Introduction
Studies investigating the role of tumor-associated
antigens (TAAs) in cancer diagnosis and progression have focused on
the MUC1 antigen, which is overexpressed in most solid epithelial
and non-solid hematological tumors including multiple myeloma
(1–3). Soluble MUC1 (sMUC1) levels were
reported to be elevated in sera of patients with solid tumors such
as breast carcinoma, and non-solid tumors such as multiple myeloma,
using FDA approved sMUC1 assays CA15.3 and CA27.29 directed to the
extracellular tandem repeat array (TRA) domain (3,4).
However, despite the wide tumor distribution of MUC1, use of the
CA15.3 and CA27.29 markers is confined to monitoring the prognosis
and response to treatment in patients with advanced breast cancer
(5) and it is not sensitive enough
to be used for early diagnosis.
In contrast, naturally generated autoantibodies to
TAAs are detectable even before the tumor is clinically apparent
(6), and due to their lower
fluctuation and longer half-life in the blood, they may be more
appropriate for cancer diagnosis. Anti-MUC1 TRA autoantibody levels
were shown to be higher in patients with breast cancer (7) and lower in multiple myeloma (3) patients compared to healthy
individuals. One suggested cause for the variations in multiple
myeloma patients relates to the presence of immune complexes
between sMUC1 and the endogenously generated anti-MUC1 TRA
antibodies (3). Bypassing this
limitation can potentially be achieved by searching for anti-MUC1
autoantibodies generated to domains flanking the TRA.
Along with others, we have demonstrated the
preferred immunogenicity of the signal peptide (SP) domain of MUC1
(8–10). This short domain has promiscuous
binding to multiple major histocompatibility complex (MHC) class I
and II, a characteristic that supports its process and presentation
on the cell surface of various tumor cells and antigen presenting
cells (10). In the present study,
we questioned whether MUC1’s SP domain preferred MHC class II
binding, coupled with in silico prediction of B-cell
epitopes, could lead to the induction of a natural anti-SP humoral
response in multiple myeloma patients.
Materials and methods
Naïve donors and cancer patients
Blood samples (3 ml) were drawn from 15 naïve
healthy volunteers, 18–60 years of age and 27 patients with
multiple myeloma, 50–75 years of age. The study patients included
14 with progressive disease under treatment, 7 with active disease
under treatment and 6 at best response off therapy. The study was
approved by the Institutional Ethics Committees of the
participating hospitals.
In silico prediction software
Binding predictions were performed for the MHC class
II (HLA-DRB1) alleles that are most prevalent worldwide. However,
to have a defined population, we focused only on the Caucasian
population. MHC class II binding prediction was carried out using
Propred http://www.imtech.res.in/raghava/propred/ (11) and Immune Epitope www.immuneepitope.org (12). B-cell epitope prediction was
evaluated with Kolaskar and Tongaonkar antigenicity score using
immune epitope http://tools.immuneepitope.org/tools/bcell/DisplayResultServlet
(13). In all prediction methods,
only binders >5% were analyzed.
Peptide synthesis
MUC1-SP-L, MUC1-SP-M, TB-Rv0476/4941-SP-L were
synthesized by fully automated, solid-phase, peptide synthesis
using fluorenylmethyloxycarbonyl (Fmoc)/tBu-strategy and
Rink-amide-polystyrene resin at EMC Microcollections, Germany,
while MUC1-TRA-L was synthesized using the same methodology at GL
Biochem, China. The purity and identity of all peptides was
>95%, as determined by HPLC and MS analysis.
ELISA for detecting serum levels of sMUC1
and anti-MUC1 IgG antibodies
Soluble MUC1 levels were evaluated in ELISA plates
(F96 Maxisorp, Nunc, Denmark) using commercial anti-MUC1 TRA
monoclonal antibodies (mAbs) (clones M4H2 and M2F1 HRP-conjugate)
and the ELISA kit (HyTest, Finland) according to the manufacturer’s
protocol. MUC1 levels were evaluated using 7 double dilutions of
100 μl of patient sera starting at 1:5. For a MUC1-positive
control, we used dilutions (starting at 1:5) of supernatant
collected from the DA3-GTRUNK transfected cell line, producing high
levels of sMUC1 containing the TRA domain (14). The ELISA plates were developed with
TMB/E solution (Southern Biotech, USA) according to the
manufacturer’s protocol. The reaction was terminated by adding 50
μl/well of 10% sulfuric acid. Results were measured at 450 nm. For
this assay, we used specific titer rather than absolute
concentration, as we did not have the appropriate pure antigen as a
standard.
