Introduction
Antitumor immunity mainly involves T cell-mediated
cytoimmunity. T cell activation requires two signals. The first
signal is specific. The second signal is non-specific and is
provided by co-stimulatory molecules on the antigen presenting cell
(APC) and corresponding ligands on the T cell. The absence of the
second signal may lead to T cell anergy (1). The dendritic cells (DCs), as highly
efficient and specialized APCs, are particularly crucial for the T
cell-mediated immune response. The 4-1BB/4-1BBL receptor/ligand
pair in the immune system is another significant co-stimulatory
signal other than CD28/ B7, which plays an important role in immune
response and tumor immunity, and also plays a part in cell
adhesion, antigen presentation, T cell co-stimulation and signal
transduction (2). In our study, we
prepared a DC vaccine transfected with co-stimulatory molecule
4-1BBL transgenic murine gastric cancer cell total ribonucleic acid
(RNA), in which the double signals may enhance the antitumor
activity.
Materials and methods
Materials and reagents
A total of 615 mice (6–8 weeks old) were purchased
from the laboratory animal center of the Chinese Academy of Medical
Science (Tianjin, China). Mice were maintained in a
specific-pathogen-free environment. All animal studies were
approved by the ethics committee of the authors’ institutional
ethics committee. The murine forestomach carcinoma (MFC) cell
strain was purchased from the Chinese Academy of Medical Science
(Shanghai, China). The pMKITneo/4-1BBL plasmid was kindly provided
by Professor Hideo Yagita. Lipofectamine 2000 was purchased from
Invitrogen Corp. (Carlsbad, CA, USA). IL-12 and IFN-γ ELISA kits
were purchased from NeoBioscience Technology Company (Beijing,
China).
MFC culture and 4-1BBL gene
transfection
The DH5α bacterial strain containing the
pMKITneo/4-1BBL plasmid was incubated in Lysogeny broth (LB). The
plasmid DNA was extracted using a Wizard plus SV minipreps DNA
purification system (Promega, Shanghai, China) and was digested by
the restriction endonucleases XhoI and NotI, and then
the identity of the plasmid was verified by electrophoresis on a
0.7% agarose gel.
MFC cell lines were cultured in RPMI-1640
supplemented with 10% fetal calf serum (FCS) in a humidified
atmosphere containing 5% CO2 at 37°C.
The MFC cell line was transfected with the plasmid
pMKITneo/4-1BBL using Lipofectamine 2000, and cultured in medium
containing 0.4 g/l G418 to screen 4-1BBL-expressing stable cell
lines, MFC/4-1BBL.
RNA preparation and semi-quantitative
reverse transcription polymerase chain reaction (RT-PCR)
Total mRNAs of the MFC and MFC/4-1BBL cell lines
were purified using TRIzol (Invitrogen) according to the
manufacturer's instructions. The clear 28S and 18S ribosomal RNA
bands indicated a good quality of RNA. The total mRNA was
subsequently amplified by RT-PCR. The PCR protocol included an
initial denaturation step at 95°C for 5 min, followed by 35 cycles
with denaturation at 94°C for 45 sec, annealing at 56°C for 30 sec
and extension at 72°C for 45 sec. The primer used were as follows:
forward, 5′-TCACTCGAGATGGACCAGCACACACTTGATG-3′; and reverse,
5′-GGCTGTTGGGTACCCTTACTCGCCGGCG AAT-3′. The length of the amplified
product was 616 bp. β-actin was used as the internal control. The
identity of the PCR products was verified by electrophoresis on a
1.5% agarose gel.
Generation of myeloid DCs
Myeloid DCs were isolated from the bone marrow of
615 mice as previously described (21). Briefly, the 615 mice were
sacrificed and immersed in 75% ethanol for 2 min. The femur and
tibia were removed by aseptic surgery. The bones were then crushed
and washed repeatedly with RPMI-1640 to remove bone marrow. The
RPMI-1640 was centrifuged at 1,000 rpm for 5 min. Erythrocyte lysis
buffer was added into the supernatant to remove red blood cells.
The isolated myeloid DC cells were cultured in 6-well plates and
maintained in the RPMI-1640 supplemented with 10% FCS, 10 ng/ml
GM-CSF and 10 ng/ml IL-4. The morphology of the cultured DCs,
including shape and cell size, was carefully observed and verified
by an inverted microscope on days 1, 3, 5 and 7 of isolation.
