Introduction
Stroke is among the most common causes of mortality
and disability worldwide (1), and is
defined as loss of blood supply to the brain by thrombotic, embolic
or hemorrhagic events (2). The main
consequences include local hypoxia, mitochondria dysfunction and
increases in intracellular calcium, all of which lead to neuronal
death and subsequent tissue damage due to oxygen and glucose
deprivation (3). According to a
report by the American Heart Association in 2015, up to 85% of
strokes are ischemic, and 12% are hemorrhagic (4).
Recently, neuropsychiatric disorders resulting from
cerebrovascular diseases have gained increasing attention. An
epidemiological study across a diverse population suggested that
post-stroke depression (PSD) was among the common complications of
stroke, with an incidence rate of more than 30% within 5 years of
stroke occurrence (5). In addition,
PSD has been reported to be accompanied by sleep disorders,
cognitive impairments, and may also be a predictor of poor
recovery, high mortality and morbidity, and a cause of considerable
health care costs (6-8). The
underlying mechanism of PSD is not clearly understood, although
several studies have suggested that it is associated with altered
neurotrophic signaling, dysregulation of the
hypothalamus-pituitary-adrenal axis, neuroinflammation and
biological amine neurotransmitter disorders (1,8,9). Drutel et al (10) reported that aryl hydrocarbon receptor
(AHR) nuclear translocator protein 2 (ARNT2) may serve a vital role
in neuron survival and cell proliferation by forming heterodimers
with hypoxia-inducible factor (HIF)-1α, a factor involved in
adaptation to hypoxia.
ARNT2 is established as a member of the basic
helix-loop-helix (bHLH) superfamily of transcription factors, and
shares an amino acid sequence of almost 90% similarity with ARNT
(11). It is primarily expressed in
the central nervous system and developing kidney (12), forming a dimer with other basic
helix-loop-helix Per-Arnt-Sim homology (bHLH-PAS) transcription
factors including AHR and HIF-1α when responding to xenobiotics and
hypoxia, respectively (13). It has
been suggested that under hypoxic conditions, HIF-1α alternatively
binds with ARNT or ARNT2, also designated as HIF-1β and HIF-2β,
respectively, and subsequently activates the hypoxic response
element, leading to the expression of target genes involved in
glycolysis, erythropoiesis and angiogenesis (14,15).
Interestingly, a study involving a population from a high altitude
area of Ethiopia demonstrated an association between ARNT2 and
hemoglobin levels (16), which aided
to further clarify the key role of this protein in the hypoxia
response in humans.
Magnetic resonance images have shown that
hippocampal volume is reduced in patients with depression (17,18).
Depression following stroke events is associated with cognitive
dysfunction, which may be indirectly associated with the damage to
the hippocampus (19-21).
In hypoxic environments, neurons, particularly CA1 pyramidal
neurons, are susceptible and vulnerable to post-ischemia cell death
(22). It has also been reported that
the hippocampal CA3 region regulates learning and memory abilities,
particularly the normal operations of learning and memory processes
(23). Although Valen et al
(24) reported that ARNT2 was highly
and preferentially expressed in the hippocampus, and previous
research indicated that ARNT2 was also potently expressed in
ischemic brain tissue in rats (16),
less is known about its expression levels in PSD rats. Therefore,
the present study aimed to investigate the levels of ARNT2 in the
hippocampus of PSD rats, as well as the potential association
between ARNT2 levels and cognitive behavioral function.
Materials and methods
Animals and groups
Male Sprague-Dawley rats (n=32) aged 8 weeks,
weighing 160±20 g, were purchased from the Experimental Animal
Center of Zhengzhou University, Zhengzhou, China. Rats were housed
at 22±2˚C with 57±2% relative humidity under a 12-h light/dark
cycle, and were given food and water ad libitum except
during specific times of the experiment. All experiments were
conducted in accordance with the National Research Council: Guide
for the Care and Use of Laboratory Animals (8th edition), and the
rat experimental procedures were approved by the Laboratory Animal
Management Commission of the Henan Key Laboratory of Biological
Psychiatry, Xinxiang, China.
Rats were allowed to adapt to the environment for 1
week before baseline body weight (BW) and sucrose consumption were
measured, and open field tests (OFTs) were performed to ensure the
homogeneity of rats. Rats were then randomly divided into four
groups (n=8/group): A control group; a chronic unpredictable mild
stress (CUMS) group; a middle cerebral artery occlusion (MCAO)
group; and an MCAO+CUMS group. All rats in the MCAO and MCAO+CUMS
groups underwent MCAO surgery, while the rats in the CUMS group
were treated with a 4-week CUMS regiment. Finally, rats in the
MCAO+CUMS group were exposed to the same 4-week CUMS regiment
following surgery.
