Introduction
Aquaporin-2 (AQP2), a member of the AQP family,
plays a vital role in regulating the water balance in the body.
This protein is primarily observed in the collecting duct of the
kidneys (1). A significantly
increased expression of Aqp2 mRNA has been observed in the
kidneys of rats with spontaneous hypertension (2,3).
Similarly, clinical observations indicate that the urine AQP2
concentration is increased in hypertensive patients with low renin
levels (4). These findings
highlight a correlation between AQP2 expression and the occurrence
of hypertension.
AQP2 is regulated by the neurohypophysial hormone
arginine vasopressin (AVP or AVP-V2) through the AVP receptor on
AQP2 (3). The AVP receptor is
regulated, in part, by angiotensin II (AngII), a peptide hormone
that induces vasoconstriction (5).
AngII is formed by the conversion of angiotensin I (AngI) by
angiotensin-converting enzyme (ACE) (6). ACE inhibitors, including imidapril,
block the conversion of AngI to AngII (7). A previous study of another ACE
inhibitor, enalapril, indicated that treatment of hypertensive rats
with enalapril reduces the overexpression of AQP2 and AVP-V2 in the
kidneys (8). However, it is not
known whether imidapril is able to regulate the expression or
secretion of AQP2.
In the current study, changes in AQP2 expression in
the kidneys, AQP2 concentration in the urine and plasma AVP
concentration in hypertensive rats were detected by molecular,
immunological and biochemical techniques prior to and following the
administration of imidapril. These analyses are likely to advance
our understanding of the regulation of AQP2 by ACE inhibitors in
hypertension.
Materials and methods
Rats
Pathogen-free male Wistar rats (body weight, 120–140
g) were purchased from Vital River Lab Animal Technology (Beijing,
China). Rats were housed three or four per cage and maintained at
constant temperature (22±2°C) and constant humidity (55±5%) with a
12-h light/dark cycle. Food and water were available ad
libitum. To induce hypertension, rats were gavaged at 40
mg/kg/day with N-nitro-L-nitro-arginine-methyl-ester (L-NAME;
Sigma, St. Louis, MO, USA), a nitric oxide synthase inhibitor. On
week 3, caudal arterial blood pressure (BP) and heart rate were
determined with an RBP-I-type rat BP and heart rate machine
(Clinical Research Institute, China-Japan Friendship Hospital,
Beijing, China). Rats with systolic BP >18.67 kPa (140 mmHg; 24
rats) were then randomized to either the control group or the
imidapril treatment group (n=12 per group). Following measurements
of BP and body mass, rats in the treatment group received an
intragastric injection of 2.5 mg/kg/day imidapril (Tianjin Tanabe
Seiyaku Co., Ltd., China) at 09:00 daily for 8 weeks. Control rats
received an equal volume of water in the same manner. BP and body
mass measurements were performed once a month.
Specimens
Rats were placed in metabolic cages prior to
decapitation to collect 24-h urine output. Freshly collected urine
was centrifuged at low temperature at 2,000 rpm for 5–10 min. The
supernatant was collected and centrifuged at low temperature at
2,000 rpm for 75 min using an ultrafiltration tube (centriplus-10;
Millipore, Billerica, MA, USA) to concentrate the urine to <1
ml. Concentrated urine was stored at −70°C for future use. Rats
were anesthetized intraperitoneally with 10% chloral hydrate at 3
ml/kg and decapitated following blood collection. Kidneys were
rapidly removed and washed with phosphate-buffered saline (PBS).
The renal medulla was separated according to gross anatomy and
placed in a tube containing liquid nitrogen. Blood samples (1 ml)
were collected in 0.13 mM disodium ethylenediamine tetraacetate
(EDTA-2Na) solution (20 ml/l whole blood) and centrifuged at 4°C at
3,000 rpm for 10 min. Plasma was collected and stored at −70°C for
detection of AVP concentration. The urine osmolality was detected
by the freezing point inhibition method and serum Na+
concentration.
Plasma AVP concentration
The plasma AVP concentration was measured by
radioimmunoassay using a plasma AVP radioimmunoassay kit (DSL
Biological Products, Webster, TX, USA) according to manufacturer’s
instructions. The FM-2000γ immune counter (Xi’an Kaipu Electrical,
China) was used to detect radiolabeling.