RNA transfection of DCs
The mouse DCs were cultured for 5 days. Then, 2 h
prior to transfection, DCs were washed with RPMI-1640 serum-free
medium 3 times and the cell density was adjusted to
1×106 cells/ml using RPMI-1640. For each transfection
sample, the complex was prepared as follows: 10 μg MFC/4-1BBL RNA
were diluted in 500 μl serum-free RPMI-1640, and 10 μl
Lipofectamine 2000 were diluted in 500 μl serum-free RPMI-1640. The
complex was incubated for 5 min at room temperature. The diluted
RNA was combined with diluted Lipofectamine 2000, mixed gently and
incubated at 37°C in a 5% CO2 incubator for 30 min. The
complex was added to the DC plates at 37°C in a 5% CO2
incubator. The medium was changed after 4 h, and the incubation of
the DCs was continued.
ELISA assays were performed to measure IL-12 levels
in the supernatant of immature DCs, DCs and MFC/4-1BBL/DCs.
The mRNAs of MFC/4-1BBL/DCs and DCs were purified
using TRIzol, followed by amplification using RT-PCR. All products
were analyzed by electrophoresis on 1.5% agarose gels.
Isolation and culture of mouse
lymphocytes
The 615 mice were sacrificed. The spleen was removed
on a 200-mesh sterilized copper net, and ground repeatedly. After
washing with phosphate-buffered saline (PBS), single spleen cell
suspension was collected and centrifuged. The pellet was mixed with
PBS and mouse hydroxypropylmethyl cellulose at a 1:1 ratio,
followed by centrifugation at 1,500 rpm for 15 min. After washing 3
times with PBS, the isolated lymphocytes were cultured in the
RPMI-1640 supplemented with 10% FCS.
MTT lymphocyte proliferation
Mixed lymphocyte reactions were performed using
mature DCs or MFC/4-1BBL/DCs as stimulator cells, and T lymphocytes
as responder cells. Stimulator cells were incubated with 25 μg/ml
mitomycin C (MMC) at 37°C for 30 min and cell density was adjusted
to 4×104/ml, 2×104/ml or 1×104/ml.
Three repeats of each cell density were plated in 96-well
plates.
The allogeneic lymphocytes at a density of
2×105/ml were used as responder cells. The lymphocytes
were added into the stimulator cells. The responder cells or
stimulator cells alone were used as the control. A total of 4 h
prior to detection, MTT was added into the cells and incubated for
4 h in the dark, followed by addition of DMSO. The optical density
(OD) at 570 nm was measured using the ELISA machine.
T cell proliferation rate = [experimental OD -
(responder cell OD + stimulator cell OD)]/responder cell OD x
100%.
MTT tumor killing activity of
DC-activated cytotoxic T lymphocytes (CTLs)
A tumor killing activity assay was performed using T
cells together with mature DCs or MFC/4-1BBL/ DCs as effector
cells, and MFC cells as target cells. DCs or MFC/4-1BBL/DCs
(2×105/ml) were cultured in 12-well plates together with
allogeneic lymphocytes (2×105/ml) in the RPMI-1640
containing 10 ng/ml IL-2 on day 1, 3 and 5 of culture. The
stimulator to responder cells at the ratio of 1:20, 1:10 or 1:5
were mixed and added in 96-well plates. Three repeats of each cell
density were performed. The stimulator cells or responder cells
alone were used as the control. A total of 4 h prior to detection,
MTT was added to the cells and incubated for 4 h in the dark,
followed by addition of DMSO. The OD at 570 nm was measured by the
ELISA machine.
Kill rate = [experimental OD – (effector cells OD +
target cells OD)]/effector cells OD x 100%.
The IFN-γ levels in the supernatant of the DCs or
MFC/4-1BBL/DCs were measured using the ELISA kit.