Soluble MUC1 SP levels were evaluated in ELISA
plates (F96 Maxisorp) coated for 2 h with 5 μg/ml of the anti-SP
polyclonal antibodies raised in rabbits to the 17-mer MUC1-SP-M.
Next, dilutions of 100 μl of patient sera, (starting at 1:5) were
incubated for 2 h. For detection we used 1 μg/ml of
biotinconjugated anti-MUC1-SP-M antibodies for 1 h, followed by 1 h
incubation at 25°C with streptovidin HRP (BioLegent, USA) diluted
1:10000. At the final step, the ELISA plates were developed with
TMB/E solution as described above. As a standard for sMUC1 SP, we
used dilutions, starting from 2.5 μg/ml of MUC1’s 21-mer SP domain,
MUC1-SP-L (10).
To evaluate the level of anti-MUC1 TRA and SP
antibodies in sera, ELISA plates (F96 Maxisorp) were coated with 50
μl of MUC1 peptide at 5 μg/ml in carbonate buffer and incubated
overnight at 4°C. Evaluated serum samples were then diluted 1:100,
plus 7 additional dilutions in PBS with 0.5% gelatin and incubated
for 2 h at 25°C. Next, 50 μl/well of the appropriate secondary
anti-IgG antibody HRP-conjugate (Jackson ImmunoResearch, USA) was
added at a final dilution of 1:10000 in a blocking buffer and
incubated for 1 h at 25°C. Plates were then developed with TMB/E
solution as described above. As a positive standard for anti-MUC1
TRA antibodies, we used dilutions starting with 10 μg/ml of the
anti-MUC1 TRA mAb H23 (15),
raised against the human breast cancer cell line T47D (15) and recognized the TRA epitope APDTRP
on the non-glycosylated form of MUC1. As a positive standard for
anti-MUC1 SP antibodies, we used dilutions beginning with 10 μg/ml
of anti-MUC1-SP-M rabbit polyclonal antibodies. In this assay,
serum levels for anti-MUC1 TRA autoantibodies in naïve donors were
X≤274 μg/ml and for anti-MUC1 SP auto-antibodies, X≤210.2 μg/ml,
based on the average plus standard deviation value determined in 15
naïve healthy individuals.
Statistical analysis
Results were statistically analyzed with Student’s
t-test, Fisher’s exact test or in the case of correlations, with
Pearson correlation coefficient. In all methods, the minimum level
of significance for a two-tailed test was set at P<0.05.
Results
MUC1-derived SP epitope has high
antigenic profile and promiscuous MHC class II binding
Selection of appropriate immunodominant antigens for
the induction of a natural humoral response was performed using
various in silico prediction methods, as described in
Materials and methods. Using these tools, we searched the MUC1 SP
domain for sequences with the following properties: superior B-cell
antigenicity profile and preferred CD4+ T-cell
activation via high binding properties to a defined list of
abundant MHC class II alleles. The outcome of our search was the
selection for further use of the 17-mer peptide MUC1-SP-M
containing amino acids 10–21 in MUC1 SP (Table I). As a control from the known MUC1
TRA domain, we used the 25-mer MUC1 TRA-derived peptide MUC-TRA-L
(16,17). As a non-MUC1 SP-derived control, we
used a 19-mer SP sequence TB-Rv0476/4941-SP-L, which included the
entire SP domain of the Mycobacterium tuberculosis (MTb)
antigen Rv0476/4941 (18)
(Table I).
 | Table I.List of peptides used in this
study. |
Table I.
