Introduction
Coronary heart disease (CHD) is the leading cause of
death in industrialized countries, and its prevalence is rapidly
increasing in China. Recent research has shown that inflammation
plays a critical role in CHD pathogenesis and in other
manifestations of atherosclerosis (1). Experimental evidence has confirmed
that activated T cells are implicated in atherogenesis (2,3).
Numerous studies have implicated TNFSF4/TNFRSF4 in
the activation of T cells, as well as that of macrophages (4). TNFSF4, also known as OX40L, is
primarily expressed by activated B cells, vessel endothelial cells,
macrophages, dendritic cells and certain activated T cells. This
generates costimulatory signals by interacting with TNFRSF4 on
activated T lymphocytes and enhances the proliferation and
differentiation of T lymphocytes and the development and survival
of memory T cells (5). Thus, it is
suggested that the TNFSF4-TNFRSF4 pathway plays a key role in
atherosclerosis through its participation in T cell activation.
Numerous genetic studies have indicated a
relationship between TNFSF4-TNFRSF4 and cardiovascular diseases. In
2006, Ria et al confirmed that genetic variation in
TNFRSF4 was associated with myocardial infarction (MI) in a
Swedish population (6).
Furthermore, Mashimo et al found an association between
TNFRSF4 gene polymorphisms and essential hypertension
(7). There is current evidence
showing that TNFSF4 is the gene underlying the
atherosclerosis susceptibility locus 1 (Ath1) in mice, and that
genetic polymorphisms in TNFSF4 are associated with MI and
the severity of coronary artery disease (CAD) in humans (8). A subsequent study by Malarstig et
al suggested that TNFSF4 variants are associated with
the risk of incident atherothrombosis and venous thromboembolism in
Caucasions (9). A recent study
demonstrated that interruption of the TNFSF4-TNFRSF4 pathway
attenuates atherogenesis in LDL receptor-deficient mice (10). However, despite the convincing
evidence of disease association, a study involving a Chinese Han
population is lacking. The present study aimed to replicate
previous findings. We investigated the association between genetic
variations in the TNFSF4 gene and CHD in a Chinese Han
population.
Materials and methods
Study subjects
Subjects with Han Chinese ethnicity (n=2180) were
included in this case-control study. A total of 1059 patients with
clinically manifested CHD, 682 males and 377 females, were
recruited from Qilu Hospital of Shandong University between
September 2006 and March 2010. The CHD patients were confirmed by
coronary angiography to have >50% stenosis in 1 or more coronary
arteries. Patients who had a history of hyperthyroidism, secondary
hypertension, chronic liver disease, chronic renal disease, acute
infection and hematologic diseases were excluded from the study. A
total of 1121 unrelated subjects (714 males and 407 females) were
randomly selected as controls from a health check-up center in Qilu
Hospital during the same period. The controls were free of CHD
according to medical history and showed no clear ischemic changes
by electrocardiography or symptoms of chest pain. Subjects with
congestive heart failure, peripheral vascular disease, rheumatic
heart disease, pulmonary heart disease, chronic kidney and hepatic
disease were excluded.
A structured questionnaire based on interviews and
clinical examinations was employed to characterize the patients and
the controls. These included details of medical history, family
history of CHD and other traditional risk factors of CHD, such as
drug intake, cigarette smoking and alcohol consumption. Blood
pressure, weight, height, waistline and hip circumference were also
checked, and body mass index and waist-to-hip ratio were
calculated. The clinical and demographic characteristics of the
patients are shown in Table I.
Genomic DNA was extracted from peripheral blood leukocytes by a
standard salting-out method. This study was approved by the Ethics
Committee of Shandong University School of Medicine, and informed
consent was obtained from the participants.
 | Table I.Clinical characteristics of the
coronary heart disease patients and the control subjects. |
Table I.