Statistical analysis
All statistical calculations were carried out using
SPSS 13.0. T-tests and ANOVA were used to compare the different
groups. All values are indicated as the means ± standard errors.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Verification of the pMKITneo/4-1BBL
plasmid
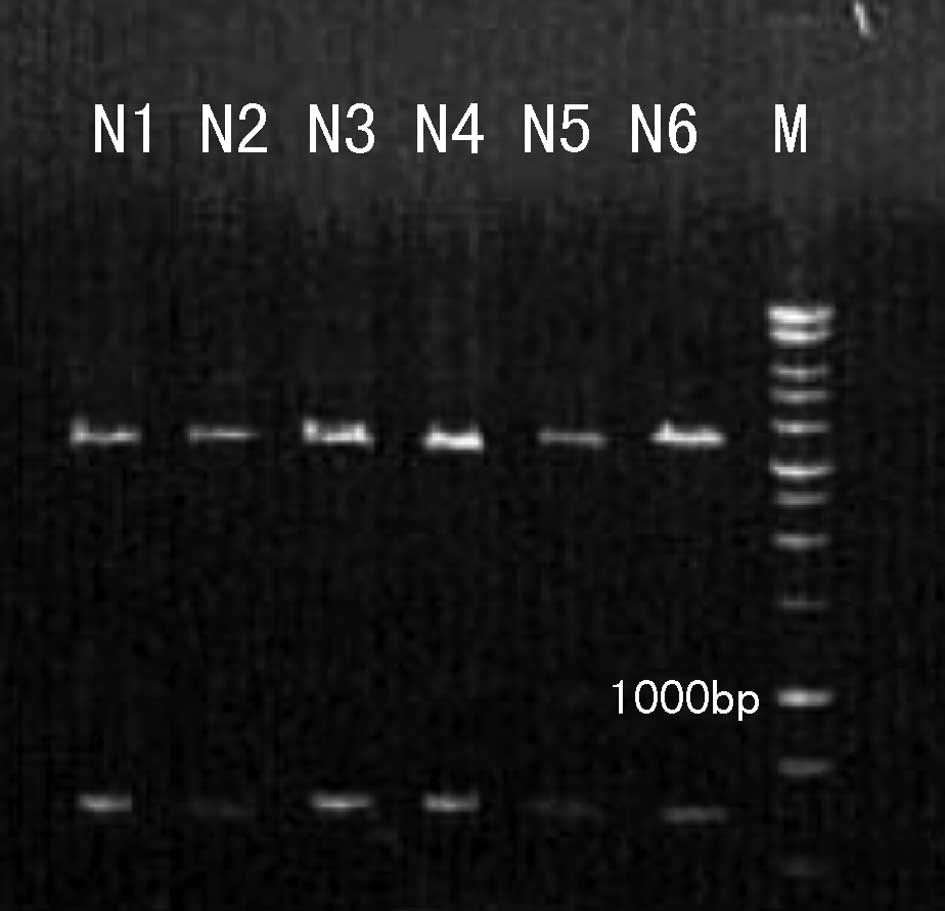
The purified pMKITneo/4-1BBL plasmid was digested by
XhoI and NotI. A 927 bp DNA fragment of the inserted
4-1BBL coding sequence was released from the 5.8kb vector plasmid
(Fig. 1). Our results indicate
that the pMKITneo/4-1BBL plasmid was successfully purified.
Observation of DC morphology
The morphology of DCs was measured using an inverted
microscope. On day 1, the majority of DCs were round or oval-shaped
with a small size, and were attached well to the plate. On day 3,
the cell number was significantly increased with an enlarged cell
size, and some cells had grown as clustered colonies. On day 5, the
number of cells was further increased and the cells were a similar
size to that on day 3. Some cells showed burr-like protuberance,
and the cell colonies were markedly increased. On day 7, the
majority of the cells were detached from the plate with the nucleus
deviated. A large number of burr-like protrusions were observed in
characteristic irregular cells. Some cells contained round, star
and dendritic-like protrusions. Notably, 12 h post-transfection,
the morphology of cells returned to the state prior to
transfection. Cells transfected with MFC total RNA carrying the
4-1BBL gene were round, and showed slightly reduced size, fewer
surface protrusions and increased cell particles (Fig. 2).
MTT lymphocyte proliferation pre- and
post-transfection
The ratio of stimulator cells to responder cells was
1:20, 1:10 or 1:5. The effects of the MFC/4-1BBL/DC group on T cell
proliferation (0.093±0.012, 0.187±0.013, 0.187±0.013) were
significantly higher than that of the DC group (0.060±0.014,
0.148±0.017, 0.195±0.015) (P<0.05). We also noted that the
higher stimulator cell to responder cell ratio appeared to show
stronger T cell proliferation (Table
I).
 | Table I.The comparison of T cell
proliferation in the different DC groups (n=3, x̄±s). |
Table I.
