Macrophages are distributed throughout the body in
various tissues and organs and show a high degree of heterogeneity
and diversity (1). Several specific
markers expressed on macrophage surfaces have been used to identify
different subsets, such as F4/80, CD68, SRA-1 and CD169(2). CD169+ macrophages are a
unique subset of macrophages distributed across multiple tissues
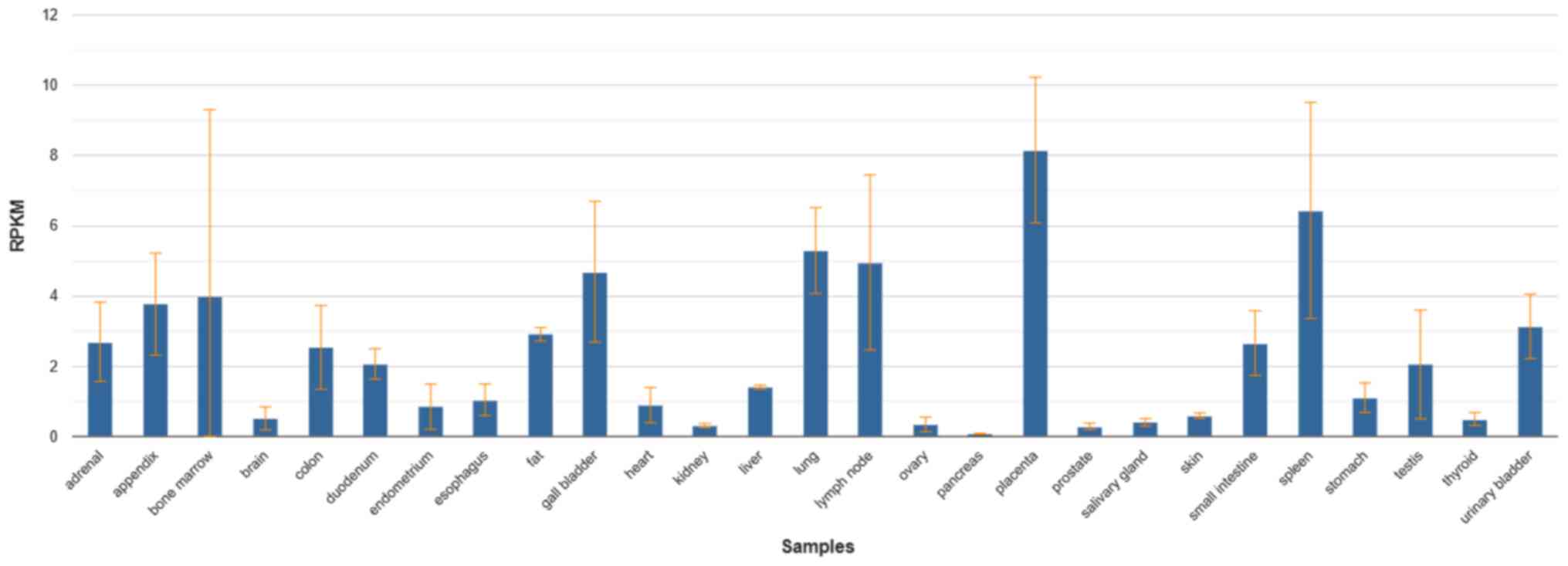
and organs of the human body. The results in the NCBI database
showed that the CD169 molecules were expressed in 27 different
tissues of the human body, such as the spleen, lymph node, small
intestine, liver, lung, heart, kidney, colon, bone marrow and
placenta, with a particularly high expression in the placenta,
spleen, lymph nodes, lungs and bone marrow. The expression of CD169
also changes in these organs when the organ becomes diseased
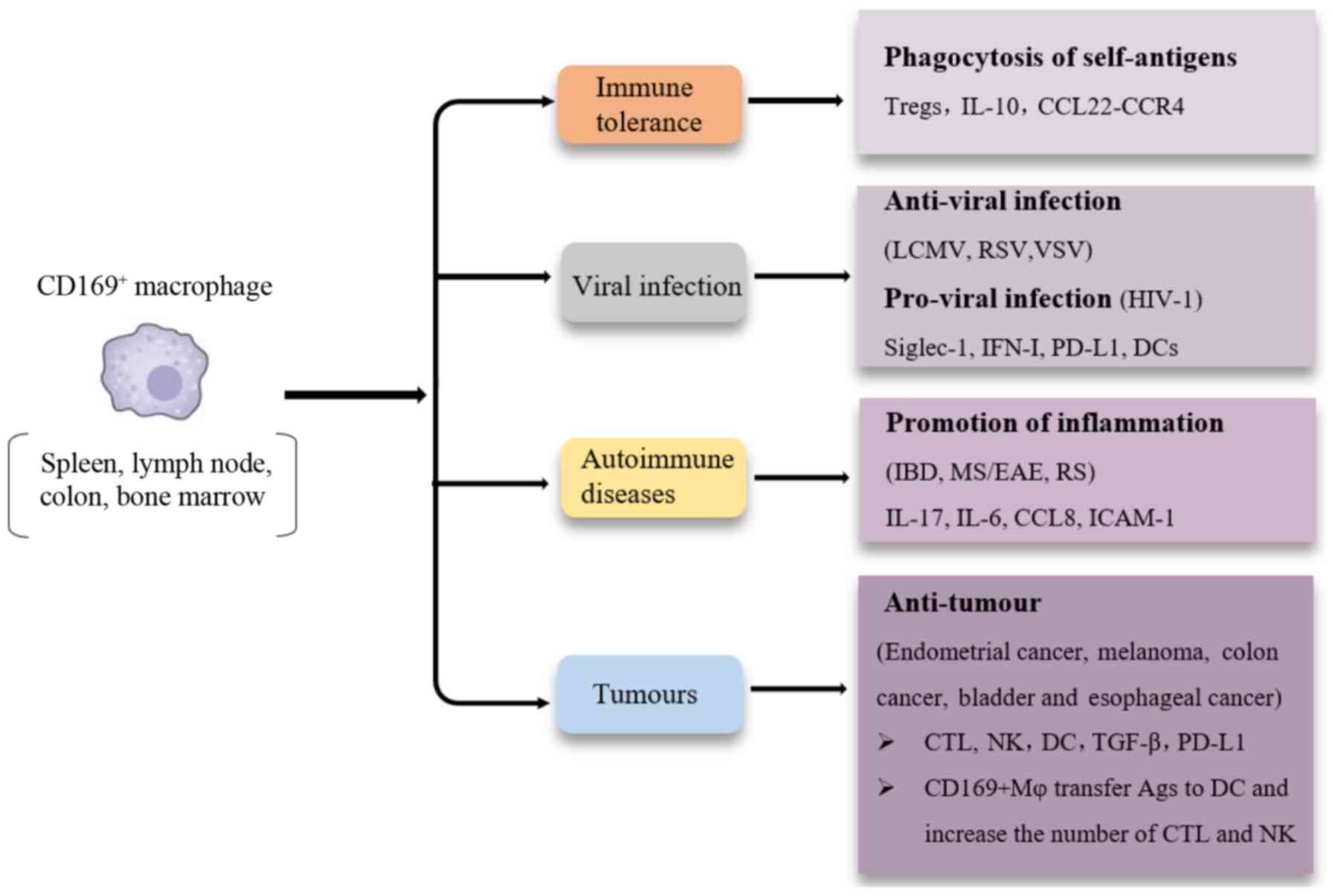
(Fig. 1) (3). Studies on CD169+ macrophages
show its unique roles in certain diseases. CD169+
macrophages exhibit a unique location distribution, primarily in
the secondary lymphoid organs where the blood and lymph enter and
leave, and express the unique CD169 molecule on their surface
(2). Unlike M1 and M2 macrophages,
CD169+ macrophages can interact directly with T cells, B
cells and dendritic cells (DC) through CD169 molecules to
participate in immune regulation (4).