Clinical characteristics of the
coronary heart disease patients and the control subjects.
| Parameter | Total subjects
| Male
| Female
|
|---|
| Case | Control | P-value | Case | Control | P-value | Case | Control | P-value |
|---|
| No. | 1059 | 1121 | | 682 | 714 | | 377 | 407 | |
| Age (years) | 60.35±10.42 | 60.82±11.21 | 0.132 | 60.52±10.21 | 60.45±10.36 | 0.299 | 60.02±10.45 | 60.98±10.88 | 0.421 |
| BMI
(kg/m2) | 26.16±5.26 | 24.65±4.38 | 0.013 | 26.12±4.13 | 24.17±5.22 | 0.032 | 25.92±7.30 | 24.25±5.93 | 0.029 |
| SBP (mmHg) | 132.23±15.12 | 121.21±9.45 | 0.002 | 133.75±12.23 | 120.23±9.56 | 0.001 | 131.92±11.45 | 120.10±10.20 | 0.004 |
| DBP (mmHg) | 78.25±10.12 | 75.45±9.72 | 0.011 | 76.59±12.23 | 75.12±7.52 | 0.045 | 74.12±13.14 | 73.35±9.65 | 0.038 |
| Glu (mmol/l) | 5.92±1.43 | 5.19±0. 89 | 0.038 | 5.87±1.48 | 5.45±0.78 | 0.133 | 5.85±1.81 | 5.02±1.19 | 0.041 |
| TC (mmol/l) | 4.98±1.01 | 4.75±0.63 | 0.021 | 4.74±1. 13 | 4.69±0.83 | 0.235 | 5.10±1.12 | 4.86±0.29 | 0.154 |
| TG (mmol/l) | 1.97±1.23 | 1.11±0.59 | 0.004 | 1.93±0.87 | 1.08±0.83 | 0.021 | 2.08±1.01 | 1.08±0.62 | 0.001 |
Single-nucleotide polymorphism
selection
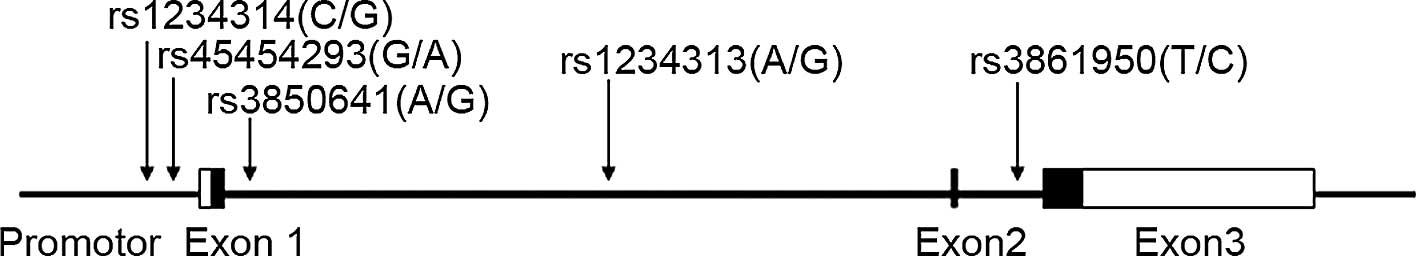
The single-nucleotide polymorphisms (SNPs) examined
here (rs3850641, rs1234313 and rs3861950) were as previously
investigated in the two case-control samples from Sweden (8). The two other SNPs (rs45454293 and
rs1234314) are located in the 5′-flanking region and may represent
potential functional variants. One of them (rs45454293) was
previously associated with incident venous thromboembolism
(9). The locations of the SNPs are
shown in Fig. 1.
Single-nucleotide polymorphism
genotyping
Initially, the SNPs were genotyped using the
PCR-RFLP method. SNPs rs3850641, rs1234314 and rs3861950 were
genotyped using the TaqMan SNP genotyping method with
assay-on-demand probes and primers (C_2955962_10 for rs1234314,
C_26492316_10 for rs3850641 and C_8920839_10 for rs3861950; Applied
Biosystems, Foster City, CA, USA) in the further recruited samples.