MCAO
Rats were fasted overnight prior to the day of the
experiment but were allowed free access to tap water. MCAO was
conducted as previously described (25). Briefly, anesthesia was induced with
10% chloral hydrate [350 mg/kg BW, intraperitoneally (i.p.); Dalian
Meilun Biological Technology Co., Ltd., Dalian, China] and a
ventral side incision was made to expose the left carotid
bifurcation, the left common carotid artery (CCA), the left
internal carotid artery (ICA) and the left external carotid artery
(ECA). The left ECA and proximal CCA were ligated with a 4-0 silk
suture, while the left ICA was occluded by a microvascular clip; a
small incision was made in the CCA proximal from the left carotid
bifurcation. The microvascular clip was then removed and a nylon
thread was inserted through the left CCA and left ECA to occlude
the middle cerebral artery. Rats in the control and CUMS groups
underwent the same surgery as above excluding threading of the
nylon through the left CCA and left ECA. Following surgery, rats
were selected according to Longa score, as previously described
(25).
CUMS procedure
Following recovery for 1 week, rats in the CUMS
groups that underwent surgery were subjected to a CUMS procedure
adapted from Willner (26) with minor
modifications. A total of seven different stressors were used:
Water/food deprivation for 24 h; behavioral restriction for 2 h;
wet cage for 24 h; electric foot shocks (average 60 V, 10 sec
duration, average 1 shock/5 sec); forced ice water swimming (4˚C,
5-15 min); and tail pinch for 1-2 min (27,28). Rats
in the CUMS and MCAO+CUMS groups were housed in separate cages.
Stressors were individually administered and separated from one
another by a 1-2 day period over a total of 28 days to account for
adaptation of rats to the CUMS regimen.
Behavioral tests BW
The BW of rats was measured prior to surgery and on
days 1, 7 and 28 thereafter, and was obtained only during normal
eating regiments (29).
Sucrose preference test
The animals were allowed to consume water and 1%
sucrose solution for 12 h following 24 h food and water deprivation
on the day prior to surgery and on days 7 and 28 thereafter. All
other components of the test were performed as previously described
(21,25).
OFT
The device for this test consisted of an open-field
response box (100x100x50 cm with opaque black metal walls; RWD Life
Science, Shenzhen, China) and an automatic data acquisition and
processing system (Panlab Smart version 2.5.16; Panlab, Barcelona,
Spain). Behavior, in terms of horizontal and vertical movements,
was determined in the open-field response box over 5 min to measure
spontaneous activity, independent ability to explore and cognitive
function (30).
Immunohistochemistry
Rats were anesthetized with 10% chloral hydrate (350
mg/kg, i.p.) and perfused with 0.9% saline followed by 4%
paraformaldehyde. The samples of 4 rats per group were used for
immunohistochemistry. Brains were removed and dehydrated, after
which brain tissue was embedded in Tissue-Tek® O.C.T.
Compound (Sakura Finetek USA, Inc., Torrance, CA, USA). Frozen
tissue was then sectioned at 15-µm thickness on a microtome (Leica
CM1850; Leica Microsystems GmbH, Wetzlar, Germany), and
immunohistochemistry was performed using a rabbit SP-9001 detection
kit (OriGene Technologies, Inc., Beijing, China) according to the
manufacturer's instructions. Briefly, sections were blocked with 3%
H2O2 in deionized water at room temperature
for endogenous peroxidase ablation, rinsed in phoshphate buffered
saline (PBS) three times and blocked in blocking buffer (normal
goat serum for 10-15 min at room temperature). Following removal of
blocking buffer, sections were incubated with primary antibody in
PBS for 1 h at 37˚C, washed three times with PBS, and incubated
with biotinylated secondary antibody (immunoglobulin G,
anti-rabbit) for 10-15 min at room temperature. Sections were then
incubated with horseradish enzyme-labeled streptavidin for a
further 10-15 min at 37˚C. ARNT-positive staining in the CA1 and
CA2 hippocampal fields was observed under an optical microscope
(Leica DM2000; Leica Microsystems GmbH). All data were analyzed
with Image-Pro plus 6.0 (Media Cybernetics, Inc., Rockville, MD,
USA).
Reverse transcription-polymerase chain
reaction (RT-PCR)
All tissue preparation steps for the isolation of
mRNA was performed as previously described (25). The samples not subject to
immunohistochemistry were used (n=4 rats per group). Primer pairs
for GAPDH and ARNT2 were designed based on sequences
in the PubMed database (https://www.ncbi.nlm.nih.gov/pubmed/). The primers for
ARNT2 were forward, 5'-ACCAGCGAGACGGGCTGTCA-3' and reverse,
5'-GTGCCCGGCAGGGAATGGAC-3', and for GAPDH were forward,
5'-GGGCTCTCTGCTCCTCCCTCT-3' and reverse,
5'-CCGTTGAACTTGCCGTGGGT-3'. The cycling parameters for PCR were as
follows: An initial cycle (pre-denaturation step) at 94˚C for 3
min, followed by 35 cycles at 94˚C for 15 sec, 68˚C for 30 sec and
72˚C for 30 sec and 72˚C for 10 min. Band intensities were measured
and analyzed using an GAS7001B gel image analysis software (UVIband
10.02; UVItec, Cambridge, UK). The expression of target mRNA was
measured based on its quantity relative to that of GADPH (relative
quality=target gene optical density/GAPDH optical density).