Macrophages usually maintain immune homeostasis by
phagocytosis of foreign particles and production of
anti-inflammatory factors, such as IL-10(64). CD169+ macrophages do not
exhibit changes in mice lacking MyD88-mediated Toll receptor
signalling, and in mice in which bacterial flora have been
eradicated, despite the high levels of bacterial flora in the colon
and the importance of TLR signalling in mucosal homeostasis
(26,65). However, several studies have shown
that activated CD169+ macrophages are involved in
inflammatory responses during several autoimmune diseases. In the
colon, CD169+ macrophages are reported to promote
colitis progression in a dextran sulfate sodium (DSS)-induced IBD
model (66,67). The symptoms of colitis in DSS-induced
mice were significantly alleviated in CD169+
macrophage-deficient CD169-DTR mice. Our previous study showed that
the numbers of CD169+ macrophages in the mesenteric
lymph nodes (mLN) and abdominal cavity were higher in DSS-induced
colitis mice than in the WT mice, with higher expression of mLN
inflammatory cytokines, such as IL-17 and IL-6(68). At the same time, the expression
levels of the chemokine CCL22 decreased, together with decreased
CCR4-expressing Treg cells, which are critical factors for
maintaining homeostasis (63). A
further study revealed that CD169+ macrophages respond
to microbial antigens and produce CCL8 to recruit inflammatory
monocytes that exacerbate inflammation during colitis (68). Moreover, the decrease in
CD169+ macrophages in cyanidin 3-O-glucoside-treated
colitis suggests that this subset could be a potential biomarker
and therapeutic target (69).
In the past decade, the marginal zone of the mouse
spleen has been shown to serve a very important role in host
defence against pathogen infections, such as viruses (74,75).
CD169+ macrophages are reported to be the primary cell
type infected during viral infection, and they can capture viral
particles in the blood, absorb antigens, such as immune complexes
and viruses, and then present them in a complete form to follicular
B cells, inducing germinal centre B cellular responses (30,76,77).
CD169+ macrophages transfer antigens to CD8a+
DCs using the CD169 molecule, which preferentially participate in
cell contact, eventually inducing an effective CD8+ T
cell response (51). Moreover,
CD169+ macrophages have been shown to enforce viral
replication, resulting in the delivery of a large number of viral
antigens, and the amplification of T and B lymphocyte responses
(29,78). Type I IFN induced macrophages express
CD169 molecules both in vivo and in vitro (50). CD169+ macrophages can also
mediate antiviral activity by secreting type I IFN during viral
infection. Since CD169+ macrophages simultaneously
express programmed death ligands (PD-L1), the expression of IFN-I
can upregulate the expression of PD-L1, which may result in
CD8+ T cell exhaustion. The exhaustion of
CD8+ T cells is a double-edged sword. In studies of
lymphocytic choroidal meningitis virus infection in vivo,
the persistent expression of IFN-I resulted in increased IL-10 and
PD-L1 levels (79). IFN-I produced
by CD169+ macrophages during chronic infection inhibits
activation of the immune response to secondary infection (80). However, the absence of
CD169+ macrophages results in inadequate production of
IFN-I, reducing antiviral activity and persistence of the virus in
the human body. Deletion of CD169+ macrophages also
limits IFN-1 dependent PD-L1 expression. Without PD-L1, viral
replication is enhanced, and the virus persists. At the same time,
CD8+ T cell depletion is inhibited. Thus, in a mouse
model, PD-L1 deletion resulted in the development of severe
immunopathology and they died quickly following infection (79).
Similarly, mice infected with respiratory syncytial
virus (RSV) also showed that the number of CD169+
macrophages localized in the alveoli increased significantly
(81). CD169-diphtheria toxin
receptor (DTR) mice revealed that the secretion of IFN-β, IL-6 and
TNF-α decreased when CD169+ macrophages were absent,
whereas CD169+ macrophage deletion reduced the
aggregation of effector CD8+ T cells to the lungs
following RSV mucosal infection. Overall, regulating the number of
CD169+ macrophages to enhance the immune response to RSV
infection may be a novel therapeutic strategy (82).
Some studies of retroviral HIV revealed that the
expression of CD169 induced by IFN-I could promote cis-infection in
bone marrow cells and target HIV to DC-mediated trans-infection
pathways (82-86).
Siglec-1 on the surface of CD169+ macrophages can
recognize gangliosides in the lipid membrane of the virus, capture
HIV particles, and further transmit the viral signal to DCs,
leading to the infection of CD4+ T cells and reducing
the antiviral effect of IFN-I (87,88).