For the PCR-RFLP, the gene sequences harboring the five sites were
obtained from Genbank. Primers were designed with Primer Premier
5.0. The primers and restriction enzymes were: rs1234314 forward,
5′-tatc tgctgggtgcctcatg and reverse, 5′-gtcagacactgtgttagatg,
ScrF I; rs45454293 forward, 5′-ttttagtggtaaagggtacctggtgtct
and reverse, 5′-ataagctttgaagattatttctttctttgagct, Sac I;
rs3850641 forward, 5′-tttgaagctttgagtcactgatatacctggtctaccaa and
reverse, 5′-gcacgcacacattgctccgctatta, Mfe I; rs1234313
forward, 5′-cactgatgggcatttgggtt and reverse,
5′-ccactgccttgccatacct, Hpy CH4 IV; rs3861950 forward,
5′-caccctttgcccatagttc and reverse, 5′-ccttcagggagatgagataaa,
Hin cII. PCR was carried out in a 10 μl reaction volume
containing 25 ng of genomic DNA, 5 pmol of each primer, 100 μmmol/l
of each dNTP, 2 μl 5 X PCR buffer and 1 U Taq DNA polymerase.
Amplification was carried out with an initial denaturation step at
94°C for 5 min, followed by 35 cycles of denaturation at 94°C for
40 sec, annealing at a suitable temperature for 40 sec, extension
at 72°C for 50 sec with a final extension at 72°C for 10 min. The
PCR products containing the SNPs were digested with the suitable
restriction enzyme (NEB, Beijing, China). The products were
separated by electrophoresis on 1.5–2.5% agarose gel. The
genotyping accuracy in the samples was confirmed by direct
sequencing of the PCR products for certain randomly chosen
samples.
Statistical analysis
Genotype frequencies of the SNPs detected were
tested for Hardy-Weinberg equilibrium. Variations in genotype and
allelic frequencies between case and control groups, odds ratios
(OR), 95% confidence intervals (CI), and logistic regression
analysis were calculated using Plink 1.07. Continuous variables
were displayed as the mean ± standard deviation (SD), and the
comparison of continuous variables was carried out using the
Student's t-test. A p-value <0.05 was considered significant.
Bonferroni correction was used to correct the multiple test. For
the 5 SNPs each having 3 model tests included in the study, the
adjusted P value significance threshold was 0.05/(5*3)=0.003. Power
calculations were performed with the Power calculator for genetic
association analysis by Menashe et al (11). For rs3850641, we referred to a
study by Wang et al (8) in
which the relative risk of rs3850641 in myocardial infarction was
1.4. For the other SNPs, the power was calculated under the
assumption of a disease prevalence of 0.10, D-prime of 1, dominant
model and using the allelic frequency of each SNP as marker
frequency.
Results
Clinical and demographic characteristics
of the cases
The body mass index (BMI), systolic blood pressure
(SBP), diastolic blood pressure (DBP), glucose (Glu), total
cholesterol (TC) and triglyceride (TG) all demonstrated significant
variations between the cases and controls. According to the
examination results, >85% of the cases had a history of MI.
Compared with the controls, the cases had higher SBP, DBP, Glu, and
TG levels. The clinical characteristics of the study subjects are
shown in Table I.
Association of the polymorphisms with
coronary heart disease
The distributions of all five SNPs were in
Hardy-Weinberg equilibrium (P>0.05) in both the CHD and control
groups. The distributions of genotypic and allelic frequencies of
these SNPs in each group are shown in Tables II and III. Neither genotypes nor alleles
differed between the case and control groups in the overall
distribution (all P>0.05). Meanwhile, we established the
recessive, dominant models and Cochran-Armitage trend test using
PLINK; no significant differences in the models were detected
between the two groups (all P>0.05).
| Table II.Genotype frequencies and allelic
frequency distribution of rs4545293 and rs1234313. |
Table II.
Genotype frequencies and allelic
frequency distribution of rs4545293 and rs1234313.
