Introduction
Luteolin (3',4',5,7-tetrahydroxyflavone; Fig. 1), is a natural, dietary flavonoid
commonly present at high concentrations in several types of fruits,
vegetables and medicinal herbs (1,2).
Flavonoids are polyphenols that serve an important role in
defending plant cells against microorganisms, insects and UV
irradiation (3). A number of
previous studies have demonstrated that luteolin possesses
beneficial pharmacological properties, including anti-oxidant,
anti-inflammatory, anti-microbial and anti-cancer properties
(4,5). Some of these properties could be
functionally associated with each other. For example, the
anti-inflammatory effects of luteolin may also be associated to its
anti-cancer properties (6).
Infection with Helicobacter pylori (H.
pylori), a bacterial carcinogen, is the greatest risk factor
for development of gastric cancer, which is the fifth most common
malignant disease and the third most common cause of
cancer-associated mortality worldwide (7). It has been reported that ~75% of global
cases of gastric cancer and 5.5% of malignancies worldwide may be
attributed to H. pylori-induced inflammation and injury
(8,9). However, the mechanisms that regulate
cancer development in response to this organism are not well
defined.
Altered glycosylation is considered to be one of the
most common characteristic features of cancer, and is the most
common post-translational modification of proteins occurring during
neoplastic transformation (10).
MUC1 mucin, transmembrane glycoprotein is expressed on the apical
surfaces of most epithelia, including the stomach, and is the
primary O-glycosylated protein of epithelial tissues. Generally,
MUC1 is composed of two subunits, a long N-terminal extracellular
domain and a short C-terminal tail, which remain associated through
hydrogen bonds (11). The
extracellular domain can be released from the cell surface by an
additional proteolytic cleavage event, which has been implicated in
the pathogenesis of inflammatory conditions and cancer (12,13).
Release of the N-terminal domain can be induced via activation of
specific enzymes, including the matrix metalloprotease a
disintegrase and metalloprotease (ADAM) metallopeptidase domain 17
(ADAM-17) (14,15).
In cancer, the O-glycan chains attached to
glycoproteins are often truncated and they commonly contain Tn
(GalNAcα1-O-Ser/Thr) and T (Galβ1-3GalNAcα1-O-Ser/Thr) antigens and
their sialylated forms (sTn and sT) (16). The aforementioned antigens are often
markers of poorly differentiated adenocarcinomas and mucinous
carcinoma, and their increased occurrence is associated with highly
proliferative tumors, metastasis, and poor clinical outcomes
(11).
The majority of types of gastric cancer are end
products of an inflammatory process. Chronic H. pylori
infection is characterized by inflammation of the gastric mucosa
and is the major cause of chronic gastritis (9,17,18).
IL-8 serves an important role in the response of epithelial cells
to H. pylori infection, as well as in the pathological
processes which ultimately result in gastric diseases. IL-8 is a
chemokine that is specific to neutrophil granulocyte chemotaxis,
and has been shown to be associated with the histological severity
of gastritis. IL-8 secretion is typically regulated by the
transcription factor NF-κB, and H. pylori can induce IL-8
expression by activating the NF-κB signaling pathway in gastric
epithelial cells (19). IL-10
represents anti-inflammatory cytokines in general; there are
several studies regarding the multifunctional roles of IL-10,
including both its immunosuppressive and anti-angiogenic effects,
and its varied roles in the pathogenesis, progression, metastasis,
and development of several types of cancer (20,21).
The primary purpose of the present study was to
determine the influence of luteolin on cancer-associated MUC1
(together with Tn and sT antigens), ADAM-17, the metalloprotease
involved in the release the extracellular domain of MUC1, IL-8 and
IL-10, which are associated with inflammation, and NF-κB, a
transcription factor regulating the expression levels of a number
of human genes. All these factors are potentially involved in
cancer development. The present experiments were performed on H.
pylori infected, gastric cancer CRL-1739 cells.
Materials and methods
Ethical approval and consent
The research protocol used in the present study was
approved by the Ethics Committee of Medical University of Białystok
(Białystok, Poland) and was performed in accordance with the 2008
Declaration of Helsinki (22).
Written informed consent was obtained from the patient.
Bacteria and cell culture
One laboratory H. pylori strain from the
Department of Microbiology of the Medical University of Białystok
(Białystok, Poland) was used in the present study. It was isolated
from gastric epithelial cells of a patient with gastritis. Prior to
the beginning of treatment, the scrapings were collected from the
prepyloric area and the body of the stomach under endoscopic
examination. Immediately afterwards, the scrapings were transferred
to the transport medium Portagerm pylori (bioMerieux SA) and
homogenized. Subsequently, the bacteria were cultured on Pylori
Agar and Columbia Agar supplemented with 5% sheep blood (bioMerieux
SA) for 7 days at 37˚C under microaerophilic conditions using a
Genbag microaer (bioMerieux SA). Bacteria were identified based on
the colony morphology, using the Gram method (23). Additionally, the activities of
bacterial urease, catalase and oxidase were determined as described
previously (24,25). To determine the H. pylori
species, an ELISA test (cat. no. HpAg48; EQUIPAR) was used.
Subsequently the bacteria were sub-cultured under the same
conditions, suspended at 1.2x109 bacteria/ml in PBS and
added to growing gastric cancer cells at a multiplicity of
infection of 10 for 24 h.
Gastric adenocarcinoma cells (CRL-1739; American
Type Culture Collection) were cultured in F-12 medium containing
10% heat inactivated FBS (Thermo Fisher Scientific, Inc), 100 U/ml
penicillin and 100 µg/ml streptomycin (Sigma-Aldrich; Merck KGaA)
at 37˚C with 5% CO2. Cells were seeded in 6-well plates.
A total of 24 h prior to H. pylori treatment, the cell
medium was changed to antibiotic-free F-12 medium. Subsequently,
media were supplemented with 30 µM luteolin alone or with bacteria,
and cultured for 24 h. The cells were washed with PBS and lysed at
4˚C using RIPA buffer (Sigma-Aldrich; Merck KGaA) supplemented with
protease inhibitors (1:200; Sigma-Aldrich; Merck KGaA). Culture
media and lysates were centrifuged at 1,000 x g for 5 min at 4˚C,
and supernatants were aliquoted, stored at -70˚C and used for
ELISA. For reverse transcription-quantitative PCR (RT-qPCR), the
monolayers were washed three times with sterile 10 mM PBS, and cell
membranes were disrupted using a sonicator (Sonics Vibra Cell;
Sonics & Materials, Inc). Aliquots of the homogenate were used
for RNA isolation. Cells not treated with either luteolin or H.
pylori addition were used as a control.
Cell viability assay
Cell viability assessment was performed as
previously described by Carmichael et al (26), using MTT (Sigma-Aldrich; Merck KGaA).
Confluent cells were cultured for 24 h with various concentrations
of luteolin (20-100 µM; Sigma-Aldrich; Merck KGaA) in six-well
plates, washed in PBS and incubated for 4 h in 1 ml MTT solution
(0.5 mg/ml PBS) at 37˚C with 5% CO2. Absorbance of the
converted dye in living cells was measured at a wavelength of 570
nm. Cell viability of gastric cancer cells in the presence of
luteolin was calculated as a percentage of the control cells.
ELISA for MUC1, and the Tn and sT
antigens
To assess the expression levels of MUC1 mucin, an
ELISA with an anti-MUC1 monoclonal antibody (BC2; Abcam; cat. no.
ab89492) was used according to the manufacturer's protocol. A total
of 50 µl cell lysates (100 µg protein/ml) or 50 µl media (1:100)
were coated on microtiter plates (NUNC F96; Maxisorp; Thermo Fisher
Scientific, Inc.) at room temperature overnight. Following blocking
with 100 µl 1% blocking reagent for ELISA, (Roche Diagnostics) and
three washes with 100 µl PBS with 0.05% Tween-20 (Sigma-Aldrich;
Merck KGaA), the plates were incubated with 100 µl anti-MUC1
antibody (1:600) for 2 h and horseradish peroxidase conjugated
rabbit anti-mouse IgG (Sigma-Aldrich; Merck KGaA) for 1 h at room
temperature. The colored reaction was developed by incubation with
100 µl 2,2'-azino-bis(3-ethylbenzthiazoline-6-sulfonic acid)
(Sigma-Aldrich; Merck KGaA) liquid substrate for horseradish
peroxidase. Absorbance at 405 nm was measured after 30-45 min.