Statistical analysis
Data were presented as the mean ± standard
error of the mean and were analyzed using SPSS 13.0 software (SPSS
Inc., Chicago, IL, USA). All data were analyzed by one-way analysis
of variance (ANOVA) followed by Fisher's least significant
difference post hoc tests. Pearson's correlation analysis was
performed to analyse the relationship between target gene
expression and behavioral performance. A two-tailed P-value of
<0.05 was considered to indicate statistical significance.
Results
Behavioral assessment BW
Rats in the MCAO and MCAO+CUMS groups gained less
weight than controls at 7 and 28 days after surgery (P<0.01).
After 28 days of the CUMS regimen, rats in the CUMS group exhibited
significantly lower BW compared with the controls (P<0.01),
while rats in the MCAO+CUMS group gained significantly less weight
compared with those in the MCAO group (P<0.01; Table I).
 | Table IResults of weight changes in
rats. |
Table I
Results of weight changes in
rats.
| | BW, g |
|---|
| | | Postoperative
day |
|---|
| Group | Baseline | 7 | 28 |
|---|
| Control | 278.38±9.89 | 299.19±9.15 | 360.69±16.39 |
| MCAO | 277.50±21.93 |
194.31±27.13a |
295.44±41.45a |
| CUMS | 273.00±21.66 | 279.75±16.66 |
273.25±20.98a |
| MCAO+CUMS | 279.75±31.57 |
208.31±21.50a |
249.56±38.44a,b |
| F-value | 0.134 | 55.126 | 18.718 |
| P-value
(ANOVA) | 0.939 | <0.001 | <0.001 |
Sucrose preference
On day 7 after surgery, sucrose preference in the
MCAO and MCAO+CUMS groups was decreased compared with in the
control group, albeit non-significantly (P>0.05); likewise the
sucrose preference index of the CUMS group did not differ
significantly compared with the control group (P>0.05). After 28
days of the CUMS regimen, the CUMS and MCAO+CUMS groups exhibited
significantly decreased sucrose preference indexes compared with
the control group (P<0.01), although no significant difference
was determined between the MCAO and control groups (P>0.05).
Meanwhile, the CUMS and MCAO+CUMS groups displayed significantly
decreased sucrose preference indexes compared with the MCAO group
(P<0.01; Table II).
| Table IISucrose preference index. |
Table II
Sucrose preference index.
| | Sucrose preference
index |
|---|
| | | Postoperative
day |
|---|
| Group | Baseline | 7 | 28 |
|---|
| Control | 0.854±0.023 | 0.735±0.010 | 0.668±0.014 |
| MCAO | 0.890±0.014 | 0.708±0.017 | 0.633±0.007 |
| CUMS | 0.873±0.011 | 0.710±0.020 |
0.457±0.015a,b |
| MCAO+CUMS | 0.874±0.020 | 0.698±0.025 |
0.406±0.025a,b |
| F-value | 0.660 | 0.683 | 60.314 |
| P-value
(ANOVA) | 0.584 | 0.570 | <0.001 |
OFT
The OFT assessed horizontal motion distance traveled
and number of rearings of each rat. On day 7, rats in the MCAO and
MCAO+CUMS groups showed significantly less horizontal movement
compared with controls (P<0.01), although rats in the CUMS group
did not (P>0.05; Table III).
Similarly, rats in the MCAO and MCAO+CUMS groups performed a
significanlty lower number of rearings compared with the control
group (P<0.01), while rats in the CUMS group exhibited no
significant difference compared with the controls (P>0.05;
Table IV). After 4 weeks of the CUMS
regiment, rats in the CUMS and MCAO+CUMS groups exhibited
significantly less horizontal movement (P<0.05) and a lower
number of rearings (P<0.01) compared with controls (Tables III and IV).
| Table IIIHorizontal movement in the OFT. |
Table III
Horizontal movement in the OFT.
| | Movement, cm/5
min |
|---|
| | | Postoperative
day |
|---|
| Groups | Baseline | 7 | 28 |
|---|
| Control |
10,526.62±746.72 |
10,798.12±1,155.86 |
10,674.85±1,914.86 |
| MCAO |
9,855.28±2,037.12 |
2,578.43±718.61a |
10,980.34±939.91 |
| CUMS |
10,621.12±2,139.89 |
10,543.91±1,975.83 |
6,630.28±4,449.65b |
| MCAO+CUMS |
10,432.58±1,332.78 |
2,504.37±1,220.60a |
6,385.03±3,738.11b |
| F-value | 0.343 | 97.340 | 5.216 |
| P-value
(ANOVA) | 0.794 | <0.001 | 0.005 |
| Table IVNumber of rearings in the OFT. |
Table IV
Number of rearings in the OFT.