List of peptides used in this
study.
| VXL IDa | Published ID
(ref)b | Length (-mer) | Position | Sequence | Target
antigen/Domainc |
|---|
VXL100
MUC1-SP-L | MUC1-SP-L (10) | 21 | 1–21 |
MTPGTQSPFFLLLLLTVLTVV-NH2 | MUC1 (SP) |
VXL3A
MUC1-SP-M | - | 17 | 10–21 |
KKFLLLLLTVLTVVKKK | MUC1 (SP) |
VXL25
MUC1-TRA-L | BLP25 (16,17) | 25 | 130–154 |
STAPPAHGVTSAPDTRPAPGSTAPP | MUC1 (TRA) |
VXL211
TB-Rv0476/4941-SP-L | VXL211 (18) | 19 | 1–19 |
MLVLLVAVLVTAVYAFVHA-NH2 | Uncharacterized
protein Rv0476/4941 (SP) |
Endogenously generated anti-MUC1
antibodies in naïve donors
Our initial aim was to determine the presence of
endogenous anti-MUC1 SP IgG antibodies in naïve donors and multiple
myeloma patients. Tables II and
III summarize the results in 15
naïve healthy donors and 27 multiple myeloma cancer patients,
respectively, at different disease stages. A baseline of 210.2 μg/
ml for anti-MUC1 SP and 274 μg/ml for anti-MUC1 TRA was set for
positive autoantibody expression levels based on their expression
levels in naïve donors, as described in Materials and methods.
Interestingly, anti-MUC1 SP autoantibody serum levels were
significantly lower (P<0.008, t-test) than those of anti-MUC1
TRA IgG autoantibodies, suggesting a lower antigen expression
profile for SP vs. TRA in the naïve donors.
| Table II.sMUC1 and anti-MUC1 autoantibody
levels in naïve donors. |
Table II.
sMUC1 and anti-MUC1 autoantibody
levels in naïve donors.
| Sample no. | Sample
characterization | sMUC1 TRA
(μg/ml)a | sMUC1 SP
(μg/ml)a | Anti-MUC1 TRA Ab
(μg/ml)b | Anti-MUC1 SP Ab
(μg/ml)b |
|---|
| 1 | Naïve healthy
donor | Negative | Negative | 224 | 178 |
| 2 | Naïve healthy
donor | Negative | Negative | 265 | 159 |
| 3 | Naïve healthy
donor | Negative | Negative | 184 | 145 |
| 4 | Naïve healthy
donor | Negative | Negative | 161 | 185 |
| 5 | Naïve healthy
donor | Negative | Negative | 205 | 185 |
| 6 | Naïve healthy
donor | Negative | Negative | 230 | 200 |
| 7 | Naïve healthy
donor | Negative | Negative | 281 | 216 |
| 8 | Naïve healthy
donor | Negative | Negative | 318 | 10 |
| 9 | Naïve healthy
donor | Negative | Negative | 105 | 110 |
| 10 | Naïve healthy
donor | Negative | Negative | 320 | 166 |
| 11 | Naïve healthy
donor | Negative | Negative | 203 | 10 |
| 12 | Naïve healthy
donor | Negative | Negative | 147 | 200 |
| 13 | Naïve healthy
donor | Negative | Negative | 200 | 184 |
| 14 | Naïve healthy
donor | Negative | Negative | 134 | 100 |
| 15 | Naïve healthy
donor | Negative | Negative | 152 | 10 |
| Average | | | | 208.60 | 137.20 |
| Standard
deviation | | | | 65.35 | 72.97 |
| Table III.sMUC1 and anti-MUC1 autoantibody
levels in multiple myeloma patients. |
Table III.
sMUC1 and anti-MUC1 autoantibody
levels in multiple myeloma patients.
