Introduction
Proinflammatory cytokines, such as IL-1β, IFN-γ and
TNF-α play an important role in the damage of insulin-producing
pancreatic β cells (1). It has
been well documented that these inflammatory cytokines induce the
apoptotic or necrotic destruction of β cells. Recent study has
shown that attenuation/prevention of pancreatic β-cell damage
induced by inflammatory cytokines is an important step for
treatment of diabetes (2).
Prunella vulgaris (P. vulgaris) is a
perennial herb with worldwide distribution and has been used as a
traditional medicine to reduce sore throat, alleviate fever and
accelerate wound healing in China for many years (3,4).
P. vulgaris contains different bioactive components,
including complex carbohydrates, hydrophobic metabolites
(triterpenes), polysaccharides and Rosmarinic acid (5,6).
Numerous studies have reported that extracts of P. vulgaris
have properties which are involved in anti-oxidative stress,
anti-microbial invasion (7,8),
anti-inflammatory responses (9)
and anti-DNA damage and caspase-3 activity (10). We previously reported that PVAE
significantly reduces postprandial hyperglycemia in ICR mice
(11). However, it is unclear
whether PVAE has a protective effect on inflammatory
cytokine-induced pancreatic β cell damage.
IL-1β induces pancreatic β cell apoptosis,
stimulates JNK phosphorylation and activates nuclear factor-κB
(NF-κB) (12), which results in
increases in inflammatory cytokine expression. In contrast,
inhibition of JNK activation protects β cells from IL-1β-induced
apoptosis (13). In the present
study, we investigated whether PVAE prevents IL-1β-induced
pancreatic β cell apoptosis and attenuates IL-1β-increased NF-κB
activation. We observed that PVAE significantly prevented
IL-1β-induced pancreatic β cell damage and attenuated
IL-1β-stimulated NF-κB activation and inflammatory cytokine
expression. Our data indicate that PVAE may have a significant
benefit for diabetes patients.
Materials and methods
Preparation of PVAE
Prunella vulgaris plants were collected in
Nanjing, China and identified by Dr Junsong Li (College of
Pharmacy, Nanjing University of Chinese Medicine). A voucher
specimen was deposited at the College of Pharmacy, Nanjing
University of Chinese Medicine. The procedure of PVAE preparation
was previously described by Kim et al (14) with modification. Briefly, P.
vulgaris was extracted with distilled water at 70°C for 5 h.
The extracts were filtered through Whatman No. 1 filter paper and
the filtrates were lyophilized. The final dried extracts were
dissolved in saline and filtered using a 0.45-μm syringe prior to
use in the in vitro experiments.
Cell culture and treatment
INS-1 pancreatic β cells were obtained from the
American Type Culture Collection (ATCC, Manassas, VA). INS-1 cells
were maintained in RPMI-1640 (Cellgro, USA) medium containing 10%
FBS, streptomycin (100 µg/ml) and penicillin (100 U/ml) at 37°C in
an incubator with a humidified atmosphere of 5% CO2
(15). To examine the cytotoxicity
of the PVAE preparation, INS-1 cells were plated at
1×104/ml and treated with increasing concentrations of
PVAE at 0.01, 0.1, 1, 10 and 100 μg/ml for 48 h. To investigate the
protective effect of PVAE on IL-1β-induced cell damage, INS-1 cells
(2×106/ml) were treated with PVAE at 100 µg/ml 30 min
before the cells were stimulated with IL-1β (10 ng/ml) for 48 h.
There were three replicates in each group. The supernatants were
harvested for measurement of LDH activity. The cells were harvested
for preparing the nuclear and cytoplasmic proteins as described
previously (16).
Cell cytotoxicity assay
The cell viability was measured using the MTT assay
(16). The LDH activity was
measured according to the manufacturer's instructions
(Sigma-Aldrich).
Western blot analysis
Western blots were performed as described previously
(17). Briefly, the cellular
proteins were separated by SDS-polyacrylamide gel electrophoresis
and transferred onto Hybond ECL membranes (Amersham Pharmacia,
Piscataway, NJ). The ECL membranes were incubated with the
appropriate primary antibody [anti-P-IκB, anti-IκB, anti-P-P38,
anti-p38, anti-P-JNK, anti-JNK, anti-P-ERK, anti-ERK, anti-cleaved
caspase-3, anti-Fas, anti-FasL, anti-Bax, anti-Bcl-2, anti-GAPDH
(Cell Signaling Technology, Inc.)], respectively, followed by
incubation with peroxidase-conjugated second antibodies (Cell
Signaling Technology, Inc.). The membranes were analyzed by the ECL
system (Amersham Pharmacia). The signals were quantified by G:Box
(Syngene).