AQP2 expression in the kidney
Immunohistochemistry
Tissues near the renal medulla were fixed in neutral
formalin for 24 h, then treated using conventional histological
methods for paraffin embedding. Paraffin-embedded tissues were
sectioned at 4 μm with a paraffin slicer (Leica RM2016,
Wetzlar, Germany) and collected on glass slides. Sections were
dewaxed with xylenes and rehydrated with an alcohol gradient.
Antigen repair was performed with citric acid solution and
microwave heating. Room-temperature slides were treated with 3%
hydrogen peroxide to block activity of endogenous peroxidase.
Sections were then covered with non-specific serum in a humidified
box and incubated at room temperature. The primary antibody against
AQP2 (rabbit anti-rat/mouse IgG; Calbiochem, San Diego, CA, USA)
was applied to sections, which were then incubated in a humidified
box at 4°C overnight. For the negative control, goat serum was used
in place of the primary antibody. After washing three times with
PBS, 50 μl biotin-labeled secondary antibody (Santa Cruz
Biotechnology Inc., Santa Cruz, CA, USA) were added and the
sections were incubated at room temperature. After washing three
times with PBS, 50 μl ready-to-use streptavidin-horseradish
peroxidase (HRP, SP kit; Maixin Inc., Fuzhou, China) was added to
the sections, which were then incubated at 37°C for 30 min.
3,3′-Diaminobenzidine (DAB) substrate (Maixin Inc.) was applied to
develop color. Staining results were observed under a light
microscope (Olympus, Tokyo, Japan) before termination of the
reaction. Sections were then counterstained with hematoxylin,
dehydrated and mounted for visualization. Positive staining appears
as brownish-yellow puncta in the cell membranes and cytoplasm of
the kidney specimens. Image-Pro Plus 6.0 image analysis software
(Media Cybernetics Inc., Silver Spring, MD, USA) was used to detect
staining. Five visual fields were randomly selected to determine
the percentage of positively-stained cells.
RT-PCR
Total RNA was extracted from 100 mg tissue near the
renal medulla using a TRIzol RNA extraction kit (Takara Biotech
Co., Ltd., Dalian, China). The A260/A280 ratio of the total RNA
(752C spectrophotometer Shanghai Spectrum Instruments Co., Ltd.,
Shanghai, China) from collected samples was calculated to be
118–210. Total RNA (2 μg) was used for reverse transcription
to synthesize cDNA using M-MuLV (MBI Fermentas, Burlington, Canada)
as reverse transcriptase in a 20-μl reaction. cDNA (2
μl) was amplified by the GeneAmp PCR system with 18S rRNA
(forward: 5′-CGACGGACCCATTCGAACGTCT-3′ and reverse:
5′-GCTATTGGAGCTGGAATTACCG-3′) and Aqp2 (forward:
5′-CATGTCTCCTTCCTTCGAGC-3′ and reverse: 5′-TTGTGGAGAGCATTGACAGC-3′)
primers and Taq polymerase (Bio-Asia Diagnostics Co., Ltd.,
Shanghai, China). Based on the published sequence of Aqp2
(GenBank Accession No. NM_000486), the primers were designed and
synthesized by Takara Biotech Co., Ltd. The following reaction
conditions were used: initial incubation for 5 min at 95°C,
followed by 30 cycles of 30 sec at 94°C, 30 sec at 60°C, 60 sec at
72°C and 7 min at 72°C. PCR products were separated in ethidium
bromide 2% sepharose gels and visualized with a gel imaging system.
The expected sizes of the amplification products were 131 bp for
Aqp2 and 312 bp for 18S rRNA. 18S rRNA was used as the
internal standard to perform semi-quantitative analysis for PCR
products, for which relative absorbance was measured using image
analysis software.