| Patient no. | Patient status | sMUC1 TRA
(titer)a | sMUC1 SP
(titer) | Anti-MUC1 TRA Ab
(μg/ml)b | Anti-MUC1 SP Ab
(μg/ml)b | Anti-MUC1 TRA Ab
(Positivity) | Anti-MUC1 SP Ab
(Positivity) |
|---|
| 1 | Active disease;
under therapy | Negative | Negative | 440 | 740 | + | + |
| 2 | Progressive
disease; under therapy | Negative | Negative | 183 | 150 | - | - |
| 3 | At best response;
off therapy | 1:10 | Negative | 382 | 312 | + | + |
| 4 | Active disease;
under therapy | 1:5 | Negative | 190 | 350 | - | + |
| 5 | Active disease;
under therapy | 1:10 | Negative | 85 | 50 | - | - |
| 6 | Active disease;
under therapy | Negative | Negative | 97 | 130 | - | - |
| 7 | At best response;
off therapy | 1:10 | Negative | 355 | 456 | + | + |
| 8 | At best response;
off therapy | Negative | Negative | 211 | 581 | - | + |
| 9 | Active disease;
under therapy | Negative | Negative | 132 | 1036 | - | + |
| 10 | At best response;
off therapy | Negative | Negative | 158 | 746 | - | + |
| 11 | Progressive
disease; under therapy | Negative | Negative | 238 | 525 | - | + |
| 12 | Progressive
disease; under therapy | Negative | Negative | 63 | 500 | - | + |
| 13 | At best response;
off therapy | Negative | Negative | 500 | 1000 | + | + |
| 14 | Active disease;
under therapy | 1:10 | Negative | 292 | 884 | + | + |
| 15 | Progressive
disease; under therapy | 1:40 | Negative | 980 | 3400 | + | + |
| 16 | At best response;
off therapy | Negative | Negative | 761 | 1500 | + | + |
| 17 | Progressive
disease; under therapy | 1:20 | Negative | 91 | 100 | - | - |
| 18 | Progressive
disease; under therapy | Negative | Negative | 728 | 424 | + | + |
| 19 | Progressive
disease; under therapy | 1:10 | Negative | 795 | 339 | + | + |
| 20 | Progressive
disease; under therapy | Negative | Negative | 120 | 80 | - | - |
| 21 | Progressive
disease; under therapy | 1:20 | Negative | 3200 | 812 | + | + |
| 22 | Progressive
disease; under therapy | Negative | Negative | 291 | 594 | + | + |
| 23 | Progressive
disease; under therapy | Negative | Negative | 500 | 732 | + | + |
| 24 | Progressive
disease; under therapy | 1:10 | Negative | 500 | 191 | + | - |
| 25 | Progressive
disease; under therapy | 1:20 | Negative | 860 | 950 | + | + |
| 26 | Active disease;
under therapy | 1:20 | Negative | 195 | 162 | - | - |
| 27 | Progressive
disease; under therapy | 1:5 | Negative | 222 | 976 | - | + |
| Percentage of
patients with positive anti-MUC1 SP- and MUC1 TRA-specific
antibodies levels | 14/27 | 20/27 |
Endogenously generated anti-MUC1
antibodies in patients with multiple myeloma
We next evaluated the levels of anti-MUC1 SP and
MUC1 TRA IgG autoantibodies in 27 multiple myeloma patients.
Although we observed elevated levels of anti-MUC1 TRA
autoantibodies in the entire group of 27 patients vs. the 15 naïve
donors, the differences were not significant (P=0.111, t-test).
Unexpectedly, we observed an elevation of up to 24-fold in the
concentration of autoantibodies to the MUC1 SP domain in most
patients (Table III). This
elevation was highly significant (P<0.004, t-test) in the entire
group of 27 multiple myeloma vs. the 15 naïve donors, in spite of
the small group size and disease stage variability. Anti-MUC1 SP
autoantibodies had antigen (i.e., MUC1) rather than general SP
specificity, since no reactivity to the non-TAA SP
TB-Rv0476/4941-SP-L was observed in several patients (data not
shown). Importantly, a high concentration of anti-MUC1 SP
autoantibodies was also present in patients with minimal disease
(characterized as ‘patients at best response off therapy’), four of
which had undetectable sMUC1 levels, suggesting a potential role
for these autoantibodies in monitoring minimal disease and
plausibly disease onset. The preferred immunogenicity of MUC1 SP
vs. TRA domains can be further demonstrated in the same group, in
which the concentration of the anti-SP autoantibodies was
significantly higher (P<0.03, t-test) than that of the anti-MUC1
TRA levels. In these patients, we ruled out any potential influence
on sMUC1 levels by anti-MUC1 TRA autoantibodies with a dedicated
ELISA that analyzed the amount of sMUC1 in antigen-antibody
complexes (data not shown). This supports the immunodominant
properties of MUC1 SP regarding antibody production as predicted
in silico.
