Introduction
Immunoglobulin A nephropathy (IgAN) is the most
common form of primary glomerulonephritis (GN). Its hallmark is
deposition of polymeric IgA1 in the glomerular mesangium
accompanied by mesangial proliferative GN (1). The mechanisms involved in mesangial
IgA deposition and the initiation of inflammatory glomerular injury
remain unclear. The degree of histopathologic injury is extremely
variable; this is reflected in the varied pace and severity of
clinical presentation noted in this disease. Genetic factors
contribute to the development of IgAN and the progression to renal
failure, including the rate of IgA deposition, the phenotype of
mesangial response and the risk of progressive renal failure
(2).
Bone morphogenetic proteins (BMPs) are the largest
group of members of the transforming growth factor β (TGFB)
superfamily and were originally identified as bone-inducer proteins
(3). In recent years, the
importance of BMP signaling has been linked to the morphogenesis,
cell proliferation and differentiation, apoptosis and patterning of
various organs (4). In the kidney,
several BMPs are expressed during organogenesis of the metanephric
kidney, and in postnatal life BMPs have been shown to be important
in both preservation of kidney function and resistance to injury
(5).
BMP2 is a specific member of the BMPs and is highly
regulated in the kidney (6).
Moreover, high affinity binding sites for BMP2 have been identified
in kidney epithelial cells (7). A
few studies have suggested associations between BMP2 and renal
diseases. In mesangial cells, BMP2 has been shown to inhibit
platelet-derived growth factor (PDGF)-induced c-fos gene
transcription and epidermal growth factor (EGF)-induced Elk
transcription as well as DNA synthesis mediated by
mitogen-activated protein kinase (MAPK) (8,9). In
podocytes, BMP2 induces reactive oxygen species via up-regulation
of Id-1 expression, which subsequently inhibits cellular functions
(10). Notably, together with
BMP7, BMP2 has been demonstrated to have the same effect on renal
fibrosis by promoting catabolism of type I TGFB receptors (6).
However, the association between IgAN and the BMP2
gene has not yet been reported. With regard to the associations
between IgAN and BMPs, only BMP7 has been reported by Chan et
al, who demonstrated that BMP7 has a beneficial effect on IgAN
by ameliorating IgAN-derived polymeric IgA-induced tumor necrosis
factor (TNF) (anti-inflammatory effect) and lymphotoxin α (TNF
superfamily, member 1, LTA) synthesis (anti-fibrotic effect) in
human mesangial cells (11).
In the present study, we investigated the
association between IgAN and polymorphisms of the BMP2 gene.
Materials and methods
Subjects
We examined 187 Korean pediatric patients with
biopsy-confirmed IgAN and 262 healthy controls. Almost all of the
patients were identified through a urinary screening program at
schools in Korea. Most of the patients showed no symptoms of renal
disease and were thought to be at a relatively early stage of the
disease. Patients that fulfilled one or more of the following
criteria underwent a renal biopsy: i) unexplained prolonged
isolated hematuria or proteinuria with a duration of >12 months;
ii) concomitant hematuria and proteinuria with a duration of >3
months; iii) hematuria or proteinuria with decreased serum C3 and
C4 levels; or iv) decreased renal function. Healthy controls were
also recruited based on the findings of a routine screening. The
screening included the completion of a questionnaire that included
the following: symptoms and medical history, blood pressure,
electrocardiography, abdominal sonography and laboratory tests,
such as complete blood count, fasting glucose level, total
cholesterol, triglyceride levels, high-density lipoprotein
cholesterol levels, rheumatoid factor, hepatitis B and C viral
markers, hemoglobin A1C, liver enzymes, blood urea nitrogen,
creatinine, electrolytes and a urinalysis (protein, occult blood
and glucose). Controls with any abnormal results were excluded. In
this study, we did not age-match controls and patients. One of the
confounding factors in genetic analysis is the presence of
subclinical IgAN in supposedly normal control populations (2). IgAN is known to develop in young
adults in their twenties. For this reason, healthy controls >20
years of age were needed to exclude the possibility of subclinical
IgAN. Therefore, the presence of an age gap between the two groups
appears unavoidable. This study was approved by the Ethics Review
Committee of the Medical Research Institute, Kyung Hee University
Medical Center, Seoul, Korea. Written informed consent was obtained
from all of the subjects.