| | No. of rearings/5
min |
|---|
| | | Postoperative
day |
|---|
| Groups | Baseline | 7 | 28 |
|---|
| Control | 20.00±4.57 | 20.50±2.33 | 20.38±8.03 |
| MCAO | 20.75±5.04 |
7.50±4.21a | 20.50±3.85 |
| CUMS | 20.25±6.67 | 21.00±3.70 |
10.13±10.23a |
| MCAO+CUMS | 19.63±2.39 |
7.13±5.77a |
8.00±4.87a |
| F-value | 0.074 | 27.496 | 6.757 |
| P-value
(ANOVA) | 0.974 | <0.001 | 0.001 |
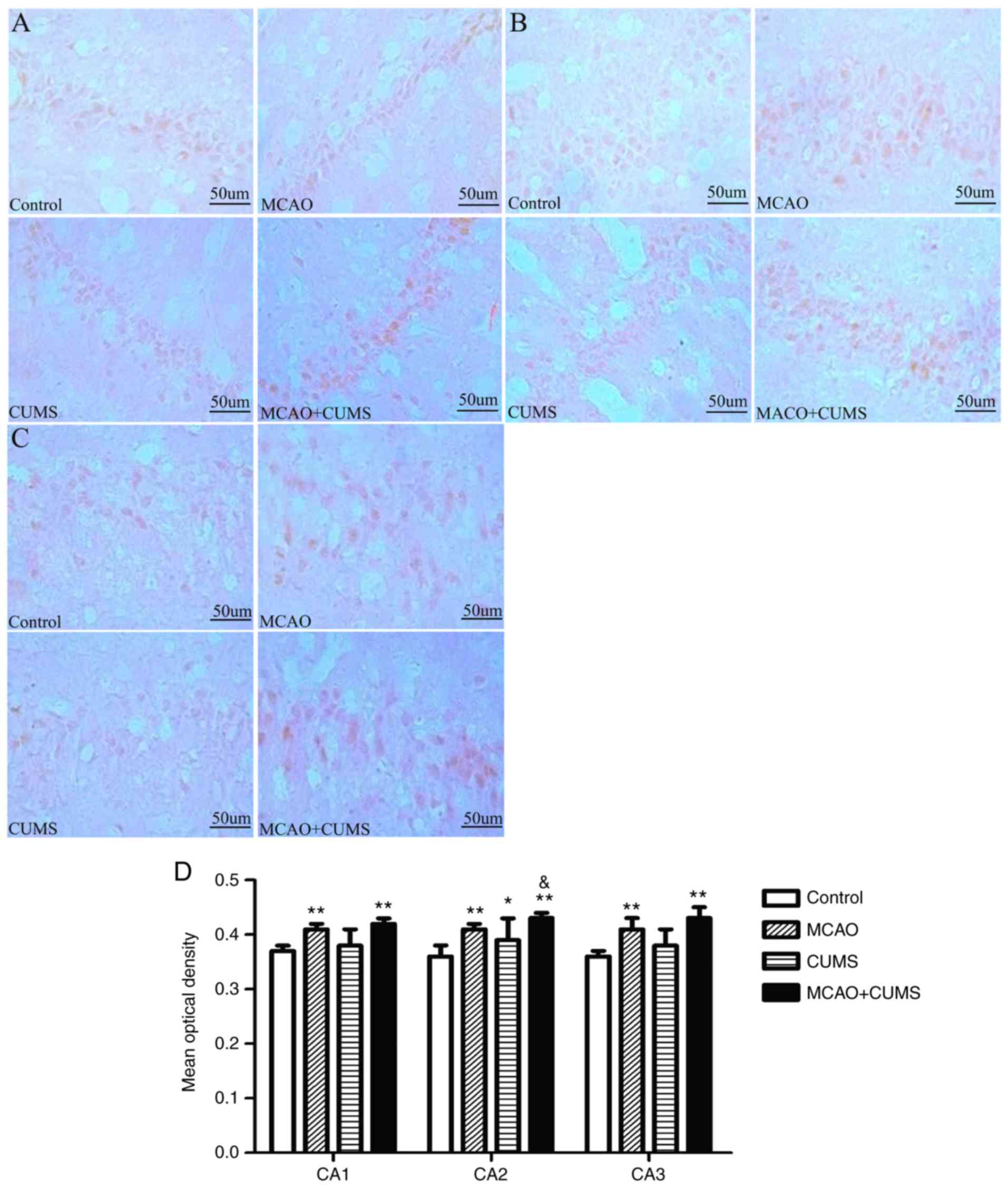
ARNT2-positive staining
ANOVA demonstrated that the mean optical density of
ARNT2-positive neurons in the CA1 and CA3 hippocampal areas of MCAO
and MCAO+CUMS rats was significantly higher than in the control
group (P<0.05; Fig. 1A,C and D).
Although staining in the CUMS group revealed more ARNT2-positive
neurons in CA1/3 than controls, the difference was not significant
(P>0.05; Fig. 1A,C and D).