We further analyzed the percentage of patients with
positive anti-MUC1 SP- and MUC1 TRA-specific titers. A positive
response was set for the average plus one standard deviation as
described in Materials and methods. Results in this analysis
(Table III) further supported our
initial observation on the elevated production of anti-MUC1 SP
autoantibodies. In particular, most patients, 20/27 (74%) had
positive anti-MUC1 SP-specific autoantibodies, while only 14/27
(51.8%) had positive anti-MUC1 TRA-specific autoantibodies.
Although these differences were not significant (P=0.158, Fisher’s
exact test), they showed a positive trend for the selectivity of
MUC1’s SP vs. TRA domain in multiple myeloma patients.
Expression levels of soluble MUC1 SP in
naïve donors and multiple myeloma patients
Previous studies have presented sMUC1 expression in
multiple myeloma patients, but none focused on the SP domain of
MUC1. This is a significant point, especially in light of the
knowledge about immune complexes between the sMUC1 containing the
TRA domain and anti-MUC1 TRA autoantibodies and their effect on
sMUC1 levels. We therefore questioned whether the high anti-MUC1 SP
autoantibody levels were merely an outcome of the immuno-dominant
properties of MUC1’s SP and/or a consequence of the minimal
interference by plausible novel sMUC1 variants containing the SP
domain. Our results (Tables II and
III) confirmed a negative
expression of sMUC1 containing the SP domain in all serum samples
of both healthy naïve donors and multiple myeloma cancer patients
and ruled out the presence of immune complexes around MUC1 SP
autoantibodies.
Correlation between serum levels of sMUC1
and anti-MUC1 autoantibodies
After finding elevated levels of anti-MUC1 TRA and
SP antibodies in the majority of the multiple myeloma patients, we
sought to explore any interactions between sMUC1 and the anti-MUC1
SP autoantibody levels in multiple myeloma patients at different
disease stages. We also wanted to reconfirm past observations in
multiple myeloma patients vis-à-vis an inverse correlation between
the levels of sMUC1 and anti-MUC1 TRA autoantibodies. In line with
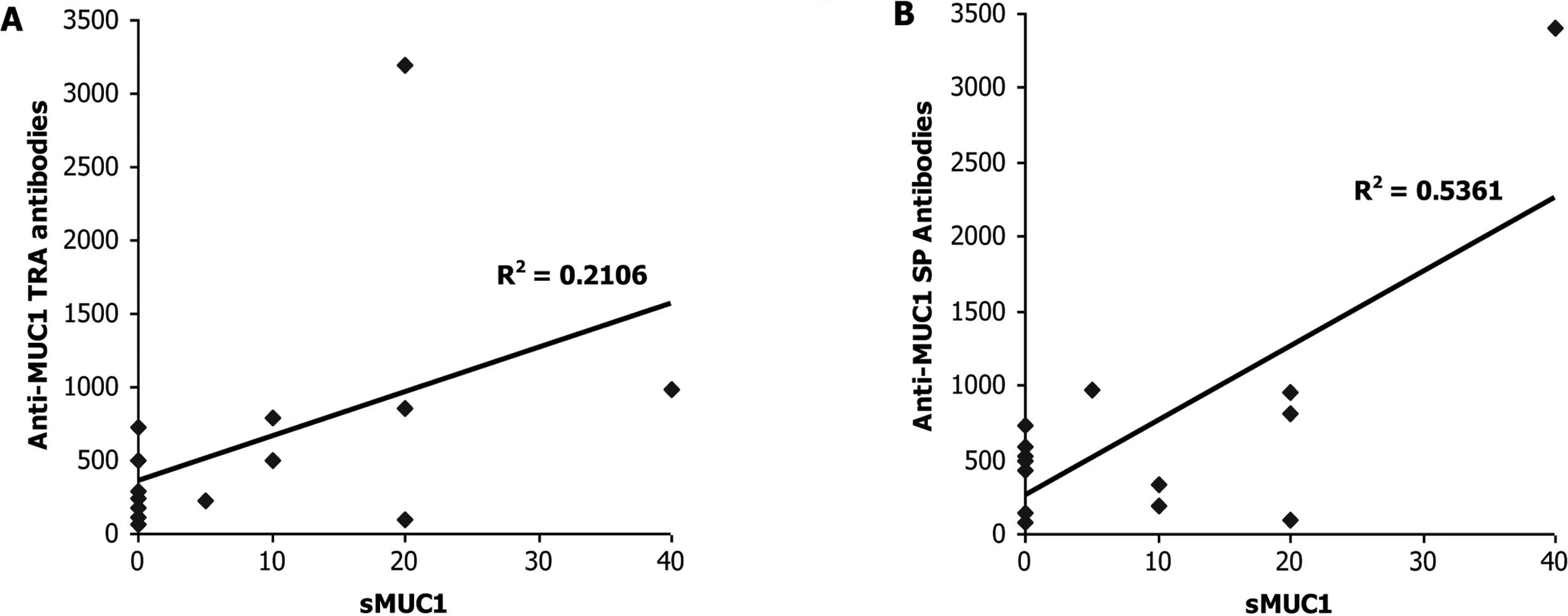
previous publications, we found no significant correlation
(R2=0.2106, P= 0.469, Pearson correlation) between sMUC1
levels and autoantibodies to MUC1 TRA among the group of 14
multiple myeloma patients characterized with ‘progressive disease
under therapy’ (Fig. 1A). In
contrast, a significant positive correlation (R2=0.5361,
