Introduction
The extracellular matrix (ECM) is considered to play
an important role in the stability of tissues and in regulating the
growth and differentiation of cells (1,2).
Synthesis, accumulation and catabolism of the ECM occur during
wound healing and during the initiation and progression of numerous
diseases (3).
Moreover, it is generally acknowledged that the ECM
does not function as a mere passive scaffold for connective tissue
within organ architecture. It also plays an ‘informational’ role
through a network of interactions between cells and signal
molecules which is of primary importance in the control of cellular
proliferation and motility during histogenesis for the maintenance
of tissue homeostasis and in cancer development.
Prolapse of the pelvic organs is a common disease
affecting the lives of millions of women. Women with prolapse
suffer from chronic pelvic pain and pressure, urinary and fecal
incontinence, sexual dysfunction and social isolation. Despite the
high prevalence of this disease and the devastating impact on the
lives of women, very little is known about its pathophysiology.
Recently, it has been suggested that a structural
defect in the vagina and its supportive tissues, such as a decrease
in collagen content or a change in collagen subtypes, is one of the
mechanisms that predisposes a woman to prolapse. Thus, there have
been many studies throughout the literature in which collagen is
analyzed in vaginal and uterine-supporting ligament biopsies
procured at the time of a repair of prolapse in patients with
prolapse or at the time of a hysterectomy in patients without
prolapse. The comparison of these samples has provided conflicting
results (4–13).
Trauma or pathology may lead to altered responses to
mechanical stresses placed on connective tissue, producing changes
in the ECM. Alterations in collagen synthesis and collagen types
are causally related to connective tissue disorders, such as
inguinal hernia (14),
genito-urinary prolapse and urinary stress incontinence (15,16).
The uterine cervices of patients with prolapse are dramatically
altered when compared to those without prolapse. Therefore, the
precise control of ECM metabolism in the uterine cervix is critical
for the pathophysiology of prolapse uteri.
In the present study, we investigated the
composition of the various types of collagen, the major component
of ECM, in human uterine cervical tissues of prolapse uteri by
sodium dodecyl sulfate-polyacrylamide gel electrophoresis
(SDS-PAGE).
Materials and methods
This project was approved by the Committee on
Investigations Involving Human Subjects of Wakayama Medical
College. Informed consent was obtained from each subject after the
purpose and nature of the study had been fully explained.
Patients and tissues
Sixteen uterine cervical samples were obtained from
postmenopausal Japanese women (59–85 years of age) with uterine
prolapse, who had undergone a vaginal hysterectomy and
colpoperineoplasty, and were defined as the prolapse group. A
Pelvic Organ Prolapse Quantification examination (17) was performed on the patients with
uterine prolapse to define the stage of prolapse. All patients with
uterine prolapse showed stage II or greater prolapse (POP-Q).
As the control group, 20 uterine cervical samples
were obtained from postmenopausal Japanese women (58–82 years of
age) without uterine prolapse, who had undergone a abdominal
hysterectomy for a benign indication other than prolapse.
Variables, including age, postmenopausal period, parity and BMI,
were not significantly different in the two groups. None of the
patients received any hormone therapy before surgery.
The uterine cervical length was measured from the
internal to external os along the cervical canal, and values are
expressed in millimeters.
SDS-PAGE of pepsin-solubilized collagens
from the uterine cervical tissues
Minced samples of human uterine cervical tissues
were washed overnight in cold distilled water and freed of blood.
Tissues were homogenized with a Polytron homogenizer in 50 volumes
of 0.5 M acetic acid that contained 1 mg/ml pepsin (Sigma Chemical
Co., St. Louis, MO, USA). Collagens were extracted with constant
stirring for 24 h at 4°C. Each solution was centrifuged at 39,000 ×
g for 1 h at 4°C. Collagens were reextracted from the pellet under
the same conditions as described above for 48 h.
The supernatants corresponding to individual samples
were then combined, and collagens were precipitated by addition of
4.0 M NaCl to a final concentration of 2.0 M. Each precipitate was
dissolved in 0.5 M acetic acid, and the solution was dialyzed
against 0.02 M Na2HPO4. Precipitated
collagens were redissolved in 0.5 M acetic acid, dialyzed
exhaustively against 0.05 M acetic acid and finally
lyophilized.