Wells treated with BSA (Sigma-Aldrich; Merck KGaA) were used as
negative controls.
To assess the expression levels of GalNAc-R (Tn
antigen) and NeuAcα2-3Gal (sT antigen), an ELISA-like test with
biotinylated lectins (Vector Laboratories, Inc.; VVA lectin cat.
no. B-1235; MAA II lectin cat. no. B-1265) at a concentration of 5
µg/ml was performed. VVA lectin (from Vicia villosa with
binding preference to GalNAc) (27)
and MAA II lectin (from Maackia amurensis with binding
preference to NeuAcα2-3Gal) (28)
were used. A total of 50 μl medium (dilution 1:100) was used to
coat microtiter plates at room temperature overnight. Blocking and
washing steps were performed as mentioned above. Subsequently, the
plates were incubated with 100 µl proper lectins (2 h) and 100 µl
horseradish peroxidase avidin D (Vector Laboratories) for 1 h at
room temperature. The colored reaction was developed as described
above. All samples were analyzed in triplicate in three independent
experiments.
ELISA for IL-8
IL-8 expression was quantitatively determined using
a commercially available ELISA capture and detection antibody kit
(BD OptEIATM Set Human IL-8; BD Biosciences; cat. no. 2654KI)
according to the manufacturer's protocol. Briefly, microwells of a
96-well plate were coated with 100 µl capture antibody (1:250) in
bicarbonate buffer (pH 9.5; 0.1 M) and incubated overnight at 4˚C.
The plates were washed three times with 200 µl PBS with Tween-20
(0.05%) between all the steps. Unbound sites were blocked with 200
µl PBS with 10% FBS (Sigma-Aldrich; Merck KGaA). Subsequently the
cell culture media (50 µl; 1:100) were added and plates were
incubated for 2 h at room temperature, followed by incubation with
biotinylated detection antibody (1:250), peroxidase-labeled
streptavidin and tetramethylbenzidine substrate (Sigma-Aldrich;
Merck KGaA). Absorbance at 450 nm was measured after 30 min and
IL-8 levels were determined from a standard curve prepared with
serial dilutions of purified chemokines. All samples were analyzed
in triplicate or quadruplicate, in three independent tests, and
standard curves were plotted for each plate.
RT-qPCR
Total RNA was isolated using Total RNA Mini Plus
Concentrator (A&A Biotechnology) according to the
manufacturer's protocol. The concentration and purity of RNA was
determined by spectrophotometry using a Nanodrop 2000 (Thermo
Fisher Scientific, Inc.). First-strand cDNA was synthesized from 1
µg total RNA using a Tetro cDNA Synthesis kit (Bioline; Meridian
Bioscience). The reaction mixture (volume, 20 µl) containing 1 µl
oligo(dT)18 primer, 1 µl dNTP mixture (10 mM each), 5 µl 5X RT
Buffer, 1 µl RiboSafe RNase Inhibitor (10 U/µl), 1 µl Tetro Reverse
Transcriptase (200 U/µl) and diethylpyrocarbonate-treated water was
incubated for 30 min at 45˚C and then inactivated at 85˚C for 5
min. qPCR assay was performed using a CFX96 Real-time system
(Bio-Rad Laboratories, Inc.) and a SensiFASTTM SYBR kit (Bioline;
Meridian Bioscience). The reaction mixtures contained 2 µl twice
diluted cDNA template, 0.8 µl of each primer (10 µmol/l), 10 µl 2X
SensiFAST SYBR mix and nuclease-free water to a final volume of 20
µl. Forward and reverse primer sequences are listed in Table I. The primers were synthesized by
Genomed. GAPDH was used as the housekeeping gene. The thermocycling
conditions were: 95˚C for 1 min to activate the DNA polymerase,
followed by 40 cycles of denaturation for 10 sec at 95˚C, annealing
for 15 sec at 60˚C and extension for 20 sec at 72˚C. The reaction
was then subjected to a melting protocol from 55 to 95˚C in 0.2˚C
increments and 1 sec holding at each increment to assess the
specificity of the amplified products. Single product formation was
confirmed by melting point analysis and agarose gel
electrophoresis. As a negative control, water was used instead of
mRNA samples. Samples were run in triplicate and the
2-ΔΔCq method was used to calculate the relative
expression (29). The relative gene
expression levels were standardized to those measured for the
untreated control.
 | Table ISequences of primers used in the
present study. |
Table I
Sequences of primers used in the
present study.
| Gene | Forward primer,
5'-3' | Reverse primer,
5'-3' |
|---|
| MUC1 |
TGCCTTGGCTGTCTGTCAGT |
GTAGGTATCCCGGGCTGGAA |
| NF-κB |
TACTCTGGCGCAGAAATTAGGTC |
CTGTCTCGGAGCTCGTCTATTTG |
| IL-8 |
TAGCAAAATTGAGGCCAAGG |
AAACCAAGGCACAGTGGAAC |
| IL-10 |
TGGTGAAACCCCGTCTCTAC |
CTGGAGTACAGGGGCAGTAT |
| ADAM-17 |
ACCTGAAGAGCTTGTTCATCGAG |
CCATGAAGTGTTCCGATAGATGTC |
| GAPDH |
GTGAACCATGAGAAGTATGACAA |
CATGAGTCCTTCCACGATAC |
Statistical analysis
Experimental data are presented as the mean ±
standard deviation of three experimental repeats. A one-way ANOVA
followed by a Duncan's post hox test was used to analyze
differences between the control and specific treatment groups.
Statistica version 10.0 (StatSoft, Inc.) was used to statistically
analyze the data. P<0.05 was considered to indicate a
statistically significant difference.
Results
Cell viability
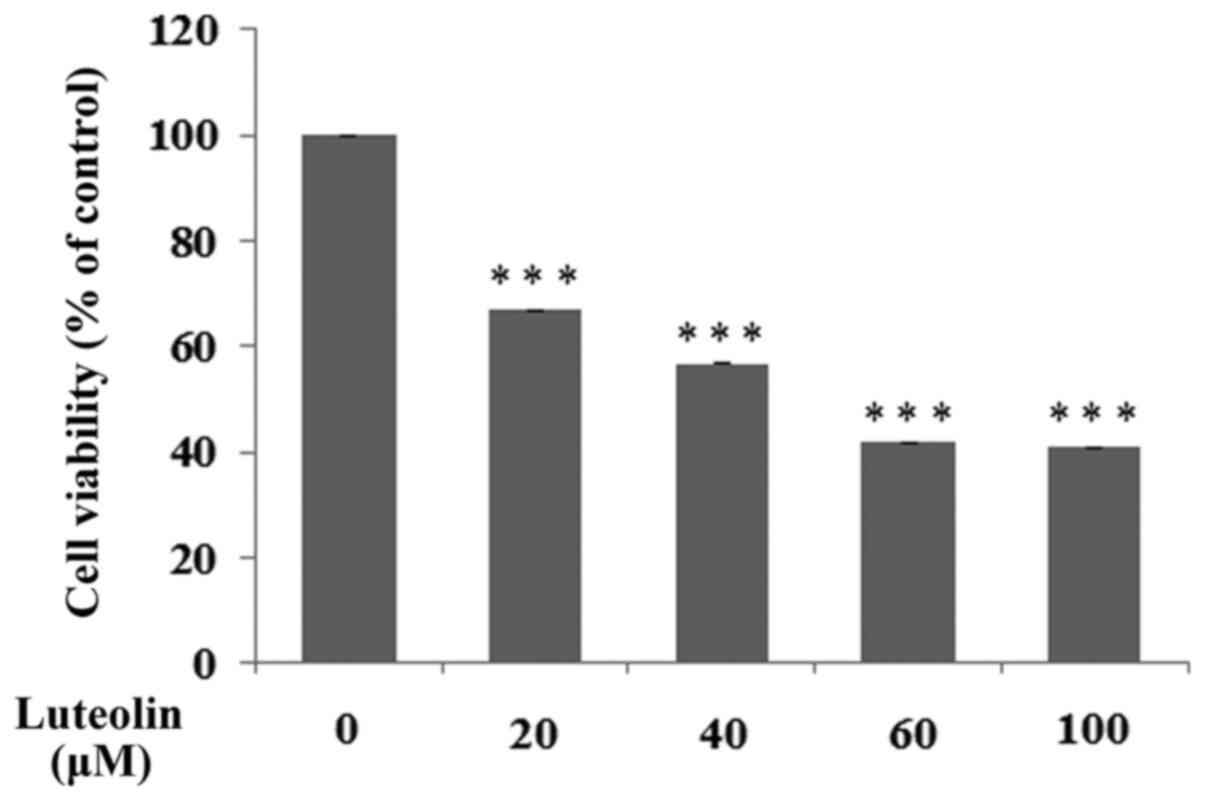
The effect of luteolin on a number of selected
biochemical factors was examined in H. pylori infected
gastric cancer CRL-1739 cells. In all experiments, 30 µM luteolin
was used as a concentration, which was below the IC50
(Fig. 2).