Electrophoretic mobility shift assay
(EMSA)
NF-κB binding activity was examined in the nuclear
protein preparation using a LightShift Chemiluminescent EMSA kit
(Thermo Scientific) according to the instructions of the
manufacturer.
ELISA
The levels of cytokines (TNF-α and IL-6) in the
supernatants were measured using commercially available ELISA kits
(Peprotech, USA) according to the instructions provided by the
manufacturer.
Statistics
Results are expressed as means ± SEM. Statistical
significance was determined by one-way analysis of variance (ANOVA)
followed by the Tukey-Kramer multiple comparisons test. A
significant value was defined as p<0.05.
Results
PVAE attenuates IL-1β-induced
cytotoxicity in INS-1 cells
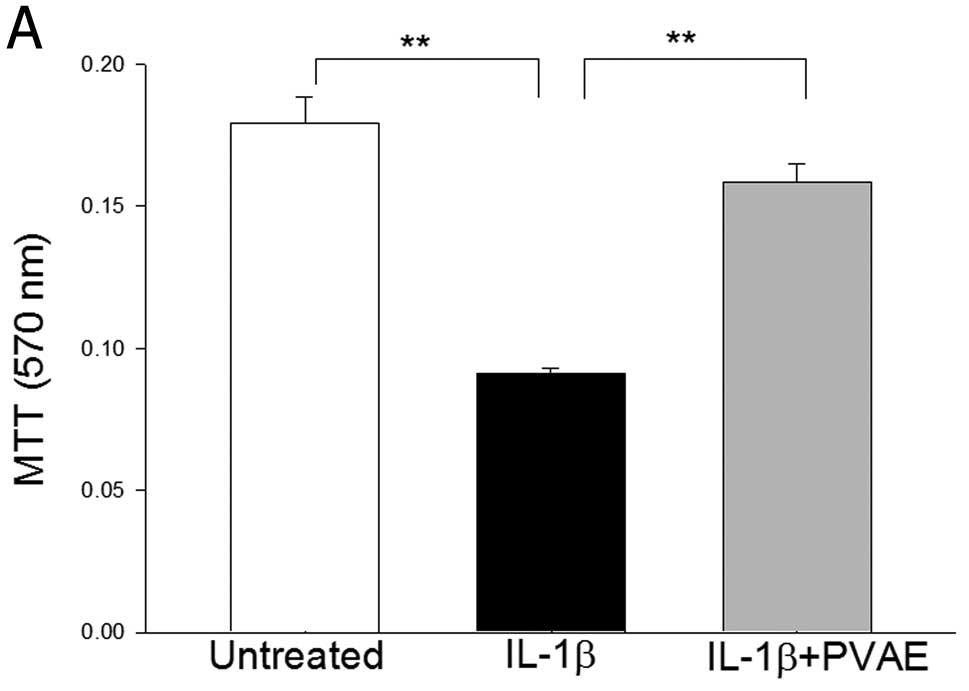
We examined the effect of PVAE on IL-1β-induced
cytotoxicity in INS-1 pancreatic β cells. INS-1 cells were treated
with IL-1β (10 ng/ml) in the presence and absence of PVAE (100
µg/ml) for 48 h. Cell cytotoxicity was examined by MTT and LDH
activity assay (16),
respectively. IL-1β treatment significantly induced cell death
(Fig. 1A) and increased LDH
activity by 1.51-fold (Fig. 1B)
compared with the untreated cells. In contrast, PVAE administration
significantly attenuated IL-1β-induced cell death (Fig. 1A) and reduced IL-1β-increased LDH
activity (Fig. 1B). The data
suggest that PVAE has a protective effect on IL-1β-induced
cytotoxicity in INS cells.
We also examined whether PVAE itself was cytotoxic
to INS-1 cells. INS-1 cells were treated with PVAE at increasing
concentrations for 48 h. The cell viability was measured by MTT. As
shown in Fig. 1C, PVAE treatment
did not significantly induce cell death compared with untreated
control, suggesting that PVAE does not have cytotoxic effects on
INS cells.