| Total subjects
(n=1148)
| Male (n=764)
| Female (n=384)
|
|---|
| dbSNPID | Genotype allele | Case (n=547) | Control (n=601) | OR (95% CI) | P-value | Case (n=386) | Control (n=378) | OR (95% CI) | P-value | Case (n=161) | Control (n=223) | OR (95% CI) | P-value |
|---|
| rs45454293 | GG | 424 (0.78) | 447 (0.74) | 1.00 | | 302 (0.78) | 275 (0.73) | 1.00 | | 122 (0.76) | 172 (0.77) | 1.00 | |
| AG | 111 (0.20) | 141 (0.23) | 1.20 (0.91–1.60) | | 75 (0.19) | 93 (0.24) | 1.36 (0.96–1.92) | | 36 (0.22) | 48 (0.22) | 0.95 (0.58–1.54) | |
| AA | 12 (0.02) | 13 (0.03) | 1.03 (0.46–2.28) | 0.43 | 9 (0.03) | 10 (0.03) | 1.22 (0.49–3.05) | 0.21 | 3 (0.02) | 3 (0.01) | 0.71 (0.14–3.57) | 0.90 |
| G | 959 (0.88) | 1035 (0.86) | 1.00 | | 679 (0.88) | 643 (0.85) | 1.00 | | 280 (0.87) | 392 (0.88) | 1.00 | |
| A | 135 (0.12) | 167 (0.14) | 1.15 (0.90–1.46) | 0.27 | 93 (0.12) | 113 (0.15) | 1.28 (0.96–1.72) | 0.10 | 42 (0.13) | 54 (0.12) | 0.92 (0.60–1.41) | 0.70 |
| rs1234313 | AA | 245 (0.45) | 272 (0.45) | 1.00 | | 173 (0.45) | 175 (0.46) | 1.00 | | 72 (0.45) | 97 (0.44) | 1.00 | |
| AG | 240 (0.44) | 250 (0.42) | 0.94
(0.73–1.20) | | 166 (0.43) | 146 (0.39) | 0.87
(0.64–1.18) | | 74 (0.46) | 104 (0.46) | 1.04
(0.68–1.60) | |
| GG | 62 (0.11) | 79 (0.13) | 1.15
(0.79–1.67) | 0.57 | 47 (0.12) | 57 (0.15) | 1.20
(0.77–1.86) | 0.34 | 15 (0.09) | 22 (0.10) | 1.09
(0.53–2.24) | 0.97 |
| A | 730 (0.67) | 794 (0.66) | 1.00 | | 512 (0.66) | 496 (0.66) | 1.00 | | 218 (0.68) | 298 (0.67) | 1.00 | |
| G | 364 (0.33) | 408 (0.34) | 1.03
(0.87–1.23) | 0.73 | 260 (0.34) | 260 (0.34) | 1.03
(0.84–1.28) | 0.77 | 104 (0.32) | 148 (0.33) | 1.04
(0.77–1.41) | 0.80 |
| Table III.Genotype frequencies and allelic
frequency distribution of three polymorphisms. |
Table III.
Genotype frequencies and allelic
frequency distribution of three polymorphisms.
| Total subjects
(n=2180)
| Male (n=1396)
| Female (n=784)
|
|---|
| dbSNPID | Genotype
allele | Case (n=1059) | Control
(n=1121) | OR (95% CI) | P-value | Case (n=682) | Control
(n=713) | OR (95% CI) | P-value | Case (n=377) | Control
(n=407) | OR (95% CI) | P-value |
|---|
| rs1234314 | CC | 381 (0.36) | 370 (0.33) | 1.00 | | 243 (0.36) | 236 (0.33) | 1.00 | | 138 (0.37) | 134 (0.33) | 1.00 | |
| CG | 476 (0.45) | 527 (0.47) | 1.14
(0.94–1.38) | | 308 (0.45) | 328 (0.46) | 1.10
(0.86–1.39) | | 168 (0.44) | 199 (0.49) | 1.22
(0.89–1.67) | |
| GG | 202 (0.19) | 224 (0.20) | 1.14
(0.90–1.45) | 0.34 | 131 (0.19) | 150 (0.21) | 1.18
(0.88–1.58) | 0.53 | 71 (0.19) | 74 (0.18) | 1.07
(0.72–1.61) | 0.45 |