Patient subgroups
To determine the nature of the association between
single nucleotide polymorphisms (SNPs) of BMP2 and the patient
phenotype and histopathological findings, patients were divided
into subgroups according to the following laboratory findings: i)
the presence of proteinuria at the time of the initial renal biopsy
(proteinuria ≤4 or >4 mg/m2/h); ii) the presence of
gross hematuria. Subgroups were also characterized on the basis of
the following: iii) the presence of podocyte foot process
effacement upon renal biopsy; iv) disease stage as assessed by
pathological markers upon renal biopsy, such as interstitial
fibrosis, tubular atrophy or global sclerosis (subjects who had one
or more markers were placed in the advanced group and subjects
without any of the pathological markers were placed in the early
group); and v) the degree of mesangial proliferation (subjects with
normal to mild focal mesangial proliferation were allocated to the
mild group and those with mild diffuse to severe diffuse mesangial
proliferation were allocated to the advanced group). All the
demographic characteristics of the IgAN patients are documented in
Table I.
 | Table I.Demographic characteristics of the
participating subjects. |
Table I.
Demographic characteristics of the
participating subjects.
| IgAN | Control |
|---|
| No. of subjects
(n) | 187 | 262 |
| Age (mean ± SD),
years | 38.2±13.8 | 12.4±4.5 |
| Proteinuria
(n)a | | |
| >4
mg/m2/h | 88 | |
| ≤4
mg/m2/h | 99 | |
| Gross hematuria
(n) | | |
| + | 35 | |
| – | 152 | |
| Podocyte foot process
effacement (n) | | |
| + | 78 | |
| – | 109 | |
| Advanced pathological
grading (n)b | | |
| Advanced | 23 | |
| Mild | 164 | |
| Mesangial
proliferation (n)c | | |
| Advanced | 65 | |
| Mild | 122 | |
SNP selection and genotyping
We searched for coding SNPs (cSNPs) in the BMP2 gene
based on the results of database searches (http://www.ncbi.nlm.nih.gov/SNP, dbSNP BUILD 131).
Among the SNPs in the coding regions, we excluded those with
heterozygosity <0.1 (rs2273073, rs79417223, rs2273074,
rs34183594 and rs11545591) or without a genotype frequency
(rs13037675). Finally, rs1049007 (synonymous, Ser87Ser) and
rs235768 (missense, Arg190Ser) were selected. The heterozygosities
of rs1049007 and rs235768 were 0.397 and 0.382, respectively.
DNA was isolated from peripheral blood samples using
the Roche DNA Extraction kit (Roche, Indianapolis, IN, USA). SNP
genotyping was performed by direct sequencing. Genomic DNA was
amplified using primers specific for each cSNP of the BMP2 gene
(Table II). The samples were
sequenced using ABI PRISM 3730XL analyzer (PE Applied Biosystems,
USA). Sequence data were analyzed using SeqManII software (DNAStar,
Madison, WI, USA).
| Table II.Sequences of the primers used for each
SNP in the BMP2 gene. |
Table II.
Sequences of the primers used for each
SNP in the BMP2 gene.
| SNP | Sequence (5′-3′) | Product size
(bp) | Temperature (°C) |
|---|
| rs235768 | Sense |
TTATCACCTCAGCAGAGCTTCA | 375 | 58 |
| Anti-sense |
GGCCAAAAGTTACTAGCAATGG | | |
| rs1049007 | Sense |
GACGAGGTCCTGAGCGAGTTCG | 339 | 58 |
| Anti-sense |
TACAGAAGCAAGAGTGGAAACG | | |
Statistical analysis
For the case-control association study, all cSNPs in
the cases and controls were assessed for Hardy-Weinberg equilibrium
(HWE) using SNPstats software (Biostatistics and Bioinformatics
Unit, Barcelona, Spain) (12).
