Introduction
Diabetic cardiomyopathy (DCM) is a distinct, chronic
complication of diabetes, particularly in patients with
long-standing, poorly controlled hyperglycemia, and is a notable
cause of heart failure and cardiovascular disease (CVD),
characterized by pathological changes of the cardiac structure and
function. It occurs independently of other cardiovascular risk
factors, such as hypertension or coronary artery disease (1)1 and contributes to
CVD-induced mortality and morbidity in diabetic patients (2). During the early stages of CVD,
increased lipid metabolism and impaired glucose metabolism are
initiated, which induce excessive oxidative stress and
inflammation. Excessive reactive oxygen species production and the
activation of excessive oxidative stress and inflammation lead to
myocardial damage and remodeling, typically impairing the diastolic
function first. With the progression of the disease, systolic
dysfunction develops and heart failure occurs (3).
Scutellaria baicalensis Georgi, particularly
its primary active flavonoid, baicalin¸ offers therapeutic effects
against cardiotoxicity (4),
myocardial ischemia (5),
obesity-induced cardiac dysfunction (6) and autoimmune myocarditis (7)- and doxorubicin-induced cardiotoxicity
(8). S. baicalensis can
treat CVD by specifically inhibiting apoptosis, decreasing
oxidative stress and inflammation and preventing cardiac fibrosis
(9-11).
It targets pro-inflammatory markers (TNF-α, IL-6, IL-1β and IL-8)
while increasing the levels of the anti-inflammatory cytokines
(IL-10) in myocardial tissues, which results in improved cardiac
function (12). Baicalin protects
against DCM by acting on multiple, complex and interrelated
pathways rather than a single target (13,14).
Therefore, key pathways and therapeutic targets involved in its
mechanism of action require a more comprehensive investigation and
further exploration.
Network pharmacology is an accessible method for
systematically identifying multi-target and -pathway mechanisms of
drug action and is used in numerous studies to determine active
compounds and underlying mechanisms (15,16).
It enables the identification of potential drug-target-disease
interactions, facilitating the exploration of novel therapeutic
applications. The present study employed the network pharmacology
approach to evaluate the therapeutic action of baicalin on DCM,
focusing on the interactions between candidate drug- and
disease-related targets (17).
Furthermore, the potential molecular mechanism of action underlying
the protective effects of baicalin on DCM was explored using
network pharmacology and in vitro assessments.
Materials and methods
Common target of baicalin and DCM
prediction
Swiss target prediction (swisstargetprediction.ch/) was used to discover
potential targets for baicalin. The candidate genes of DCM were
screened from the Comparative Toxicogenomics Database (ctdbase.org) and the GeneCards database (genecards.org/). The threshold for inclusion was set
at an inference score >30. Duplicate targets were eliminated,
and the shared targets were entered into the Venny diagram 2.1
online platform (bioinfogp.cnb.csic.es/tools/venny/index.html.) to
visualize the overlapping targets. Potential candidate targets were
imported into the WebGestalt server (version 2024; webgestalt.org/) for Gene Ontology (GO) (18) and Kyoto Encyclopedia of Genes and
Genomes (KEGG) enrichment analyses (kegg.jp/KEGG).
Molecular docking
Molecular docking analysis was performed as
previously described (19). The 2D
chemical structure of baicalin was retrieved from the PubChem
(pubchem.ncbi.nlm.nih.gov) database and
prepared for molecular docking by adding hydrogen atoms, followed
by free energy minimization using the CB-Dock2 platform (2022;
cadd.labshare.cn/cb-dock2/php/index.php). The crystal
structures of protein tyrosine phosphatase non-receptor type 22
(PTPN22) and tumor necrosis factor (TNF)-α were retrieved from the
Protein Databank (rcsb.org/) and modified by adding
hydrogen atoms and eliminating water molecules. The CB-Dock2
platform was used to calculate the binding affinity energy. PyMol
software (version 2.6; pymol.org/) and
BIOVIA Discovery Studio Visualizer (version 2021; discover.3ds.com/discovery-studio-visualizer-download)
were used to visualize the interactions between the ligand and
receptor molecules.
Culture of H9c2 cardiomyocyte cells
and small interfering (si)RNA transfection
H9c2 (Cellverse Co., Ltd.) cells were cultured in
DMEM (HyClone; Cytiva) with 10% fetal bovine serum (Sigma-Aldrich;
Merck KGaA) and 1% penicillin-streptomycin and were maintained at
37˚C in a 5% CO2 environment. The cells were grouped as
follows: Control (5.5 mM glucose), high-glucose (HG; 33 mM
glucose), HG + with low-dose baicalin (50 µM; HG + BL), HG +
high-dose baicalin (100 µM; HG + BH) and HG + PTPN22-siRNA. All
treatment was performed at 37˚C for 24 h. Cells were transfected
with siRNA (Shanghai GenePharma Co., Ltd) once they reached a
confluence of 60-80%. Lipo3000-B reagent was diluted with Opti-MEM
(Gibco; Thermo Fisher Scientific, Inc.) and mixed thoroughly. siRNA
(Table SI) was diluted to 50 nM
with Opti-MEM. Lipo3000-A reagent (Invitrogen; Thermo Fisher
Scientific, Inc.; cat. no. L3000001) and Lipo3000-B reagent were
added at room temperature for 5-15 min. The contents of the tubes
were added to the cells. The cell culture medium was replaced with
normal medium for subsequent experiments 6 h after transfection.
The samples were collected 48 h later for subsequent experimental
detection.
Cell viability
Cell viability was detected using Cell Counting
Kit-8 (Abbkine Scientific Co., Ltd.), according to the
manufacturer's instructions. After being exposed to baicalin (50 or
100 µM) for 48 h at 37˚C, CCK-8 solution was added for 1 h at 37˚C.
The absorbance of the resulting solution was evaluated with a
microplate reader at a wavelength of 450 nm.
Calcein-AM staining
Staining of cells was performed according to the
manufacturer's instructions of the Calcein-AM staining kit
(Beyotime) at 37˚C in the dark for 30 min. After incubation,
fluorescence can be directly detected using a fluorescence
microplate reader (excitation wavelength: 494 nm, emission
wavelength: 517 nm).
Reverse transcription-quantitative
(RT-q)PCR
Total RNA was extracted from cells using the Total
RNA Isolation Kit (cat. no. RE-030111, Fuji) according to the
manufacturer's instructions. Total RNA were reverse-transcribed
into complementary DNA using the 1st Strand cDNA Synthesis Kit
(cat. no. KY01, KeyCloud Biotech), according to the manufacturer's
protocol. RT-qPCR was performed on an ABI QuantStudio™ 12 K Flex
real-time PCR system (Applied Biosystems; Thermo Fisher Scientific,
Inc., Waltham, MA, USA) using the 2x SYBR Green qPCR Master Mix
(cat. no. KY02, KeyCloud Biotech, China). The primer sequences for
collagen I, transforming growth factor-β1 (TGF-β1), connective
tissue growth factor (CTGF), C-C motif chemokine ligand 20 (CCL20),
colony-stimulating factor 1 (CSF1), CSF2, and the housekeeping gene
glyceraldehyde-3-phosphate dehydrogenase (GAPDH) are listed in
Table SII. Thermocycling
conditions were as follows: initial denaturation at 95˚C for 30
sec, followed by 40 cycles of denaturation at 95˚C for 5 sec and
combined annealing/extension at 60˚C for 30 sec. Relative mRNA
expression levels were quantified using the 2-ΔΔCq method with
GAPDH as the endogenous reference gene (20). All target gene expression levels
were normalized to the average value of the control group.