| C | 1238 (0.58) | 1267 (0.57) | 1.00 | | 794 (0.58) | 800 (0.56) | 1.00 | | 444 (0.59) | 467 (0.57) | 1.00 | |
| G | 880 (0.42) | 975 (0.43) | 1.08
(0.96–1.22) | 0.20 | 570 (0.42) | 628 (0.44) | 1.09
(0.94–1.27) | 0.24 | 310 (0.41) | 347 (0.43) | 1.06
(0.87–1.30) | 0.54 |
| rs3850641 | AA | 773 (0.73) | 784 (0.70) | 1.00 | | 504 (0.74) | 489 (0.68) | 1.00 | | 269 (0.71) | 295 (0.72) | 1.00 | |
| AG | 254 (0.24) | 314 (0.28) | 1.22
(1.00–1.48) | | 157 (0.23) | 209 (0.29) | 1.37
(1.08–1.75) | | 97 (0.26) | 105 (0.26) | 0.99
(0.72–1.36) | |
| GG | 32 (0.03) | 23 (0.02) | 0.71
(0.41–1.22) | 0.05 | 21 (0.03) | 16 (0.02) | 0.79
(0.40–1.52) | 0.02 | 11 (0.03) | 7 (0.02) | 0.58
(0.22–1.52) | 0.53 |
| A | 1800 (0.85) | 1882 (0.84) | 1.00 | | 1165 (0.85) | 1187 (0.83) | 1.00 | | 635 (0.84) | 695 (0.85) | 1.00 | |
| G | 318 (0.15) | 360 (0.16) | 1.08
(0.92–1.28) | 0.34 | 199 (0.15) | 241 (0.17) | 1.19
(0.97–1.46) | 0.09 | 119 (0.16) | 119 (0.15) | 0.91
(0.69–1.20) | 0.52 |
| rs3861950 | TT | 911 (0.86) | 953 (0.85) | 1.00 | | 593 (0.87) | 592 (0.83) | 1.00 | | 318 (0.84) | 361 (0.89) | 1.00 | |
| TC | 137 (0.13) | 157 (0.14) | 1.10
(0.86–1.40) | | 82 (0.12) | 115 (0.16) | 1.40
(1.04–1.91) | | 55 (0.15) | 42 (0.10) | 0.67
(0.44–1.03) | |
| CC | 11 (0.01) | 11 (0.01) | 0.96
(0.41–2.22) | 0.76 | 7 (0.01) | 7 (0.01) | 1.00
(0.35–2.87) | 0.09 | 4 (0.01) | 4 (0.01) | 0.88
(0.22–3.55) | 0.19 |
| T | 1959 (0.92) | 2063 (0.92) | 1.00 | | 1268 (0.93) | 1299 (0.91) | 1.00 | | 691 (0.92) | 764 (0.94) | 1.00 | |
| C | 159 (0.08) | 179 (0.08) | 1.07
(0.86–1.34) | 0.56 | 96 (0.07) | 129 (0.09) | 1.31
(1.00–1.73) | 0.05 | 63 (0.08) | 50 (0.06) | 0.72
(0.49–1.06) | 0.09 |
Case-only association study
A case-only study was conducted to determine the
effect of TNFSF4 variations on disease severity. The
patients were divided into three subgroups according to the numbers
of coronary arteries involved. The frequencies of the five SNPs in
each group were compared. As shown in Table IV, there were no significant
differences in the distribution of genotypic and allelic
frequencies between each group. Although the p-value for rs3861950
between single-vessel and double-vessel, and single-vessel vs.
triple-vessel was <0.05, the significance threshold was not
reached after adjustment.
| Table IV.Genotype frequencies and allelic
frequencies in cases with a different number of coronary artery
involved vessels. |
Table IV.
Genotype frequencies and allelic
frequencies in cases with a different number of coronary artery
involved vessels.