Logistic regression analysis was used to calculate the odds ratios
(OR), 95% confidence intervals (CIs) and p-values. For multiple
logistic regression analyses to determine associations of single
SNPs with IgAN, multiple inheritance models, including codominant 1
(major allele homozygotes vs. heterozygotes), codominant 2 (major
allele homozygotes vs. minor allele homozygotes), dominant (major
allele homozygotes vs. minor allele homozygotes plus heterozygotes)
and recessive (major allele homozygotes plus heterozygotes vs.
minor allele homozygotes) models were used. For comparisons between
the patients with IgAN and healthy controls, the data were adjusted
for gender only, and for comparisons among the IgAN subgroups,
gender and age were controlled. Allelic frequencies of each cSNP
were compared by Pearson's Chi-square test. A linkage
disequilibrium (LD) block of polymorphisms was tested using
Haploview version 4.1 (Broad Institute, Cambridge, MA, USA).
SNPstats and HapAnalyzer Pro version 1.0 (http://www.hap.ngri.go.kr), and HelixTree (Golden
Helix Inc., Bezemen, MT, USA) were used to analyze the association
between SNPs and haplotypes. Statistical analyses were performed
using the SPSS statistics 18.0 software package (SPSS Inc.,
Chicago, IL, USA), and a p-value of <0.05 was considered
statistically significant. To avoid chance findings due to multiple
testing, the Bonferroni correction was applied by lowering the
significance levels to p=0.025 (p=0.05/2) for the two SNPs and for
the two haplotypes.
Results
We investigated whether or not BMP2 gene
polymorphisms are related to IgAN by genotyping two cSNPs in Korean
children. The genetic distributions of all cSNPs in this study were
in agreement with HWE (p>0.05).
As shown in Table
III, the genotyping data revealed that two cSNPs in the coding
region were significantly associated with IgAN. The rs235768 was
associated with IgAN in the codominant model II [p=0.01; OR (95%
CI), 0.08 (0.01–0.57)] and in the recessive model [p=0.0002; OR
(95% CI), 0.07 (0.01–0.55)]. A significant association was also
observed between IgAN and rs1049007 in the codominant model II
[p=0.02; OR (95% CI), 0.16 (0.04–0.70)] and in the recessive model
[p=0.0023; OR (95% CI), 0.16 (0.04–0.69)]. These results remained
significant after Bonferroni correction (p<0.025). However,
allelic frequencies were not associated with IgAN (Table III). Comparisons of the subgroups
did not show any significant associations with the cSNPs (data not
shown).
| Table III.Logistic regression analysis of
genotype and allelic frequencies of BMP2 gene polymorphisms in IgAN
patients and controls. |
Table III.
Logistic regression analysis of
genotype and allelic frequencies of BMP2 gene polymorphisms in IgAN
patients and controls.