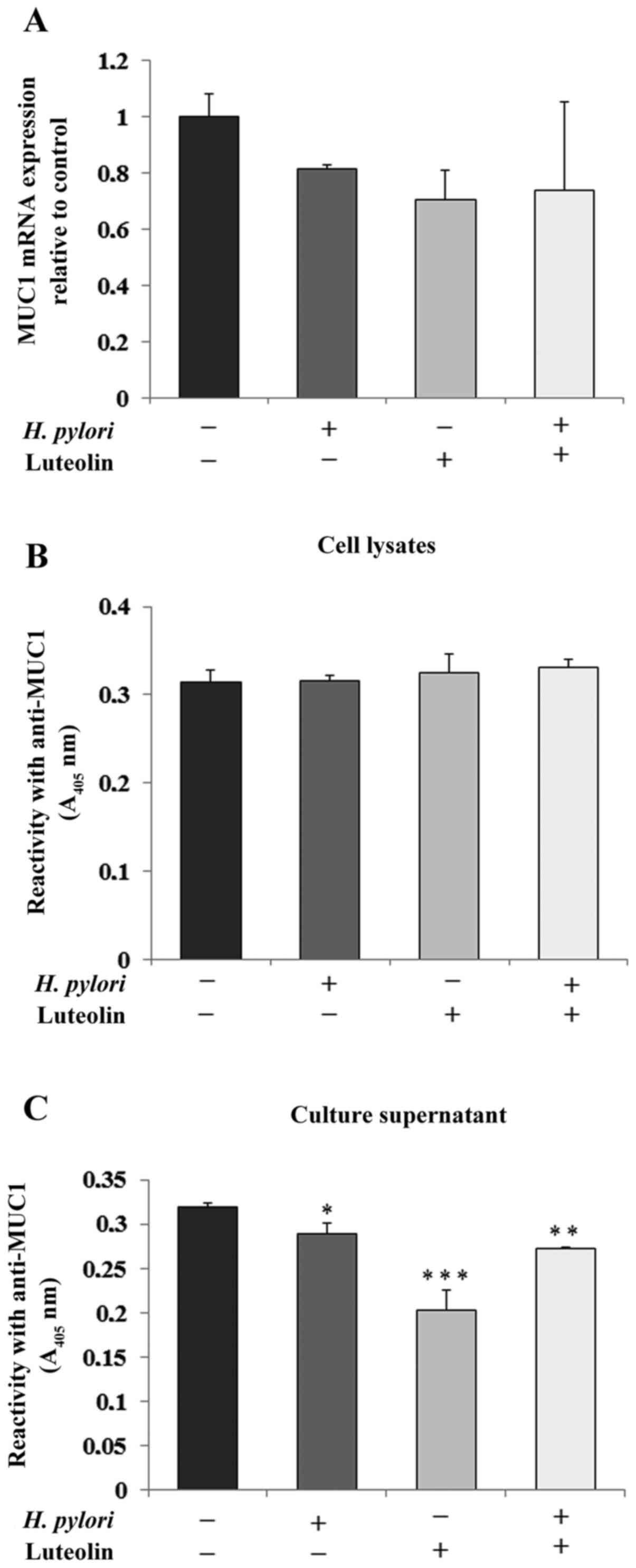
Determination of MUC1
MUC1 is the primary epithelial glycoprotein present
in the gastric epithelium (11). As
shown in Fig. 3A, H. pylori,
luteolin and H. pylori combined with luteolin did not have
significant effects on MUC1 mRNA expression compared with the
untreated control. There was no significant change in MUC1
expression in the cell lysates (Fig.
3B). However, the examined factors notably reduced MUC1
extracellular domain expression in culture medium (Fig. 3C).
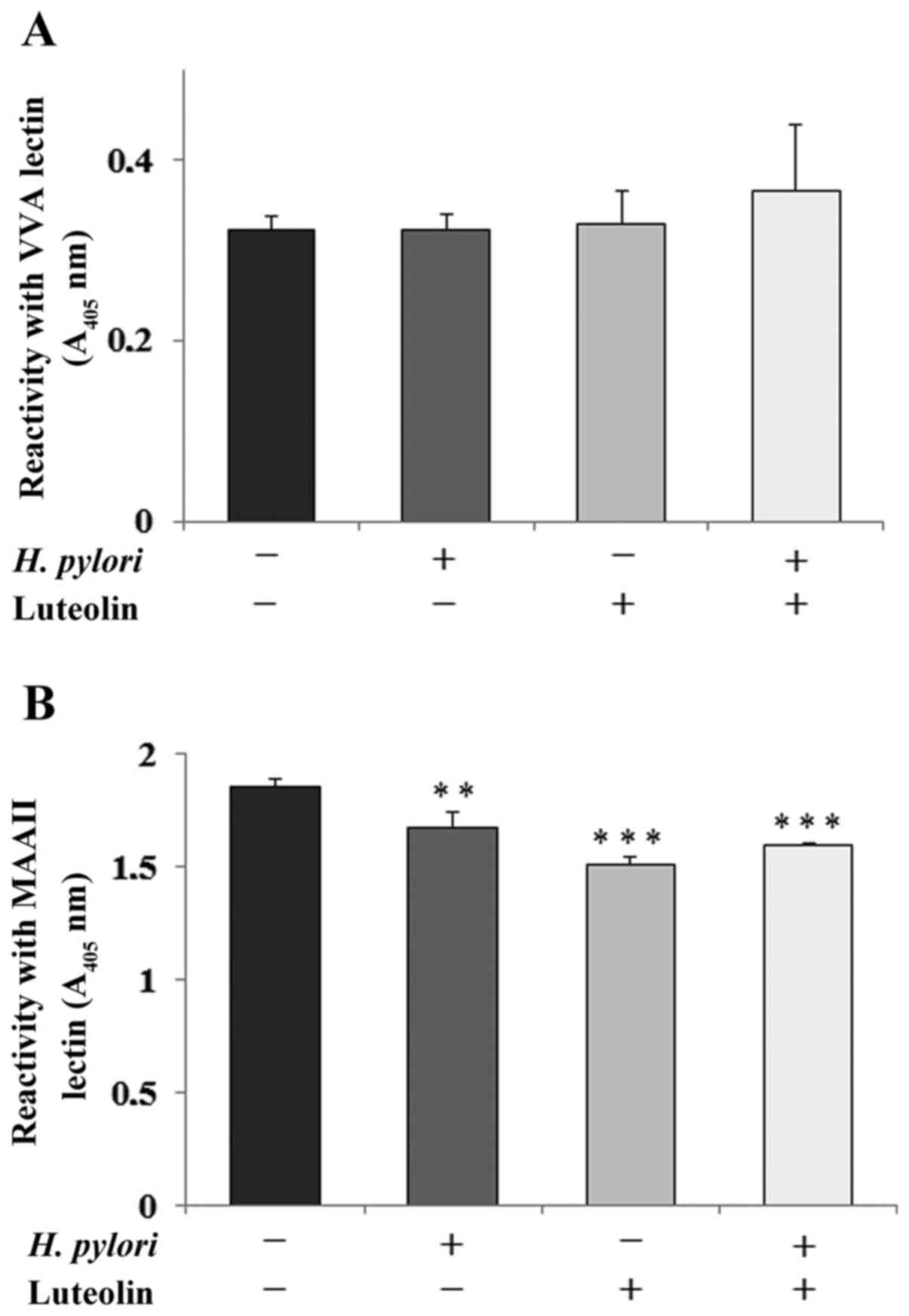
Determination of Tn and sT
antigens
GalNAcα-R (Tn antigen) and NeuAcα2-3Galβ1-3GalNAc-R
(sT antigen) are amongst the core sugar structures that are present
on the extracellular domain of MUC1 mucin (11). The expression levels of Tn antigen
(based on reactivity with VVA lectin) were not significantly
affected by bacteria or luteolin (Fig.