Apoptosis analysis
Cell apoptosis was assessed using the TUNEL assay
kit (cat. no. KTA2011; Abbkine Scientific Co., Ltd.) according to
the manufacturer's instructions. Cells were fixed with 4%
paraformaldehyde at room temperature for 30 min and rinsed with PBS
for 5 min, followed by permeabilization using 0.02 µg/µl proteinase
K. TUNEL detection solution was then applied for 10 min at room
temperature. slides were washed by PBS at room temperature twice
for 5 min each time. Nuclei were stained with 0.05 µg/µl DAPI
solution at room temperature for 10 min in the dark. Mounting
medium was antifade mounting medium (Beijing Solarbio Science &
Technology Co., Ltd.). A fluorescence microscope was used to
determine the proportion of TUNEL-positive cells (UltraVIEW VoX
& IX81, Olympus Corporation). 3 fields of view were randomly
selected in each group.
Immunofluorescence staining
analysis
Cells were fixed with 4% paraformaldehyde at room
temperature for 15 min. Heat-induced antigen retrieval at 100˚C was
performed, followed by washing with PBS (pH 7.4). The slices were
treated with 3% BSA (Sigma) at room temperature for 2 h and
incubated with primary antibody (GSDMD-N, Proteintech, cat. No
66387-1-Ig, 1:200) overnight at 4˚C and FITC-labeled goat
anti-mouse IgG (1:10,000; cat.no A0568; Beyotime Biotechnology) for
1 h at room temperature. DAPI was added in the dark at room
temperature for 5 min. Images were captured by a Nikon fluorescence
microscope (NIKON ECLIPSE TI-SR, Nikon Corporation). Image analysis
was performed using ImageJ software (version 1.54k, National
Institutes of Health).
Western blotting
Protein was extracted from cells using RIPA lysis
buffer (cat. no. BL504A; Biosharp) supplemented with phosphatase
and protease inhibitors. The protein concentrations in the
supernatants were assessed by the BCA Protein Assay kit (Beyotime
Biotechnology). Equal amounts of proteins (50 µg/lane) were
separated using 10-12% SDS-PAGE, transferred to a PVDF membrane,
blocked with 5% BSA (Sigma-Aldrich; Merck KGaA) at room temperature
for 2 h and then incubated with primary antibodies as follows: CTGF
(cat. no. 25474-1-AP; 1:2,000), collagen I (cat. no. 67288-1-Ig;
1:1,000), TGF-β (cat. no. 21898-1-AP; all Proteintech Group, Inc.;
1:2,000), GSDMD (gasdermin D)-N (cat. no. bs-14287R; BIOSS), PTPN22
(cat. no. 11783-1-AP; both 1:1,000), apoptosis-associated
speck-like protein containing a CARD (ASC; cat. no. 10500-1-AP;
both Proteintech Group, Inc.; 1:20,000), NLRP3 (cat. no. ab263899;
Abcam), Caspase-1 (cat. no. AF5418; Affinity Biosciences; both
1:1,000), IL-1β (cat. no. 16806-1-AP; 1:5,000), IL-18 (cat. no.
10663-1-AP; 1:10,000), TNFα (cat. no. 17590-1-AP; 1:1,000),
phosphorylated (p-)p38, p38 (cat. no. 28796-1-AP and 66234-1-Ig;
both 1:3,000), p-ERK, ERK (cat. nos. 28733-1-AP and 16443-1-AP; all
Proteintech Group, Inc.; both 1:2,000), p-JNK, JNK (cat. no.
ABP50351and ABP51663; both Abbkine Scientific Co., Ltd.; both
1:1,000) and GAPDH (cat. no. 60004-1-Ig; Proteintech Group, Inc.;
1:50,000) at 4˚C with gentle shaking overnight. Immunoreactive
bands were revealed by HRP-conjugated anti-rabbit or anti-mouse IgG
(Beyotime Biotechnology; cat. no. A0208 and A0216; 1:1,000) for 1h
at room temperature. Protein bands were visualized with an enhanced
chemiluminescence (ECL) kit (Abbkine). Protein bands were detected
and analyzed by fully automatic chemiluminescence imaging system
(5200Multi, Tanon). Densitometry was analyzed by Tanon GIS 1D
(version 4.2; 5200Multi, Tanon).
Statistical analysis
All data are presented as the mean ± standard
deviation of three independent experimental repeats and were
analyzed with GraphPad Prism 9.5 (Dotmatics). Data were analyzed by
one-way ANOVA followed by Tukey's post hoc test. P<0.05 was
considered to indicate a statistically significant difference.
Results
Potential targets of baicalin against
DCM
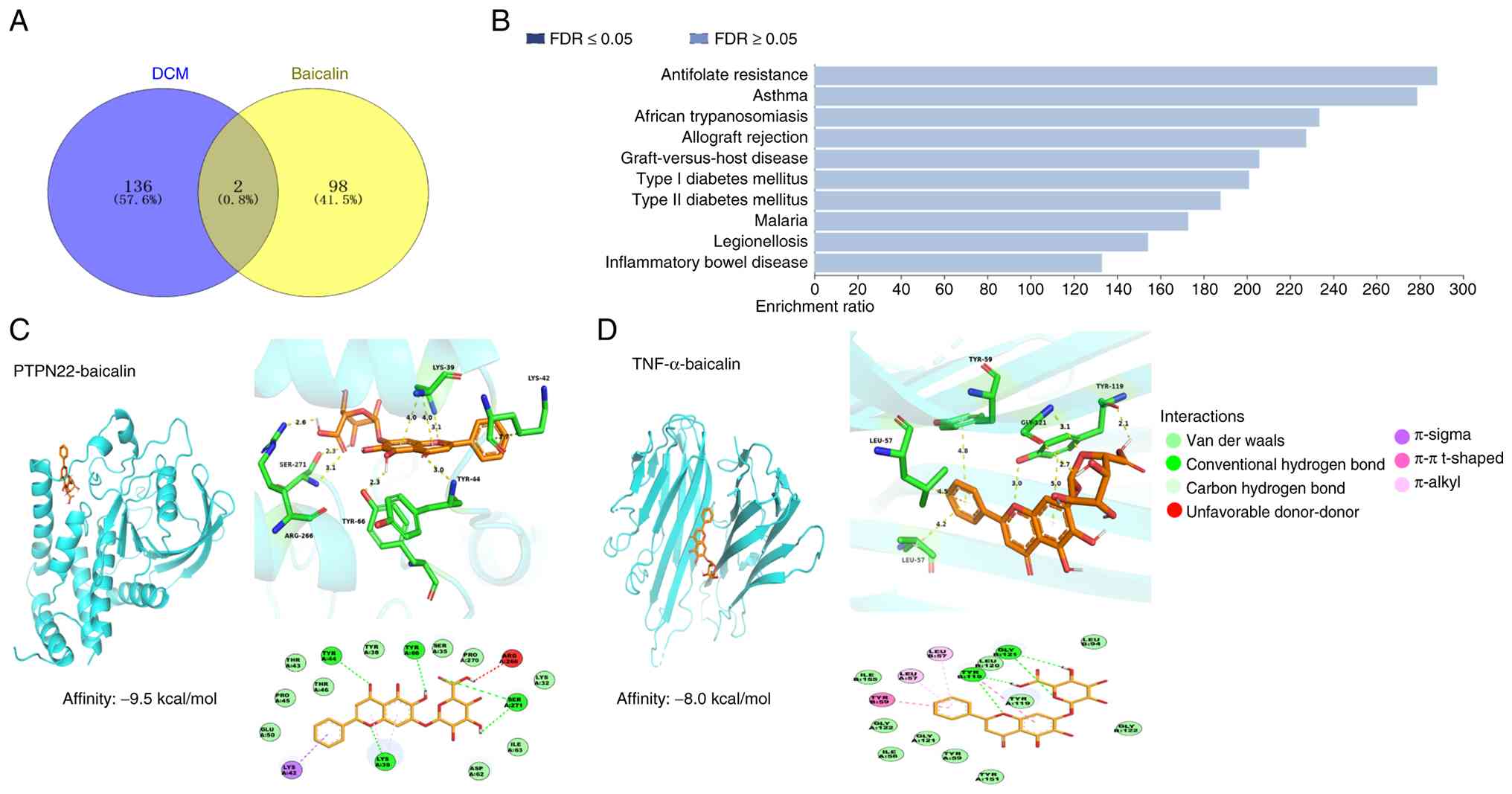
A total of 138 common targets associated with DCM
and 100 targets associated with baicalin were identified (Table SIII). PTPN22 and TNF were shared
targets between baicalin and DCM (Fig.