| SNP | Type | Control, n (%) | IgAN, n (%) | Model | OR (95% CI) | p-value |
|---|
| rs235768 (missense
Arg190Ser) | Genotype | | | | | |
| TT | 165 (62.98) | 123 (65.77) | Codominant I | 1.10 (0.73–1.65) | 0.6500 |
| TA | 79 (30.15) | 63 (33.70) | Codominant II | 0.08
(0.01–0.57) | 0.0100 |
| AA | 18 (6.87) | 1 (0.53) | Dominant | 0.91 (0.61–1.35) | 0.6200 |
| | | Recessive | 0.07
(0.01–0.55) | 0.0002 |
| Allele | | | | | |
| T | 409 (78.05) | 309 (82.62) | | 0.75 (0.53–1.05) | 0.0900 |
| A | 115 (21.95) | 65 (17.38) | | | |
| rs1049007
(synonymous Ser87Ser) | Genotype | | | | | |
| GG | 168 (64.12) | 127 (67.91) | Codominant I | 1.02
(0.68–0.04) | 0.9200 |
| GA | 77 (29.39) | 58 (31.02) | Codominant II | 0.16
(0.04–0.70) | 0.0200 |
| AA | 17 (6.49) | 2 (1.07) | Dominant | 0.86
(0.58–1.29) | 0.4700 |
| | | Recessive | 0.16
(0.04–0.69) | 0.0023 |
| Allele | | | | | |
| G | 413 (78.82) | 312 (83.42) | | 0.74
(0.52–1.04) | 0.0800 |
| A | 111 (21.18) | 62 (16.58) | | | |
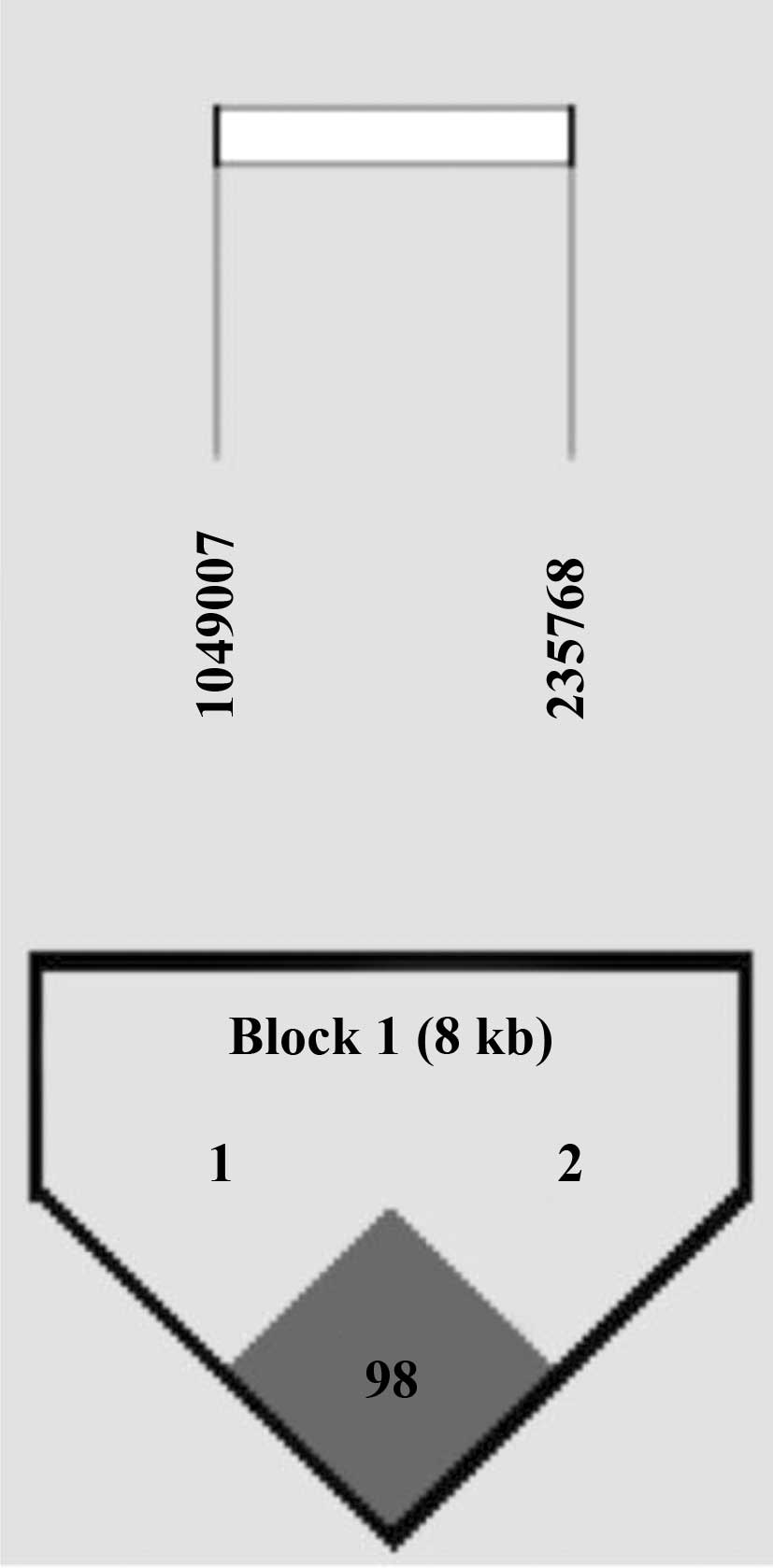
Through LD block analysis using the Gabriel method
(13), we found that an LD block
was made between two cSNPs (Fig.