Western blotting
Kidney tissues were homogenized with histone
solution using a Polytron high-speed homogenizer. Proteins were
denatured by boiling samples for 3 min. Proteins were separated via
12% sodium dodecyl sulfate (SDS)-polyacrylamide gel electrophoresis
prior to transfer to a nitrocellulose membrane. Proteins on
nitrocellulose membranes were blocked with 5% skimmed milk at 37°C
for 2 h, then AQP2 primary antibody (rabbit anti-rat/mouse IgG) was
added to the membrane for incubation at 4°C overnight. Membranes
were washed with Tris-buffered saline with Tween-20 (TBST) at 37°C
three times for 10 min each. Biotin-labeled secondary antibody
(Santa Cruz Biotechnology Inc.) was added prior to incubation at
37°C for 40 min. Following three more washes with TBST, the
staining color was developed by the addition of an enhanced
chemiluminescence (ECL) reagent for 3–5 min. A gel imaging system
was used to visualize and compare the protein bands. Expression was
normalized against β-actin, which was used as an internal
control.
AQP2 concentration in urine
The AQP2 concentration in the urine was determined
by enzyme-linked immunosorbent assay (ELISA) according to
previously published methods (9,10).
Briefly, the square matrix titration method was used to detect the
optimum working concentration of antibodies; the primary antibody
(rabbit anti-AQP2 purified polyclonal antibody) was optimized at 2
mg/l and the secondary antibody (HRP goat anti-rabbit IgG; New
England Biolabs, Beijing, China) was optimized at 1:1000. The AQP2
positive control, polypeptide standard preparation coupled with
bovine serum albumin (BSA; Alpha Diagnostic, San Antonio, TX, USA),
was diluted with 0.05% SDS-PBS at a 1:1 ratio. Then, 100 μl
diluted urine samples and 100 μl standard preparation were
added to a 96-well plate. Negative control wells contained 100
μl 0.05% SDS. Plates were pre-coated at 37°C for 30 min,
then incubated at 4°C overnight. Each well was washed with 0.05%
PBS with Tween-20 (PBST) and incubated with 100 μl blocking
solution (3% BSA-PBS) at 37°C for 45 min. Next, 0.25% BSA-PBS was
added to each well (diluted to 100 μl). The primary antibody
against AQP2 (2 mg/l) was added to the wells and plates were
incubated at 37°C for 2 h. Following four 1-min washes with PBST,
the secondary antibody diluted with 0.25% BSA-PBS was incubated in
the wells at 37°C for 90 min. Following another four washes with
PBST, 100 μl fresh 0.01% TMB substrate buffer was added to
each well for incubation at 37°C for 15 min. Finally, 2 μM
H2SO4 was added to each well to terminate the
reaction. Absorbance was measured at 450 nm on a microplate reader
(ELX800 enzyme-linked immunosorbent detector; Dio-Tek Instruments
Inc., Winooski, VT, USA). The standard curve was determined using
the absorbance of the standard solution.
Statistical analysis
SPSS 17.0 statistical software (SPSS Inc., Chicago,
IL, USA) was used for statistical analyses. Measurement data are
expressed as the mean ± standard deviation. The independent-sample
t-test was used to analyze and compare differences in the
intergroup indices. α=0.05 and P<0.05 were considered to
indicate a statistically significant difference.
Results
Imidapril affects BP, urine output and
urine osmolality in hypertensive rats
All experimental rats were induced to exhibit
hypertension, then randomly assigned to a control group (water) or
treatment group (imidapril). Following treatment, the rats were
assessed for changes in BP and kidney function (Table I). Compared with rats in the
control group, rats treated with imidapril exhibited decreased
systolic BP, increased 24-h urine output and decreased urine
osmolality (P<0.05). However, no significant difference was
observed in Na+ level between the two groups.
 | Table IComparison of systolic pressure,
Na+ concentration, 24-h urine output and urine osmotic
pressure in hypertensive rats left untreated or treated with
imidapril. |
Table I
Comparison of systolic pressure,
Na+ concentration, 24-h urine output and urine osmotic
pressure in hypertensive rats left untreated or treated with
imidapril.
| Treatment group | n | Systolic pressure
(mmHg) | Na+
(mmol/l) | 24-h urine volume
(ml) | Osmotic pressure
(mOsm/kg H2O) |
|---|
| Control | 8 | 155.1±17.6 | 146.3±6.8 | 11.3±2.1 | 1818.6±118.6 |
| Imidapril | 8 | 132.3±20.1 | 151.3±6.3 | 17.0±2.2 | 1311.8±77.4 |
| t-value | | 2.965 | 1.862 | 6.653 | 12.393 |
| P-value | | 0.007 | 0.076 | 0.001 | 0.001 |
Effects of imidapril on AQP2 in the
kidneys of hypertensive rats
To determine whether the modulating effects of
imidapril on kidney function in hypertensive rats involved changes
in AQP2 expression, we used molecular and immunological techniques
to assess the expression of Aqp2 mRNA and the protein
product. Semi-quantitative mRNA expression was determined by RT-PCR
in hypertensive rats in the control and imidapril-treated groups
and was normalized against 18S rRNA expression. Compared with the
control group (0.73±0.07), relative Aqp2 expression in
imidapril-treated rat kidneys was significantly lower (0.46±0.07,
t=9.263, P=0.001; Fig. 1).