| rs1234314 | rs45454293 | rs3850641 | rs1234313 | rs3861950 |
|---|
| Single-vessel
(266) | CC | 93 (0.35) | GG | 109 (0.78) | AA | 186 (0.70) | AA | 55 (0.40) | TT | 221 (0.83) |
| CG | 125 (0.47) | AG | 27 (0.19) | AG | 69 (0.26) | AG | 64 (0.46) | TC | 40 (0.15) |
| GG | 48 (0.18) | AA | 3 (0.03) | GG | 11 (0.04) | GG | 20 (0.14) | CC | 5 (0.02) |
| C | 311 (0.58) | G | 245 (0.88) | A | 441 (0.83) | A | 174 (0.63) | T | 482 (0.91) |
| G | 221 (0.42) | A | 33 (0.12) | G | 91 (0.17) | G | 104 (0.37) | C | 50 (0.09) |
| Double-vessel
(290) | CC | 107 (0.37) | GG | 114 (0.78) | AA | 209 (0.72) | AA | 73 (0.50) | TT | 255 (0.88) |
| CG | 130 (0.45) | AG | 29 (0.20) | AG | 70 (0.24) | AG | 58 (0.40) | TC | 26 (0.09) |
| GG | 53 (0.18) | AA | 3 (0.02) | GG | 11 (0.04) | GG | 15 (0.10) | CC | 9 (0.03) |
| C | 344 (0.60) | G | 257 (0.88) | A | 488 (0.84) | A | 204 (0.70) | T | 536 (0.92) |
| G | 236 (0.40) | A | 35 (0.12) | G | 92 (0.16) | G | 88 (0.30) | C | 44 (0.08) |
| Triple-vessel
(501) | CC | 185 (0.37) | GG | 194 (0.76) | AA | 371 (0.74) | AA | 116 (0.46) | TT | 436 (0.87) |
| CG | 220 (0.44) | AG | 54 (0.21) | AG | 115 (0.23) | AG | 115 (0.45) | TC | 60 (0.12) |
| GG | 96 (0.19) | AA | 6 (0.03) | GG | 15 (0.03) | GG | 23 (0.09) | CC | 5 (0.01) |
| C | 596 (0.59) | G | 442 (0.87) | A | 857 (0.86) | A | 347 (0.68) | T | 932 (0.93) |
| G | 412 (0.41) | A | 66 (0.13) | G | 145 (0.14) | G | 161 (0.32) | C | 70 (0.07) |
| Control (1121) | CC | 370 (0.33) | GG | 447 (0.74) | AA | 784 (0.70) | AA | 272 (0.45) | TT | 953 (0.85) |
| CG | 527 (0.47) | AG | 141 (0.23) | AG | 314 (0.28) | AG | 250 (0.42) | TC | 157 (0.14) |
| GG | 224 (0.20) | AA | 13 (0.03) | GG | 23 (0.02) | GG | 79 (0.13) | CC | 11 (0.01) |
| C | 1267 (0.57) | G | 1035 (0.86) | A | 1882 (0.84) | A | 794 (0.66) | T | 2063 (0.92) |
| G | 975 (0.43) | A | 167 (0.14) | G | 360 (0.16) | G | 408 (0.34) | C | 179 (0.08) |
|
| S vs. C | | | | | | | | | | |
| P-value | | 0.42 | | 0.37 | | 0.56 | | 0.27 | | 0.29 |
| OR | | 1.08 | | 1.20 | | 0.93 | | 0.86 | | 0.84 |
| 95% CI | | (0.89–1.31) | | (0.80–1.78) | | (0.72–1.19) | | (0.66–1.13) | | (0.60–1.16) |
| D vs. C | | | | | | | | | | |
| P-value | | 0.22 | | 0.39 | | 0.91 | | 0.22 | | 0.75 |
| OR | | 1.12 | | 1.18 | | 1.01 | | 1.19 | | 1.06 |
| 95% CI | | (0.93–1.35) | | (0.80–1.75) | | (0.79–1.30) | | (0.90–1.57) | | (0.75–1.49) |
| T vs. C | | | | | | | | | | |
| P-value | | 0.16 | | 0.62 | | 0.25 | | 0.37 | | 0.32 |
| OR | | 1.11 | | 1.08 | | 1.13 | | 1.11 | | 1.16 |
| 95% CI | | (0.96–1.29) | | (0.80–1.47) | | (0.92–1.39) | | (0.89–1.38) | | (0.87–1.54) |
| S vs. D | | | | | | | | | | |
| P-value | | 0.77 | | 0.99 | | 0.58 | | 0.07 | | 0.28 |
| OR | | 0.97 | | 0.98 | | 0.91 | | 1.39 | | 0.79 |
| 95% CI | | (0.76–1.23) | | (0.60–1.64) | | (0.67–1.25) | | (0.98–1.96) | | (0.52–1.21) |
| D vs. T | | | | | | | | | | |
| P-value | | 0.94 | | 0.68 | | 0.46 | | 0.65 | | 0.66 |
| OR | | 1.01 | | 0.91 | | 0.90 | | 1.08 | | 0.91 |
| 95% CI | | (0.82–1.24) | | (0.59–1.41) | | (0.68–1.19) | | (0.79–1.47) | | (0.62–1.35) |
| S vs. T | | | | | | | | | | |
| P-value | | 0.80 | | 0.65 | | 0.17 | | 0.59 | | 0.09 |
| OR | | 0.97 | | 1.12 | | 0.82 | | 0.92 | | 0.72 |
| 95% CI | | (0.79–1.20) | | (0.71–1.73) | | (0.62–1.09) | | (0.67–1.26) | | (0.50–1.06) |
Power calculation
For rs3850641, our sample size had the power of
>90% when α was 0.05 under that assumption. For the other four
SNPs, our sample size had ≥80% power to detect the effect of a
relative risk of 1.4.