The comparison of T cell
proliferation in the different DC groups (n=3, x̄±s).
| Stimulation to
responding ratio (%)
|
|---|
| 1:20 | 1:10 | 1:5 |
|---|
| Group | | | |
| DC | 0.060±0.014 | 0.148±0.017 | 0.195±0.015 |
|
MFC/4-1BBL/DC | 0.093±0.012 | 0.187±0.013 | 0.240±0.021 |
| P-value | 0.040 | 0.035 | 0.039 |
MTT tumor killing activity of
DC-activated CTLs pre- and post-transfection
The ratio of target to effector cells was 1:20, 1:10
or 1:5. The effects of the MFC/4-1BBL/DC group tumor cell kill rate
(0.533±0.092, 0.321±0.018, 0.218±0.025) were higher than that of
the DC group (0.342±0.023, 0.267±0.0243, 0.147±0.031) (P<0.05).
Consistent with the proliferation results, higher target cell to
effector cell ratio showed a stronger tumor cell killing rate
(Table II).
| Table II.The activation of cytotoxic T
lymphocyte (CTL) killing rate of tumor in the different DC groups
(n=3, x̄±s). |
Table II.
The activation of cytotoxic T
lymphocyte (CTL) killing rate of tumor in the different DC groups
(n=3, x̄±s).
| Target/effector
cell ratio (%)
|
|---|
| 1:20 | 1:10 | 1:5 |
|---|
| Group | | | |
| DC+T cell | 0.342±0.023 | 0.267±0.0243 | 0.147±0.031 |
| MFC/4-1BBL/DC+T
cell | 0.533±0.092 | 0.321±0.018 | 0.218±0.025 |
| P value | 0.025 | 0.036 | 0.037 |
The secretion of IL-12 and IFN-γ
We measured IL-12 and IFN-γ secretion in the culture
medium of different groups using ELISA analysis. We found that the
IL-12 levels for the DC/ MFC/4-1BBL group were 20.240±2.494 pg/ml,
17.088±3.933 pg/ ml for the DC group, and 10.288±2.390 pg/ ml for
the immature DC group (P<0.05). The IFN-γ levels were
9.451±2.925 pg/ml for the DC/ MFC/4-1BBL group and 5.979±1.639
pg/ml for the DC group (P<0.05).
Discussion
Gastric cancer is one of the most common malignant
tumors. Tumor immune escape and tolerance are major problems in the
treatment of the disease. Gene therapy provides a novel clinic
strategy to enhance the immune function of tumor immunity. Upon
activation by sensitized DCs, auxiliary T lymphocytes and CTLs are
capable of killing tumors, which is the theoretical basis of tumor
vaccines (2–4). There are a number of reports on DC
vaccines in the treatment of melanoma, ovarian, colon (5), breast (6) and gastric cancer (7). Although some of them have been used
in clinical antitumor treatment and have shown certain curative
effects, we should have a better understanding of the patients,
including human leukocyte antigen (HLA) phenotype and antigenic
peptide epitopes. Recently, investigators have focused on DC
vaccines loaded with total RNA with full tumor messages, and the
advantages of this method are: no advance knowledge of the
patient’s HLA phenotype is required; total RNA contains multiple
antigens and multiple epitopes of certain proteins that may be
presented by different HLA types, therefore including a variety of
tumor antigen encoded information; genetic material is easily
available but not integrated into the host chromosome, so more
secure; little damage to the DC is conductive to maintaining the
activity and presenting function of the cell; RNA may be isolated
from a small amount of tumor tissue and may be amplified to provide
an adequate amount of material; comprehensive immune responses are
induced to prevent antigenic variation and immune escape due to
antigenic variation (8–10).
The 4-1BB/4-1BBL receptor/ligand pair is another
crucial co-stimulatory signal besides CD28/B7 in immune response
and tumor immunity. It activates resting T cells separately or in
coordination with CD28. It plays a significant role to maintain the
existence and memory response of T lymphocytes (11). The secondary response of
CD8+ T cells is highly dependent on 4-1BBL, which is
also the case for some, though not all, secondary CD4+ T
cell responses. Localized expression of immunostimulatory ligands
using gene transfer approaches may reduce the risk of excessive,
generalized immune activation observed in some clinical trials of
systemically delivered antibodies (12,13).
Our investigation on DCs transfected with total RNA from murine
gastric carcinoma cells with 4-1BBL gene in vitro, reduces
the possibility of immune escape. It is a safe and easy method and
has a potential clinical application (9,12).