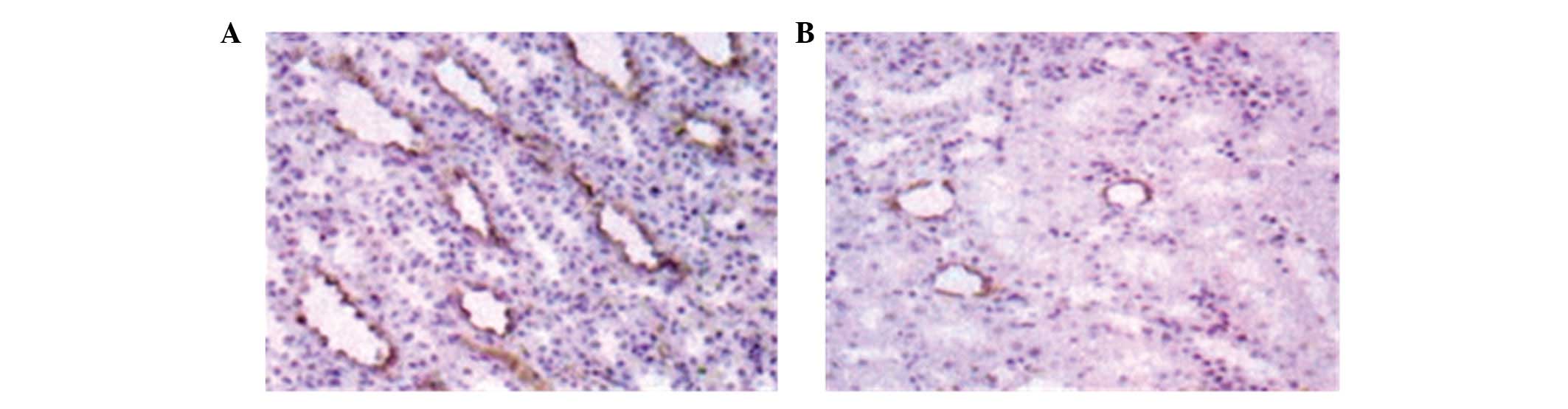
Immunohistochemistry against AQP2 was performed on
sections from hypertensive rat kidneys. As expected, AQP2 was
detected predominantly in cells surrounding the collecting tube in
tissues near the renal medulla (Fig.
2). However, AQP2 staining in the kidneys of imidapril-treated
rats was significantly lighter (Fig.
2B) compared with that of rats in the control group.
Additionally, the positively stained area (0.46±0.07) of kidney
sections from rats in the imidapril group was significantly smaller
compared with that of the control group (0.80±0.08, t=11.154,
P=0.001; Fig. 3).
Given the reduced staining intensity observed by
immunohistochemistry, we sought to quantify AQP2 expression in rat
kidneys using western blot analysis. AQP2 expression was normalized
against β-actin expression. Relative AQP2 expression in the kidneys
of imidapril-treated rats (0.76±0.06) was significantly lower
compared with that of rats in the control group (t=10.371, P=0.001;
Fig. 4).
Imidapril affects plasma AVP and urine
AQP2 concentration in hypertensive rats
Imidapril treatment significantly altered the
concentration of AVP in the plasma and of AQP2 in the urine of
hypertensive rats (Table II).