Additionally, rats in the CUMS group exhibited increased ARNT2
staining in the CA2 area compared with controls (P<0.05;
Fig. 1B and D); and decreased ARNT2 staining in CA2
compared with MCAO+CUMS rats (P<0.01; Fig. 1B and D).
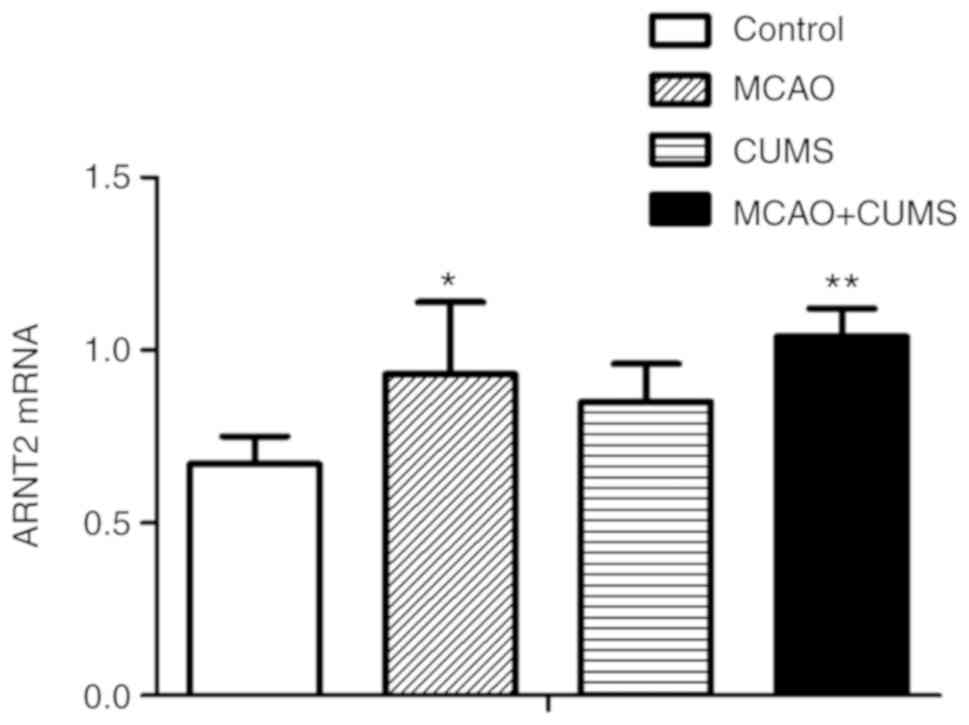
PCR of ARNT2 mRNA
ARNT2 mRNA levels in the hippocampi of MCAO and
MCAO+CUMS rats were significantly increased compared with in
controls (P<0.05 and <0.01, respectively); while there was no
significant difference between the MCAO and MCAO+CUMS groups
(P>0.05). Additionally, ARNT2 mRNA expression was higher in the
hippocampi of rats in the MCAO+CUMS group compared with in CUMS
rats, albeit non-significantly (P>0.05; Fig. 2).
Partial correlation analysis
results
The sucrose preference index and horizontal movement
distance in the OFT were positively correlated with ARNT mRNA level
(r=0.547 and 0.485; P<0.01). ARNT mRNA expression was also
positively correlated with body weight (r=0.067) and number of
rearings (r=0.014), though this was not significant (P>0.05;
Table V).
| Table VCorrelation (r-values) between rat
behavioral traits and ARNT mRNA expression. |
Table V
Correlation (r-values) between rat
behavioral traits and ARNT mRNA expression.
| | Body weight | Sucrose preference
index | Horizontal
movement | Number of
rearings |
|---|
| ARNT mRNA
expression | 0.067 | 0.547a | 0.485a | 0.014 |
Discussion
The results of the current study illustrated that
ARNT2 expression in the hippocampus differed among rats following
control, CUMS, MCAO and MCAO+CUMS treatments, which is consistent
with a previous study describing the effect of transcription
factors from the bHLH super-family on post-stroke depression
(25). The increased expression of
ARNT2 detected in the hippocampi of rats has previously been
confirmed to be associated with the vulnerability of distinct
dopaminergic projections to hypoxia (16,31), but
to our knowledge, the increased expression in the MCAO+CUMS group,
representing a model of PSD, is a novel finding. Additionally, the
observation that there was a trend toward increased ARNT2 levels in
the MCAO+CUMS group compared with in the MCAO group is also novel.
These alterations in ARNT2 expression were also confirmed at the
protein level, which is consistent with a previous study that found
HIF-1α-mediated increases in ARNT2 protein expression in neuronal
PC12 cells (32). By contrast,
another study detected decreased ARNT2 expression in response to
HIF-1α signaling in oral squamous cell carcinoma-derived cell lines
(33), which may be attributed to
variable cell types and altered cell cycle progression. Although
these conflicting findings warrant further research, the focus of
the current study was primarily on the change of ARNT2 in a PSD
model, and its specific mechanism is still unclear, worthy for
further study.