1A). KEGG) analysis did not identify any significant pathways
associated with the effects of baicalin on DCM (Fig. 1B).
Molecular docking
Baicalin exhibited a binding affinity of -9.5
kcal/mol for PTPN22 (Fig. 1C).
Based on 3D and 2D interaction analysis, baicalin had a strong
affinity for PTPN22, with key residues GLU50, PRO45, THR43, THR46,
TYR38, SER35, PRO270, LYS32, ILE63 and ASP62 facilitating
stabilization primarily through van der Waals interactions.
Additionally, baicalin formed stable hydrogen bonds with TYR44,
TYR66, SER271 and LYS39. The stable binding of baicalin to PTPN22
involved π-alkyl interactions with LYS39 and π-sigma interactions
with LYS42. However, an unfavorable donor-donor interaction was
found with the ARG266 residue. Baicalin demonstrated a binding
affinity of -8.0 kcal/mol for TNF (PDB ID: 5MU8; Fig. 1D). The 3D and 2D interaction maps
indicated that baicalin bound TNF primarily through van der Waals
interactions, with key residues including TYR151, TYR59, GLY121,
ILE58, GLY122, ILE155, LEU120, TYR119, LEU94 and GLY122 forming the
binding pocket. The stable binding of baicalin to TNF was achieved
through hydrogen bonds with TYR119 and GLY121. Additionally, the
stable binding involved π-alkyl interactions with the residues
LEU57 and LEU57. Finally, baicalin interacted with TNF through π-π
T-shaped interactions with the TYR59 and TYR119 residues.
Baicalin suppresses the secretion of
HG-induced fibrotic and inflammatory markers in vitro
Western blot and RT-qPCR analyses of H9c2 cell
lysates demonstrated that HG significantly elevated the expression
levels of fibrotic markers, such as collagen I, TGF-β and CTGF
(Fig. 2A-G), as well as
inflammatory markers, including CCL20, CSF1, and CSF2 (Fig. 2H-J). However, BH significantly
counteracted these changes compared with HG group.
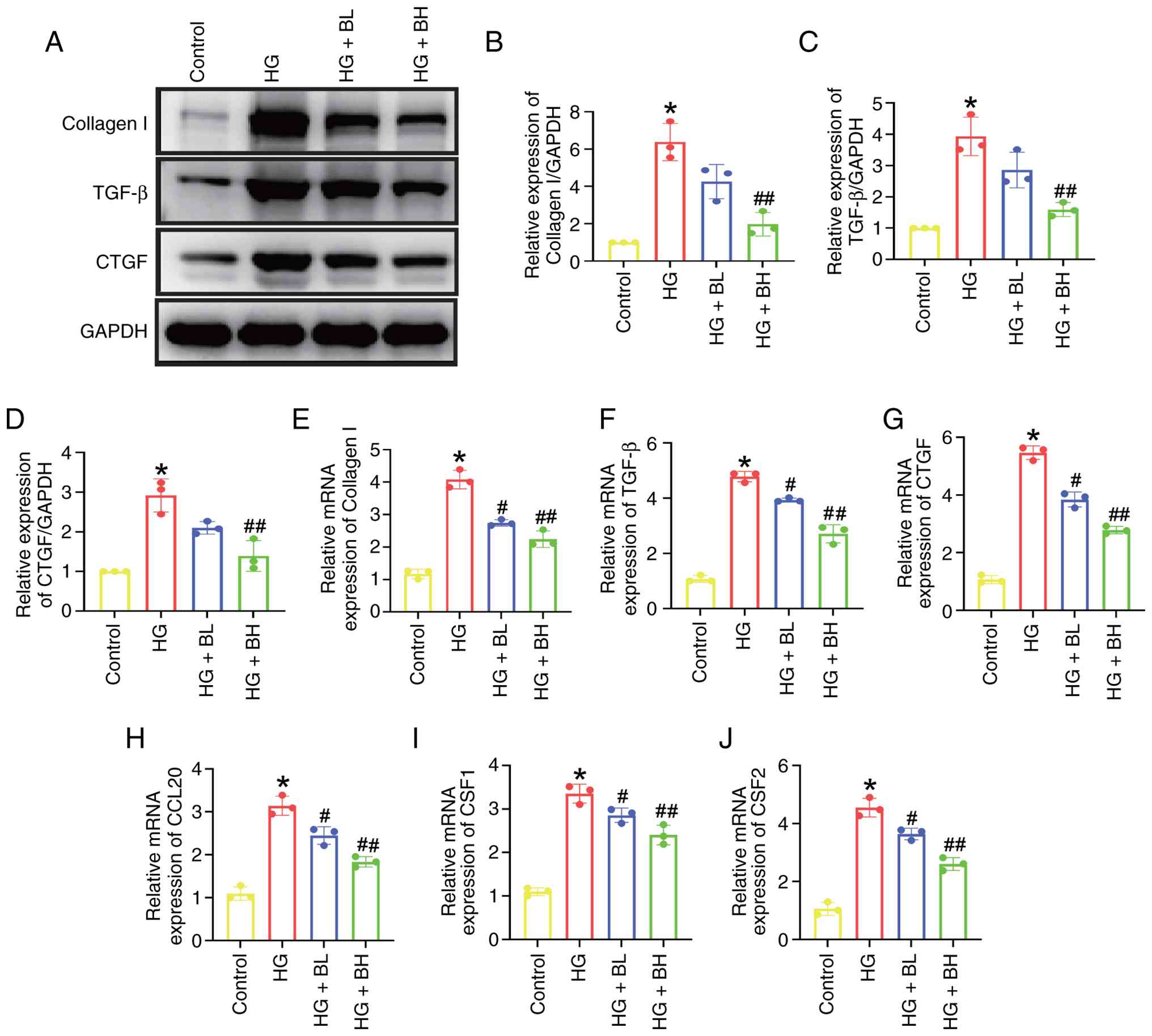 | Figure 2Baicalin suppresses HG-induced
release of fibrotic and inflammatory markers in H9c2 cells. (A)
Western blotting analysis of (B) collagen I, (C) TGF-β and (D) CTGF
protein expression in H9c2 cells challenged with HG (33 mM glucose)
in the presence or absence of baicalin (BL: 50 µmol; BH: 100 µmol)
for 24 h. Quantitative analysis of (E) collagen I, (F) TGF-β, (G)
CTGF, (H) CCL20, (I) CSF1 and (J) CSF2 mRNA levels detected by
reverse transcription-quantitative PCR. *P<0.05 vs.