4A). Sialylation of T antigen (detected by MAAII lectin) was
significantly inhibited by flavonoid and H. pylori addition
separately and by both agents combined, compared with the control
(Fig. 4B).
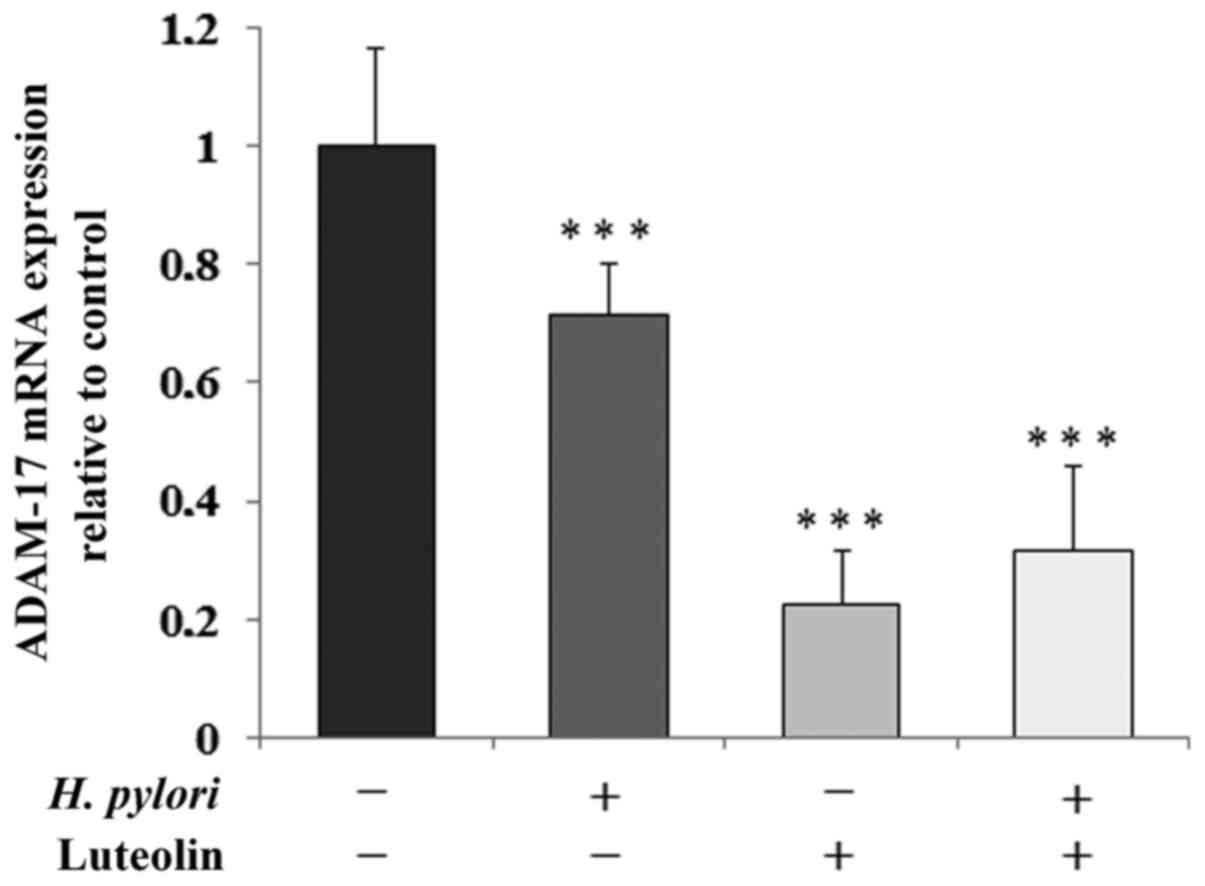
Determination of ADAM-17 levels
ADAM-17 is one of the sheddases that is responsible
for the release of MUC1 extracellular domain. In all conditioned
cultures, sheddase was markedly inhibited by the examined factors
at the mRNA level (by 29, 78 and 68% for gastric cancer cells
treated with H. pylori, luteolin independently and combined,
respectively) compared with the untreated control (Fig. 5).
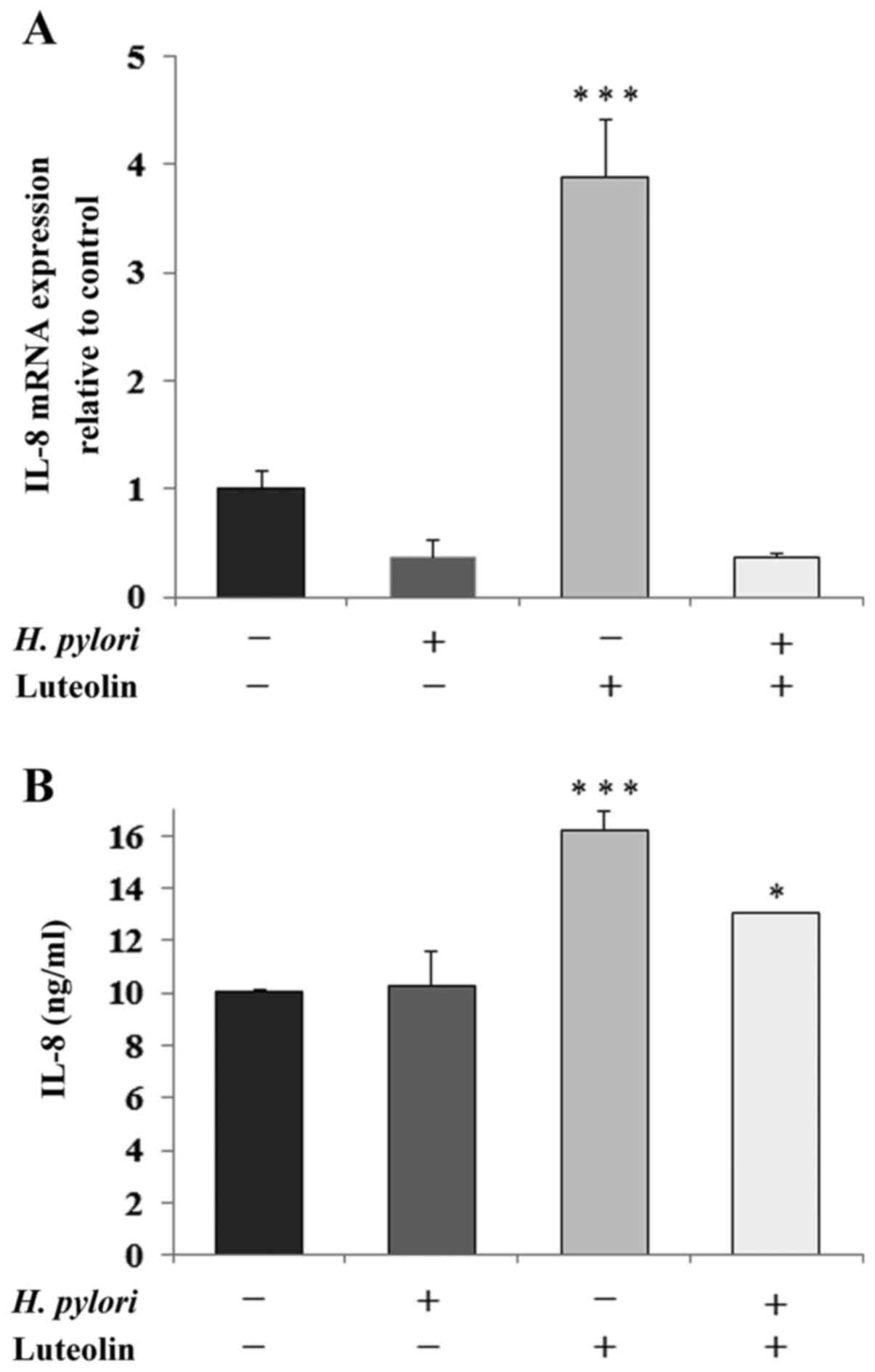
Determination of IL-8 and IL-10
IL-8 is a proinflammatory chemokine that is
considered to be an important regulatory factor in the tumor