Moreover, Siglec-1 induces the formation of a virus-containing
compartment and enhances macrophage-to-T cell transmission of
HIV-1(83). Siglec-1 expression on
pre-DCs amongst blood DCs promotes attachment and fusion of viral
particles and mediates the replication-independent transfer of
HIV-1 to activated primary T lymphocytes (20). Whether CD169+ macrophages
serve an antiviral role or promote viral replication during viral
infection is dependent on the genetic characteristics of the virus
and the location of CD169+ macrophages. Altogether, the
roles and mechanisms of CD169+ macrophages in humans
infected with viruses still requires further study.
The production of cytotoxic T lymphocytes (CTLs) in
tumour targeting CTLs is considered to be key in inducing
antitumour immunity (89,90). A previous study reported that CTLs
and NK cells were activated by the subcutaneous injection of
apoptotic tumour cells and they exhibited anti-tumour immunity
effects (91). Antigen-presenting
cells are critical for the activation of CTLs by capturing tumour
cell-related antigens, which are primarily released from apoptotic
tumour cells (92).
CD169+ macrophages in lymph nodes and spleen were
reported to present apoptotic tumour antigens. Additionally,
intravenous injection of apoptotic tumour cells may be different
from those obtained by subcutaneous injection of apoptotic tumour
cells (93). Furthermore, tumour
antigen-specific CD8+ T cell activation and subsequent
anti-tumour immune function in CD169+
macrophage-deficient mice was severely impaired (13,59).
Not applicable.
This work was supported by The Natural Science
Foundation of Shandong Province (grant no. ZR2017MH003), and
supported in part by the National Key Research and Development
Program of China, the Ministry of Science and Technology (grant no.
2016YFE0127000).
Not applicable.
YL, YX and CHQ wrote the manuscript. CHQ edited the
manuscript. All authors read and approved the final manuscript.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
|
1
|
Varol C, Mildner A and Jung S:
Macrophages: Development and tissue specialization. Annu Rev
Immunol. 33:643–675. 2015.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Martinez-Pomares L and Gordon S:
CD169+ macrophages at the crossroads of antigen
presentation. Trends Immunol. 33:66–70. 2012.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Fagerberg L, Hallström BM, Oksvold P,
Kampf C, Djureinovic D, Odeberg J, Habuka M, Tahmasebpoor S,
Danielsson A, Edlund K, et al: Analysis of the human
tissue-specific expression by genome-wide integration of
transcriptomics and antibody-based proteomics. Mol Cell Proteomics.
13:397–406. 2014.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Chávez-Galán L, Olleros ML, Vesin D and
Garcia I: Much more than M1 and M2 macrophages, there are also
CD169(+) and TCR(+) macrophages. Front Immunol.
6(263)2015.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Li W, Wang Y, Zhao H, Zhang H, Xu Y, Wang
S, Guo X, Huang Y, Zhang S, Han Y, et al: Identification and
transcriptome analysis of erythroblastic island macrophages. Blood.
134:480–491. 2019.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Komohara Y, Ohnishi K and Takeya M:
Possible functions of CD169-positive sinus macrophages in lymph
nodes in anti-tumor immune responses. Cancer Sci. 108:290–295.
2017.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Bao G, Han Z, Yan Z, Wang Q, Zhou Y, Yao
D, Gu M, Chen B, Chen S, Deng A and Zhong R: Increased Siglec-1
expression in monocytes of patients with primary biliary cirrhosis.
Immunol Invest. 39:645–660. 2010.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Strömvall K, Sundkvist K, Ljungberg B,
Halin Bergström S and Bergh A: Reduced number of CD169(+)
macrophages in pre-metastatic regional lymph nodes is associated
with subsequent metastatic disease in an animal model and with poor
outcome in prostate cancer patients. Prostate. 77:1468–1477.
2017.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Biesen R, Demir C, Barkhudarova F, Grün
JR, Steinbrich-Zöllner M, Backhaus M, Häupl T, Rudwaleit M,
Riemekasten G, Radbruch A, et al: Sialic acid-binding Ig-like
lectin 1 expression in inflammatory and resident monocytes is a
potential biomarker for monitoring disease activity and success of
therapy in systemic lupus erythematosus. Arthritis Rheum.
58:1136–1145. 2008.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Zhang J, Xu J, Zhang RX, Zhang Y, Ou QJ,
Li JQ, Jiang ZZ, Wu XJ, Fang YJ and Zheng L: CD169 identifies an
activated CD8(+) T cell subset in regional lymph nodes that
predicts favorable prognosis in colorectal cancer patients.
Oncoimmunology. 5(e1177690)2016.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Nycholat CM, Rademacher C, Kawasaki N and
Paulson JC: In silico-aided design of a glycan ligand of
sialoadhesin for in vivo targeting of macrophages. J Am Chem Soc.
134:15696–15699. 2012.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Oetke C, Vinson MC, Jones C and Crocker
PR: Sialoadhesin-deficient mice exhibit subtle changes in B- and
T-cell populations and reduced immunoglobulin M levels. Mol Cell
Biol. 26:1549–1557. 2006.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Edgar LJ, Kawasaki N, Nycholat CM and
Paulson JC: Targeted delivery of antigen to activated CD169(+)
macrophages induces bias for expansion of CD8(+) T cells. Cell Chem
Biol. 26:131–136.e4. 2019.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Ravishankar B, Shinde R, Liu H, Chaudhary
K, Bradley J, Lemos HP, Chandler P, Tanaka M, Munn DH, Mellor AL
and McGaha TL: Marginal zone CD169+ macrophages
coordinate apoptotic cell-driven cellular recruitment and
tolerance. Proc Natl Acad Sci USA. 111:4215–4220. 2014.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Panduro M, Benoist C and Mathis D: Tissue
tregs. Annu Rev Immunol. 34:609–633. 2016.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Sakaguchi S, Yamaguchi T, Nomura T and Ono
M: Regulatory T cells and immune tolerance. Cell. 133:775–787.