A notable finding was that the hippocampi isolated
from the MCAO and MCAO+CUMS groups exhibited increased levels of
ARNT2 protein expression as well as mRNA expression, reflecting its
potential protective role in resisting stress and promoting neuron
survival. Ischemic brain damage leads to secondary ischemic brain
tissue hypoxia, brain edema, degeneration, tissue softening,
necrosis and inflammation, which in turn induce apoptosis and cell
death (34).
Behavioral tests were also utilized to investigate
the association between the expression of ARNT2 and behavioral
performance. On day 7 after surgery, rats that underwent MCAO
gained less weight, consumed slightly less sucrose solution and
showed reduced activity, albeit temporarily, as a result of the
procedure. After 28 days, rats with MCAO appeared to somewhat
recover, although the BW of MCAO rats remained significantly lower
compared with that of control rats, potentially as a result of a
systemic response to the surgical injury. Moreover, the BW and
sucrose consumption of rats subjected to the CUMS regiment were
significantly decreased compared with in controls; further,
MCAO+CUMS rats gained less weight and consumed less sucrose
solution compared with MCAO rats. Collectively, these results
indicate successful establishment of the depression and PSD
models.
OFT was performed to measure spontaneous activity
and independent exploratory behavior, which has previously been
used to evaluate depressive behavior (35). In the current study, CUMS and
MCAO+CUMS rats subjected to a 4 week-CUMS regiment moved shorter
distances and performed a lower number of rearings compared with
controls. Furthermore, MCAO+CUMS rats moved shorter distances and
performed less rearings than those of the CUMS group, although
these differences were not significant. Overall these results
indicate that cerebral ischemia together with stress may serve a
role in behavioral performance. Interestingly, ARNT2 was
significantly elevated in the MCAO and MCAO+CUMS groups within
penumbra regions of the hippocampus, and therefore may be part of a
protective response to this type of injury. Moreover, levels of
ARNT were increased, which was previously observed in white blood
cells of patients with depression (36). Expectedly, MCAO+CUMS rats also
exhibited higher levels of ARNT2 compared with MCAO and CUMS rats.
However, the behavioral performance of MCAO+CUMS rats was
significantly worse compared with the other groups. Several factors
may account for this. First, it may be that the protective role of
ARNT2 is limited to compensation for damaged tissue, via
upregulation in the hippocampi; however, the regulation underlying
this ARNT2 expression has yet to be fully elucidated. Second, the
HIF-1α-dependent feed-forward loop that promotes the upregulation
of ARNT2, thus inhibiting apoptosis and necrosis of neurons
(31), may be prolonged in disease
states in order to ensure the survival of neurons in the brain.
Maltepe et al (15)
demonstrated that ARNT2/HIF-1α complexes associated with p53 and
B-cell lymphoma 2 family members to modulate apoptosis in response
to hypoxia/ischemia in neurons and other tissues. It has also been
indicated that chronic stress has a negative impact on
neurogenesis, with a previous study showing that cell proliferation
and survival, as well as neuronal differentiation, declined in
stressed rodents (25). Therefore, it
may be worthwhile investigating whether and how the survival and
apoptosis of neurons in the prefrontal cortex and hippocampus are
affected in PSD.
Previously, our group demonstrated that the bHLH-PAS
factor, NPAS4, which dimerizes with ARNT2 and subsequently binds to
brain-derived neurotrophic factor promoter-I to modulate
nerve-excitability-dependent transcription, was significantly
decreased in CUMS and MCAO+CUMS rats (25). Therefore, it is possible that ARNT2 is
involved in mechanisms underlying gene-environment interactions in
psychiatric models, but the specific biological signaling pathways
require further investigation. Additionally, further research is
warranted to clarify HIF-1 activity in neurons, using
ARNT2-deficient rats or ARNT2 antagonists/agonists. Nevertheless,
the results of the present study regarding the expression of ARNT2
in the hippocampus may serve as a basis for further study into the
etiology of psychiatric disorders following cerebral ischemia.
Acknowledgements
Not applicable.
Funding
The current study was supported by the National
Natural Science Foundation of China (grant no. 81471349), Henan
Science and Technology Development Project (grant no.
162102310489), the Natural Science Foundation of Henan Province
(grant no. 162300410224), Research on the Open Group of Phychiatric
Medicine in Xinxiang Medical College (grant no. 2016PN-KFKT-09),
the Training Plan for Young Excellent Teachers in Colleges and
Universities of Henan (grant no. 2016GGJS-106) and the Science and
Technology Project of Xinxiang (grant no. CXGG17030), and
additional support was provided by the 2016 Graduate Scientific
Research Innovation Support Program of Xinxiang (grant no.
YJSCX201625Y).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
WW made substantial contributions to conception and
design of the study. LZ performed the experiments on animal
behavior. SG was responsible for the polymerase chain reaction and
immunohistochemistry experiments. XW collected the data. PF
analyzed and interpreted data. CP drafted the manuscript. XZ
revised the manuscript critically for important intellectual
content. WL and JM ensured that questions related to the accuracy
or integrity of any part of the work were appropriately
investigated and resolved. ZZ and JS gave final approval of the
version to be published.