PVAE attenuates IL-1β-activated
FasL/Fas-mediated apoptotic signaling in INS-1 cells
Next, we examined how PVAE attenuates IL-1β-induced
cell death. It is well known that IL-1β can induce apoptosis in β
cells (18). Therefore, we
examined the protective effect of PVAE on IL-1β-induced apoptosis
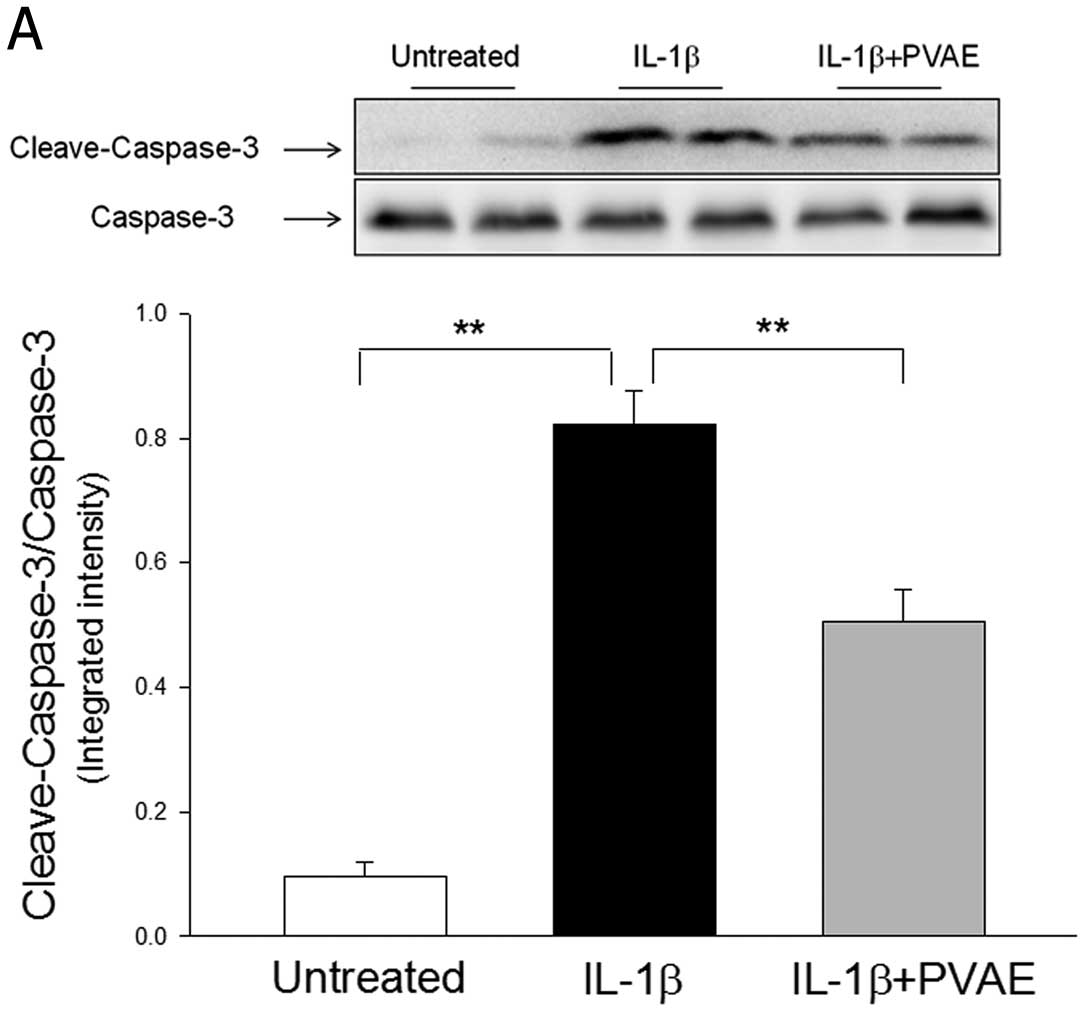
in INS-1 cells. Caspase-3 activity is a specific marker for
apoptosis (19). Fig. 2A shows that IL-1β treatment
significantly increased caspase-3 activity by 7.6-fold compared
with control cells. In the PVAE-treated cells, IL-1β-increased
caspase-3 activity was significantly reduced by 38.4%.
Activation of apoptotic signaling can be induced by
the mitochondria pathway and the death receptor pathway (1). Fig.
2B shows that IL-1β treatment did not alter the levels of Bax
and Bcl-2, indicating that IL-1β-induced INS-1 cell apoptosis was
not mediated by the mitochondria pathway. PVAE administration also
did not affect the levels of Bax and Bcl-2 in INS-1 cells (Fig. 2B). However, IL-1β stimulation
significantly increased the levels of Fas and FasL in INS-1 cells
compared with untreated cells. PVAE administration significantly
attenuated IL-1β-increased Fas and FasL (Fig. 2C). The data indicate that PVAE
attenuates IL-1β-induced apoptosis in INS-1 cells, in part, by
regulating the Fas/FasL apoptotic signaling pathway.