2008.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Sakaguchi S, Ono M, Setoguchi R, Yagi H,
Hori S, Fehervari Z, Shimizu J, Takahashi T and Nomura T:
Foxp3+ CD25+ CD4+ natural
regulatory T cells in dominant self-tolerance and autoimmune
disease. Immunol Rev. 212:8–27. 2006.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Wu C, Rauch U, Korpos E, Song J, Loser K,
Crocker PR and Sorokin LM: Sialoadhesin-positive macrophages bind
regulatory T cells, negatively controlling their expansion and
autoimmune disease progression. J Immunol. 182:6508–6516.
2009.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Ramos-Leví AM and Marazuela M:
Pathogenesis of thyroid autoimmune disease: The role of cellular
mechanisms. Endocrinol Nutr. 63:421–429. 2016.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Hashimoto K, Nishihara E, Matsumoto M,
Matsumoto S, Nakajima Y, Tsujimoto K, Yamakage H, Satoh-Asahara N,
Noh JY, Ito K, et al: Sialic acid-binding immunoglobulin-like
lectin1 as a novel predictive biomarker for relapse in Graves'
disease: A multicenter study. Thyroid. 28:50–59. 2018.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Ruffin N, Gea-Mallorquí E, Brouiller F,
Jouve M, Silvin A, See P, Dutertre CA, Ginhoux F and Benaroch P:
Constitutive Siglec-1 expression confers susceptibility to HIV-1
infection of human dendritic cell precursors. Proc Natl Acad Sci
USA. 116:21685–21693. 2019.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Fraschilla I and Pillai S: Viewing Siglecs
through the lens of tumor immunology. Immunol Rev. 276:178–191.
2017.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Shiota T, Miyasato Y, Ohnishi K,
Yamamoto-Ibusuki M, Yamamoto Y, Iwase H, Takeya M and Komohara Y:
The clinical significance of CD169-positive lymph node macrophage
in patients with breast cancer. PLoS One.
11(e0166680)2016.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Saunderson SC, Dunn AC, Crocker PR and
McLellan AD: CD169 mediates the capture of exosomes in spleen and
lymph node. Blood. 123:208–216. 2014.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Camara A, Cordeiro OG, Alloush F, Sponsel
J, Chypre M, Onder L, Asano K, Tanaka M, Yagita H, Ludewig B, et
al: Lymph node mesenchymal and endothelial stromal cells cooperate
via the RANK-RANKL cytokine axis to shape the sinusoidal macrophage
niche. Immunity. 50:1467–1481. 2019.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Hiemstra IH, Beijer MR, Veninga H,
Vrijland K, Borg EG, Olivier BJ, Mebius RE, Kraal G and den Haan
JM: The identification and developmental requirements of colonic
CD169+ macrophages. Immunology. 142:269–278. 2014.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Lescoat A, Ballerie A, Augagneur Y,
Morzadec C, Vernhet L, Fardel O, Jégo P, Jouneau S and Lecureur V:
Distinct properties of human M-CSF and GM-CSF monocyte-derived
macrophages to simulate pathological lung conditions in vitro:
Application to systemic and inflammatory disorders with pulmonary
involvement. Int J Mol Sci. 19(894)2018.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Vance J, Santos A, Sadofsky L, Morice A
and Cervantes J: Effect of high glucose on human alveolar
macrophage phenotype and phagocytosis of mycobacteria. Lung.
197:89–94. 2019.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Friedrich SK, Lang PA, Friebus-Kardash J,
Duhan V, Bezgovsek J and Lang KS: Mechanisms of lymphatic
system-specific viral replication and its potential role in
autoimmune disease. Clin Exp Immunol. 195:64–73. 2019.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Xu HC, Huang J, Khairnar V, Duhan V,
Pandyra AA, Grusdat M, Shinde P, McIlwain DR, Maney SK, Gommerman
J, et al: Deficiency of the B cell-activating factor receptor
results in limited CD169+ macrophage function during
viral infection. J Virol. 89:4748–4759. 2015.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Kikuchi K, Iida M, Ikeda N, Moriyama S,
Hamada M, Takahashi S, Kitamura H, Watanabe T, Hasegawa Y, Hase K,
et al: Macrophages switch their phenotype by regulating Maf
expression during different phases of inflammation. J Immunol.
201:635–651. 2018.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Yao H, Zhang Y, Xie B, Shang Y, Yuan S and
Zhang J: Sleep-restriction inhibits neurogenesis through decreasing
the infiltration of CD169(+) macrophages to ischemic brain after
stroke. Neuroscience. 431:222–236. 2020.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Spaulding E, Fooksman D, Moore JM, Saidi
A, Feintuch CM, Reizis B, Chorro L, Daily J and Lauvau G:
STING-licensed macrophages prime type I IFN production by
plasmacytoid dendritic cells in the bone marrow during severe
plasmodium yoelii malaria. PLoS Pathog. 12(e1005975)2016.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Chavez M, Silvestrini MT, Ingham ES, Fite
BZ, Mahakian LM, Tam SM, Ilovitsh A, Monjazeb AM, Murphy WJ,
Hubbard NE, et al: Distinct immune signatures in directly treated
and distant tumors result from TLR adjuvants and focal ablation.
Theranostics. 8:3611–3628. 2018.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Ferreira RC, Guo H, Coulson RM, Smyth DJ,
Pekalski ML, Burren OS, Cutler AJ, Doecke JD, Flint S, McKinney EF,
et al: A type I interferon transcriptional signature precedes
autoimmunity in children genetically at risk for type 1 diabetes.
Diabetes. 63:2538–2550. 2014.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Rose T, Szelinski F, Lisney A, Reiter K,
Fleischer SJ, Burmester GR, Radbruch A, Hiepe F, Grützkau A, Biesen
R and Dörner T: Siglec1 is a biomarker of disease activity and
indicates extraglandular manifestation in primary Sjögren's
syndrome. RMD Open. 2(e000292)2016.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Seu KG, Papoin J, Fessler R, Hom J, Huang
G, Mohandas N, Blanc L and Kalfa TA: Unraveling macrophage
heterogeneity in erythroblastic islands. Front Immunol.