Studies using knockout mice have found that the
secondary response of CD8+ T cells was highly dependent
upon 4-1BBL. Though not all CD4+ T cells in the
secondary response depended on the 4-1BBL, lasting tumor-specific
CD4+ T cells may effectively lead to the response of CTL
effector cells (14). 4-1BBL is an
important co-stimulatory molecule in the T cell immune response,
which is capable of promoting the amplification of tumor-specific T
cells (14,15). The mature DCs transfected with
4-1BBL are capable of secreting large amounts of IL-12. It is known
that IL-12 is one of the most important factors to promote helper T
cell conversion to Th1 subsets and it is also a CTL differentiation
factor (16,17). The production of IL-12 is a
critical step in the CTL activation process. It promotes CTL
proliferation and generation of the protein perforin, induces the
secretion of IFN-γ and mediates the Th1 response. The 4-1BBL ligand
also co-stimulates T cells and results in the secretion of IFN-γ
with CD28 (11), which is capable
of promoting T cell differentiation to the Th1 type cells and
preventing cytokine secretion in the Th2 cell (16).
Patients with malignant tumors can produce DCs with
low expression of MHCI molecules, low secretion of IL-12 and
reduced ability to uptake antigen and to stimulate T cells. Our
results suggest that the effect on T cell proliferation in
MFC/4-1BBL/DC groups was stronger than that in the DC group, and
with the increased ratio of stimulator cells to responder cells,
the T cell proliferation was enhanced, and the levels of IL-12 in
the MFC/4-1BBL/DC group were significantly higher. Moreover, the
tumor killing ability by CTLs activated with transfected DCs
(MFC/4-1BBL/DC group) was stronger, and the levels of IFN-γ of in
the MFC/4-1BBL/ DC groups were significantly higher than in other
groups. 4-1BBL-transfected DCs may provide dual signals; therefore,
their ability to promote T cell proliferation is significantly
enhanced compared with non-transfected DCs, and that is induced to
secrete IFN-γ, which directly activates CD8+ T cells and
further enhances the specific tumor cell killing effect.
The results of our study are consistent with ones
from previous studies (18,19).
Therefore, the success of therapeutic vaccines will require
formulations that are effective in: i) generating new immune
responses, ii) boosting the existing immune responses, and iii)
overcoming immune evasion mechanisms (20–22).
In conclusion, the results from our study suggest
that the transfected DC vaccine enhances T lymphocyte proliferation
and induces CTL cells to kill gastric cancer and promote the
secretion of more IL-12 and IFN-γ. The results reported here
provide the experimental basis for the clinical treatment of
gastric cancer.
Acknowledgements
This study was supported by the Hebei
Natural Sciences Foundation (No. C2008000966).
References
|
1.
|
SW LeeY ParkT SoBS KwonH CheroutreRS
MittlerM CroftIdentification of regulatory functions for 4-1BB and
4-1BBL in myelopoiesis and the development of dendritic cellsNat
Immunol9917926200818604213
|
|
2.
|
C Reis e SousaDendritic cells in a mature
ageNat Rev Immunol6476483200616691244
|
|
3.
|
AK Thomas-KaskelH VeelkenActive
immunotherapy of prostate cancer with a focus on dendritic
cellsActas Urol Esp31668679200717896564
|
|
4.
|
J BanchereauAK PaluckaDendritic cells as
therapeutic vaccines against cancerNat Rev
Immunol5296306200515803149
|
|
5.
|
JS YuG LiuH YingWH YongKL BlackCJ
WheelerVaccination with tumor lysate-pulsed dendritic cells elicits
antigen-specific, cytotoxic T-cells in patients with malignant
gliomaCancer
Res6449734979200410.1158/0008-5472.CAN-03-350515256471
|
|
6.
|
T ChanA SamiA El-GayedX GuoJ XiangHER-2/
neu-gene engineered dendritic cell vaccine stimulates stronger
HER-2/neu-specific immune responses compared to DNA vaccinationGene
Ther1313911402200610.1038/sj.gt.330279716724093
|
|
7.
|
E HuarteJR Cubillos-RuizYC NesbethUK
ScarlettDG MartinezRJ BuckanovichF BenenciaRV StanT KelerP SarobeCL
SentmanJR Conejo-GarciaDepletion of dendritic cells delays ovarian
cancer progression by boosting antitumor immunityCancer
Res6876767684200818768667
|
|
8.
|
JE Moyron-QuirozJ Rangel-MorenoL HartsonK
KusserMP TigheKD KlonowskiL LefrançoisLS CauleyAG HarmsenFE LundTD
RandallPersistence and responsiveness of immunologic memory in the
absence of secondary lymphoid
organsImmunity25643654200617045819
|
|
9.