P<0.048, Pearson correlation) was found between sMUC1 serum
levels and anti-MUC1 SP autoantibodies among this group of 14
multiple myeloma patients (Fig.
1B). These results were unexpected, especially since they were
obtained from a relatively small patient population without
selecting for key parameters such as age and gender, which can
influence the expression levels of MUC1 and anti-MUC1 antibodies.
No significant correlation was found in a similar analysis
performed on the other patients with multiple myeloma classified as
‘active disease under therapy’ or ‘patients at best response off
therapy’ with both anti-MUC1 SP and anti-MUC1 TRA autoantibodies
(data not shown).
Discussion
Despite the wide distribution and high
immunogenicity of MUC1 TAA and anti-MUC1 autoantibodies, their
utilization in cancer diagnosis and prognosis is still limited
(5). Whereas over the last three
decades, studies with sMUC1 have concentrated on the TRA domain
(19–22), here we suggest for the first time a
novel strategy focused on non-TRA, anti-MUC1 SP IgG
autoantibodies.
While naïve donors had significantly lower IgG
autoantibody levels to MUC1 SP compared to MUC1 TRA epitopes, the
levels in multiple myeloma patients were significantly higher than
that of MUC1 TRA. This outcome was related to the immunodominant
properties of SP also described in previous publications (9,10).
Baty and Lazdunski reported the induction of preferential IgG titer
to the small (Mr 2600) alkaline phosphatase SP domain (23) following vaccination with the much
larger (Mr 45600) precursor protein. In a different study, Harboe
et al reported the production of high polyclonal antibodies
in rabbits to a 13-mer fragment from the SP domain of the MTb
antigen, MPT83 (24).
The specific nature of the antigen leading to the
generation of anti-MUC1 SP autoantibodies is still unclear. In this
study, we showed the absence of the sMUC1 SP domain in serum
samples of naïve donors and multiple myeloma patients. Yet, it is
still possible that in multiple myeloma patients, undetectable
levels of SP fragments from debris of tumor cells could reach the
blood stream. An alternative source for MUC1 SP is cell surface
expression. We and others demonstrated the expression of MUC1 SP in
association with MHC class I and II in the context of cancer cells
(8–10). Since antibody production to
MHC-peptide complexes is not trivial due to the various binding
restrictions of different MHC alleles, a third, still unknown
mechanism could be the primary resource for the MUC1 SP
antigen.