The solubility of the tissue collagen from each
uterine cervical tissue sample was estimated by comparing the
hydroxyproline content of the initial homogenate to that of the
final solution of collagen (18).
Type V collagen was isolated by salt precipitation from pepsin
digests of human uterine cervical tissues by the methods described
elsewhere (19,20). The extracted type V collagen was
also lyophilized.
Estimation of the relative abundance of the α1(III)
chain and α1(V) chain was performed by interrupted gel
electrophoresis (21).
Electrophoresis was performed in an 8% polyacrylamide slab gel
(Sigma Chemical Co.). The gel and electrode buffers consisted of
0.1 M phosphate buffer, pH 7.2, containing 0.1% SDS (Nacalai
Tesque, Inc., Kyoto, Japan), as previously described (22). Lyophilized samples of collagen and
type V collagen were dissolved at a concentration of 0.2 mg/ml and
denatured by heating in the gel buffer that contained 1% SDS at
60°C for 30 min.
Aliquots of 25 μl of each solution of denatured
collagens and 5 μl of denatured type V collagen were applied to the
gel and subjected to electrophoresis at 80 mA. After 1.5 h, the
current was switched off and sample wells were filled with a
solution of 20% β-mercaptoethanol (Wako Chemical Co., Osaka, Japan)
in gel buffer, and the β-mercaptoethanol was allowed to diffuse
into the gel for 1 h to cleave the intramolecular disulfide bonds
of type III collagen, [α1(III)]3. Then, electrophoresis was resumed
and allowed to continue for another 1 h. Each collagen α chain was
stained with Coomassie brilliant blue (Sigma Chemical Co.) and
quantitated by densitometry.
The relative amounts of α1(III) or α1(V) chains were
calculated by dividing the intensities of the band areas under
densitometric peaks of α1(III) and of α1(V) by that of α1(I). In
this method, the α chains derived from type IV collagen could not
be evaluated because of degradation of this collagen by pepsin.
Statistical analysis
The uterine cervical length was expressed as the
mean ± SD. Densitometric data were expressed as the mean ± SEM.
Mean values were compared by the Student’s t-test or analysis of
variance using a StatView software program on a Macintosh computer.
Two-tailed P-values <0.05 were considered statistically
significant.
Results
The uterine cervix of prolapse uteri were
significantly longer compared to those of the postmenopausal uterus
(Table I).
 | Table I.Relative abundance of the α1(III) and
α1(V) chains of collagen as compared to the α1(I) chain in the
human uterine cervix tissues of the postmenopausal uterus as
control and the prolapse uteri. |
Table I.
Relative abundance of the α1(III) and
α1(V) chains of collagen as compared to the α1(I) chain in the
human uterine cervix tissues of the postmenopausal uterus as
control and the prolapse uteri.
| No. of patients | Cervical length
(mm) | Relative ratio of
collagen α chain
|
|---|
| α1(III)/α1(I) | α1(V)/α1(I) |
|---|
| Control | 16 | 28±5 | 0.18±0.05 | 0.02±0.01 |
