction of the reactive lipid
species 4-Hydroxy-trans-2-nonenal (HNE) at lysine residue 97
directly inhibits kinase activity. J Biol Chem. 287:42400–42406.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
El-Mir MY, Nogueira V, Fontaine E, Avéret
N, Rigoulet M and Leverve X: Dimethylbiguanide inhibits cell
respiration via an indirect effect targeted on the respiratory
chain complex I. J Biol Chem. 275:223–228. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Owen MR, Doran E and Halestrap AP:
Evidence that metformin exerts its anti-diabetic effects through
inhibition of complex 1 of the mitochondrial respiratory chain.
Biochem J. 348:607–614. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Shackelford DB, Abt E, Gerken L, Vasquez
DS, Seki A, Leblanc M, Wei L, Fishbein MC, Czernin J, Mischel PS
and Shaw RJ: LKB1 inactivation dictates therapeutic response of
non-small cell lung cancer to the metabolism drug phenformin.
Cancer Cell. 23:143–158. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zhou W, Marcus AI and Vertino PM:
Dysregulation of mTOR activity through LKB1 inactivation. Chin J
Cancer. 32:427–433. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Calamaras TD, Lee C, Lan F, Ido Y, Siwik
DA and Colucci WS: The lipid peroxidation product
4-hydroxy-trans-2-nonenal causes protein synthesis in cardiac
myocytes via activated mTORC1-p70S6K-RPS6 signaling. Free Radic
Biol Med. 82:137–146. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Kuwabara Y, Horie T, Baba O, Watanabe S,
Nishiga M, Usami S, Izuhara M, Nakao T, Nishino T, Otsu K, et al:
MicroRNA-451 exacerbates lipotoxicity in cardiac myocytes and
high-fat diet-induced cardiac hypertrophy in mice through
suppression of the LKB1/AMPK pathway. Circ Res. 116:279–288. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Dolinsky VW, Chakrabarti S, Pereira TJ,
Oka T, Levasseur J, Beker D, Zordoky BN, Morton JS, Nagendran J,
Lopaschuk GD, et al: Resveratrol prevents hypertension and cardiac
hypertrophy in hypertensive rats and mice. Biochim Biophys Acta.
1832:1723–1733. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Pillai VB, Sundaresan NR, Kim G, Gupta M,
Rajamohan SB, Pillai JB, Samant S, Ravindra PV, Isbatan A and Gupta
MP: Exogenous NAD blocks cardiac hypertrophic response via
activation of the SIRT3-LKB1-AMP-activated kinase pathway. J Biol
Chem. 285:3133–3144. 2010. View Article : Google Scholar : PubMed/NCBI
|