Results in the current study suggested a
MUC1-specific rather than SP-specific autoantibody titer. Previous
reports showed either cross reactivity between different SP
sequences when a large precursor was used as an antigen (23) or antigen specificity when a linear
SP fragment was used (24). In our
opinion, these differences stem from the nature of the sequence
used for vaccination and its ability to form conformation
structures. A larger sequence, such as the one used by Baty and
Lazdunski (23), forms
conformational structures that lead to cross-reactivity between
different SPs. We plausibly overcame this conformation structure by
using only amino acids 10–21 from the total 21-mer of SP domain of
MUC1 and by adding several lysine residues at the N and C terminal
ends of this peptide, which form a linear structure. In summary, we
hypothesize that in addition to the antigen-specific antibodies
recognizing a linear sequence, there are potentially SP-specific
autoantibodies that recognize a SP conformation motif.
Past studies on multiple myeloma reported a negative
correlation between the levels of sMUC1 and anti-MUC1 TRA
autoantibodies (3). We confirmed
these findings, but also presented a statistically significant,
positive correlation between sMUC1 and anti-MUC1 SP autoantibody
levels in multiple myeloma patients with progressive disease under
therapy.
This is the first report on the presence of
autoantibodies to the MUC1 SP domain in cancer patients. Additional
analysis needs to be performed on a larger, homogeneous patient
population, especially at disease onset, before firm conclusions
can be made. One such analysis should involve the existence of IgM
autoantibodies to the MUC1 SP domain and the switch from IgM to
IgG. This would assist us to better understand the entire process
of anti-MUC1 SP antibody formation. Moreover, in-depth analysis
needs to be performed in breast cancer patients where the CA15.3
marker is still in use.
Abbreviations:
|
MHC,
|
major histocompatibility complex; mAb,
monoclonal antibody;
|
|
SP,
|
signal peptide;
|
|
sMUC1,
|
soluble MUC1;
|
|
TRA,
|
tandem repeat array;
|
|
TAA,
|
tumor-associated antigen
|
References
|
1.
|
Croce MV, Isla-Larrain MT, Demichelis SO,
Gori JR, Price MR and Segal-Eiras A: Tissue and serum MUC1 mucin
detection in breast cancer patients. Breast Cancer Res Treat.
81:195–207. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
2.
|
Luminari S, Goldaniga M, Ceccherelli F,
Guffanti A, Bombardieri E, Marcheselli R, et al: Prevalence and
prognostic significance of sMUC-1 levels in plasma cell dyscrasias.
Br J Haematol. 121:772–774. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
3.
|
Treon SP, Maimonis P, Bua D, Young G, Raje
N, Mollick J, et al: Elevated soluble MUC1 levels and decreased
anti-MUC1 antibody levels in patients with multiple myeloma. Blood.
96:3147–3153. 2000.PubMed/NCBI
|
|
4.
|
Reddish MA, MacLean GD, Poppema S, Berg A
and Longenecker BM: Pre-immunotherapy serum CA27.29 (MUC-1) mucin
level and CD69+ lymphocytes correlate with effects of
Theratope sialyl-Tn-KLH cancer vaccine in active specific immuno
therapy. Cancer Immunol Immunother. 42:303–309. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
5.
|
Gion M, Mione R, Leon AE and Dittadi R:
Comparison of the diagnostic accuracy of CA27.29 and CA15.3 in
primary breast cancer. Clin Chem. 45:630–637. 1999.
|
|
6.
|
Lu H, Goodell V and Disis ML: Humoral
immunity directed against tumor-associated antigens as potential
biomarkers for the early diagnosis of cancer. J Proteome Res.
7:1388–1394. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7.
|
MacLean GD, Reddish MA, Koganty RR and
Longenecker BM: Antibodies against mucin-associated sialyl-Tn
epitopes correlate with survival of metastatic adenocarcinoma
patients undergoing active specific immunotherapy with synthetic
STn vaccine. J Immunother Emphasis Tumor Immunol. 19:59–68. 1996.
View Article : Google Scholar
|
|
8.
|
Brossart P, Heinrich KS, Stuhler G, Behnke
L, Reichardt VL, Stevanovic S, et al: Identification of
HLA-A2-restricted T-cell epitopes derived from the MUC1 tumor
antigen for broadly applicable vaccine therapies. Blood.