Discussion
Atherosclerosis is characterized by a complex
multifactorial pathophysiology. It was formerly considered a
disease of lipid accumulation, yet it actually involves an ongoing
inflammatory response. Recent research has demonstrated that
inflammation plays a pivotal role in the pathogenesis of
atherosclerotic coronary artery disease. Vascular inflammation
plays a predominant role in the initiation, progression and final
steps of atherosclerosis (12).
Numerous studies have shown that the TNFSF4-TNFRSF4 pathway plays a
critical role in the pathogenesis of cardiovascular diseases.
We first investigated the association of the
TNFSF4 gene and CHD using 547 patients and 601 controls. No
significant differences were found in the genotypic and allelic
frequency distribution between the two groups in the Chinese Han
population studied. Moreover, even after adjusting for age, gender,
BMI, SBP, DBP, Glu, TC and TG, no significant differences were
found for the SNPs typed. To exclude falsenegative results due to
the small sample size, we genotyped rs3850641, rs1234313 and
rs3861950 in additional samples using the TaqMan assay method. In
the Stockholm Coronary Atherosclerosis Risk Factor (SCARF) study,
the minor allele of SNP rs3850641 in TNFSF4 was found to be
associated with increased risk of MI in women (8). However, in our study, significant
differences in genotypes were not found in either gender for the
five SNPs, even after adjusting for age, gender, BMI, SBP, DBP,
Glu, TC and TG.
In a previous study, the minor allele of rs3850641
in TNFSF4 was significantly more frequent in individuals
with MI than in the controls in two independent human cohorts
(8), while no significant
difference was observed in our samples, and the frequency of the
minor allele was even slightly higher in the controls. Also, no
significant difference was found between the allelic frequencies in
our data and the Han Chinese in Beijing (HCB) data from Hapmap.
Analysis of the linkage disequilibrium (LD) structure using
Haploview showed there is not much difference in the LD of the
TNFSF4 gene between HCB and Utah residents of northern and
western European ancestry.
A number of possibilities may account for the lack
of association between the SNPs and CHD in this case-control study,
including diagnostic heterogeneity, sample size, population
stratification and various genetic backgrounds in the various
populations. To increase the homogeneity of our patients, we also
performed a case-only association study; the genotypic and allelic
frequencies did not indicate any trend in difference between the
groups. With our sample size, we expected a power of at least 80%
in detecting an effect of relative risk ≥1.4 for each of the SNPs
even at α=0.05. Therefore, our failure to detect an association
between CHD and these five SNPs may not have been due to the sample
size.
Our results suggest that the SNPs studied in the
TNFSF4 gene are unlikely to contribute to the CHD risk in
the Chinese Han population. These results were consistent with
those of Koch et al who did not find any association between
the TNFSF4 gene and MI in a German population (13). In addition, Olofsson et al,
in an association study of TNFSF4 gene variations with the
risk for ischemic stroke using expression analysis, no significant
association was found (14).
In conclusion, the present study aimed to determine
whether the TNFSF4 gene is associated with CHD in a Chinese
Han population. We did not find any significant association between
five SNPs studied and CHD in our samples. Our results suggest that
the TNFSF4 gene is unlikely to be a major susceptibility
gene for CHD in our population.