environment. Fig. 6 shows a marked
stimulatory effect of luteolin on IL-8 expression at both the mRNA
(Fig. 6A) and protein (Fig. 6B) levels compared with the untreated
control. The protein expression levels of IL-8 were also stimulated
by the simultaneous action of both examined factors compared with
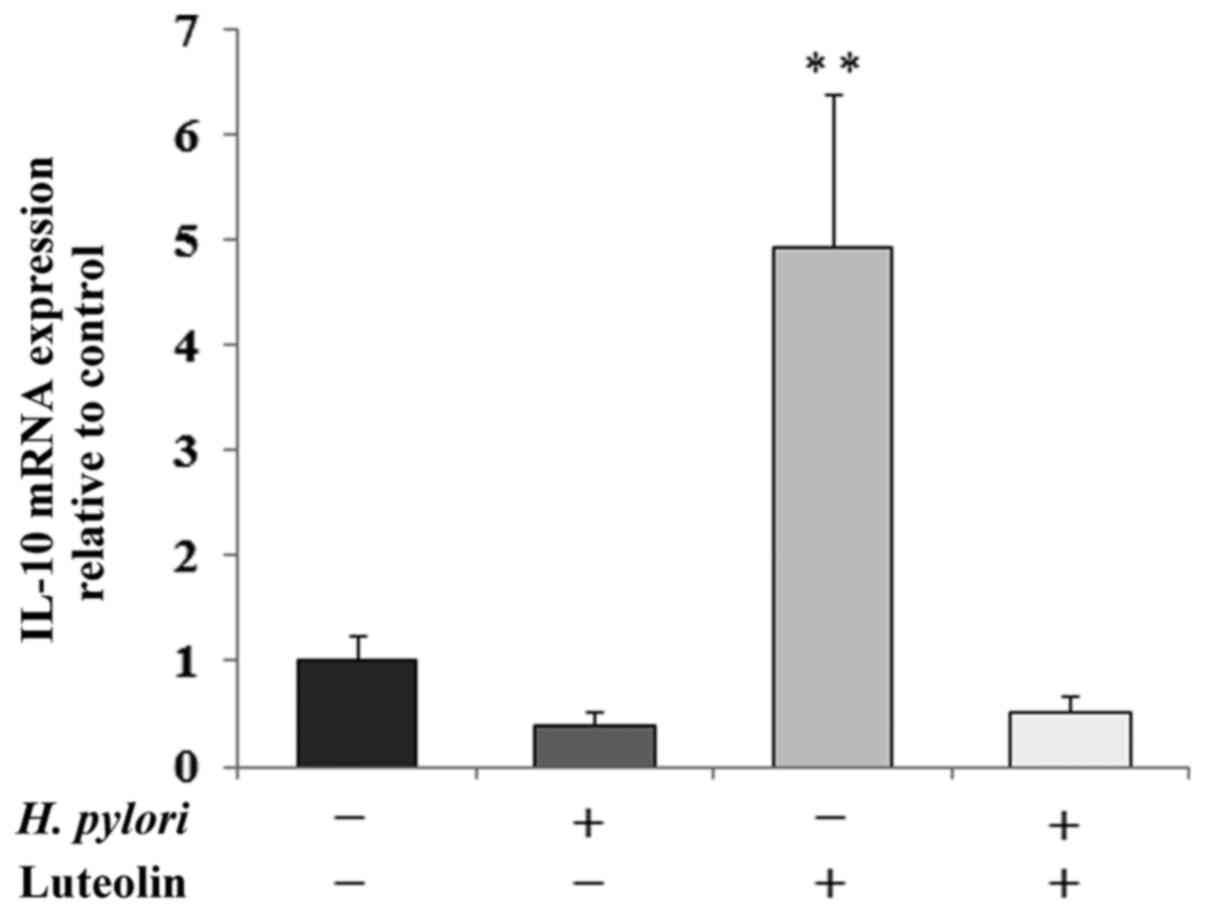
the control. IL-10 is an inhibitory cytokine which can help tumor
cells evade the immune system to avoid destruction by cell-mediated
immune mechanisms. A marked increase in IL-10 mRNA expression
following luteolin treatment was observed (Fig. 7).
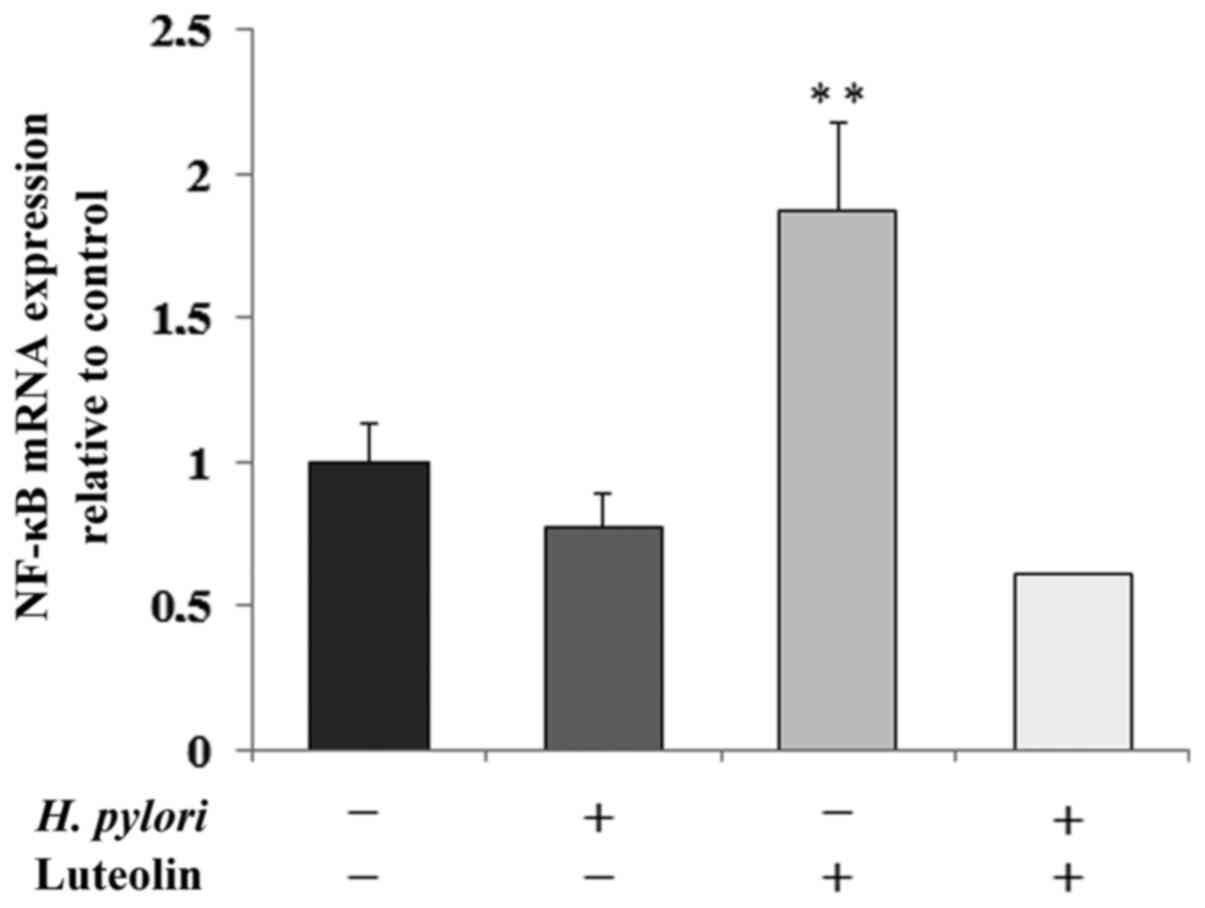
Determination of NF-κB expression
NF-κB is a transcription factor modulated by
numerous stimuli, and is able to regulate the expression levels of
several human genes. As shown in Fig.