93:4309–4317. 1999.PubMed/NCBI
|
|
9.
|
Carmon L, El-Shami KM, Paz A, Pascolo S,
Tzehoval E, Tirosh B, et al: Novel breast-tumor-associated
MUC1-derived peptides: characterization in Db-/- x beta2
microglobulin (beta2m) null mice transgenic for a chimeric
HLA-A2.1/Db-beta2 micro-globulin single chain. Int J Cancer.
85:391–397. 2000. View Article : Google Scholar
|
|
10.
|
Kovjazin R, Volovitz I, Kundel Y,
Rosenbaum E, Medalia G, Horn G, et al: ImMucin: a novel therapeutic
vaccine with promiscuous MHC binding for the treatment of
MUC1-expressing tumors. Vaccine. 29:4676–4686. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11.
|
Singh H and Raghava GP: ProPred:
prediction of HLA-DR binding sites. Bioinformatics. 17:1236–1237.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
12.
|
Vita R, Zarebski L, Greenbaum JA, Emami H,
Hoof I, Salimi N, et al: The immune epitope database 2.0. Nucleic
Acids Res. 38:D854–D862. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13.
|
Kolaskar AS and Tongaonkar PC: A
semi-empirical method for prediction of antigenic determinants on
protein antigens. FEBS Lett. 276:172–174. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
14.
|
Horn G, Gaziel A, Wreschner DH,
Smorodinsky NI and Ehrlich M: ERK and PI3K regulate different
aspects of the epithelial to mesenchymal transition of mammary
tumor cells induced by truncated MUC1. Exp Cell Res. 315:1490–1504.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
15.
|
Keydar I, Chou CS, Hareuveni M, Tsarfaty
I, Sahar E, Selzer G, et al: Production and characterization of
monoclonal antibodies identifying breast tumor-associated antigens.
Proc Natl Acad Sci USA. 86:1362–1366. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
16.
|
Palmer M, Parker J, Modi S, Butts C,
Smylie M, Meikle A, et al: Phase I study of the BLP25 (MUC1
peptide) liposomal vaccine for active specific immunotherapy in
stage IIIB/IV non-small cell lung cancer. Clin Lung Cancer.
3:49–58. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
17.
|
Sangha R and North S: L-BLP25: a
MUC1-targeted peptide vaccine therapy in prostate cancer. Expert
Opin Biol Ther. 7:1723–1730. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
18.
|
Kovjazin R, Volovitz I, Daon Y,
Vider-Shalit T, Azran R, Tsaban L, et al: Signal peptides and
trans-membrane regions are broadly immunogenic and have high
CD8+ T cell epitope densities: implications for vaccine
development. Mol Immunol. 48:1009–1018. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19.
|
Apostolopoulos V and McKenzie IF: Cellular
mucins: targets for immunotherapy. Crit Rev Immunol. 14:293–309.
1994. View Article : Google Scholar : PubMed/NCBI
|
|
20.
|
Graham RA, Burchell JM and
Taylor-Papadimitriou J: The polymorphic epithelial mucin: potential
as an immunogen for a cancer vaccine. Cancer Immunol Immunother.
42:71–80. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
21.
|
Ho SB, Niehans GA, Lyftogt C, Yan PS,
Cherwitz DL, Gum ET, et al: Heterogeneity of mucin gene expression
in normal and neoplastic tissues. Cancer Res. 53:641–651.
1993.PubMed/NCBI
|
|
22.
|
Tang CK, Katsara M and Apostolopoulos V:
Strategies used for MUC1 immunotherapy: human clinical studies.
Expert Rev Vaccines. 7:963–975. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23.
|
Baty D and Lazdunski C: An anti-(signal
peptide) antibody: purification, properties and use as a
conformational probe. Eur J Biochem. 102:503–507. 1979. View Article : Google Scholar : PubMed/NCBI
|
|
24.
|
Harboe M, Whelan AO, Ulvund G, McNair J,
Pollock JM, Hewinson RG, et al: Generation of antibodies to the
signal peptide of the MPT83 lipoprotein of Mycobacterium
tuberculosis. Scand J Immunol. 55:82–87. 2002. View Article : Google Scholar : PubMed/NCBI
|