Control; #P<0.05, ##P<0.05 vs. HG. HG,
high-glucose; CTGF, connective tissue growth factor; BL, baicalin
low; BH, baicalin high; CCL, C-C motif chemokine ligand; CSF,
colony stimulating factor. |
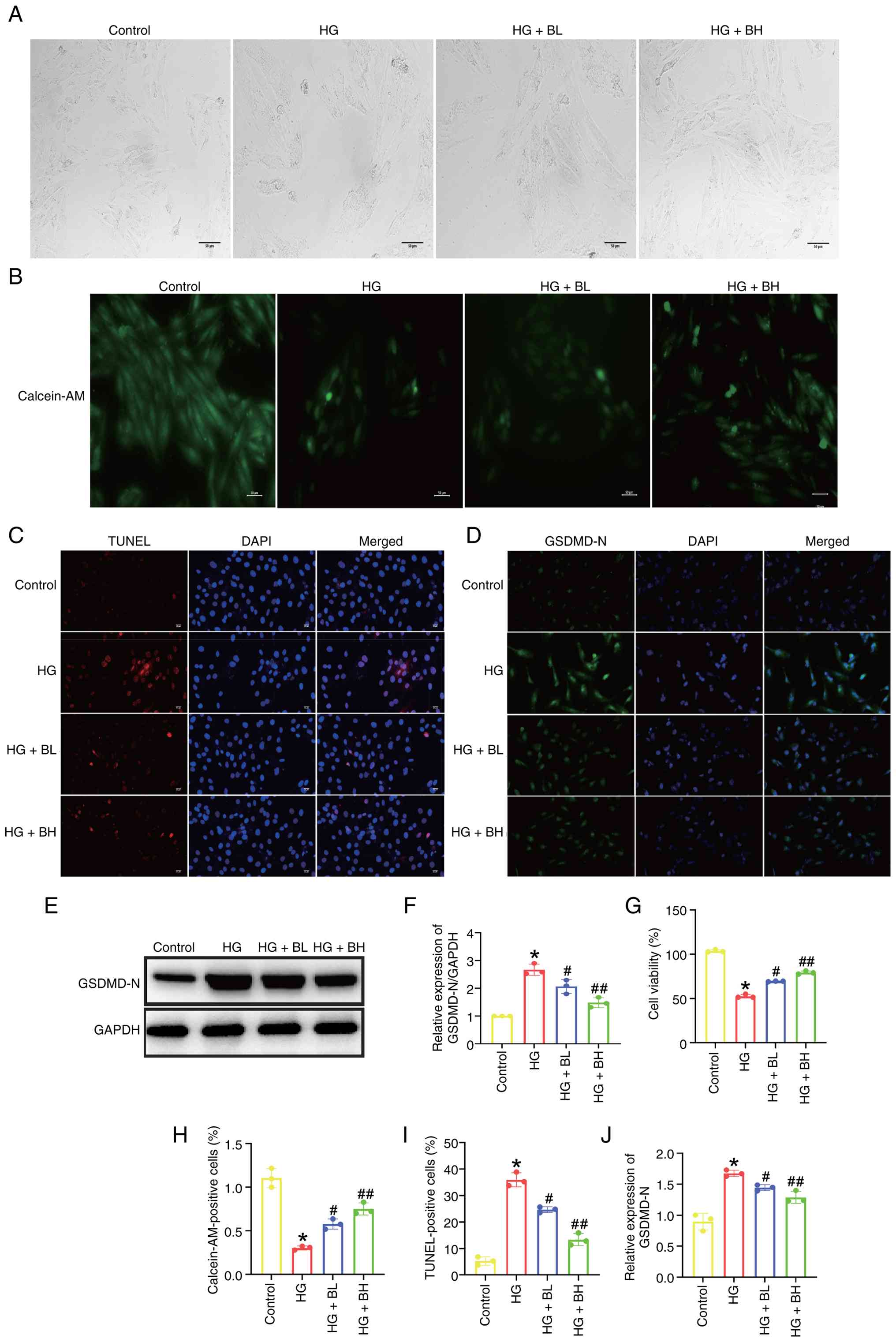
Baicalin mitigates HG-induced
pyroptosis in vitro
Fig. 3A displays
HG-induced characteristic morphological changes in H9c2 cells,
including swelling and rupture of the cell membrane. Calcein-AM
staining revealed increased membrane damage resulting from HG
exposure (Fig. 3B and H). CCK-8 assay showed that baicalin
protected H9c2 cells against HG-induced cell death (Fig. 3G), a finding corroborated by TUNEL
staining (Fig. 3C and I). Additionally, baicalin was found to
reduce the expression of the pyroptosis marker-GSDMD-N, increased
by exposure to HG (Fig. 3D-F and
J).
Baicalin inhibits the activation of
the TNF and PTPN22 signaling pathways induced by HG in vitro
The present study aimed to verify the effects of
baicalin on the TNF and PTPN22 signaling pathways in vitro.
BH decreased the elevated TNF-α expression as well as p38, ERK and
JNK phosphorylation levels compared with HG (Fig. 4A-E). BH significantly decreased the
elevated expression levels associated with the PTPN22 signaling
pathway, including PTPN22, NLRP3, ASC, caspase, IL-1β, and IL-18
compared with HG group (Fig.
4F-L).
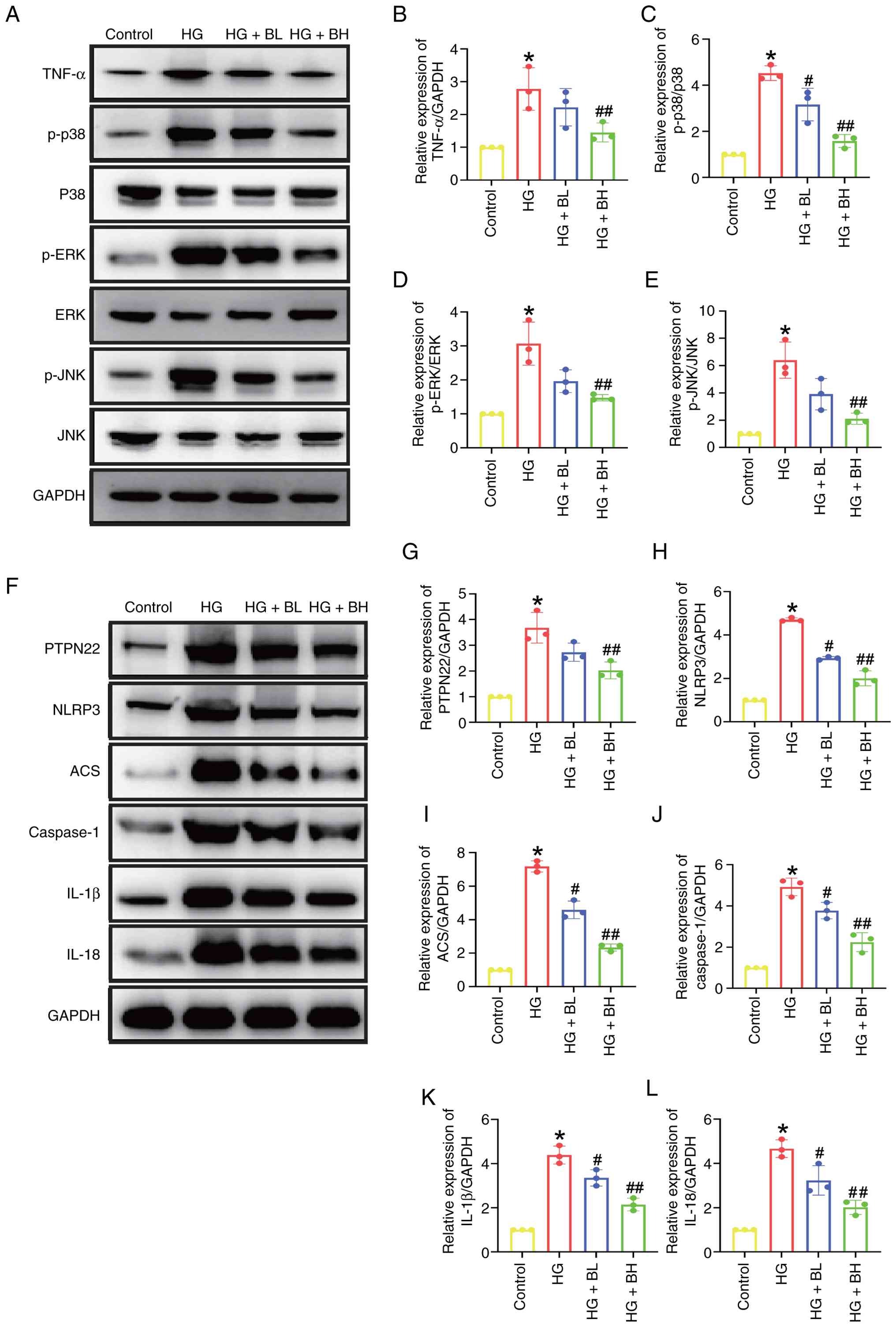 | Figure 4Baicalin inhibits HG-induced
activation of TNF and PTPN22 signaling pathways in H9c2 cells. H9c2
cells were treated with HG, or baicalin (BL: 50 µm; BH: 100 µm) for
24 h. (A) Western blotting of (B) TNF-α expression and
phosphorylation levels of (C) p38, (D) ERK and (E) JNK. (F) Western
blot analysis of (G) PTPN22, (H) NLRP3, (I) ASC, (J) caspase, (K)
IL-1β and (L) IL-18 expression. *P<0.05 vs. Control;
#P<0.05, ##P<0.05 vs. HG. HG,
high-glucose; BL, baicalin low; BH, baicalin high; p-,
phosphorylated; PTPN, protein tyrosine phosphatase non-receptor
type; ASC, apoptosis-associated speck-like protein containing a
CARD.. |
PTPN22 inhibition attenuated
HG-induced pyroptosis in vitro
The role of PTPN22 in HG-induced pyroptosis was
investigated by PTPN22-siRNA transfection. The successful knockdown
of PTPN22 in cells was confirmed by western blot analysis (Fig. S1). Based on the results, the siRNA
PTPN22#2 was selected for subsequent studies. Based on the findings
from the morphological analysis (Fig.