Acknowledgements
This work was supported by grants of
the National Natural Science Foundation of China (no. 30671956),
the National Basic Research Program of China (973 Program) grant
2007CB512001 and the National High-Tech Research and Development
Program of China; grant no. 2006AA02A406.
References
|
1.
|
Hansson GK: Inflammation, atherosclerosis,
and coronary artery disease. N Engl J Med. 352:1685–1695. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
2.
|
Hansson GK, Libby P, Schonbeck U and Yan
ZQ: Innate and adaptive immunity in the pathogenesis of
atherosclerosis. Circ Res. 91:281–291. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
3.
|
De Boer OJ, Becker AE and van der Wal AC:
T lymphocytes in atherogenesis-functional aspects and antigenic
repertoire. Cardiovasc Res. 60:78–86. 2003.PubMed/NCBI
|
|
4.
|
Walker LS, Gulbranson-Judge A, Flynn S,
Brocker T and Lane PJ: Co-stimulation and selection for T-cell help
for germinal centres: The role of CD28 and OX40. Immunol Today.
21:333–337. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
5.
|
Weinberg AD: Ox40: targeted immunotherapy
– implications for tempering autoimmunity and enhancing vaccines.
Trends Immunol. 23:102–109. 2002.
|
|
6.
|
Ria M, Eriksson P, Boquist S, Ericsson CG,
Hamsten A and Lagercrantz J: Human genetic evidence that OX40 is
implicated in myocardial infarction. Biochem Biophys Res Commun.
339:1001–1006. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
7.
|
Mashimo Y, Suzuki Y, Hatori K, Tabara Y,
Miki T, Tokunaga K, Katsuya T, Ogihara T, Yamada M, Takahashi N,
Makita Y, Nakayama T, Soma M, Hirawa N, Umemura S, Ohkubo T, Imai Y
and Hata A: Association of TNFRSF4 gene polymorphisms with
essential hypertension. J Hypertens. 26:902–913. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8.
|
Wang X, Ria M, Kelmenson PM, Eriksson P,
et al: Positional identification of TNFSF4, encoding OX40 ligand,
as a gene that influences atherosclerosis susceptibility. Nat
Genet. 37:365–372. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
9.
|
Malarstig A, Eriksson P, Rose L, Diehl KA,
Hamsten A, Ridker PM and Zee RY: Genetic variants of tumor necrosis
factor superfamily, member 4 (TNFSF4), and risk of incident
atherothrombosis and venous thromboembolism. Clin Chem. 54:833–840.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
10.
|
Van Wanrooij EJ, van Puijvelde GH, de Vos
P, Yagita H, van Berkel TJ and Kuiper J: Interruption of the
TNFRSF4/TNFSF4 (OX40/OX40l) pathway attenuates atherogenesis in
low-density lipoprotein receptor-deficient mice. Arterioscler
Thromb Vasc Biol. 27:204–210. 2007.PubMed/NCBI
|
|
11.
|
Menashe I, Rosenberg PS and Chen BE: PGA:
Power calculator for case-control genetic association analyses. BMC
Genet. 9:362008. View Article : Google Scholar : PubMed/NCBI
|
|
12.
|
Hansson GK, Robertson AK and
Soderberg-Naucler C: Inflammation and atherosclerosis. Annu Rev
Pathol. 1:297–329. 2006. View Article : Google Scholar
|
|
13.
|
Koch W, Hoppmann P, Mueller JC, Schomig A
and Kastrati A: Lack of support for association between common
variation in TNFSF4 and myocardial infarction in a German
population. Nat Genet. 40:1386–1388. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14.
|
Olofsson PS, Soderstrom LA, Jern C, Sirsjo
A, Ria M, Sundler E, de Faire U, Wiklund PG, Ohrvik J, Hedin U,
Paulsson-Berne G, Hamsten A, Eriksson P and Hansson GK: Genetic
variants of TNFSF4 and risk for carotid artery disease and stroke.
J Mol Med. 87:337–346. 2009. View Article : Google Scholar : PubMed/NCBI
|