| Prolapse uteri | 20 | 37±8a | 0.06±0.03b | 0.01±0.01 |
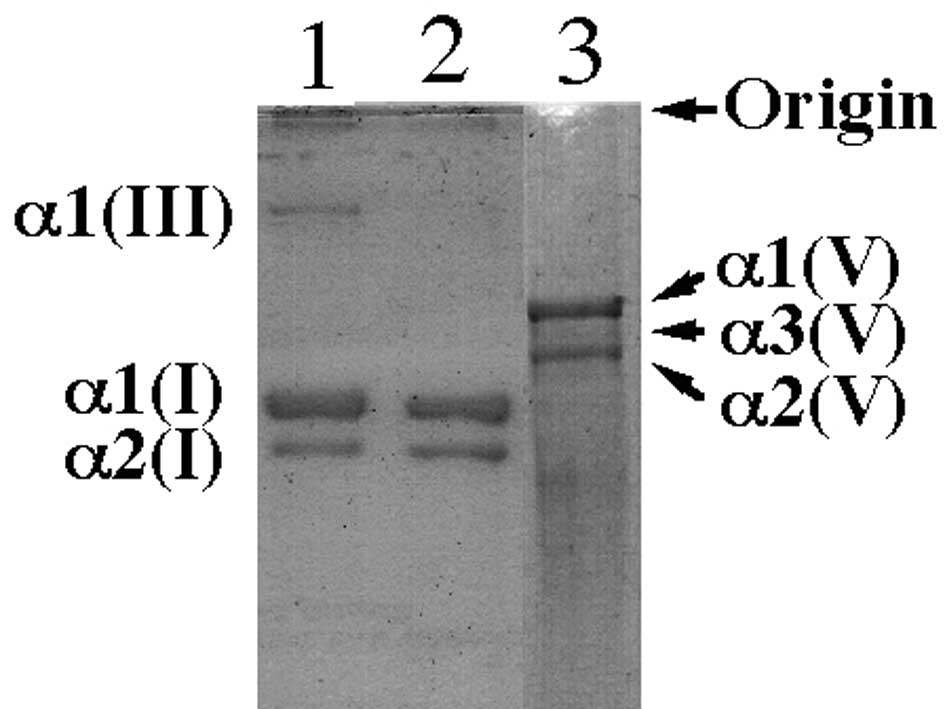
Although the relative levels of α1(I) and α1(V) were
similar in the uterine cervical tissues in the two groups (Fig. 1), those of α1(III) were decreased
in the uterine cervical tissues of the prolapse uteri compared to
those of the normal postmenopausal uterine cervix (Fig. 1). The mean ratio of the intensity
of the band of α1(III) to that of α1(I) in the uterine cervix
tissues of prolapse uteri was significantly lower than that in the
normal postmenopausal uterine tissues (P<0.01). The data are
summarized in Table I.
Discussion
In the present study, we investigated changes in the
composition of the ECM, including type I, III and V collagens, in
the human uterine cervical tissues obtained from the women with or
without uterine prolapse. We were able to solubilize 70–85% of
collagen in the human cervical tissues, as measured by reference to
levels of hydroxyproline (data not shown). Therefore, it was
postulated that the extracted collagen may accurately reflect the
entire complement of collagen in the tissues.
Human cervix is made up mainly of fibrous connective
tissues in which collagen (23)
and glycosaminoglycans (24)
predominate. The physiological properties of the cervix depend on
the interplay between collagen and glycosaminoglycan molecules
(25). Therefore, collagen may
play a pivotal role in the structure and function of human uterine
cervix.
Although type I and type III collagens are commonly
found in combination, the ratios of type III to type I collagen in
the tissues of human cervix of the patients with uterine prolapse
were significantly lower than those of the patients without
prolapse. Our findings suggest at least two possible mechanisms for
decreased type III collagen expression in the uterine cervices with
uterine prolapse. First, the reduced type III collagen expression
in the pelvic organ supporting system involving uterine cervix may
induce genital prolapse. However, these present findings conflict
with those of previous reports (4–13).
Second, the pelvic supporting system is relaxed in
patients with uterine prolapse. Therefore, a progressively
increasing mechanical load may reduce focal type III collagen
expression in uterine cervical tissues.
It has been suggested that the ECM in human cervical
tissues may be controlled by collagenases, hormones, such as
estrogens (26–28), dehydroepiandrosterone (29) and prostaglandins (30,31),
and/or cytokines, such as IL-I (32) and IL-8 (33–37).
Recent studies have shown that nitric oxide (NO) may
participate in the biochemical and anatomical changes in human and
animal cervical tissues (38–42).
In addition, it has been suggested that NO directly controls
various metalloproteinases (43).
A low estrogen level is commonly recognized in postmenopausal women
with or without uterine prolapse. Therefore, in regards to the
causes of uterine prolapse, we may suggest not only lack of
estrogen, but also other factors, such as NO.