5A) and Calcein-AM (Fig. 5B and
L) staining, HG-induced cell
swelling and membrane rupture were reduced when PTPN22 was
silenced. PTPN22-siRNA decreased cell apoptosis (Fig. 5C and M) and the expression of GSDMD-N (Fig. 5D and N). Additionally, PTPN22 inhibition
effectively blocked the NLRP3 signaling pathway (Fig. 5E-K).
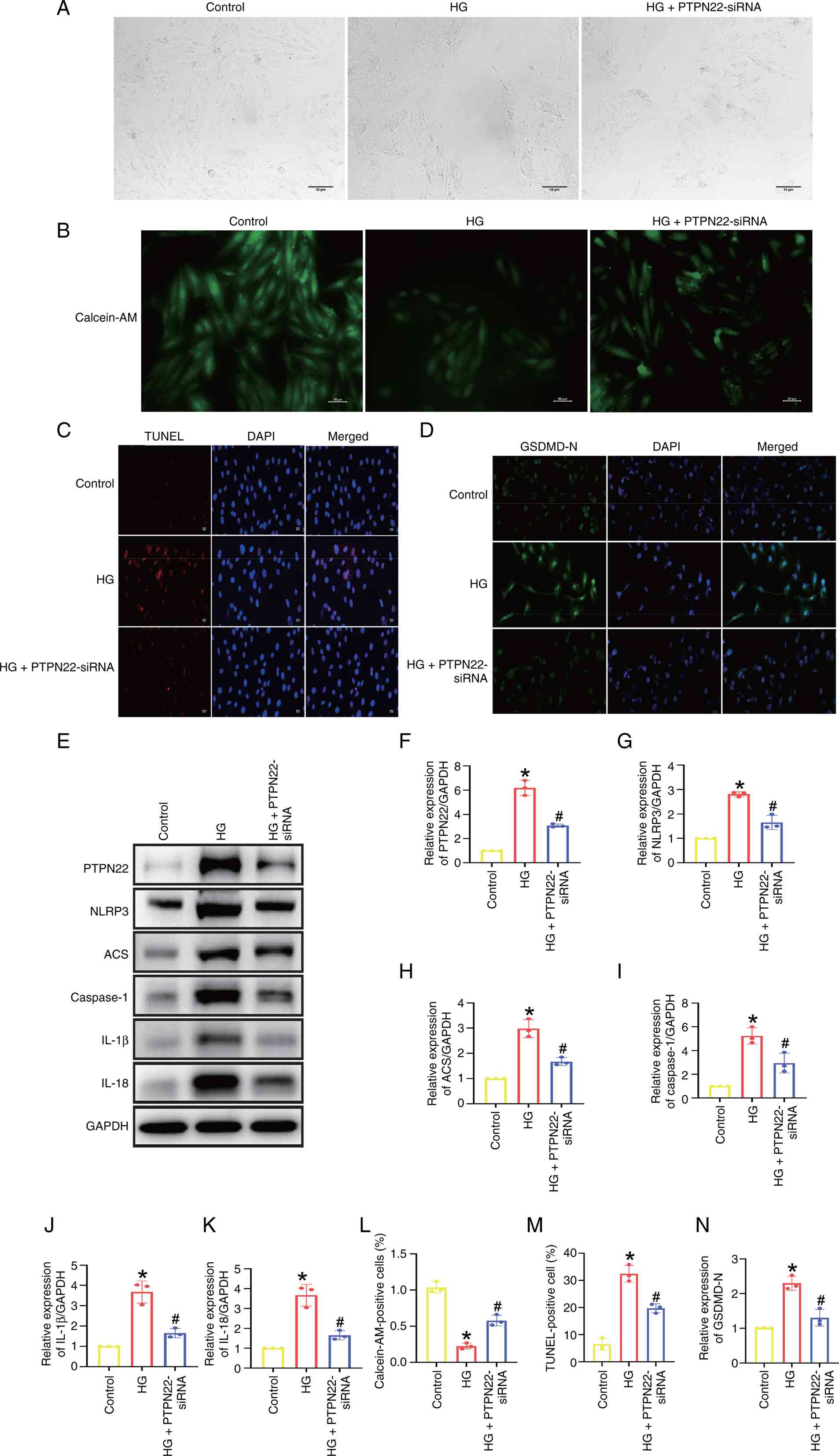 | Figure 5PTPN22 inhibition attenuates
HG-induced pyroptosis in H9c2 cells. H9c2 cells were treated with
HG or PTPN22-siRNA for 6 h following 48 h incubation in normal
culture medium. (A) Morphological changes of H9c2 cells. Scale bar,
50 µm. (B) Calcein-AM, (C) TUNEL and (D) GSDMD-N immunofluorescence
staining of H9c2 cells. Scale bar, 20 µm. (E) Western blot analysis
of (F) PTPN22, (G) NLRP3, (H) ASC, (I) caspase, (J) IL-1β and (K)
IL-18 expression. Quantitative analysis of (L) Calcein-AM staining,
(M) TUNEL-positive cells and (N) GSDMD-N immunofluorescence
staining. *P<0.05 vs. control; #P<0.05
vs. HG. HG, high-glucose; GSDM, gasdermin; BL, baicalin low; BH,
baicalin low; si, small interfering; PTPN, protein tyrosine
phosphatase non-receptor type; ASC, apoptosis-associated speck-like
protein containing a CARD. |
Discussion
The present study investigated the underlying
mechanisms of the protective effects of baicalin against DCM. Using
network pharmacology, two important targets linked to
baicalin-induced therapeutic effects on DCM were identified,
namely, PTPN22 and the TNF-α pathway. The binding interactions of
baicalin with PTPN22 and TNF-α were confirmed using molecular
docking. Baicalin was shown to protect against HG-induced injury by
inhibiting the pyroptosis pathway, decreasing inflammation and
mitigating fibrotic responses. It serves as a therapeutic agent by
blocking the activation of TNF and PTPN22 pathways. Furthermore,
the inhibition of PTPN22 suppresses this detrimental signaling,
decreasing HG-induced pyroptosis and cellular damage.
DCM is characterized by structural and functional
abnormalities, including inflammation, oxidative stress, cardiac
lipotoxicity, mitochondrial dysfunction and endoplasmic reticulum
stress (21), which collectively
cause cardiomyocyte death and fibrosis. These processes lead to
left ventricular remodeling and decreased ejection fraction,
typically resulting in severe heart failure and high mortality
rates (22). Numerous studies have
demonstrated that diabetes mellitus increases both the
susceptibility and incidence of heart failure by around 2.5-fold;
notably, this figure doubles in female patients, irrespective of
age and accompanying conditions such as coronary artery disease and
dyslipidemia (23-27).
In vivo studies have shown baicalin-induced therapeutic
effects against DCM, revealing that it improves cardiac function in
DCM via the regulation of autophagy (28,29).