PVAE decreases IL-1β-increased
phosphorylation of JNK in INS-1 cells
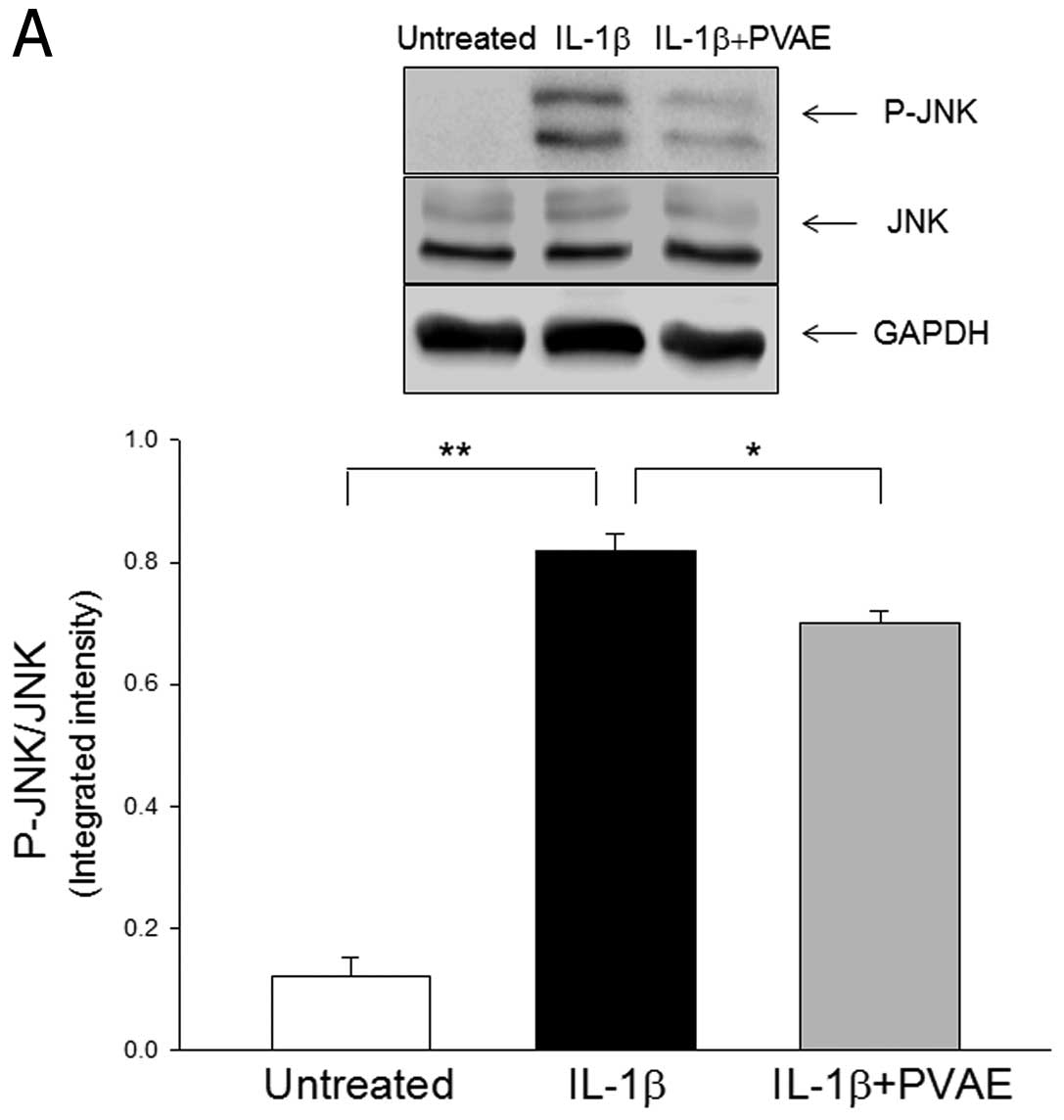
It is well known that the JNK signaling pathway
plays a critical role in activating apoptosis (20). We examined whether the
anti-apoptotic effect of PVAE involves blunting JNK activation. We
observed that IL-1β treatment significantly increased JNK
phosphorylation by 5.7-fold compared with untreated cells (Fig. 3). In the presence of PVAE, however,
IL-1β-increased levels of phosphorylated JNK was significantly
attenuated (Fig. 3A). However,
IL-1β treatment did not increase the levels of p38 and ERK
phosphorylation in the INS-1 cells (Fig. 3B and C). PVAE administration also
did not affect IL-1β-mediated changes in phosphorylation of p38 and
ERK in INS-1 cells (Fig. 3B and
C). The data suggest that the anti-apoptotic effect of PVAE is
mediated, in part, by attenuating IL-1β-activated JNK
signaling.