The ECM of the uterine cervix is a fiber-reinforced
compositive viscoelastic material made up of fibrillar collagen
(two thirds type I, one third type III) and proteoglycans
(hyaluronic acid, chondroitin sulfate, keratan sulfate and dermatan
sulfate) (44,45). Thus, our study suggests that a
decreased ratio of type III collagen and an increased ratio of type
I collagen in the uterine cervix may induce elongation of the
cervix in terms of uterine prolapse. Type III collagen belongs to a
family of fibril-forming collagens that are found throughout the
body. Each of the fibrillar collagens has a unique structural
profile that in turn imparts tissues with distinct physical
properties (46,47). This collagen, forming smaller
fibers, predominates in tissues that require increased flexibility
and distensibility and are subject to periodic stress, such as the
vasculature. Therefore, decreased expression of type III collagen,
the main macromolecular component of the ECM of the cervix, may be
a key event in the development of uterine prolapse.
In conclusion, these changes in the composition of
collagens may result in the morphologic and functional
characteristics of human uterine cervix with uterine prolapse.
This study furthers the understanding of the
pathophysiology of human uterine cervix in terms of ECM metabolism.
Further research is required to elucidate the mechanisms regulating
the expression of genes of other types of collagen or
metalloproteinases in the uterine cervix during the premenopausal
period and pathological status.
References
|
1.
|
Lin CQ and Bissell MJ: Multi-faceted
regulation of cell differentiation by extracellular matrix. FASEB
J. 7:737–743. 1993.PubMed/NCBI
|
|
2.
|
Madri JA and Basson MD: Extracellular
matrix-cell interactions: dynamic modulators of cell, tissue and
organism structure and function. Lab Invest. 66:519–521.
1992.PubMed/NCBI
|
|
3.
|
Haralson MA: Extracellular matrix and
growth factors: an integrated interplay controlling tissue repair
and progression to disease. Lab Invest. 69:369–372. 1993.PubMed/NCBI
|
|
4.
|
Goepel C, Heller L, Methfessel HD and
Koelbl H: Periurethral connective tissue stastus of postmenopausal
women with genital prolapse with and without stress incontinence.
Acta Obstet Gynecol Scand. 82:659–664. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
5.
|
Soderberg MW, Falconer C, Bystrom B,
Malmstrom A and Ekman G: Young women with genital prolapse have a
low collagen concentration. Acta Obstet Gynecol Scand.
83:1193–1198. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
6.
|
Jackson SR, Avery NC, Tarlton JF, Eckford
SD, Abrams P and Bailey AJ: Changes in the metabolism of collagen
in genitourinary prolapse. Lancet. 347:1658–1661. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
7.
|
Liapis A, Bakas P, Pafiti A,
Frangos-Plemenos M, Arnoyannaki N and Creatsas G: Changes of
collagen type III in female patients with genuine stress
incontinence and pelvic floor prolapse. Eur J Obstet Gynecol Reprod
Biol. 97:76–79. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
8.
|
Makinen J, Kahari VM, Soderstrom KO,
Vuorio E and Hirvonen T: Collagen synthesis in the vaginal
connective tissue of patients with and without uterine prolapse.
Eur J Obstet Gynecol Reprod Biol. 24:319–325. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
9.
|
Kokcu A, Yanik F, Cetinkaya M, Alper T,
Kandemir B and Malatyalioglu E: Histopathological evaluation of the
connective tissue of the vaginal fascia and the uterine ligaments
in women with and without pelvic relaxation. Arch Gynecol Obstet.
226:75–78. 2002. View Article : Google Scholar
|
|
10.
|
Chen BH, Wen Y, Li H and Polan ML:
Collagen metabolism and turnover in women with stress urinary
incontinence and pelvic prolapse. Int Urogynecol J Pelvic Floor
Dysfunct. 13:80–87. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
11.
|
Takano CC, Girao MJ, Sartori MG, et al:
Analysis of collagen in parametrium and vaginal apex of women with
and without uterine prolapase. Int Urogynecol J Floor Dysfunct.
13:342–345. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
12.
|
Ewies A, Al-Azzawi F and Thompson J:
Changes in extracellular matrix proteins in the cardinal ligaments
of post-menopausal women with or without prolapse: a computerized
immunohistomorphometric analysis. Hum Reprod. 18:2189–2195. 2003.