8, NF-κB mRNA expression was significantly increased by
luteolin treatment by 87% compared with the untreated control.
Discussion
Gastric cancer is one of the most common malignances
worldwide, and is a serious risk to human health due to its
asymptomatic course of development, nonspecific symptoms during the
early stages and the high mortality rates associated with advanced
stages (7,9). Furthermore, there are limited
efficacious therapeutic strategies for treating advanced gastric
cancer (30,31). Therefore, it remains a priority to
develop novel therapeutic reagents for treatment of gastric cancer.
Long-term consumption of fruit and vegetables reduces the risk of
cancer (32). Therefore,
identification of natural phytochemicals as potential anticancer
agents may improve treatment as they are typically less toxic than
chemotherapeutic agents, and thus, this approach has gained
increasing attention (3,33,34).
Luteolin is a dietary flavonoid that has been
reported to inhibit the development and progression of several
different types of tumors (35,36). Pu
et al (36) reported that
luteolin reduced cell viability, induced cell cycle arrest, colony
formation, proliferation and migration, and promoted apoptosis of
gastric cancer MKN45 and BGC823 cells. According to Zang et
al (37) luteolin inhibited
gastric cancer progression via suppression of Notch 1 signaling and
reversal of epithelial-mesenchymal transition in gastric cancer
Hs-746T and MKN28 cells. Due to these promising results regarding
the use of luteolin as an anti-cancer agent, the present study
attempted to examine its effect on a number of other factors
potentially involved in gastric cancer development. Since H.
pylori is considered to be a causative agent of gastric cancer
(8) H. pylori-infected
gastric cancer cells were used, as well as the cells not treated
with these bacteria.
Cancers are diverse and complex diseases based on
multiple etiologies and various cell targets. A number of these
targets are associated with cancer development, however, not all
the relations have been identified. A possible target for cancer
cells is MUC1 mucin with truncated O-glycans (11,38). The
present study examined the effects of luteolin on a number of
selected factors, which were all cancer-related; however not all of
these appeared to be associated with cancer development or
progression in the present study.
MUC1 mucin is considered to be an oncoprotein based
on evidence which suggests its cancer-promoting function. MUC1 can
activate anti-apoptotic proteins (Bcl-xL) (39), attenuate apoptosis execution pathways
(40,41) and may contribute to metastasis
(42). Furthermore, MUC1 is involved
in H. pylori infection development via direct interaction
with bacterial adhesins (17,43). To
the best of our knowledge, there are no studies assessing the
effects of luteolin on MUC1 mucin. The present study showed the
inhibitory effects of bacteria and luteolin on the expression
levels of the extracellular domain of MUC1 mucin, with a more
potent effect being observed following luteolin treatment. Notably,
this effect was not observed at the mRNA level. It has been
suggested that ADAM-17 is involved in the release of MUC1
extracellular domain into the culture medium (14,15). The
results of the present study appear to support this hypothesis,
since ADAM-17 mRNA expression was associated increased MUC1 release
into the culture medium. Additionally, ADAM-17 (also known as
TNF-α-converting enzyme) has been identified to function as a
signaling scissor in the tumor microenvironment, and thus
contributes to tumorigenesis and tumor progression (44). Upregulation of ADAM-17 is associated
with the progression of non-small cell lung cancer (45) or the promotion of breast cancer
tumorigenesis by regulating cell proliferation, angiogenesis,
invasion and apoptosis (46).
Upregulation of ADAM-17 contributes to the progression of gastric
cancer and is associated with a poor prognosis (47,48).
Therefore, the enzyme is considered to be a potential target for
treatment of cancer, as well as an indicator for predicting
therapeutic outcomes. The present study revealed the inhibitory
effect of luteolin on ADAM-17 gene expression, which appears to
support the hypothesis regarding the potential anti-cancer effects
of the examined flavonoids. Thus, it is hypothesized that luteolin
is involved in the inhibition of degradation of ECM components by
decreasing ADAM-17 expression.
In several types of cancer, O-glycan chains attached
to glycoproteins, including MUC1 mucin, the primary O-glycoprotein
of gastric epithelium, commonly contain short carbohydrate forms,
Tn and T antigens and their sialylated derivatives. Their increased
occurrence is associated with highly proliferative tumors,
metastasis, and poor clinical outcomes (11,16).
Loss or acquisition of glycans affects interactions of MUC1 and
other cellular proteins implicated in the migratory and metastatic
activities of cancer cells. Santos-Silva et al (49) showed that 53.2% of gastric carcinomas
express the sT antigen, indicating that sialylation may contribute
to the low frequency of cases observed with T antigen expression.
The synthesis of the sT structure stops further processing and
elongation of the carbohydrate chain (16). Yu et al (50) showed that the cancer-associated T
antigen on MUC1 is the natural ligand of galectin-3, a galactose
binding protein, which after connecting to MUC1, initiates
MUC1-dependent intracellular signaling in cancer cells and
facilitates adhesion of cancer cells to each other and to
endothelial cells (51). Therefore,
inhibition of sT structure expression by luteolin, which was
observed in the present study, may be associated with its possible
anti-cancer activity.
It has been stated that the interaction between
H. pylori with the gastric epithelium induces the production
of IL-8, which is a proinflammatory cytokine and chemotaxin for
neutrophils and mononuclear cells, and this can lead to chronically
activated gastritis (19). The
majority of gastric cancer cases are the end products of an
inflammatory process (19,20,52). A
significant association between high expression levels of IL-8 in
the gastric mucosa and the risk of gastric cancer has been reported
previously (19,53). Furthermore, H. pylori can
stimulate the secretion of IL-10, which has been recently
recognized as the most potent anti-inflammatory cytokine, and also
as the agent enabling cancer immune surveillance and tumor
rejection (54). The results of the
present study were not consistent with the aforementioned results,
as H. pylori reduced IL-8 and IL-10 expression, in contrast
to the previous studies. One possible explanation for this result
could be that the action of the pathogen on gastric cancer cells
was too short (24 h). Notably, in the present study, luteolin
clearly increased expression of both cytokines at the mRNA level
and IL-8 at the protein level as well. These results are not
consistent with those reported in other studies which demonstrated
the inhibition of IL-8 upregulation by the examined flavonoid
(55,56). However, there is also at least one
report regarding the stimulating effects of luteolin on IL-8
expression. Lee et al (57)
showed that relatively low concentrations of luteolin (20 and 40
µM) increased the IL-8 levels, and only a high concentration of
flavonoid (80 µM) was shown to decrease the expression levels of
IL-8 in lung cancer cells.
It has been reported that direct contact of H.
pylori and gastric cancer cells induces NF-κB activation
(58). However, this result was not
observed in the present study, and it was suggested that this could
be due to the exposure of gastric cells to the pathogen being too
short as mentioned above. Notably, luteolin induced NF-κB mRNA
expression, which is not consistent with the general tendency of
anti-inflammatory actions of the flavonoid via the NF-κB signaling
pathway (58). At the current stage
of the present study, this discrepancy cannot be explained.
However, an association between the effect of luteolin on nuclear
factor and IL-8 expression was revealed, in agreement with the
general tendency of NF-κB to stimulate chemokine production
(58).
Based on the preliminary results of the present
study, it may be assumed that luteolin may be used as an adjuvant
for treatment of gastric cancer. This hypothesis is particularly
based on the outcomes concerning MUC1 extracellular domain, mRNA
ADAM-17 and sT antigen expression. There are some discrepancies
between the results of the present study and the results of
previous studies, for example regarding IL-8, IL-10 and NF-κB
levels. This requires further exploration in future experiments.