8(1140)2017.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Falchi M, Varricchio L, Martelli F,
Masiello F, Federici G, Zingariello M, Girelli G, Whitsett C,
Petricoin EF III, Moestrup SK, et al: Dexamethasone targeted
directly to macrophages induces macrophage niches that promote
erythroid expansion. Haematologica. 100:178–187. 2015.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Jacobsen RN, Forristal CE, Raggatt LJ,
Nowlan B, Barbier V, Kaur S, van Rooijen N, Winkler IG, Pettit AR
and Levesque JP: Mobilization with granulocyte colony-stimulating
factor blocks medullar erythropoiesis by depleting
F4/80(+)VCAM1(+)CD169(+)ER-HR3(+)Ly6G(+) erythroid island
macrophages in the mouse. Exp Hematol. 42:547–561.e4.
2014.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Chow A, Huggins M, Ahmed J, Hashimoto D,
Lucas D, Kunisaki Y, Pinho S, Leboeuf M, Noizat C, van Rooijen N,
et al: CD169+ macrophages provide a niche promoting
erythropoiesis under homeostasis and stress. Nat Med. 19:429–436.
2013.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Kaur S, Raggatt LJ, Millard SM, Wu AC,
Batoon L, Jacobsen RN, Winkler IG, MacDonald KP, Perkins AC, Hume
DA, et al: Self-repopulating recipient bone marrow resident
macrophages promote long-term hematopoietic stem cell engraftment.
Blood. 132:735–749. 2018.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Gbotosho OT, Kapetanaki MG, Ross M, Ghosh
S, Weidert F, Bullock GC, Watkins S, Ofori-Acquah SF and Kato GJ:
Nrf2 deficiency in mice attenuates erythropoietic stress-related
macrophage hypercellularity. Exp Hematol. 84:19–28.e4.
2020.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Zhang RR and Zhu XF: Relationship between
macrophages and erythropoiesis. Zhongguo Dang Dai Er Ke Za Zhi.
18:94–99. 2016.PubMed/NCBI View Article : Google Scholar : (In Chinese).
|
|
44
|
Batoon L, Millard SM, Wullschleger ME,
Preda C, Wu AC, Kaur S, Tseng HW, Hume DA, Levesque JP, Raggatt LJ
and Pettit AR: CD169(+) macrophages are critical for osteoblast
maintenance and promote intramembranous and endochondral
ossification during bone repair. Biomaterials. 196:51–66.
2019.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Asano K, Kikuchi K and Tanaka M: CD169
macrophages regulate immune responses toward particulate materials
in the circulating fluid. J Biochem. 164:77–85. 2018.PubMed/NCBI View Article : Google Scholar
|
|
46
|
De Schryver M, Leemans A, Pintelon I,
Cappoen D, Maes L, Caljon G, Cos P and Delputte PL: Comparative
analysis of the internalization of the macrophage receptor
sialoadhesin in human and mouse primary macrophages and cell lines.
Immunobiology. 222:797–806. 2017.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Louie DAP and Liao S: Lymph Node
Subcapsular sinus macrophages as the frontline of lymphatic immune
defense. Front Immunol. 10(347)2019.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Heath WR, Kato Y, Steiner TM and Caminschi
I: Antigen presentation by dendritic cells for B cell activation.
Curr Opin Immunol. 58:44–52. 2019.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Veninga H, Borg EG, Vreeman K, Taylor PR,
Kalay H, van Kooyk Y, Kraal G, Martinez-Pomares L and den Haan JM:
Antigen targeting reveals splenic CD169+ macrophages as
promoters of germinal center B-cell responses. Eur J Immunol.
45:747–757. 2015.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Grabowska J, Lopez-Venegas MA, Affandi AJ
and den-Haan JMM: CD169+ macrophages capture and
Dendritic cells instruct: The interplay of the gatekeeper and the
general of the immune system. Front Immunol. 9(2472)2018.PubMed/NCBI View Article : Google Scholar
|
|
51
|
Van Dinther D, Veninga H, Iborra S, Borg
EGF, Hoogterp L, Olesek K, Beijer MR, Schetters STT, Kalay H,
Garcia-Vallejo JJ, et al: Functional CD169 on macrophages mediates
interaction with Dendritic cells for CD8(+) T Cell cross-priming.
Cell Rep. 22:1484–1495. 2018.PubMed/NCBI View Article : Google Scholar
|
|
52
|
Barral P, Polzella P, Bruckbauer A, van
Rooijen N, Besra GS, Cerundolo V and Batista FD: CD169(+)
macrophages present lipid antigens to mediate early activation of
iNKT cells in lymph nodes. Nat Immunol. 11:303–312. 2010.PubMed/NCBI View Article : Google Scholar
|
|
53
|
Kawasaki N, Vela JL, Nycholat CM,
Rademacher C, Khurana A, van Rooijen N, Crocker PR, Kronenberg M
and Paulson JC: Targeted delivery of lipid antigen to macrophages
via the CD169/sialoadhesin endocytic pathway induces robust
invariant natural killer T cell activation. Proc Natl Acad Sci USA.
110:7826–7831. 2013.PubMed/NCBI View Article : Google Scholar
|
|
54
|
Covarrubias R, Wilhelm AJ and Major AS:
Specific deletion of LDL receptor-related protein on macrophages
has skewed in vivo effects on cytokine production by invariant
natural killer T cells. PLoS One. 9(e102236)2014.PubMed/NCBI View Article : Google Scholar
|
|
55
|
Trahtemberg U and Mevorach D: Apoptotic
cells induced signaling for immune homeostasis in macrophages and
Dendritic cells. Front Immunol. 8(1356)2017.PubMed/NCBI View Article : Google Scholar
|
|
56
|
Vives-Pi M, Rodríguez-Fernández S and
Pujol-Autonell I: How apoptotic β-cells direct immune response to
tolerance or to autoimmune diabetes: A review. Apoptosis.
20:263–272. 2015.PubMed/NCBI View Article : Google Scholar
|
|
57
|
Tanaka M and Miyake Y: Apoptotic cell
clearance and autoimmune disorder. Curr Med Chem. 14:2892–2897.
2007.PubMed/NCBI View Article : Google Scholar
|
|
58
|
Miyake Y, Asano K, Kaise H, Uemura M,
Nakayama M and Tanaka M: Critical role of macrophages in the
marginal zone in the suppression of immune responses to apoptotic
cell-associated antigens. J Clin Invest. 117:2268–2278.