View Article : Google Scholar
|
|
13.
|
Moalli PA, Shand SH, Zyczynski HM, Gordy
SC and Meyn LA: Remodeling of vaginal connective tissue in patients
with prolapse. Obstet Gynecol. 106:953–963. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
14.
|
Friedman D, Boyd C, Norton P and Deak S:
Altered type III collagen synthesis in patients with inguinal
hernia. Am Surg. 218:754–760. 1993.PubMed/NCBI
|
|
15.
|
Ulmsten U, Ekman G, Giertz G and Malmstrom
A: Different biochemical composition of connective tissue in
continent and stress incontinent women. Acta Obstet Gynecol Scand.
66:455–457. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
16.
|
Versi E, Cardozo L, Bribcat M, Cooper D,
Montgomery J and Studd J: Corelation of urethral physiology and
skin collagen in postmenopausal women. Br J Obstet Gynecol.
95:147–152. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
17.
|
Bump RC, Mattiasson A, Bo K, et al: The
standardization of terminology of female pelvic organ prolapse and
pelvic floor dysfunction. Am J Obstet Gynecol. 175:10–17. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
18.
|
Kivirikko KI and Prockop DJ: Hydroxylation
of proline in synthetic polypeptides with purified protocollagen
hydroxylase. J Biol Chem. 242:4009–4012. 1967.PubMed/NCBI
|
|
19.
|
Furuto DK and Miller EJ: Isolation of a
unique collagenous fraction from limited pepsin digests of human
placental tissue. J Biol Chem. 255:290–295. 1980.PubMed/NCBI
|
|
20.
|
Miller EJ and Rhodes RK: Preparation and
characterization of the different types of collagen. Methods
Enzymol. 82:33–64. 1982. View Article : Google Scholar
|
|
21.
|
Sykes B, Puddle B, Francis M and Smith R:
The estimation of two collagen from human dermis by interrupted gel
electrophoresis. Biochem Biophys Res Commun. 72:1472–1480. 1976.
View Article : Google Scholar : PubMed/NCBI
|
|
22.
|
Laemmli UK: Cleavage of structural
proteins during the assembly of the head of bacteriophage T4.
Nature. 227:680–685. 1970. View
Article : Google Scholar : PubMed/NCBI
|
|
23.
|
Danforth DN: The fibrous nature of the
human cervix, and its relation to isthmic segment in gravid and non
gravid uteri. Am J Obstet Gynecol. 53:541–560. 1974.PubMed/NCBI
|
|
24.
|
Kitamura K, Ito A, Mori Y and Hirakawa S:
Glycosaminoglycans of human uterine cervix: heparan sulfate
increase with reference to cervical ripening. Biochem Med.
23:159–167. 1980. View Article : Google Scholar : PubMed/NCBI
|
|
25.
|
Wallis RM and Hiller K: Regulation of
collagen dissolution in human cervix by oestradiol-17b and
progesterone. J Reprod Fertil. 62:55–61. 1981. View Article : Google Scholar : PubMed/NCBI
|
|
26.
|
Sato T, Ito A, Mori Y, Yamashita K,
Hayakawa T and Nagase H: Hormonal regulation of collagenolysis in
uterine cervical fibroblasts. Modulation of synthesis of
procollagenase, prostromelysin and tissue inhibitor
metalloproteinases (TIMP) by progesterone and oestradiol 17 b
Biochem J. 275:645–650. 1991.PubMed/NCBI
|
|
27.
|
Rajabi MR, Dodge GR, Solomon S and Poole
AR: Immunochemical and immunohistochemical evidence of
estrogen-mediated collagenolysis as a mechanism of cervical
dilatation in the guinea pig at partrition. Endocrinology.
128:371–378. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
28.
|
Mochizuki M, Honda T, Deguchi M, Morikawa
H and Tojo S: A study of the effect of dehydroepiandrosterone
sulfate on so-called cervical ripening. Acta Obstet Gynecol Scand.
57:397–401. 1978. View Article : Google Scholar : PubMed/NCBI
|
|
29.
|
Uldbjerg N, Forman A, Petersen L, et al:
Biochemical changes of the uterus and cervix during pregnancy.