PVAE attenuates IL-1β-increased NF-κB
activation in INS-1 cells
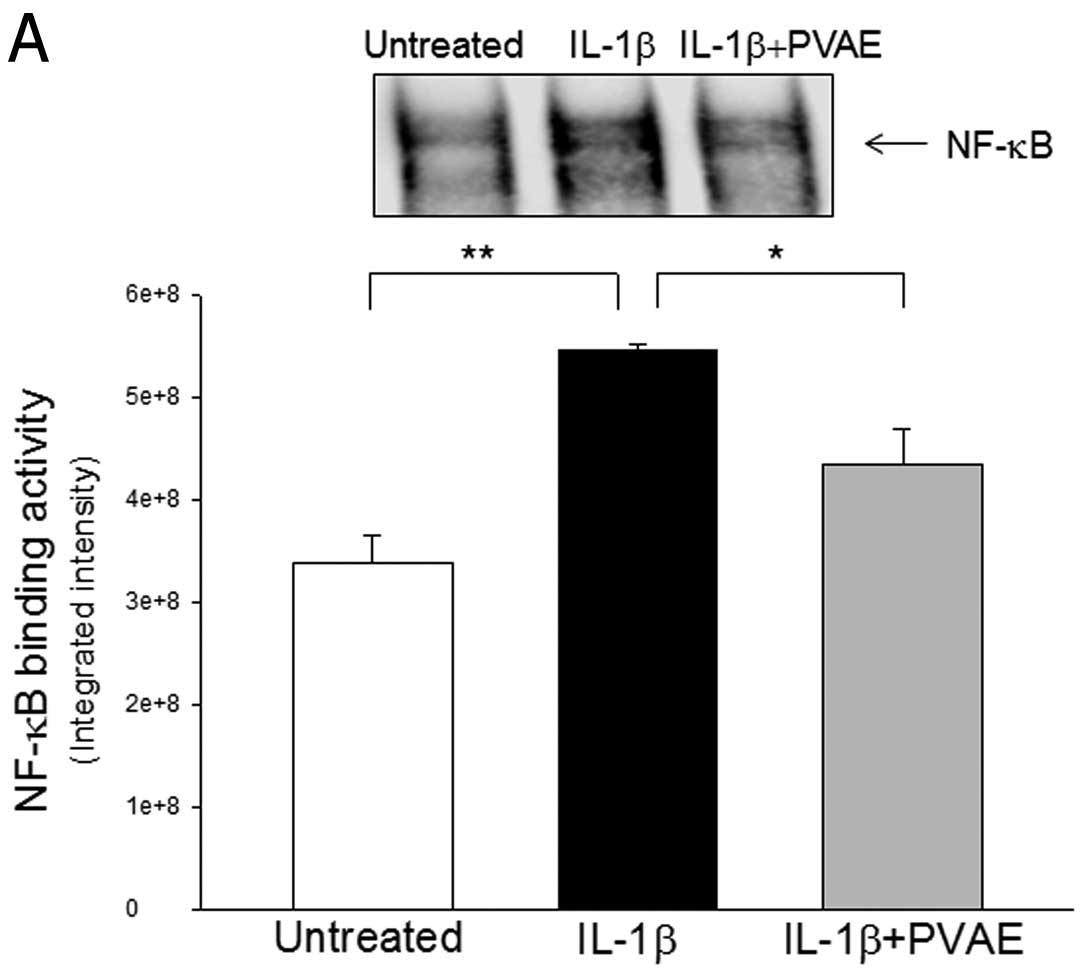
IL-1β is an important inflammatory mediator of
diabetes and stimulates NF-κB activation which controls
inflammatory cytokine gene expression (21). We examined the effect of PVAE on
IL-1β-stimulation of NF-κB activation in INS-1 cells. It was found
that IL-1β treatment significantly increased NF-κB binding activity
(Fig. 4A) by 61.1% and IκBα
phosphorylation (Fig. 4B) by
2.5-fold compared with untreated cells. In contrast, PVAE
administration significantly attenuated IL-1β-increased levels of
phosphorylated IκBα and NF-κB binding activity, respectively.