Baicalin is also reported to treat DCM by reducing activation of
the purinergic receptor P2Y, G Protein Coupled 12 receptor and
NRF2/P62 in vivo (30,31). However, the mechanisms by which
baicalin decreases HG-induced damage in H9c2 cells are unknown. The
present study demonstrated baicalin decreased pyroptosis and
fibrotic and inflammatory markers in vitro.
Diabetes is marked by persistent inflammation, which
serves a role in the progression of DCM. Elevated levels of
inflammatory cytokines, particularly TNF-α, IL-6, and IL-1β, are
key drivers of DCM, promoting structural damage through cardiac
remodeling and fibrosis (32). TNF
initiates the recruitment of receptor-interacting protein kinase 1
and cell division cycle 37 and heat shock protein 90 to form a
complex that produces active IKK complexes (33). This activation stimulates the
kinases p38, ERK and JNK, ultimately leading to the upregulation of
pro-inflammatory genes (34). An
increase in genes associated with the TNF signaling pathway,
including CCL20, CSF1 and CSF2, has been observed during the
inflammatory response (35). By
targeting key proteins involved in inflammatory signaling,
therapeutic approaches for the treatment of DCM may be established.
Here, baicalin showed therapeutic potential in DCM by directly
binding and inhibiting TNF-α. It suppressed HG-induced inflammation
by downregulating TNF-α expression and inhibiting downstream p38,
ERK and JNK phosphorylation. Baicalin also decreased expression of
pro-inflammatory factors such as CCL20, CSF1 and CSF2, mitigating
HG-induced injury. Therefore, baicalin modulated TNF-α pathways in
DCM.
Pyroptosis is a pro-inflammatory form of programmed
cell death defined by GSDM-mediated plasma membrane rupture
(36). Pyroptosis has been
identified as a key factor in DCM (37-39).
When activated, NLRP3 oligomers are formed, triggered by stimuli,
including microbes, particulate matter and damage-associated
molecules. This process facilitates the recruitment of the adaptor
protein (ASC) and caspase-1(40),
resulting in the cleavage of GSDMD. Caspase-1 converts pro-IL-1β
and pro-IL-18 into their active forms. The N-terminal domain of
GSDMD forms pores that cause cell swelling, promoting IL-1β and
IL-18 release. Baicalin reduces inflammatory diseases by inhibiting
pyroptosis (41). However, its
specific role in DCM requires further study. Here, treatment with
baicalin in vitro alleviated HG-induced pyroptosis. PTPN22
encodes the lymphoid-specific tyrosine phosphatase, a crucial
regulator in hematopoietic cells (42). The present study demonstrated that
baicalin can bind to PTPN22 and inhibit the HG-induced upregulation
of PTPN22 expression. While KEGG analysis did not identify any
significant signaling pathways associated with baicalin-induced
therapeutic effects on DCM, PTPN22 is involved in controlling the
activation of the NLRP3 inflammasome and the IL-1β pathway
(43). Moreover, baicalin
suppressed the expression of GSDMD, PTPN22, NLRP3, ASC, caspase,
IL-1β and IL-18, indicating that PTPN22 may induce DCM via the
pyroptosis pathwayPTPN22 inhibition also blocked pyroptosis and the
NLRP3 signaling pathway, suggesting PTPN22 regulated pyroptosis via
the NLRP3 signaling pathway.
Expression of PTPN22 is inhibited by TNF-α in
peripheral blood mononuclear cells (44). PTPN22/22 loss of function increases
TNF-α secretions in Jurkat T cells (45). Upregulation of TNF-α is observed in
PTPN22 1858C/T single-nucleotide polymorphism in rheumatoid
arthritis (46). However, the exact
mechanism of the interaction between PTPN22 and TNF-α remains
unknown.
There are several limitations to the present study.
The absence of additional databases may limit the scope of target
screening. The mechanism of baicalin in the treatment of diabetic
cardiomyopathy was not fully identified in the present study, since
only two common targets and no significant pathways were found with
network pharmacology. Future studies should expand the scope of
target prediction by introducing more databases to explore the
multi-target therapeutic mechanism of baicalin in treating DCM.
Furthermore, the present study was only validated in H9c2
cardiomyocytes; to increase clinical translational value, the
physiological significance of the identified targets (PTPN22 and
TNF-α) and associated pathways must be evaluated in in vivo
models. In addition, the lack of a positive control makes it
difficult to evaluate the relative efficacy and clinical relevance
of baicalin. A positive control group should be included in future
in vitro and in vivo studies to verify the
therapeutic effects and clinical potential of baicalin against
DCM.
The primary reason for not introducing additional
databases in the network pharmacology-based target screening
process was to ensure the reliability and consistency of the
predicted targets. The present study selected Swiss Target
Prediction) for baicalin target prediction, which is commonly used
in network pharmacology studies of traditional Chinese medicine
monomers and has high target prediction accuracy (47-49).
This database covers the main target information of baicalin,
including its potential binding proteins and associated signaling
molecules, which met the needs of target screening for the present
study. In addition, the present study considered homogeneity and
avoidance of redundant information between databases. Many existing
target prediction databases have overlapping target information and
introducing too many additional databases may lead to redundant
target screening results, increase the complexity of target
intersection analysis and introduce false-positive targets due to
differences in database prediction algorithms and data sources.
This would not only fail to improve the comprehensiveness of target
screening but also affect the accuracy and efficiency of
experimental verification. Additionally, the present study used a
relatively high cutoff value (>30) for target screening, which
decreased the number of targets that were originally screened,
which had an impact on the number of common targets. Furthermore,
the present study aimed to verify the therapeutic effect and
underlying mechanisms of baicalin on DCM through in vitro
experiments; network pharmacology was used as an auxiliary tool to
screen potential targets and guide subsequent experimental design.
The in vitro results supported the validity of the network
pharmacology predictions.
Although KEGG pathway analysis did not reveal any
significant enriched pathways, the two identified common targets
(PTPN22 and TNF-α) are associated with the pathogenesis of DCM.
PTPN22 is associated with myocardial inflammation (50), oxidative stress (51) and cardiomyocyte apoptosis (52), which are key pathological processes
of DCM. PTPN22 can regulate the activation of T cells and release
of inflammatory factors, thereby participating in the progression
of DCM (53). Similarly, TNF-α is a
classical pro-inflammatory cytokine that plays a pivotal role in
the development of DCM by inducing myocardial inflammation,
promoting cardiomyocyte pyroptosis and impairing cardiac function
(54). These well-documented
associations between the two targets and DCM pathogenesis support
their potential as key therapeutic targets of baicalin in DCM
(55,56). In addition, the selection of PTPN22
and TNF-α was based on the reliability of network pharmacology
prediction results. Although the present study only identified two
common targets due to the limitations of database selection and
high screening cutoff value, these targets were consistently
predicted by the databases. The present study verified the binding
ability of baicalin to PTPN22 and TNF-α through molecular docking
experiments, which showed strong binding affinity, indicating that
baicalin directly bound these two targets and exerted regulatory
effects. The molecular docking verification further confirms the
rationality of selecting these two targets. Furthermore,
considering the multi-target and multi-pathway characteristics of
traditional Chinese medicine monomers, the lack of enriched KEGG
pathways does not mean that the identified targets are invalid.
Traditional Chinese medicine monomers typically exert therapeutic
effects by regulating multiple targets and signaling pathway
networks, rather than relying on a single or significantly enriched
pathways. Here, baicalin inhibited the expression of PTPN22 and
TNF-α and suppressed the downstream inflammatory and pyroptosis
pathways mediated by these targets. These results not only confirm
the regulatory effect of baicalin on the two targets but also
explain the potential therapeutic mechanism of baicalin in treating
DCM from the perspective of target regulation.
In conclusion, the primary cell response to baicalin
was suppression of inflammation and pyroptosis. By regulating the
activation of the PTPN22 and TNF-α signaling pathways, baicalin
inhibited inflammation and pyroptosis in DCM. TNF-α and PTPN22 were
key factors in the development of DCM and PTPN22 was involved in
pyroptosis. Therefore, dual targeting of these two pathways may
represent a promising strategy for treating DCM by treating
HG-induced pyroptosis with baicalin.