2007.PubMed/NCBI View Article : Google Scholar
|
|
59
|
Asano K, Nabeyama A, Miyake Y, Qiu CH,
Kurita A, Tomura M, Kanagawa O, Fujii S and Tanaka M:
CD169-positive macrophages dominate antitumor immunity by
crosspresenting dead cell-associated antigens. Immunity. 34:85–95.
2011.PubMed/NCBI View Article : Google Scholar
|
|
60
|
Ravishankar B, Liu H, Shinde R, Chandler
P, Baban B, Tanaka M, Munn DH, Mellor AL, Karlsson MC and McGaha
TL: Tolerance to apoptotic cells is regulated by indoleamine
2,3-dioxygenase. Proc Natl Acad Sci USA. 109:3909–3914.
2012.PubMed/NCBI View Article : Google Scholar
|
|
61
|
Black LV, Saunderson SC, Coutinho FP,
Muhsin-Sharafaldine MR, Damani TT, Dunn AC and McLellan AD: The
CD169 sialoadhesin molecule mediates cytotoxic T-cell responses to
tumour apoptotic vesicles. Immunol Cell Biol. 94:430–438.
2016.PubMed/NCBI View Article : Google Scholar
|
|
62
|
Qiu CH, Miyake Y, Kaise H, Kitamura H,
Ohara O and Tanaka M: Novel subset of CD8{alpha}+ dendritic cells
localized in the marginal zone is responsible for tolerance to
cell-associated antigens. J Immunol. 182:4127–4136. 2009.PubMed/NCBI View Article : Google Scholar
|
|
63
|
Hao S, Han X, Wang D, Yang Y, Li Q, Li X
and Qiu CH: Critical role of CCL22/CCR4 axis in the maintenance of
immune homeostasis during apoptotic cell clearance by splenic
CD8α(+) CD103(+) dendritic cells. Immunology. 148:174–186.
2016.PubMed/NCBI View Article : Google Scholar
|
|
64
|
Shapouri-Moghaddam A, Mohammadian S,
Vazini H, Taghadosi M, Esmaeili SA, Mardani F, Seifi B, Mohammadi
A, Afshari JT and Sahebkar A: Macrophage plasticity, polarization,
and function in health and disease. J Cell Physiol. 233:6425–6440.
2018.PubMed/NCBI View Article : Google Scholar
|
|
65
|
Detienne S, Welsby I, Collignon C, Wouters
S, Coccia M, Delhaye S, Van Maele L, Thomas S, Swertvaegher M,
Detavernier A, et al: Central role of CD169(+) lymph node resident
macrophages in the adjuvanticity of the QS-21 component of AS01.
Sci Rep. 6(39475)2016.PubMed/NCBI View Article : Google Scholar
|
|
66
|
Wang D, Li Q, Yang Y, Hao S, Han X, Song
J, Yin Y, Li X, Tanaka M and Qiu CH: Macrophage subset expressing
CD169 in peritoneal cavity-regulated mucosal inflammation together
with lower levels of CCL22. Inflammation. 40:1191–1203.
2017.PubMed/NCBI View Article : Google Scholar
|
|
67
|
Li Q, Wang D, Hao S, Han X, Xia Y, Li X,
Chen Y, Tanaka M and Qiu CH: CD169 expressing macrophage, a key
subset in mesenteric lymph nodes promotes mucosal inflammation in
dextran sulfate sodium-induced colitis. Front Immunol.
8(669)2017.PubMed/NCBI View Article : Google Scholar
|
|
68
|
Asano K, Takahashi N, Ushiki M, Monya M,
Aihara F, Kuboki E, Moriyama S, Iida M, Kitamura H, Qiu CH, et al:
Intestinal CD169(+) macrophages initiate mucosal inflammation by
secreting CCL8 that recruits inflammatory monocytes. Nat Commun.
6(7802)2015.PubMed/NCBI View Article : Google Scholar
|
|
69
|
Xia Y, Tian LM, Liu Y, Guo KS, Lv M, Li
QT, Hao SY, Ma CH, Chen YX, Tanaka M, et al: Low dose of
cyanidin-3-O-glucoside alleviated dextran sulfate sodium-induced
colitis, mediated by CD169+ macrophage pathway. Inflamm Bowel Dis.
25:1510–1521. 2019.PubMed/NCBI View Article : Google Scholar
|
|
70
|
Bogie JF, Boelen E, Louagie E, Delputte P,
Elewaut D, van Horssen J, Hendriks JJ and Hellings N: CD169 is a
marker for highly pathogenic phagocytes in multiple sclerosis. Mult
Scle. 24:290–300. 2018.PubMed/NCBI View Article : Google Scholar
|
|
71
|
Xiong YS, Cheng Y, Lin QS, Wu AL, Yu J, Li
C, Sun Y, Zhong RQ and Wu LJ: Increased expression of Siglec-1 on
peripheral blood monocytes and its role in mononuclear cell
reactivity to autoantigen in rheumatoid arthritis. Rheumatology
(Oxford). 53:250–259. 2014.PubMed/NCBI View Article : Google Scholar
|
|
72
|
Guo X, Nakamura K, Kohyama K, Harada C,
Behanna HA, Watterson DM, Matsumoto Y and Harada T: Inhibition of
glial cell activation ameliorates the severity of experimental
autoimmune encephalomyelitis. Neurosci Res. 59:457–466.
2007.PubMed/NCBI View Article : Google Scholar
|
|
73
|
Karasawa K, Asano K, Moriyama S, Ushiki M,
Monya M, Iida M, Kuboki E, Yagita H, Uchida K, Nitta K and Tanaka
M: Vascular-resident CD169-positive monocytes and macrophages
control neutrophil accumulation in the kidney with
ischemia-reperfusion injury. J Am Soc Nephrol. 26:896–906.