Medicine of the Fetus and Mother. Reece EA, Hobbins JC, Mahoney MJ
and Petrie RH: JB Lippincott Co; Philadelphia: pp. 849–868.
1992
|
|
30.
|
Kelly RW: Pregnancy maintenance and
partrition: the role of prostaglandin in manipulating the immune
and inflammatory response. Endocr Rev. 15:684–706. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
31.
|
El Maradney E, Kanayama N, Halim A,
Maehara K, Sumimoto K and Terao T: The effect of interleukin-1 in
rabbit cervical ripening. Eur J Obstet Gynecol Reprod Biol.
60:75–80. 1995.PubMed/NCBI
|
|
32.
|
Barclay CG, Brennad JE, Kelly RW and
Calder AA: Interleukin-8 production by human cervix. Am J Obstet
Gynecol. 169:625–632. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
33.
|
El Maradney E, Kanayama N, Kobayashi H, et
al: The role of hyaluronic acid as a mediator and regulator of
cervical ripening. Hum Reprod. 12:1080–1088. 1997.PubMed/NCBI
|
|
34.
|
Osmers R, Blaser J, Kuhn W and Tshesche H:
Interleukin-8 synthesis and the onset of labor. Obstet Gynecol.
86:223–229. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
35.
|
Chwalisz K, Benson M, Scholz P, Daum J,
Beier HM and Hegele-Hartung C: Cervical ripening with cytokine
interleukin-8, interleukin-1 beta and tumour necrosis factor alpha
in guinea-pigs. Hum Reprod. 9:2173–2181. 1994.PubMed/NCBI
|
|
36.
|
El Maradney E, Kanayama N, Halim A,
Maehara K, Sumimoto K and Terao T: Interleukin-8 induces cervical
ripening in rabbits. Am J Obstet Gynecol. 171:77–83.
1994.PubMed/NCBI
|
|
37.
|
Garfield RE, Saade G, Buhimschi C, et al:
Control and assessment of the uterus and cervix during pregnancy
and labour. Hum Reprod Update. 4:673–695. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
38.
|
Tschugguel W, Schneeberger C, Lass H, et
al: Human cervical ripening is associated with an increase in
cervical inducible nitric oxide synthase expression. Biol Reprod.
60:1367–1372. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
39.
|
Buhimschi I, Ali M, Jain V, Chwalisz K and
Garfield RE: Differential regulation of nitric oxide in the rat
uterus and cervix during pregnancy and labour. Hum Reprod.
11:1755–1766. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
40.
|
Chwalisz K, Shao-Qing S, Garfield RE and
Beier HM: Cervical ripening in guinea pigs after a local
application of nitric oxide. Hum Reprod. 12:2093–2101. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
41.
|
Ali M, Buhimschi I, Chwalisz K and
Garfield RE: Changes in expression of the nitric oxide synthase
isoforms in rat uterus and cervix during pregnancy and parturition.
Mol Hum Reprod. 3:995–1003. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
42.
|
Thomson AJ, Lunan CB, Cameron AD, Cameron
IT, Greer IA and Norman JE: Nitric oxide donors induce ripening of
the human uterine cervix: a randomised controlled trial. Br J
Obstet Gynaecol. 104:1054–1057. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
43.
|
Drapier JC and Bouton C: Modulation by
nitric oxide of metalloproteinase regulatory activities. Bioessays.
18:549–556. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
44.
|
Aspeden RM: Collagen organization in the
cervix and its relation to mechanical functions. Collagen Relat
Res. 8:103–112. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
45.
|
Kleissel HP, van der Rest M, Naftolin F,
Glorieux FH and de Leon A: Collagen changes in the human uterine
cervix at partruition. Am J Obstet Gynecol. 130:748–753.
1978.PubMed/NCBI
|
|
46.
|
Ottani V, Martini D, Franchi M, Ruggeri A
and Raspanti M: Hierarchical structures in fibrillar collagen.
Micron. 33:587–596. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
47.
|
Oxlund H: Relationships between the
biomechanical properties, composition and molecular structure of
connective tissues. Connect Tissue Res. 15:65–72. 1986. View Article : Google Scholar : PubMed/NCBI
|