Supplementary Material
Knockdown efficiency of three siRNAs
targeting PTPN22. (A) Western blotting of (B) PTPN22 expression
following treatment with siRNA. ***P<0.0001 vs.
control. si, small interfering; PTPN, protein tyrosine phosphatase
non-receptor type.
Sequences of the siRNAs.
Primers for reverse
transcription-quantitative PCR.
Common targets of DCM and
baicalin.
Acknowledgements
Not applicable.
Funding
Funding: The present study was supported by Science and
Technology Plan Project of Wenzhou Municipality (grant no.
Y2023414).
Availability of data and materials
The data generated in the present study may be
requested from the corresponding author.
Authors' contributions
XZ wrote the manuscript and designed the
experiments. LX analyzed data. NZ performed the experiments and
edited the manuscript. LX and NZ confirm the authenticity of all
the raw data. All authors have read and approved the final
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Xie SY, Liu SQ, Zhang T, Shi WK, Xing Y,
Fang WX, Zhang M, Chen MY, Xu SC, Fan MQ, et al: USP28 serves as a
key suppressor of mitochondrial morphofunctional defects and
cardiac dysfunction in the diabetic heart. Circulation.
149:684–706. 2024.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Dillmann WH: Diabetic cardiomyopathy. Circ
Res. 124:1160–1162. 2019.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Hölscher ME, Bode C and Bugger H: Diabetic
cardiomyopathy: Does the type of diabetes matter? Int J Mol Sci.
17(2136)2016.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Sun X, Wang X, He Q, Zhang M, Chu L, Zhao
Y, Wu Y, Zhang J, Han X, Chu X, et al: Investigation of the
ameliorative effects of baicalin against arsenic trioxide-induced
cardiac toxicity in mice. Int Immunopharmacol.
99(108024)2021.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Chan E, Liu XX, Guo DJ, Kwan YW, Leung GP,
Lee SM and Chan SW: Extract of scutellaria baicalensis georgi root
exerts protection against myocardial ischemia-reperfusion injury in
rats. Am J Chin Med. 39:693–704. 2011.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Guo L, Yang J, Yuan W, Li C, Li H, Yang Y,
Xue R and Yan K: Baicalein ameliorated obesity-induced cardiac
dysfunction by regulating the mitochondrial unfolded protein
response through NRF2 signaling. Phytomedicine.
126(155441)2024.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Xin L, Gao J, Lin H, Qu Y, Shang C, Wang
Y, Lu Y and Cui X: Regulatory mechanisms of baicalin in
cardiovascular diseases: A review. Front Pharmacol.
11(583200)2020.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Zeng Y, Liao X, Guo Y, Liu F, Bu F, Zhan
J, Zhang J, Cai Y and Shen M: Baicalin-peptide supramolecular
self-assembled nanofibers effectively inhibit ferroptosis and
attenuate doxorubicin-induced cardiotoxicity. J Control Release.
366:838–848. 2024.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Wang T, Wang S, Jia X, Li C, Ma X, Tong H,
Liu M and Li L: Baicalein alleviates cardiomyocyte death in EAM
mice by inhibiting the JAK-STAT1/4 signalling pathway.
Phytomedicine. 128(155558)2024.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Hu S, Jiang L, Yan Q, Zhou C, Guo X, Chen
T, Ma S, Luo Y, Hu C, Yang F, et al: Evidence construction of
baicalin for treating myocardial ischemia diseases: A preclinical
meta-analysis. Phytomedicine. 107(154476)2022.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Zhao F, Fu L, Yang W, Dong Y, Yang J, Sun
S and Hou Y: Cardioprotective effects of baicalein on heart failure
via modulation of Ca(2+) handling proteins in vivo and in vitro.
Life Sci. 145:213–223. 2016.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Luan Y, Sun C, Wang J, Jiang W, Xin Q,
Zhang Z and Wang Y: Baicalin attenuates myocardial
ischemia-reperfusion injury through Akt/NF-κB pathway. J Cell
Biochem. 120:3212–3219. 2019.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Yu H, Chen B and Ren Q: Baicalin relieves
hypoxia-aroused H9c2 cell apoptosis by activating
Nrf2/HO-1-mediated HIF1α/BNIP3 pathway. Artif Cells Nanomed
Biotechnol. 47:3657–3663. 2019.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Cheng Y, Yan M, He S, Xie Y, Wei L, Xuan
B, Shang Z, Wu M, Zheng H, Chen Y, et al: Baicalin alleviates
angiotensin II-induced cardiomyocyte apoptosis and autophagy and
modulates the AMPK/mTOR pathway. J Cell Mol Med.
28(e18321)2024.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Nogales C, Mamdouh ZM, List M, Kiel C,
Casas AI and Schmidt HHHW: Network pharmacology: Curing causal
mechanisms instead of treating symptoms. Trends Pharmacol Sci.
43:136–150. 2022.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Zhang P, Zhang D, Zhou W, Wang L, Wang B,
Zhang T and Li S: Network pharmacology: Towards the artificial
intelligence-based precision traditional Chinese medicine. Brief
Bioinform. 25(bbad518)2023.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Zhu N, Huang B, Zhu L and Wang Y:
Potential mechanisms of triptolide against diabetic cardiomyopathy
based on network pharmacology analysis and molecular docking. J
Diabetes Res. 2021(9944589)2021.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Denny P, Feuermann M, Hill DP, Lovering
RC, Plun-Favreau H and Roncaglia P: Exploring autophagy with gene
ontology. Autophagy. 14:419–436. 2018.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Zhao X, Huang B, Zhang J, Xiang W and Zhu
N: Celastrol attenuates streptozotocin-induced diabetic
cardiomyopathy in mice by inhibiting the ACE / Ang II / AGTR1
signaling pathway. Diabetol Metab Syndr. 15(186)2023.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-delta delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Tan Y, Zhang Z, Zheng C, Wintergerst KA,
Keller BB and Cai L: Mechanisms of diabetic cardiomyopathy and
potential therapeutic strategies: Preclinical and clinical
evidence. Nat Rev Cardiol. 17:585–607. 2020.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Ritchie RH and Abel ED: Basic mechanisms
of diabetic heart disease. Circ Res. 126:1501–1525. 2020.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Gulsin GS, Swarbrick DJ, Hunt WH, Levelt
E, Graham-Brown MPM, Parke KS, Wormleighton JV, Lai FY, Yates T,
Wilmot EG, et al: Relation of aortic stiffness to left ventricular
remodeling in younger adults with type 2 diabetes. Diabetes.
67:1395–1400. 2018.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Nichols GA, Gullion CM, Koro CE, Ephross
SA and Brown JB: The incidence of congestive heart failure in type
2 diabetes: An update. Diabetes Care. 27:1879–1884. 2004.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Thrainsdottir IS, Aspelund T, Thorgeirsson
G, Gudnason V, Hardarson T, Malmberg K, Sigurdsson G and Rydén L:
The association between glucose abnormalities and heart failure in
the population-based Reykjavik study. Diabetes Care. 28:612–616.
2005.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Aksnes TA, Kjeldsen SE, Rostrup M, Omvik
P, Hua TA and Julius S: Impact of new-onset diabetes mellitus on
cardiac outcomes in the valsartan antihypertensive long-term use
evaluation (VALUE) trial population. Hypertension. 50:467–473.
2007.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Ohkuma T, Komorita Y, Peters SAE and
Woodward M: Diabetes as a risk factor for heart failure in women
and men: A systematic review and meta-analysis of 47 cohorts
including 12 million individuals. Diabetologia. 62:1550–1560.
2019.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Zhang P, Wu H, Lou H, Zhou J, Hao J, Lin
H, Hu S, Zhong Z, Yang J, Guo H and Chi J: Baicalin attenuates
diabetic cardiomyopathy in vivo and in vitro by inhibiting
autophagy and cell death through SENP1/SIRT3 signaling pathway
activation. Antioxid Redox Signal. 42:53–76. 2025.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Xu Q, Liu Z, Wang L, Liu G, Zhao L, Yue H
and Liu Y: Baicalin protects against myocardial fibrosis through
inhibition of DOT1L/COL-1 pathway during diabetic cardiomyopathy.