PVAE attenuates IL-1β-increased
inflammatory cytokine expression by INS-1 cells
Activation of NF-κB stimulates inflammatory cytokine
expression (16). We examined
whether PVAE attenuates IL-β-increased pro-inflammatory cytokine
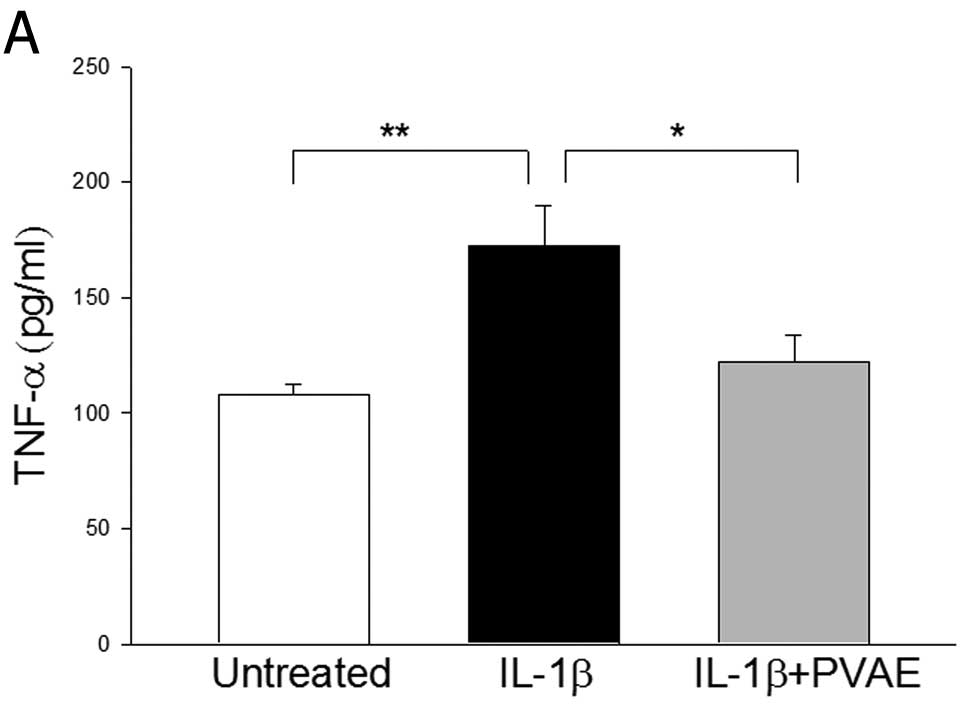
expression (Fig. 5). As expected,
IL-1β stimulation significantly increased the levels of TNF-α
(Fig. 5A) and IL-6 (Fig. 5B) in the supernatants of cultured
INS-1 cells by 59.4% and 1.1-fold, respectively. However, PVAE
administration significantly attenuated IL-1β-increased expression
of both TNF-α and IL-6 in INS-1 cells. The data indicate that PVAE
exerts an anti-inflammatory effect.
Discussion
A significant finding in the present study was that
PVAE significantly prevented IL-1β-induced pancreatic β cell
apoptosis and attenuated IL-1β-increased NF-κB activation,
resulting in decreases in IL-1β-increased TNF-α and IL-6 production
in INS-1 cells. Our data indicate that PVAE may be a novel
treatment for diabetic patients.
Prunella vulgaris has been used as a
traditional medicine for treatment of herpetic keratitis (22). Recent studies have reported that
Prunella vulgaris extracts have anti-oxidant and
anti-microbial properties (7,8). We
previously reported that administration of PVAE to rats
significantly reduced postprandial hyperglycemia in ICR mice
(11). PVAE treatment also
regulated glucose transport gene expression (SGLT-1, GLUT-2, and
Na+-K+-ATPase) in Caco-2 cells (23). It has been demonstrated that innate
immune and inflammatory responses play an important role in the
pathophysiologic mechanisms leading to the development of diabetes.
For example, inflammatory cytokines, such as TNF-α, IL-1β and IL-6,
induce pancreatic β cell apoptosis and promote inflammatory
responses in pancreatic islets, resulting in damage of β cells
(24). Therefore,
anti-inflammatory responses by pharmacological approaches will
prevent inflammatory cytokine-induced cytotoxicity in pancreatic β
cells.
We observed in the present study that PVAE
administration significantly prevented IL-1β-induced pancreatic β
cell (INS-1) cytotoxicity and apoptosis. The protective effect of
PVAE involved the attenuation of the IL-1β-increased
Fas/FasL-mediated apoptotic signaling pathway. Our observation was
supported by numerous studies. For example, Jacobsen et al
reported that IL-1β induced insulin-producing cell apoptosis
through the Fas-mediated apoptotic signaling pathway (25). We also observed that PVAE
administration significantly attenuated IL-1β-induced increases in
JNK phosphorylation in INS-1 cells. Activation of MAPK, including
JNK plays an important role in cell proliferation, differentiation
and cell death. IL-1β can activate pancreatic β cell c-Jun
N-terminal kinase (JNK), extracellular signal-regulated kinase
(ERK) and p38 (26). Larsen et
al reported that the inhibition of JNK activity by a novel
inhibitor attenuated the pancreatic β cell pro-apoptotic response
to IL-1β stimulation (13).