2015.PubMed/NCBI View Article : Google Scholar
|
|
74
|
Shinde PV, Xu HC, Maney SK, Kloetgen A,
Namineni S, Zhuang Y, Honke N, Shaabani N, Bellora N, Doerrenberg
M, et al: Tumor necrosis factor-mediated survival of CD169(+) cells
promotes immune activation during vesicular stomatitis virus
infection. J Virol. 92:e01637–e01617. 2018.PubMed/NCBI View Article : Google Scholar
|
|
75
|
Uchil PD, Pi R, Haugh KA, Ladinsky MS,
Ventura JD, Barrett BS, Santiago ML, Bjorkman PJ, Kassiotis G,
Sewald X and Mothes W: A protective role for the lectin
CD169/Siglec-1 against a pathogenic murine retrovirus. Cell Host
Microbe. 25:87–100.e10. 2019.PubMed/NCBI View Article : Google Scholar
|
|
76
|
Frederico B, Chao B, Lawler C, May JS and
Stevenson PG: Subcapsular sinus macrophages limit acute
gammaherpesvirus dissemination. J Gen Virol. 96:2314–2327.
2015.PubMed/NCBI View Article : Google Scholar
|
|
77
|
Junt T, Moseman EA, Iannacone M, Massberg
S, Lang PA, Boes M, Fink K, Henrickson SE, Shayakhmetov DM, Di
Paolo NC, et al: Subcapsular sinus macrophages in lymph nodes clear
lymph-borne viruses and present them to antiviral B cells. Nature.
450:110–114. 2007.PubMed/NCBI View Article : Google Scholar
|
|
78
|
Honke N, Shaabani N, Merches K, Gassa A,
Kraft A, Ehrhardt K, Häussinger D, Löhning M, Dittmer U, Hengel H,
et al: Immunoactivation induced by chronic viral infection inhibits
viral replication and drives immunosuppression through sustained
IFN-I responses. Eur J Immunol. 46:372–380. 2016.PubMed/NCBI View Article : Google Scholar
|
|
79
|
Shaabani N, Duhan V, Khairnar V, Gassa A,
Ferrer-Tur R, Häussinger D, Recher M, Zelinskyy G, Liu J, Dittmer
U, et al: CD169(+) macrophages regulate PD-L1 expression via type I
interferon and thereby prevent severe immunopathology after LCMV
infection. Cell Death Dis. 7(e2446)2016.PubMed/NCBI View Article : Google Scholar
|
|
80
|
Teijaro JR: Too much of a good thing:
Sustained type 1 interferon signaling limits humoral responses to
secondary viral infection. Eur J Immunol. 46:300–302.
2016.PubMed/NCBI View Article : Google Scholar
|
|
81
|
Oh DS, Oh JE, Jung HE and Lee HK:
Transient depletion of CD169(+) cells contributes to impaired early
protection and effector CD8(+) T cell recruitment against mucosal
respiratory syncytial virus infection. Front Immunol.
8(819)2017.PubMed/NCBI View Article : Google Scholar
|
|
82
|
Jans J, Unger WWJ, Vissers M, Ahout IML,
Schreurs I, Wickenhagen A, de Groot R, de Jonge MI and Ferwerda G:
Siglec-1 inhibits RSV-induced interferon gamma production by adult
T cells in contrast to newborn T cells. Eur J Immunol. 48:621–631.
2018.PubMed/NCBI View Article : Google Scholar
|
|
83
|
Hammonds JE, Beeman N, Ding L, Takushi S,
Francis AC, Wang JJ, Melikyan GB and Spearman P: Siglec-1 initiates
formation of the virus-containing compartment and enhances
macrophage-to-T cell transmission of HIV-1. PLoS Pathog.
13(e1006181)2017.PubMed/NCBI View Article : Google Scholar
|
|
84
|
Jobe O, Kim J and Rao M: The role of
Siglec-1 in HIV-1/macrophage interaction. Macrophage (Houst).
3(e1435)2016.PubMed/NCBI
|
|
85
|
Pino M, Erkizia I, Benet S, Erikson E,
Fernández-Figueras MT, Guerrero D, Dalmau J, Ouchi D, Rausell A,
Ciuffi A, et al: HIV-1 immune activation induces Siglec-1
expression and enhances viral trans-infection in blood and tissue
myeloid cells. Retrovirology. 12(37)2015.PubMed/NCBI View Article : Google Scholar
|
|
86
|
Martinez-Picado J, McLaren PJ, Erkizia I,
Martin MP, Benet S, Rotger M, Dalmau J, Ouchi D, Wolinsky SM,
Penugonda S, et al: Identification of Siglec-1 null individuals
infected with HIV-1. Nat Commun. 7(12412)2016.PubMed/NCBI View Article : Google Scholar
|
|
87
|
Akiyama H, Ramirez NP, Gibson G, Kline C,
Watkins S, Ambrose Z and Gummuluru S: Interferon-inducible
CD169/Siglec1 attenuates anti-HIV-1 effects of alpha interferon. J
Virol. 91:e00972–e00917. 2017.PubMed/NCBI View Article : Google Scholar
|
|
88
|
Yu X, Feizpour A, Ramirez NG, Wu L,
Akiyama H, Xu F, Gummuluru S and Reinhard BM:
Glycosphingolipid-functionalized nanoparticles recapitulate
CD169-dependent HIV-1 uptake and trafficking in dendritic cells.
Nat Commun. 5(4136)2014.PubMed/NCBI View Article : Google Scholar
|
|
89
|
Farhood B, Najafi M and Mortezaee K:
CD8(+) cytotoxic T lymphocytes in cancer immunotherapy: A review. J
Cell Physiol. 234:8509–8521. 2019.PubMed/NCBI View Article : Google Scholar
|
|
90
|
Joyce JA and Fearon DT: T cell exclusion,
immune privilege, and the tumor microenvironment. Science.