1). In haplotype analysis, haplotypes TG [p=0.01; OR (95% CI),
6.76 (1.55–29.50) in the dominant model] and AA [p=0.01; OR (95%
CI), 0.08 (0.01–0.59) in the recessive model] were associated with
IgAN (Table IV). The significant
association of haplotypes with IgAN remained after Bonferroni
correction (p<0.025).
| Table IV.Haplotype analysis of BMP2 gene
polymorphisms in the IgAN patients and controls. |
Table IV.
Haplotype analysis of BMP2 gene
polymorphisms in the IgAN patients and controls.
| Haplotype | Type | Control
| IgAN
| Model | OR (95% CI) | p-value |
|---|
| Frequency | n (%) | Frequency | n (%) |
|---|
| TG | H/H | 0.78 | 164 (62.36) | 0.83 | 123 (66.13) | Codominant | 1.34
(0.96–1.88) | 0.08 |
| H/- | | 81 (30.80) | | 61 (32.80) | Dominant | 6.76
(1.55–29.5) | 0.01 |
| -/- | | 18 (6.84) | | 2 (1.07) | Recessive | 1.18
(0.80–1.75) | 0.41 |
| AA | H/H | 0.21 | 17 (6.46) | 0.16 | 1 (0.54) | Codominant | 0.72
(0.51–1.02) | 0.07 |
| H/- | | 77 (29.28) | | 58 (31.18) | Dominant | 0.84
(0.56–1.24) | 0.38 |
| -/- | | 169 (64.26) | | 127 (68.28) | Recessive | 0.08
(0.01-0.59) | 0.01 |
We calculated the sample power to verify our data,
using a genetic power calculator (http://pngu.mgh.harvard.edu/~purcell/gpc/cc2.html). In
our case-control study, sample powers for the SNPs were 0.825
(rs235768, number of cases for 70% power = 175) and 0.814
(rs1049007, number of cases for 70% power = 180), respectively
(α=0.05, genotype relative risk 2-fold). Therefore, our
case-control study was sufficiently powered for rs235768 and
rs1049007 to determine a positive association.
Discussion
Bone morphogenetic proteins are multi-functional
growth factors and belong to the TGFB superfamily (3). Their functions include the regulation
of embryonic development and cellular homeostasis, such as the
regulation of proliferation, differentiation, apoptosis and
remodeling of the extracellular matrix. Dysregulation of BMPs has
been documented in several types of diseases, including cancer,
diabetes, fibrosis and renal, auto-immune and cardiovascular
diseases (4,14–17).