There were also some shortcomings of the present study. The
limitations include using only one cancer cell line and the lack of
a normal cell line. However, it has been shown in other studies
that the effects of luteolin are specific to cancer cells at the
concentrations used in the present study (59). Additional experiments, such as
apoptosis assays and cell cycle analysis, combined with the use of
additional experimental methods, such as western blotting and
immunofluorescence analysis should be performed to determine how
luteolin regulates the changes in the examined factors and to
identify the targets of the flavonoid. In future experiments, how
H. pylori affects cells growth at different multiplicities
of infection will be assessed. Other experiments where MUC1 or
ADAM-17 are knocked down will also be performed.
Experimental and clinical data suggest that
pharmacological regulation of various factors participating in
cancer development exerts several beneficial effects improving the
outcomes of different anti-cancer therapies. Dysregulation of
cancer cells results from a number of genetic alterations, which
impacts the signaling network and the control of numerous cellular
processes. Thus, therapeutic targeting of specific factors may also
elicit opposing, often unwanted effects. Therefore, extensive
investigations are required to understand the molecular signaling
pathway network for the development of synergistic complex
therapies for cancer treatments.
Acknowledgements
The authors would like to thank Ms. Joanna Wosek
(Department of Medical Chemistry, Medical University of Białystok
(Białystok, Poland) for her technical support.
Funding
The present study was supported by the Medical
University of Białystok (Białystok, Poland) (grant no.
N/ST/15/004/2203).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
IR designed the study, interpreted the data and
drafted the manuscript. MBK performed some of the experiments,
assisted in interpretation of the data and performed the
statistical analysis. KL performed some of the experiments and
assisted in interpretation of the data. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
The research protocol was approved by the Ethics
Committee of Medical University of Białystok (Białystok, Poland)
and was performed in accordance with the 2008 Declaration of
Helsinki. Written informed consent was obtained the patient.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they no competing
interests.
References
|
1
|
Xiong J, Li S, Wang W, Hong Y, Tang K and
Luo Q: Screening and identification of the antibacterial bioactive
compounds from Lonicera japonica Thunb. leaves. Food Chem.
138:327–333. 2013.PubMed/NCBI View Article : Google Scholar
|
|
2
|
López-Lázaro M: Distribution and
biological activities of the flavonoid luteolin. Mini Rev Med Chem.
9:31–59. 2009.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Birt DF, Hendrich S and Wang W: Dietary
agents in cancer prevention: Flavonoids and isoflavonoids.
Pharmacol Ther. 90:157–177. 2001.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Lin Y, Shi R, Wang X and Shen HM:
Luteolin, a flavonoid with potential for cancer prevention and
therapy. Curr Cancer Drug Targets. 8:634–646. 2008.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Nepali S, Son JS, Poudel B, Lee JH, Lee YM
and Kim DK: Luteolin is a bioflavonoid that attenuates
adipocyte-derived inflammatory responses via suppression of nuclear
factor-κB/mitogen-activated protein kinases pathway. Pharmacogn
Mag. 11:627–635. 2015.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Johnson JL and Gonzalez de Mejia E:
Interactions between dietary flavonoids apigenin or luteolin and
chemotherapeutic drugs to potentiate anti-proliferative effect on
human pancreatic cancer cells, in vitro. Food Chem Toxicol.
60:83–91. 2013.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Siegel R, Ma J, Zou Z and Jemal A: Cancer
statistics, 2014. CA Cancer J Clin. 64:9–29. 2014.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Amieva M and Peek RM Jr: Pathobiology of
Helicobacter pylori -induced gastric cancer.
Gastroenterology. 150:64–78. 2016.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Ang TL and Fock KM: Clinical epidemiology
of gastric cancer. Singapore Med J. 55:621–628. 2014.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Christiansen MN, Chik J, Lee L, Anugraham
M, Abrahams JL and Packer NH: Cell surface protein glycosylation in
cancer. Proteomics. 14:525–546. 2014.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Cascio S and Finn OJ: Intra- and
extra-cellular events related to altered glycosylation of MUC1
promote chronic inflammation, tumor progression, innovation and
metastasis. Biomolecules. 6(E39)2016.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Hollingsworth MA and Swanson BJ: Mucins in
cancer: Protection and control of the cell surface. Nat Rev Cancer.
4:45–60. 2004.PubMed/NCBI View
Article : Google Scholar
|
|
13
|
Mukhopadhyay P, Chakraborty S, Ponnusamy
MP, Lakshmanan I, Jain M and Batra SK: Mucins in the pathogenesis
of breast cancer: Implications in diagnosis, prognosis and therapy.
Biochim Biophys Acta. 1815:224–240. 2011.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Thathiah A, Blobel CP and Carson DD: Tumor
necrosis factor-α converting enzyme/ADAM 17 mediates MUC1 shedding.
J Biol Chem. 278:3386–3394. 2003.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Goth CK, Halim A, Khetarpal SA, Rader DJ,
Clausen H and Schjoldager KT: A systematic study of modulation of
ADAM-mediated ectodomain shedding by site-specific O-glycosylation.
Proc Natl Acad Sci USA. 112:14623–14628. 2015.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Fu C, Zhao H, Wang Y, Cai H, Xiao Y, Zeng
Y and Chen H: Tumor-associated antigens: Tn antigen, sTn antigen,
and T antigen. HLA. 88:275–286. 2016.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Lillehoj EP, Guang W, Ding H, Czinn SJ and
Blanchard TG: Helicobacter pylori and gastric inflammation:
Role of MUC1 mucin. J Pediatr Biochem. 2:125–132. 2012.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Wang F, Meng W, Wang B and Qiao L:
Helicobacter pylori-induced gastric inflammation and gastric
cancer. Cancer Lett. 345:196–202. 2014.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Lee KE, Khoi PN, Xia Y, Park JS, Joo YE,
Kim KK, Choi SY and Jung YD: Helicobacter pylori and
interleukin-8 in gastric cancer. World J Gastroenterol.
19:8192–8202. 2013.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Ahmad N, Ammar A, Storr SJ, Green AR,
Rakha E, Ellis IO and Martin SG: IL-6 and IL-10 are associated with
good prognosis in early stage invasive breast cancer patients.
Cancer Immunol Immunother. 67:537–549. 2018.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Fasoulakis Z, Kolios G, Papamanolis V and
Kontomanolis EN: Interleukins associated with breast cancer.
Cureus. 10(e.3549)2018.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Puri KS, Suresh KR, Gogtay NJ and Thatte
UM: Declaration of Helsinki, 2008: Implications for stakeholders in
research. J Postgrad Med. 55:131–134. 2009.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Beveridge TJ: Use of the gram stain in
microbiology. Biotech Histochem. 76:111–118. 2001.PubMed/NCBI
|
|
24
|
Uotani T and Graham DY: Diagnosis of
Helicobacter pylori using the rapid urease test. Ann Transl
Med. 3:9–7. 2015.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Bai Y, Zhang YL, Jin JF, Wang JD, Zhang ZS
and Zhou DY: Recombinant Helicobacter pylori catalase. World
J Gastroenterol. 9:1119–1122. 2003.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Carmichael J, DeGraff WG, Gazdar AF, Minna
JD and Mitchell JB: Evaluation of a tetrazolium-based semiautomated
colorimetric assay: Assessment of chemosensitivity testing. Cancer
Res. 47:936–942. 1987.PubMed/NCBI
|
|
27
|
Puri KD, Gopalakrishnan B and Surolia A:
Carbohydrate binding specificity of the Tn-antigen binding lectin
from Vicia villosa seeds (VVLB4). FEBS Lett. 312:208–212.
1992.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Redelinghuys P, Antonopoulos A, Liu Y,
Campanero-Rhodes MA, McKenzie E, Haslam SM, Dell A, Feizi T and
Crocker PR: Early murine T-lymphocyte activation is accompanied by
a switch from N-Glycolyl- to N-acetyl-neuraminic acid and
generation of ligands for siglec-E. J Biol Chem. 286:34522–34532.
2011.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2-ΔΔCT method. Methods. 25:402–408. 2001.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Zong L, Abe M, Seto Y and Ji J: The
challenge of screening for early gastric cancer in China. Lancet.
388(2606)2016.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Cervantes A, Roda D, Tarazona N, Roselló S
and Pérez-Fidalgo JA: Current questions for the treatment of
advanced gastric cancer. Cancer Treat Rev. 39:60–67.