348:74–80. 2015.PubMed/NCBI View Article : Google Scholar
|
|
91
|
Schnurr M, Scholz C, Rothenfusser S,
Galambos P, Dauer M, Röbe J, Endres S and Eigler A: Apoptotic
pancreatic tumor cells are superior to cell lysates in promoting
cross-priming of cytotoxic T cells and activate NK and gammadelta T
cells. Cancer Res. 62:2347–2352. 2002.PubMed/NCBI
|
|
92
|
Jenne L, Arrighi JF, Jonuleit H, Saurat JH
and Hauser C: Dendritic cells containing apoptotic melanoma cells
prime human CD8+ T cells for efficient tumor cell lysis. Cancer
Res. 60:4446–4452. 2000.PubMed/NCBI
|
|
93
|
Van Dinther D, Veninga H, Revet M,
Hoogterp L, Olesek K, Grabowska J, Borg EGF, Kalay H, van Kooyk Y
and den Haan JMM: Comparison of protein and peptide targeting for
the development of a CD169-based vaccination strategy against
melanoma. Front Immunol. 9(1997)2018.PubMed/NCBI View Article : Google Scholar
|
|
94
|
Asano T, Ohnishi K, Shiota T, Motoshima T,
Sugiyama Y, Yatsuda J, Kamba T, Ishizaka K and Komohara Y:
CD169-positive sinus macrophages in the lymph nodes determine
bladder cancer prognosis. Cancer Sci. 109:1723–1730.
2018.PubMed/NCBI View Article : Google Scholar
|
|
95
|
Takeya H, Shiota T, Yagi T, Ohnishi K,
Baba Y, Miyasato Y, Kiyozumi Y, Yoshida N, Takeya M, Baba H and
Komohara Y: High CD169 expression in lymph node macrophages
predicts a favorable clinical course in patients with esophageal
cancer. Pathol Int. 68:685–693. 2018.PubMed/NCBI View Article : Google Scholar
|
|
96
|
Ohnishi K, Yamaguchi M, Erdenebaatar C,
Saito F, Tashiro H, Katabuchi H, Takeya M and Komohara Y:
Prognostic significance of CD169-positive lymph node sinus
macrophages in patients with endometrial carcinoma. Cancer Sci.
107:846–852. 2016.PubMed/NCBI View Article : Google Scholar
|
|
97
|
Ohnishi K, Komohara Y, Saito Y, Miyamoto
Y, Watanabe M, Baba H and Takeya M: CD169-positive macrophages in
regional lymph nodes are associated with a favorable prognosis in
patients with colorectal carcinoma. Cancer Sci. 104:1237–1244.
2013.PubMed/NCBI View Article : Google Scholar
|
|
98
|
Saito Y, Ohnishi K, Miyashita A, Nakahara
S, Fujiwara Y, Horlad H, Motoshima T, Fukushima S, Jinnin M, Ihn H,
et al: Prognostic significance of CD169+ lymph node
sinus macrophages in patients with malignant melanoma. Cancer
Immunol Res. 3:1356–1363. 2015.PubMed/NCBI View Article : Google Scholar
|
|
99
|
Marmey B, Boix C, Barbaroux JB,
Dieu-Nosjean MC, Diebold J, Audouin J, Fridman WH, Mueller CG and
Molina TJ: CD14 and CD169 expression in human lymph nodes and
spleen: Specific expansion of CD14+CD169-
monocyte-derived cells in diffuse large B-cell lymphomas. Hum
Pathol. 37:68–77. 2006.PubMed/NCBI View Article : Google Scholar
|
|
100
|
Van Dinther D, Lopez Venegas M, Veninga H,
Olesek K, Hoogterp L, Revet M, Ambrosini M, Kalay H, Stöckl J, van
Kooyk Y and den Haan JMM: Activation of CD8+ T cell responses after
melanoma antigen targeting to CD169+ antigen presenting cells
inmice and humans. Cancers (Basel). 11(183)2019.PubMed/NCBI View Article : Google Scholar
|
|
101
|
Topf MC, Harshyne L, Tuluc M, Mardekian S,
Vimawala S, Cognetti DM, Curry JM, Rodeck U and Luginbuhl A: Loss
of CD169(+) subcapsular macrophages during metastatic spread of
head and neck squamous cell carcinoma. Otolaryngol Head Neck Surg.
161:67–73. 2019.PubMed/NCBI View Article : Google Scholar
|
|
102
|
Takeuchi H, Tanaka M, Tanaka A, Tsunemi A
and Yamamoto H: Predominance of M2-polarized macrophages in bladder
cancer affects angiogenesis, tumor grade and invasiveness. Oncol
Lett. 11:3403–3408. 2016.PubMed/NCBI View Article : Google Scholar
|
|
103
|
Iftakhar-E-Khuda I, Fair-Mäkelä R,
Kukkonen-Macchi A, Elima K, Karikoski M, Rantakari P, Miyasaka M,
Salmi M and Jalkanen S: Gene-expression profiling of different arms
of lymphatic vasculature identifies candidates for manipulation of
cell traffic. Proc Natl Acad Sci USA. 113:10643–10648.
2016.PubMed/NCBI View Article : Google Scholar
|
|
104
|
Wang B, Liu H, Dong X, Wu S, Zeng H, Liu
Z, Wan D, Dong W, He W, Chen X, et al: High CD204+
tumor-infiltrating macrophage density predicts a poor prognosis in
patients with urothelial cell carcinoma of the bladder. Oncotarget.
6:20204–20214. 2015.PubMed/NCBI View Article : Google Scholar
|
|
105
|
Li JQ, Yu XJ, Wang YC, Huang LY, Liu CQ,
Zheng L, Fang YJ and Xu J: Distinct patterns and prognostic values
of tumor-infiltrating macrophages in hepatocellular carcinoma and
gastric cancer. J Transl Med. 15(37)2017.PubMed/NCBI View Article : Google Scholar
|
|
106
|
Al Dubayee MS, Alayed H, Almansour R,
Alqaoud N, Alnamlah R, Obeid D, Alshahrani A, Zahra MM, Nasr A,
Al-Bawab A and Aljada A: Differential expression of human
peripheral mononuclear cells phenotype markers in type 2 diabetic
patients and type 2 diabetic patients on metformin. Front
Endocrinol (Lausanne). 9(537)2018.PubMed/NCBI View Article : Google Scholar
|
|
107
|
Jing W, Guo X, Wang G, Bi Y, Han L, Zhu Q,
Qiu C, Tanaka M and Zhao Y: Breast cancer cells promote CD169(+)
macrophage-associated immunosuppression through JAK2-mediated PD-L1
upregulation on macrophages. Int Immunopharmacol.
78(106012)2020.PubMed/NCBI View Article : Google Scholar
|