Compared with the control group, the plasma AVP concentration of
imidapril-treated rats was significantly reduced; by contrast, the
urine AQP2 concentration was significantly increased following
imidapril treatment (P<0.05).
| Table IIComparison of plasma AVP and AQP2
concentrations in hypertensive rats left untreated or treated with
imidapril. |
Table II
Comparison of plasma AVP and AQP2
concentrations in hypertensive rats left untreated or treated with
imidapril.
| Treatment group | n | AVP (ng/l) | AQP2
(μg/l) |
|---|
| Control | 8 | 81.9±12.0 | 12.2±1.3 |
| Imidapril | 8 | 50.2±8.6 | 19.9±3.3 |
| t-value | | 7.439 | 7.466 |
| P-value | | 0.001 | 0.001 |
Discussion
AQP2 is the key protein regulating the water
permeability of the renal collecting duct; therefore, it is
critical in maintaining the renal water balance (10,11).
AQP2 operates through short- and long-term regulatory mechanisms
(12–14). AQP2 is also the only AVP-dependent
AQP. Elevated AVP content in plasma promotes AQP2 expression in the
epithelium of the renal collecting duct, opening the water channel
and increasing water reabsorption, which results in urine
concentration and increased infiltration capacity (15).
The renin-angiotensin-aldosterone system (RAAS) is
an endocrine pathway that regulates water and electrolyte balance,
blood volume and BP through a number of hormones and enzymes
(16,17). Renin is a proteolytic enzyme that
is synthesized and secreted by the juxtaglomerular cells (18) and promotes the conversion of plasma
pro-angiotensin to AngI (19). The
conversion of AngI by ACE produces AngII, a BP-boosting protein
that promotes the vasoconstriction of small arteries and indirectly
increases BP (20). AngII also
stimulates the adrenal zona to produce greater quantities of
aldosterone, promotes the absorption of sodium and chloride ions by
distal tubules, increases blood volume and leads to increased
BP.
Imidapril, a new type of highly selective ACE
inhibitor, when administered orally, becomes the active metabolite,
imidaprilat, through liver de-esterification (20,21).
Imidaprilat inhibits the activity of ACE and prevents the
conversion of AngI to AngII. This causes peripheral vasodilatation
and reduces vascular resistance, thus producing an antihypertensive
effect (7). Additionally,
imidapril reduces aldosterone secretion, increases Na+
discharge and simultaneously reduces glomerular perfusion pressure
and increases renal blood flow, thus increasing urine volume
(22). Therefore, it is not
surprising that we identified that imidapril treatment
significantly reduces BP and urine osmolality while also increasing
24-h urine output in hypertensive rats.
By investigating the changes induced by imidapril
treatment, we sought to determine whether treatment affects the
expression of AQP2. mRNA and protein levels were reduced in
imidapril-treated hypertensive rats compared with control
hypertensive rats. Furthermore, urine AQP2 concentrations were
significantly increased following imidapril treatment. These
findings indicate that imidapril downregulates AQP2 expression in
renal tissues and increases AQP2 urine excretion. The inhibitory
effect of imidapril on AQP2 expression may involve imidapril
preventing AngII generation and subsequently reducing the
stimulatory effect of AngII on the expression of AVP-V2 receptor
mRNA, thus indirectly inhibiting the expression of AQP2. We also
identified that plasma AVP concentrations significantly decreased.
Since AQP2 is mainly located in the cytoplasm of cells and the
membrane of tubules of the renal collecting duct and a certain
amount of AQP2 protein enters the tubules and is washed away by
urine, the AQP2 concentration in urine is correlated with the
effects of AVP in plasma and AQP2 expression in the kidney.
References
|
1.
|
Nielsen S, Frør J and Knepper MA: Renal
aquaporins: key roles in water balance and water balance disorders.
Curr Opin Nephrol Hypertens. 7:509–516. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
2.
|
Nielsen S, Kwon TH, Christensen BM,
Promeneur D, Frøkiaer J and Marples D: Physiology and
pathophysiology of renal aqua-porins. J Am Soc Nephrol. 10:647–663.
1999.
|
|
3.
|
Huang JH, Xie LD, Jiang DW, et al: Effect
of vasopressin on the expression of aquaporin 2 in kidney in WKY
rats. Chin J Hypertens. 16:427–430. 2008.(In Chinese).
|
|
4.
|
Ou YS, Chen W and Kuang XB: Effect of
urinary aquaporin-2 in low renin essential hypertension. Chin J
Cardiovasc Rev. 6:55–58. 2008.(In Chinese).
|
|
5.
|
Matsukawa T and Miyamoto T: Angiotensin
II-stimulated secretion of arginine vasopressin is inhibited by
atrial natriuretic peptide in humans. Am J Physiol Regul Integr
Comp Physiol. 300:R624–R629. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6.
|
Erdös EG: Conversion of angiotensin I to
angiotensin II. Am J Med. 60:749–759. 1976.
|
|
7.
|
Hosoya K and Ishimitsu T: Protection of
the cardiovascular system by imidapril, a versatile
angiotensin-converting enzyme inhibitor. Cardiovasc Drug Rev.