Størling et al suggested that Ca(2+) plays a permissive role
in IL-1β activation of the JNK signaling pathway in
insulin-secreting cells (27).
When taken together, PVAE prevented IL-1β-induced pancreatic β cell
apoptosis via downregulation of both Fas/FasL and JNK-mediated
apoptotic signaling pathways. Thus, PVAE maintains the function of
β cells by influencing β cell proliferation and death.
Activation of the NF-κB signaling pathway
contributes to dysfunction of pancreatic islet β cells (28). A number of proinflammatory
cytokines can activate NF-κB to regulate both the survival and
death of β cells. Thus, inflammatory cytokines and chemokines play
an important role in the development of the inflammatory infiltrate
in insulitis in the early stages of diabetes (21). IL-1β has been reported to induce
NF-κB activation in INS-1 cells (28). We observed that PVAE administration
significantly protected INS-1 cells from IL-1β-induced NF-κB
activation. Moreover, PVAE prevented IL-1β-increased levels of IL-6
and TNF-α in INS-1 cells. Recent studies have shown the PVAE has an
anti-inflammatory effect. Choi et al reported that PVAE
suppressed phorbol myristate acetate (PMA)-induced NF-κB activation
(16). When considered as a whole,
PVAE exhibited a significant anti-inflammatory effect via the
inhibition of NF-κB activation.
In summary, our data suggest that PVAE has a
protective effect on IL-1β-induced pancreatic β cell apoptosis and
cytotoxicity. The mechanisms involve attenuation of IL-1β-activated
Fas/FasL and JNK apoptotic signaling pathways and prevention of
IL-1β-activated NF-κB, resulting in decreases in inflammatory
cytokine (TNF-α and IL-6) production. Thus, PVAE may be a novel
treatment for diabetic patients.
Acknowledgements
This study was supported, in part, by
the National Natural Science Foundation of China (30772851) and
Administration of Chinese Pharmaceutical Medicine in Jiangsu
Province (HZ07061).
References
|
1.
|
Grunnet LG, Aikin R, Tonnesen MF, et al:
Proinflammatory cytokines activate the intrinsic apoptotic pathway
in β-cells. Diabetes. 58:1807–1815. 2009.PubMed/NCBI
|
|
2.
|
Akerfeldt MC, Howes J, Chan JY, et al:
Cytokine-induced β-cell death is independent of endoplasmic
reticulum stress signaling. Diabetes. 57:3034–3044. 2008.
|
|
3.
|
Pinkas M, Trotin F, Peng M, et al: Use,
chemistry and pharmacology of the Chinese medicinal plants.
Fitotherapia. 55:343–353. 1994.
|
|
4.
|
Chiej R: The MacDonald Encyclopaedia of
Medicinal Plants. MacDonald Co; Edinburgh: 1984
|
|
5.
|
Tabba HD, Chang RS and Smith KM:
Isolation, purification, and partial characterization of prunellin,
an anti-HIV component from aqueous extracts of Prunella
vulgaris. Antiviral Res. 11:263–273. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
6.
|
Psotova J, Kolar M, Sousek J, et al:
Biological activities of Prunella vulgaris extracts.
Phytother Res. 17:1082–1087. 2003.
|
|
7.
|
Chiu LC, Zhu W and Ooi VE: A
polysaccharide fraction from medicinal herb Prunella
vulgaris downregulates the expression of herpes simplex virus
antigen in Vero cells. J Ethnopharmacol. 93:63–68. 2004.PubMed/NCBI
|
|
8.
|
Kageyama S, Kurokawa M and Shiraki K:
Extract of Prunella vulgaris spikes inhibits HIV replication
at reverse transcription in vitro and can be absorbed from
intestine in vivo. Antivir Chem Chemother. 11:157–164. 2000.
|
|
9.
|
Ryu SY, Oak MH, Yoon SK, et al:
Anti-allergic and anti-inflammatory triterpenes from the herb of
Prunella vulgaris. Planta Med. 66:358–360. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
10.
|
Psotova J, Svobodova A, Kolarova H, et al:
Photoprotective properties of Prunella vulgaris and
rosmarinic acid on human keratinocytes. J Photochem Photobiol B.