|
GK KoskiPA CohenRE RosesS XuBJ
CzernieckiReengineering dendritic cell-based anti-cancer
vaccinesImmunol Rev222256276200818364007
|
|
10.
|
KR JordanRH McMahanCB KemmlerJW KapplerJE
SlanskyPeptide vaccines prevent tumor growth by activating T cells
that respond to native tumor antigensProc Natl Acad Sci
USA10747884789201020133772
|
|
11.
|
P KrauseM BrucknerC UermösiE SingerM
GroettrupDF LeglerProstaglandin E(2) enhances T-cell proliferation
by inducing the costimulatory molecules OX40L, CD70, and 4-1BBL on
dendritic
cellsBlood11324512460200910.1182/blood-2008-05-15712319029446
|
|
12.
|
KH YiH NechushtanWJ BowersGR WalkerY
ZhangDG PhamER PodackHJ FederoffKA TolbaJD RosenblattAdoptively
transferred tumor-specific T cells stimulated ex vivo using herpes
simplex virus amplicons encoding 4-1BBL persist in the host and
show antitumor activity in vivoCancer
Res671002710037200710.1158/0008-5472.CAN-06-2391
|
|
13.
|
Y KuangX WengX LiuH ZhuZ ChenB JiangH
ChenAnti-tumor immune response induced by dendritic cells
transduced with truncated PSMA IRES 4-1BBL recombinant
adenovirusesCancer
Lett293254262201010.1016/j.canlet.2010.01.01120149524
|
|
14.
|
KA Shafer-WeaverSK WatkinsMJ AndersonLJ
DraperA MalyguineWG AlvordNM GreenbergAA HurwitzImmunity to murine
prostatic tumors: continuous provision of T-cell help prevents CD8
T-cell tolerance and activates tumor-infiltrating dendritic
cellsCancer Res6962576264200919622771
|
|
15.
|
M Habib-AgahiTT PhanPF
SearleCo-stimulation with 4-1BB ligand allows extended T-cell
proliferation, synergizes with CD80/CD86 and can reactivate anergic
T cellsInt Immunol1913831394200710.1093/intimm/dxm10617977894
|
|
16.
|
S RadhakrishnanKR WiehagenV PulkoV Van
KeulenWA FaubionKL KnutsonLR PeaseInduction of a Th1 response from
Th2-polarized T cells by activated dendritic cells: dependence on
TCR:peptide-MHC interaction, ICAM-1, IL-12, and IFN-gammaJ
Immunol17835833592200710.4049/jimmunol.178.6.3583
|
|
17.
|
EL GautierT HubyF Saint-CharlesB
OuzilleauJ PiraultV DeswaerteF GinhouxER MillerJL WitztumMJ
ChapmanP LesnikConventional dendritic cells at the crossroads
between immunity and cholesterol homeostasis in
atherosclerosisCirculation11923672375200910.1161/CIRCULATIONAHA.108.80753719380622
|
|
18.
|
ML DrakesSJ CzinnTG BlanchardRegulation of
murine dendritic cell immune responses by Helicobacter felis
antigenInfect Immun7446244633200610.1128/IAI.00289-0616861650
|
|
19.
|
MR JenkinsJ MinternNL La GrutaK
KedzierskaPC DohertySJ TurnerCell cycle-related a quisition of
cytotoxic mediators defines the progressive differentiation to
effector status for virus specific CD8+ T cellsJ
Immunol18138183822200810.4049/jimmunol.181.6.381818768835
|
|
20.
|
RK SharmaKG ElpekES YolcuRH SchabowskyH
ZhaoL Bandura-MorganH ShirwanCostimulation as a platform for the
development of vaccines: a peptide-based vaccine containing a novel
form of 4-1BB ligand eradicates established tumorsCancer
Res6943194326200910.1158/0008-5472.CAN-08-314119435920
|
|
21.
|
T KurodaY KitadaiS TanakaX YangN MukaidaM
YoshiharaK ChayamaMonocyte chemoattractant protein-1 transfection
induces angiogenesis and tumorigenesis of gastric carcinoma in nude
mice via macrophage recruitmentClin Cancer
Res1176297636200510.1158/1078-0432.CCR-05-0798
|
|
22.
|
SJ XieQH YanBE ShanZX FuFJ MengBD LiJH
CaiIn vitro experimental study individualised immunotherapy induced
by dendritic cells transfected with total RNA of autologous gastric
cancer cellsXi Bao Yu Fen Zi Mian Yi Xue Za Zhi2292952006
|