Sci Rep. 16(1914)2025.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Sheng X, Wang J, Guo J, Xu Y, Jiang H,
Zheng C, Xu Z, Zhang Y, Che H, Liang S, et al: Effects of baicalin
on diabetic cardiac autonomic neuropathy mediated by the P2Y12
receptor in rat stellate ganglia. Cell Physiol Biochem. 46:986–998.
2018.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Wang W, Han R, Lai L and Zhang X:
Unlocking the potential: Baicalin's apoptosis-reducing power and
activation of NRF2/P62 for alleviating diabetic cardiomyopathy in
rats. Mol Cell Toxicol. 21:183–196. 2024.
|
|
32
|
Luo W, Lin K, Hua J, Han J, Zhang Q, Chen
L, Khan ZA, Wu G, Wang Y and Liang G: Schisandrin B attenuates
diabetic cardiomyopathy by targeting MyD88 and inhibiting
MyD88-dependent inflammation. Adv Sci (Weinh).
9(e2202590)2022.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Chen G, Cao P and Goeddel DV: TNF-induced
recruitment and activation of the IKK complex require Cdc37 and
Hsp90. Mol Cell. 9:401–410. 2002.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Newton K and Dixit VM: Signaling in innate
immunity and inflammation. Cold Spring Harb Perspect Biol.
4(a006049)2012.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Qian K, Shan L, Shang S, Li T, Wang S, Wei
M, Tang B and Xi J: Manganese enhances macrophage defense against
mycobacterium tuberculosis via the STING-TNF signaling pathway. Int
Immunopharmacol. 113(109471)2022.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Coll RC, Schroder K and Pelegrín P: NLRP3
and pyroptosis blockers for treating inflammatory diseases. Trends
Pharmacol Sci. 43:653–668. 2022.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Fu Y, Shen J, Li Y, Liu F, Ning B, Zheng Y
and Jiang X: Inhibition of the PERK/TXNIP/NLRP3 axis by baicalin
reduces NLRP3 inflammasome-mediated pyroptosis in macrophages
infected with mycobacterium tuberculosis. Mediators Inflamm.
2021(1805147)2021.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Meng L, Lin H, Huang X, Weng J, Peng F and
Wu S: METTL14 suppresses pyroptosis and diabetic cardiomyopathy by
downregulating TINCR lncRNA. Cell Death Dis. 13(38)2022.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Liu P, Zhang Z, Chen H and Chen Q:
Pyroptosis: Mechanisms and links with diabetic cardiomyopathy.
Ageing Res Rev. 94(102182)2024.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Zeng C, Duan F, Hu J, Luo B, Huang B, Lou
X, Sun X, Li H, Zhang X, Yin S and Tan H: NLRP3
inflammasome-mediated pyroptosis contributes to the pathogenesis of
non-ischemic dilated cardiomyopathy. Redox Biol.
34(101523)2020.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Wei Z, Gao R, Sun Z, Yang W, He Q, Wang C,
Zhang J, Zhang X, Guo L and Wang S: Baicalin inhibits influenza A
(H1N1)-induced pyroptosis of lung alveolar epithelial cells via
caspase-3/GSDME pathway. J Med Virol. 95(e28790)2023.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Wang X, Wei G, Ding Y, Gui X, Tong H, Xu
X, Zhang S, Sun Z, Ju W, Li Y, et al: Protein tyrosine phosphatase
PTPN22 negatively modulates platelet function and thrombus
formation. Blood. 140:1038–1051. 2022.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Spalinger MR, Lang S, Gottier C, Dai X,
Rawlings DJ, Chan AC, Rogler G and Scharl M: PTPN22 regulates
NLRP3-mediated IL1B secretion in an autophagy-dependent manner.
Autophagy. 13:1590–1601. 2017.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Chang HH, Ho CH, Tomita B, Silva AA,
Sparks JA, Karlson EW, Rao DA, Lee YC and Ho IC: Utilizing a PTPN22
gene signature to predict response to targeted therapies in
rheumatoid arthritis. J Autoimmun. 101:121–130. 2019.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Shaw AM, Qasem A and Naser SA: Modulation
of PTPN2/22 function by spermidine in CRISPR-Cas9-edited
T-cells associated with crohn's disease and rheumatoid arthritis.
Int J Mol Sci. 22(8883)2021.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Ghorban K, Ezzeddini R, Eslami M, Yousefi
B, Sadighi Moghaddam B, Tahoori MT, Dadmanesh M and Salek Farrokhi
A: PTPN22 1858 C/T polymorphism is associated with alteration of
cytokine profiles as a potential pathogenic mechanism in rheumatoid
arthritis. Immunol Lett. 216:106–113. 2019.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Cao F, Guo C and Guo J: Deciphering CSU
pathogenesis: Network toxicologyand molecular dynamics of DOTP
exposure. Ecotoxicol Environ Saf. 291(117864)2025.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Shang L, Wang Y, Li J, Zhou F, Xiao K, Liu
Y, Zhang M, Wang S and Yang S: Mechanism of Sijunzi Decoction in
the treatment of colorectal cancer based on network pharmacology
and experimental validation. J Ethnopharmacol.
302(115876)2023.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Gong W, Sun P, Li X, Wang X, Zhang X, Cui
H and Yang J: Investigating the molecular mechanisms of resveratrol
in treating cardiometabolic multimorbidity: A network pharmacology
and bioinformatics approach with molecular docking validation.
Nutrients. 16(2488)2024.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Bottini N and Peterson EJ: Tyrosine
phosphatase PTPN22: Multifunctional regulator of immune signaling,
development, and disease. Annu Rev Immunol. 32:83–119.
2014.PubMed/NCBI View Article : Google Scholar
|
|
51
|
Buczyńska A, Sidorkiewicz I, Hryniewicka
J, Zbucka-Krętowska M, Dzięcioł J, Szelachowska M and Krętowski AJ:
Pregnancy-associated thyroid disorders: The role of genetic,
epigenetic, and oxidative stress factors. Rev Endocr Metab Disord.
26:679–692. 2025.PubMed/NCBI View Article : Google Scholar
|
|
52
|
Negro R, Gobessi S, Longo PG, He Y, Zhang
ZY, Laurenti L and Efremov DG: Overexpression of the
autoimmunity-associated phosphatase PTPN22 promotes survival of
antigen-stimulated CLL cells by selectively activating AKT. Blood.
119:6278–6287. 2012.PubMed/NCBI View Article : Google Scholar
|
|
53
|
Zhang K, Li Y, Ge X, Meng L, Kong J and
Meng X: Regulatory T cells protect against diabetic cardiomyopathy
in db/db mice. J Diabetes Investig. 15:1191–1201. 2024.PubMed/NCBI View Article : Google Scholar
|
|
54
|
Elmadbouh I and Singla DK: BMP-7
attenuates inflammation-induced pyroptosis and improves cardiac
repair in diabetic cardiomyopathy. Cells. 10(2640)2021.PubMed/NCBI View Article : Google Scholar
|
|
55
|
Fang T, Wang J, Sun S, Deng X, Xue M, Han
F, Sun B and Chen L: JinLiDa granules alleviates cardiac
hypertrophy and inflammation in diabetic cardiomyopathy by
regulating TP53. Phytomedicine. 130(155659)2024.PubMed/NCBI View Article : Google Scholar
|
|
56
|
Liang H, Yang S, Huang Y, Zhu Y, Wu Q, Wu
Z, Li S, Shi Y, Chen Z, Jin H and Wang X: PTPN22 as a therapeutic
target in intervertebral disc degeneration: Modulating mitophagy
and pyroptosis through the PI3K/AKT/mTOR axis. J Adv Res.
80:775–789. 2026.PubMed/NCBI View Article : Google Scholar
|