20:93–110. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
8.
|
Wong NL and Tsui JK: Upregulation of
vasopressin V2 and aquaporin 2 in the inner medullary collecting
duct of cardiomyopathic hamsters is attenuated by enalapril
treatment. Metabolism. 51:970–975. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
9.
|
Jiang RY, Xu DL, Lai WY, et al:
Quantitative measurement of urinary excretion of aquaporin-2 water
channel protein in rat by indirect ELISA. Chin J Geriat Heart Brain
Vessel Dis. 7:260–262. 2005.(In Chinese).
|
|
10.
|
Lu W, Xu D and Yin X: Development of
double antibody sandwich ELISA assay in detection of urinary
aquaporin-2 concentration. Journal of First Military Medical
University. 22:486–489. 2002.
|
|
11.
|
Hasler U, Mordasini D, Bens M, Bens M,
Bianchi M, Cluzeaud F, Rousselot M, Vandewalle A, Feraille E and
Martin PY: Long term regulation of aquaporin-2 expression in
vasopressin-responsive renal collecting duct principal cells. J
Biol Chem. 277:10379–10386. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
12.
|
Deen PM, van Balkom BW and Kamsteeg EJ:
Routing of the aquaporin-2 water channel in health and disease. Eur
J Cell Biol. 79:523–530. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
13.
|
Zhu TY, Gu Y, Huang GY, et al: Expression
of channel protein mRNA in kidney of congestive heart failure and
its significance. Chin J Nephrol. 18:438–441. 2002.(In
Chinese).
|
|
14.
|
Earm JH, Christensen BM, Frøkiaer J,
Marples D, Han JS, Knepper MA and Nielsen S: Decreased aquaporin-2
expression and apical plasma membrane delivery in kidney collecting
ducts of polyuric hypercalcemic rats. J Am Soc Nephrol.
9:2181–2193. 1998.PubMed/NCBI
|
|
15.
|
Pedersen RS, Bentzen H, Bech JN and
Pedersen EB: Effect of an acute oral lithium intake on urinary
aquaporin-2 in healthy humans with and without simultaneous
stimulation with hypertonic saline infusion. Scand J Clin Lab
Invest. 63:181–194. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
16.
|
Weir MR and Dzau VJ: The
renin-angiotensin-aldosterone system: a specific target for
hypertension management. Am J Hypertens. 12:205S–213S. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
17.
|
Sigmund CD: Divergent mechanism regulation
fluid intake and metabolism by the brain renin-angiotensin system.
Am J Physiol Integr Comp Physiol. 302:313–320. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18.
|
Kon Y: Comparative study of
renin-containing cells: histological approaches. J Vet Med Sci.
61:1075–1086. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
19.
|
Mesquita FF, Gontijo JA and Boer PA:
Expression of renin-angiotensin system signaling compounds in
material protein-restricted rates: effects on renal sodium
excretion and blood pressure. Nephrol Dial Transplant. 25:380–388.
2012. View Article : Google Scholar
|
|
20.
|
Song JC and White CM: Clinical
pharmacokinetics and selective pharmacodynamics of new angiotensin
converting enzyme inhibitors: an update. Clin Pharmacokinet.
41:207–224. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
21.
|
Guo XG, Uzui H, Mizuguchi T, Ueda T, Chen
JZ and Lee JD: Imidaprilat inhibits matrix metalloproteinase-2
activity in human cardiac fibroblasts induced by interleukin-1beta
via NO-dependent pathway. Int J Cardiol. 126:414–420. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
22.
|
Naitoh M, Suzuki H, Arakawa K, et al: Role
of kinin and renal AngII blockade in acute effects of ACE
inhibitors in low-renin hypertension. Am J Physiol. 272:H679–H687.
1997.PubMed/NCBI
|