Ethics approval and consent to
participate
All experiments involving animals were approved by
the Laboratory Animals Management Commission of the Henan Key
Laboratory of Biological Psychiatry, Xinxiang, China.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Kronenberg G, Gertz K, Heinz A and Endres
M: Of mice and men: Modelling post-stroke depression
experimentally. Br J Pharmacol. 171:4673–4689. 2014.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Robinson RG and Jorge RE: Post stroke
depression: A review. Am J Psychiatry. 173:221–231. 2016.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Nabavi SF, Habtemariam S, Di Lorenzo A,
Sureda A, Khanjani S, Nabavi SM and Daglia M: Post-stroke
depression modulation and in vivo antioxidant activity of gallic
acid and its synthetic derivatives in a murine model system.
Nutrients. 8(E248)2016.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Mozaffarian D, Benjamin EJ, Go AS, Arnett
DK, Blaha MJ, Cushman M, de Ferranti S, Després JP, Fullerton HJ,
Howard VJ, et al: Heart disease and stroke statistics-2015 update:
A report from the American Heart Association. Circulation.
131:e29–e322. 2015.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Wang S, Sun H, Liu S, Wang T, Guan J and
Jia J: Role of hypothalamic cannabinoid receptors in post-stroke
depression in rats. Brain Res Bull. 121:91–97. 2016.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Swartz RH, Bayley M, Lanctôt KL, Murray
BJ, Cayley ML, Lien K, Sicard MN, Thorpe KE, Dowlatshahi D, Mandzia
JL, et al: Post-stroke depression, obstructive sleep apnea, and
cognitive impairment: Rationale for, and barriers to, routine
screening. Int J Stroke. 11:509–518. 2016.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Nabavi SF, Dean OM, Turner A, Sureda A,
Daglia M and Nabavi SM: Oxidative stress and post stroke
depression: Possible therapeutic role of polyphenols? Curr Med
Chem. 22:343–351. 2015.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Loubinoux I, Kronenberg G, Endres M,
Schumann-Bard P, Freret T, Filipkowski RK, Kaczmarek L and
Popa-Wagner A: Post-stroke depression: Mechanisms, translation and
therapy. J Cell Mol Med. 16:1961–1969. 2012.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Nabavi SF, Turner A, Dean O, Sureda A and
Mohammad S: Post stroke depression therapy: Where are we now? Curr
Neurovasc Res. 11:279–289. 2014.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Drutel G, Héron A, Kathmann M, Gros C,
Macé S, Plotkine M, Schwartz JC and Arrang JM: ARNT2 a
transcription factor for brain neuron survival? Eur J Neurosci.
11:1545–1553. 1999.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Dougherty EJ and Pollenz RS: Analysis of
Ah receptor-ARNT and Ah receptor-ARNT2 complexes in vitro and in
cell culture. Toxicol Sci. 103:191–206. 2008.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Rajashekaran P, Pai K, Thunga R and
Unnikrishnan B: Post-stroke depression and lesion location: A
hospital based cross-sectional study. Indian J Psychiatry.
55:343–348. 2013.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Keith B, Adelman DM and Simon MC: Targeted
mutation of the murine arylhydrocarbon receptor nuclear
translocator 2 (Arnt2) gene reveals partial redundancy with Arnt.
Proc Natl Acad Sci USA. 98:6692–6697. 2001.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Corcoran SE and O'Neill LA: HIF1α and
metabolic reprogramming in inflammation. J Clin Invest.
126:3699–3707. 2016.PubMed/NCBI View
Article : Google Scholar
|
|
15
|
Maltepe E, Keith B, Arsham AM, Brorson JR
and Simon MC: The role of ARNT2 in tumor angiogenesis and the
neural response to hypoxia. Biochem Biophys Res Commun.
273:231–238. 2000.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Dela Cruz JA, Schmidt-Kastner R, Stevens
JA, Steinbusch HW and Rutten BP: Differential distribution of
hypoxia-inducible factor 1-beta (ARNT or ARNT2) in mouse substantia
nigra and ventral tegmental area. J Chem Neuroanat. 61-62:64–71.
2014.PubMed/NCBI View Article : Google Scholar
|
|
17
|
den Heijer T, Tiemeier H, Luijendijk HJ,
van der Lijn F, Koudstaal PJ, Hofman A and Breteler MM: A study of
the bidirectional association between hippocampal volume on
magnetic resonance imaging and depression in the elderly. Biol
Psychiatry. 70:191–197. 2011.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Kempton MJ, Salvador Z, Munafò MR, Geddes
JR, Simmons A, Frangou S and Williams SC: Structural neuroimaging
studies in major depressive disorder. Meta-analysis and comparison
with bipolar disorder. Arch Gen Psychiatry. 68:675–690.
2011.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Erickson KI, Prakash RS, Voss MW, Chaddock
L, Heo S, McLaren M, Pence BD, Martin SA, Vieira VJ, Woods JA, et
al: Brain-derived neurotrophic factor is associated with
age-related decline in hippocampal volume. J Neurosci.