BMP2 was originally identified as an essential
inducer for bone development and repair (18). Clinically, BMP2 has been used for
various therapeutic interventions, such as bone defects, non-union
fracture, spinal fusion, osteoporosis and root canal surgery
(3). Apart from bone disease, BMP2
has been shown to be associated with invasion and metastasis of
several tumor cell lines (15,19,20)
and is used as a prognostic marker for specific tumors (21). Moreover, it is involved in vascular
physiology as a proatherogenic mediator in the arterial wall
(4). However, limited data have
been reported for the association between BMP2 and renal diseases
(6,22,23).
In the present study, the genotypes of rs235768
(missense, Arg190Ser) and rs1049007 (synonymous, Ser87Ser) were
associated with childhood IgAN in a case-control analysis. In
addition, the TG and AA haplotypes consisting of rs235768 and
rs1049007 were associated with IgAN. However, the mechanisms by
which they are involved in the development of IgAN are not yet
known.
BMPs exert their biologic actions via transmembrane
serine/threonine kinase receptors. Binding of BMP to its receptor
stimulates oligomerization of the receptor complex. According to
the different oligomerization modes, BMP2 activates distinct
signaling pathways, including the classical Smad pathway and
non-Smad pathways, such as MAPK signaling through transcriptional
effects, and also has non-transcriptional effects, such as
cytoskeletal rearrangements (24).
Previous in vitro studies have demonstrated that BMP2
inhibits MAPK-dependent gene transcription and DNA synthesis in
mesangial cells (8,9). Further studies that discriminate
between the distinct pathways in the kidneys of IgAN patients with
such polymorphisms may aid in the understanding of IgAN
pathogenesis and present the possibility of individualized gene or
cell therapy (4).
Additionally, we analyzed the association between
BMP2 gene polymorphisms and disease progression using clinical and
histological parameters. However, neither of the two SNPs showed an
association with any of the parameters for disease progression. One
possible reason is that our patients were enrolled through urine
screening tests; therefore, most of them showed no symptoms and had
relatively mild GN. Further studies with subjects in various stages
of IgAN are required to evaluate the association with disease
progression.
In conclusion, this study is the first to evaluate
the association of BMP2 gene polymorphisms with childhood IgAN. The
rs235768 and rs1049007 cSNPs of the BMP2 gene may be associated
with IgAN in the Korean population.
Acknowledgements
This study was supported by the
Program at Kyung Hee University for the Young Researcher of Medical
Science in 2009 (KHU-20091456).
References
|
1.
|
Barratt J and Feehally J: IgA nephropathy.
J Am Soc Nephrol. 16:2088–2097. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
2.
|
Barratt J, Smith AC, Molyneux K and
Feehally J: Immunopathogenesis of IgAN. Semin Immunopathol.
29:427–443. 2007. View Article : Google Scholar
|
|
3.
|
Chen D, Zhao M and Mundy GR: Bone
morphogenetic proteins. Growth Factors. 22:233–241. 2004.
View Article : Google Scholar
|
|
4.
|
Maciel TT, Kempf H and Campos AH:
Targeting bone morphogenetic protein signaling on renal and
vascular diseases. Curr Opin Nephrol Hypertens. 19:26–31. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
5.
|
Simic P and Vukicevic S: Bone
morphogenetic proteins in development and homeostasis of kidney.
Cytokine Growth Factor Rev. 16:299–308. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6.
|
Yang YL, Liu YS, Chuang LY, et al: Bone
morphogenetic protein-2 antagonizes renal interstitial fibrosis by
promoting catabolism of type I transforming growth factor-beta
receptors. Endocrinology. 150:727–740. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7.
|
Iwasaki S, Tsuruoka N, Hattori A, Sato M,
Tsujimoto M and Kohno M: Distribution and characterization of
specific cellular binding proteins for bone morphogenetic
protein-2. J Biol Chem. 270:5476–5482. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
8.
|
Ghosh Choudhury G, Kim YS, Simon M, et al:
Bone morphogenetic protein 2 inhibits platelet-derived growth
factor-induced c-fos gene transcription and DNA synthesis in
mesangial cells. Involvement of mitogen-activated protein kinase. J
Biol Chem. 274:10897–10902. 1999.
|
|
9.
|
Ghosh Choudhury G, Jin DC, Kim Y, Celeste
A, Ghosh-Choudhury N and Abboud HE: Bone morphogenetic protein-2
inhibits MAPK-dependent Elk-1 transactivation and DNA synthesis
induced by EGF in mesangial cells. Biochem Biophys Res Commun.