84:167–174. 2006.
|
|
11.
|
Guo Y, Wu H, Gao M, et al: Effects of the
water extract of Prunella vulgaris L. on postprandial
hyperglycemia in ICR mice. J Southeast University. 25:2010.
|
|
12.
|
Karlsen AE, Heding PE, Frobøse H, et al:
Suppressor of cytokine signaling (SOCS)-3 protects β cells against
IL-1β-mediated toxicity through inhibition of multiple nuclear
factor-κB-regulated proapoptotic pathways. Diabetologia.
47:1998–2011. 2004.
|
|
13.
|
Larsen CM, Døssing MG, Papa S, et al:
Growth arrest- and DNA-damage-inducible 45β gene inhibits c-Jun
N-terminal kinase and extracellular signal-regulated kinase and
decreases IL-1β-induced apoptosis in insulin-producing INS-1E
cells. Diabetologia. 49:980–989. 2006.
|
|
14.
|
Kim SY, Kim SH, Shin HY, et al: Effects of
Prunella vulgaris on mast cell-mediated allergic reaction
and inflammatory cytokine production. Exp Biol Med (Maywood).
232:921–926. 2007.
|
|
15.
|
Jensen J, Galsgaard ED, Karlsen AE, et al:
STAT5 activation by human GH protects insulin-producing cells
against interleukin-1β, interferon-γ and tumour necrosis
factor-α-induced apoptosis independent of nitric oxide production.
J Endocrinol. 187:25–36. 2005.PubMed/NCBI
|
|
16.
|
Choi JH, Han EH, Hwang YP, et al:
Suppression of PMA-induced tumor cell invasion and metastasis by
aqueous extract isolated from Prunella vulgaris via the
inhibition of NF-κB-dependent MMP-9 expression. Food Chem Toxicol.
48:564–571. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17.
|
Ha T, Xia Y, Liu X, et al: Glucan
phosphate attenuates myocardial HMGB1 translocation in severe
sepsis through inhibiting NF-κB activation. Am J Physiol Heart Circ
Physiol. 301:H848–H855. 2011.PubMed/NCBI
|
|
18.
|
Barthson J, Germano CM, Moore F, et al:
Tumor necrosis factor-α and interferon-γ induce pancreatic β-cell
apoptosis through STAT1-mediated Bim activation. J Biol Chem.
286:39632–39643. 2011.
|
|
19.
|
Fox R and Aubert M: Flow cytometric
detection of activated caspase-3. Methods Mol Biol. 414:47–56.
2008.PubMed/NCBI
|
|
20.
|
Cui J, Zhang M, Zhang YQ, et al: JNK
pathway: diseases and therapeutic potential. Acta Pharmacol Sin.
28:601–608. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21.
|
Haddad JJ and Abdel-Karim NE: NF-κB
cellular and molecular regulatory mechanisms and pathways:
therapeutic pattern or pseudoregulation. Cell Immunol. 271:5–14.
2011.
|
|
22.
|
Zheng M: Experimental study of 472 herbs
with antiviral action against the herpes simplex virus. Zhong Xi Yi
Jie He Za Zhi. 10:39–41. 1990.(In Chinese).
|
|
23.
|
Wu H, Ha T and Gao M: Prunella
vulgaris L. extract decreasing mRNA expressions of
α-glucosidase SGLT-1, GLUT-2 and
Na+-K+-ATPase in Caco-2 cells. Zhongguo Sheng
Hua Yao Wu Za Zhi. 31:373–376. 2010.
|
|
24.
|
Han X, Sun Y, Scott S, et al: Tissue
inhibitor of metalloproteinase-1 prevents cytokine-mediated
dysfunction and cytotoxicity in pancreatic islets and β-cells.
Diabetes. 50:1047–1055. 2001.PubMed/NCBI
|
|
25.
|
Jacobsen ML, Rønn SG, Bruun C, et al:
IL-1β-induced chemokine and Fas expression are inhibited by
suppressor of cytokine signalling-3 in insulin-producing cells.
Diabetologia. 52:281–288. 2009.
|
|
26.
|
Cargnello M and Roux PP: Activation and
function of the MAPKs and their substrates, the MAPK-activated
protein kinases. Microbiol Mol Biol Rev. 75:50–83. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
27.
|
Størling J, Zaitsev SV, Kapelioukh IL, et
al: Calcium has a permissive role in interleukin-1β-induced c-jun
N-terminal kinase activation in insulin-secreting cells.
Endocrinology. 146:3026–3036. 2005.PubMed/NCBI
|
|
28.
|
Friberg J, Tonnesen MF, Heller S, et al:
Inhibition of the nuclear factor-κB pathway prevents beta cell
failure and diet induced diabetes in Psammomys obesus. PLoS
One. 5:133412010.
|