30:5368–5375. 2010.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Li G, Peskind ER, Millard SP, Chi P, Sokal
I, Yu CE, Bekris LM, Raskind MA, Galasko DR and Montine TJ:
Cerebrospinal fluid concentration of brain-derived neurotrophic
factor and cognitive function in non-demented subjects. PLoS One.
4(e5424)2009.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Zhang ZH, Wu LN, Song JG and Li WQ:
Correlations between cognitive impairment and brain-derived
neurotrophic factor expression in the hippocampus of post-stroke
depression rats. Mol Med Rep. 6:889–893. 2012. View Article : Google Scholar
|
|
22
|
Zhao DA, Bi LY, Huang Q, Zhang FM and Han
ZM: Isoflurane provides neuroprotection in neonatal hypoxic
ischemic brain injury by suppressing apoptosis. Braz J Anesthesiol.
66:613–621. 2016.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Xu Y, Tian Y, Tian Y, Li X and Zhao P:
Autophagy activation involved in hypoxic-ischemic brain injury
induces cognitive and memory impairment in neonatal rats. J
Neurochem. 139:795–805. 2016.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Valen E, Pascarella G, Chalk A, Maeda N,
Kojima M, Kawazu C, Murata M, Nishiyori H, Lazarevic D, Motti D, et
al: Genome-wide detection and analysis of hippocampus core
promoters using DeepCAGE. Genome Res. 19:255–265. 2008.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Zhang Z, Fei P, Mu J, Wang H, Li W and
Song J: Decreased expression of neuronal Per-Arnt-Sim domain
protein 4 gene in the hippocampus of a post-stroke depression rat
model. Exp Ther Med. 7:1045–1049. 2014.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Willner P: Chronic mild stress (CMS)
revisited: Consistency and behavioural-neurobiological concordance
in the effects of CMS. Neuropsychobiology. 52:90–110.
2005.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Chen Q, Ren L, Min S, Hao X, Chen H and
Deng J: Changes in synaptic plasticity are associated with
electroconvulsive shock-induced learning and memory impairment in
rats with depression-like behavior. Neuropsychiatr Dis Treat.
14:1737–1746. 2018.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Ren L, Hao X, Min S, Deng J, Chen Q, Chen
H and Liu D: Anesthetics alleviate learning and memory impairment
induced by electroconvulsive shock by regulation of NMDA
receptor-mediated metaplasticity in depressive rats. Neurobiol
Learn Mem. 155:65–77. 2018.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Araujo SM, Poetini MR, Bortolotto VC, de
Freitas Couto S, Pinheiro FC, Meichtry LB, de Almeida FP, Santos
Musachio EA, de Paula MT and Prigol M: Chronic unpredictable mild
stress-induced depressive-like behavior and dysregulation of brain
levels of biogenic amines in Drosophila melanogaster. Behav Brain
Res. 351:104–113. 2018.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Shao QY, You F, Zhang YH, Hu LL, Liu WJ,
Liu Y, Li J, Wang SD and Song MF: CSF miR-16 expression and its
association with miR-16 and serotonin transporter in the raphe of a
rat model of depression. J Affect Disord. 238:609–614.
2018.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Mandl M and Depping R: Hypoxia-inducible
aryl hydrocarbon receptor nuclear translocator (ARNT)(HIF-1β): Is
it a rare exception? Mol Med. 20:215–220. 2014.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Mandl M, Lieberum MK and Depping R: A
HIF-1α-driven feed-forward loop augments HIF signalling in Hep3B
cells by upregulation of ARNT. Cell Death Dis.
7(e2284)2016.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Kimura Y, Kasamatsu A, Nakashima D,
Yamatoji M, Minakawa Y, Koike K, Fushimi K, Higo M, Endo-Sakamoto
Y, Shiiba M, et al: ARNT2 regulates tumoral growth in oral squamous
cell carcinoma. J Cancer. 7:702–710. 2016.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Arteaga O, Álvarez A, Revuelta M,
Santaolalla F, Urtasun A and Hilario E: Role of antioxidants in
neonatal hypoxic-ischemic brain injury: New therapeutic approaches.
Int J Mol Sci. 18(E265)2017.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Kim YR, Kim HN, Pak ME, Ahn SM, Hong KH,
Shin HK and Choi BT: Studies on the animal model of post-stroke
depression and application of antipsychotic aripiprazole. Behav
Brain Res. 287:294–303. 2015.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Kajta M, Wnuk A, Rzemieniec J, Litwa E,
Lason W, Zelek-Molik A, Nalepa I, Rogóż Z, Grochowalski A and
Wojtowicz AK: Depressive-like effect of prenatal exposure to DDT
involves global DNA hypomethylation and impairment of GPER1/ESR1
protein levels but not ESR2 and AHR/ARNT signaling. J Steroid
Biochem Mol Biol. 171:94–109. 2017.PubMed/NCBI View Article : Google Scholar
|