258:490–496. 1999.PubMed/NCBI
|
|
10.
|
Pache G, Schafer C, Wiesemann S, et al:
Upregulation of Id-1 via BMP-2 receptors induces reactive oxygen
species in podocytes. Am J Physiol Renal Physiol. 291:F654–F662.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
11.
|
Chan WL, Leung JC, Chan LY, Tam KY, Tang
SC and Lai KN: BMP-7 protects mesangial cells from injury by
polymeric IgA. Kidney Int. 74:1026–1039. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12.
|
Sole X, Guino E, Valls J, Iniesta R and
Moreno V: SNPStats: a web tool for the analysis of association
studies. Bioinformatics. 22:1928–1929. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13.
|
Gabriel SB, Schaffner SF, Nguyen H, et al:
The structure of haplotype blocks in the human genome. Science.
296:2225–2229. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
14.
|
Nguyen TQ, Roestenberg P, van Nieuwenhoven
FA, et al: CTGF inhibits BMP-7 signaling in diabetic nephropathy. J
Am Soc Nephrol. 19:2098–2107. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
15.
|
Park Y, Kim JW, Kim DS, et al: The bone
morphogenesis protein-2 (BMP-2) is associated with progression to
metastatic disease in gastric cancer. Cancer Res Treat. 40:127–132.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
16.
|
Ara J, See J, Mamontov P, et al: Bone
morphogenetic proteins 4, 6, and 7 are up-regulated in mouse spinal
cord during experimental autoimmune encephalomyelitis. J Neurosci
Res. 86:125–135. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17.
|
Weiskirchen R, Meurer SK, Gressner OA,
Herrmann J, Borkham-Kamphorst E and Gressner AM: BMP-7 as
antagonist of organ fibrosis. Front Biosci. 14:4992–5012. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
18.
|
Rosen V: BMP2 signaling in bone
development and repair. Cytokine Growth Factor Rev. 20:475–480.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
19.
|
Langenfeld EM, Calvano SE, Abou-Nukta F,
Lowry SF, Amenta P and Langenfeld J: The mature bone morphogenetic
protein-2 is aberrantly expressed in non-small cell lung carcinomas
and stimulates tumor growth of A549 cells. Carcinogenesis.
24:1445–1454. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
20.
|
Kleeff J, Maruyama H, Ishiwata T, et al:
Bone morphogenetic protein 2 exerts diverse effects on cell growth
in vitro and is expressed in human pancreatic cancer in vivo.
Gastroenterology. 116:1202–1216. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
21.
|
Liu C, Tian G, Tu Y, Fu J, Lan C and Wu N:
Expression pattern and clinical prognostic relevance of bone
morphogenetic protein-2 in human gliomas. Jpn J Clin Oncol.
39:625–631. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22.
|
Baelde HJ, Eikmans M, Doran PP, Lappin DW,
de Heer E and Bruijn JA: Gene expression profiling in glomeruli
from human kidneys with diabetic nephropathy. Am J Kidney Dis.
43:636–650. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
23.
|
Dalfino G, Simone S, Porreca S, et al:
Bone morphogenetic protein-2 may represent the molecular link
between oxidative stress and vascular stiffness in chronic kidney
disease. Atherosclerosis. 211:418–423. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24.
|
Sieber C, Kopf J, Hiepen C and Knaus P:
Recent advances in BMP receptor signaling. Cytokine Growth Factor
Rev. 20:343–355. 2009. View Article : Google Scholar : PubMed/NCBI
|