2013.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Wang X, Ouyang Y, Liu J, Zhu M, Zhao G,
Bao W and Hu FB: Fruit and vegetable consumption and mortality from
all causes, cardiovascular disease, and cancer: Systematic review
and dose-response meta-analysis of prospective cohort studies. BMJ.
349(g4490)2014.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Ranjan A, Ramachandran S, Gupta N, Kaushik
I, Wright S, Srivastava S, Das H, Srivastava S, Prasad S and
Srivastava SK: Role of phytochemicals in cancer prevention. Int J
Mol Sci. 20(4981)2019.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Chikara S, Nagaprashantha LD, Singhal J,
Horne D, Awasthi S and Singhal SS: Oxidative stress and dietary
phytochemicals: Role in cancer chemoprevention and treatment.
Cancer Lett. 413:122–134. 2018.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Ma L, Peng H, Li K, Zhao R, Li L, Yu Y,
Wang X and Han Z: Luteolin exerts an anticancer effect on NCI-H460
human non-small cell lung cancer cells through the induction of
Sirt1-mediated apoptosis. Mol Med Rep. 12:4196–4202.
2015.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Pu Y, Zhang T, Wang J, Mao Z, Duan B, Long
Y, Xue F, Liu D, Liu S and Gao Z: Luteolin exerts an anticancer
effect on gastric cancer cells through multiple signaling pathways
and regulating miRNAs. J Cancer. 9:3669–3675. 2018.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Zang MD, Hu L, Fan ZY, Wang HX, Zhu ZL,
Cao S, Wu XY, Li JF, Su LP, Li C, et al: Luteolin suppresses
gastric cancer progression by reversing epithelial-mesenchymal
transition via suppression of the Notch signaling pathway. J Transl
Med. 15(52)2017.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Bhatia R, Gautam SK, Cannon A, Thompson C,
Hall BR, Aithal A, Banerjee K, Jain M, Solheim JC, Kumar S, et al:
Cancer-associated mucins: Role in immune modulation and metastasis.
Cancer Metastasis Rev. 38:223–236. 2019.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Raina D, Kharbanda S and Kufe D: The MUC1
oncoprotein activates the anti-apoptotic phosphoinositide
3-kinase/Akt and Bcl-xL pathways in rat 3Y1 fibroblasts. J Biol
Chem. 279:20607–20612. 2004.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Raina D, Ahmad R, Kumar S, Ren J, Yoshida
K, Kharbanda S and Kufe D: MUC1 oncoprotein blocks nuclear
targeting of c-Abl in the apoptotic response to DNA damage. EMBO J.
25:3774–3783. 2006.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Agata N, Ahmad R, Kawano T, Raina D,
Kharbanda S and Kufe D: MUC1 oncoprotein blocks death
receptor-mediated apoptosis by inhibiting recruitment of caspase-8.
Cancer Res. 68:6136–6144. 2008.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Rahn JJ, Chow JW, Horne GJ, Mah BK,
Emerman JT, Hoffman P and Hugh JC: MUC1 mediates transendothelial
migration in vitro by ligating endothelial cell ICAM-1. Clin Exp
Metastasis. 22:475–483. 2005.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Lindén SK, Sheng YH, Every AL, Miles KM,
Skoog EC, Florin TH, Sutton P and McGuckin MA: MUC1 limits
Helicobacter pylori infection both by steric hindrance and
by acting as a releasable decoy. PLoS Pathog.
5(e1000617)2009.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Murphy G: The ADAMs: Signaling scissors in
the tumour microenvironment. Nat Rev Cancer. 8:929–941.
2008.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Ni SS, Zhang J, Zhao WL, Dong XC and Wang
JL: ADAM17 is overexpressed in non small cell lung cancer and its
expression correlates with poor patient survival. Tumour Biol.
34:1813–1818. 2013.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Zhao J and Tang J: The role of ADAM17 in
tumorigenesis and progression of breast cancer. Tumour Biol.
37:15359–15370. 2016.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Fang W, Qian J, Wu Q, Chen Y and Yu G:
ADAM 17 expression is enhanced by FoxM1 and is a poor prognostic
sign in gastric carcinoma. J Surg Res. 220:223–233. 2017.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Li W, Wang D, Sun X, Zhang Y, Wang L and
Suo J: ADAM17 promotes lymph node metastasis in gastric cancer via
activation of the Notch and Wnt signaling pathways. Int J Mol Med.
43:914–926. 2019.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Santos-Silva F, Fonseca A, Caffrey T,
Carvalho F, Mesquita P, Reis C, Almeida R, David L and
Hollingsworth MA: Thomsen-Friedenreich antigen expression in
gastric carcinomas is associated with MUC1 mucin VNTR polymorphism.
Glycobiology. 15:511–517. 2005.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Yu LG, Andrews N, Zhao Q, McKean D,
Williams JF, Connor LJ, Gerasimenko OV, Hilkens J, Hirabayashi J,
Kasai K, et al: Galectin-3 interaction with Thomsen-Friedenreich
disaccharide on cancer-associated MUC1 causes increased cancer cell
endothelial adhesion. J Biol Chem. 282:773–781. 2007.PubMed/NCBI View Article : Google Scholar
|
|
51
|
Mori Y, Akita K, Yashiro M, Sawada T,
Hirakawa K, Murata T and Nakada H: Binding of galectin-3, a
β-galactoside-binding lectin, to MUC1 protein enhances
phosphorylation of extracellular signal-regulated kinase 1/2
(ERK1/2) and Akt, promoting tumor cell malignancy. J Biol Chem.
290:26125–26140. 2015.PubMed/NCBI View Article : Google Scholar
|
|
52
|
Peek RM Jr, Fiske C and Wilson KT: Role of
innate immunity in Helicobacter pylori-induced gastric
malignancy. Physiol Rev. 90:831–858. 2010.PubMed/NCBI View Article : Google Scholar
|
|
53
|
Yamada S, Kato S, Matsuhisa T,
Makonkawkeyoon L, Yoshida M, Chakrabandhu T, Lertprasertsuk N,
Suttharat P, Chakrabandhu B, Nishiumi S, et al: Predominant mucosal
IL-8 mRNA expression in non-cagA Thais is risk for gastric cancer.
World J Gastroenterol. 19:2941–2949. 2013.PubMed/NCBI View Article : Google Scholar
|
|
54
|
Mumm JB and Oft M: Pegylated IL-10 induces
cancer immunity: The surprising role of IL-10 as a potent inducer
of IFN-γ-mediated CD8(+) T cell cytotoxicity. Bioessays.
35:623–631. 2013.PubMed/NCBI View Article : Google Scholar
|
|
55
|
Kim JA, Kim DK, Kang OH, Choi YA, Park HJ,
Choi SC, Kim TH, Yun KJ, Nah YH and Lee YM: Inhibitory effect of
luteolin on TNF-alpha-induced IL-8 production in human colon
epithelial cells. Int Immunopharmacol. 5:209–217. 2005.PubMed/NCBI View Article : Google Scholar
|
|
56
|
Nunes C, Almeida L, Barbosa RM and
Laranjinha J: Luteolin suppresses the JAK/STAT pathway in a
cellular model of intestinal inflammation. Food Funct. 8:387–396.
2017.PubMed/NCBI View Article : Google Scholar
|
|
57
|
Lee YJ, Lim T, Han MS, Lee SH, Baek SH,
Nan HY and Lee C: Anticancer effect of luteolin is mediated by
downregulation of TAM receptor tyrosine kinases, but not
interleukin-8, in non-small cell lung cancer cells. Oncol Rep.
37:1219–1226. 2017.PubMed/NCBI View Article : Google Scholar
|
|
58
|
Sokolova O and Naumann M: NF-κB signaling
in gastric cancer. Toxins (Basel). 9:1–22. 2017.PubMed/NCBI View Article : Google Scholar
|
|
59
|
Imran M, Rauf A, Abu-Izneid T, Nadeem M,
Shariati MA, Khan IA, Imran A, Orhan IE, Rizwan M, Atif M, et al:
Luteolin, a flavonoid, as an anticancer agent: A review. Biomed
Pharmacother. 112(108612)2019.PubMed/NCBI View Article : Google Scholar
|