Introduction
Coronary artery disease and hypertension account for
the majority of the myocardial abnormalities that occur in
diabetes. However, previous studies have shown that diabetes
mellitus alters cardiac structure and function independently of
coronary artery disease and systemic hypertension, a condition
known as diabetic cardiomyopathy (DCM) (1,2). DCM
is characterized by systolic and diastolic dysfunction due to
reduced contractility, prolonged relaxation and decreased
compliance of the myocardium (3,4). The
pathological mechanism of DCM is considered to involve myocardial
apoptosis and necrosis, reactive hypertrophy, myocardial fibrosis,
endothelial dysfunction, disturbance of the management of the
metabolic cardiovascular load and cardiac autonomic neuropathy
(4–6). Although the features of DCM are
well-identified, the pathogenesis underlying the myocardial
remodeling process has not been elucidated, and no effective
treatment strategy is available.
Neuregulin-1 (NRG-1), a cardioactive growth factor
released from endothelial cells, is indispensable for cardiac
development and the structural maintenance and functional integrity
of the heart (7,8). In the adult heart, NRG-1 expression
appears to be restricted to endothelial cells near cardiomyocytes
(in the endocardium and in the myocardial microvasculature), while
it is absent from larger coronary arteries, veins and the aorta
(9). An increasing number of
studies have focused on NRG-1 and members of the ErbB family that
serve as receptors for NRG-1, in order to better understand the
role of this signaling pathway in the physiology and
pathophysiology of the heart. Based on studies of isolated cell
systems, a number of processes appear to be regulated by NRG-1/ErbB
signaling, including cell growth, myofilament structure and
organization, survival, myocyte-matrix coupling, glucose uptake and
angiogenesis (10–12).
Studies using recombinant human neuregulin-1
(rhNRG-1) containing the epidermal growth factor (EGF)-like domain
(necessary for ErbB2/ErbB4 activation) have shown that NRG-1 plays
an important role in heart performance (13,14).
Therefore, we hypothesized that the gene transfer of NRG-1 would
attenuate ventricular remodeling and improve cardiac function
through regulating cardiac apoptosis and fibrosis. To test this
hypothesis, we investigated the pathophysiological role of NRG-1 in
a rat model of DCM induced by streptozotocin (STZ).
Materials and methods
Animals
Male Sprague Dawley (SD) rats were obtained from the
Animal Center of Nanchang University (Nanchang, China). The
experiments were performed in compliance with the ARRIVE Guidelines
on Animal Research (15). All the
procedures were approved by the Institutional Animal Care and Use
Committee of Nanchang University.
Preparation of rat cardiac microvascular
endothelial cells (CMECs)
Neonatal SD rats (70–80 g) were anesthetized and the
hearts were removed and retrogradely perfused with Dulbecco’s
modified Eagle’s medium (DMEM; Gibco, Carlsbad, CA, USA) for 5 min
through the ascending aorta to remove blood cells. Following the
removal of connective tissue, the remaining left ventricles were
separately finely minced and digested with 0.2% collagenase I
(Sigma, St. Louis, MO, USA) in Hank’s balanced salt solution for 30
min at 37°C in a shaking water bath. Trypsin (0.02%; Gibco) was
then added and the mixture was incubated for an additional 5 min.
The digested solution was filtered through an 100-μm mesh
filter. The filtrate was collected and the cells were plated on
culture dishes coated with human fibronectin (Invitrogen, Carlsbad,
CA, USA) and maintained in DMEM supplemented with 20% fetal bovine
serum (FBS; HyClone, Logan, UT, USA), human vascular endothelial
growth factor (Invitrogen), human fibro-blast growth factor
(Invitrogen) and human EGF (Invitrogen). After a 3-day culture,
unattached cells were removed and fresh medium was added to the
adherent cells. The medium was replaced once every 3 days. The
cells were cultured to 80% confluence before being released with
EDTA (Sigma) and subcultured. Some of the adherent cells at passage
3 were collected for immunofluorescence analyses, which was
performed by staining using factor VIII (Solarbio, Beijing,
China).
Lentiviral construction and gene
transfer
A recombinant lentivirus containing human NRG-1
(pLV-hNRG-1) was constructed. Briefly, a full-length hNRG-1 gene
cDNA was cloned into the lentivirus shuttle plasmid vector pGC-FU,
which contains a cytomegalovirus promoter and a polyadenylation
signal of bovine growth hormone. For the construction of lentivirus
containing green fluorescent protein (GFP), a shuttle vector
containing human phosphoglycerate kinase gene promoter was used.
The control virus lacking the hNGR-1 gene was separately prepared.
Recombinant lentivirus was generated by homologous recombination
and propagated in 293T cells (Genechem, Shanghai, China). At 48 h
after transduction, the supernatant from the 293T cells was
collected and purified by cesium chloride density gradient
centrifugation and stored in 10 mmol/l Tris-HCl (pH 7.4), 1 mmol/l
MgCl2, and 10% (v/v) glycerol at −70°C. Virus titers
were determined by a plaque assay on 293T cell monolayers.
For transduction, CMECs at passage 3 were plated at
a density of 1×105/well into 6-well dishes with DMEM/20%
FBS. The cells at each well were incubated with 20 MOI NRG-1
lentivirus (NRG-1-CMEC) or 20 MOI GFP lentivirus (GFP-CMEC) for 72
h. Lentiviral infection was validated by visualization of enhanced
GFP under a fluorescence microscope (Nikon, Tokyo, Japan).
Subsequently, the medium was replaced with fresh DMEM/20% FBS and
the cells were cultured for an additional 48 h. At the end of the
incubation period, the media and cells in each well were collected
and analyzed.
ELISA for cytokines in CMEC media
The levels of hNRG-1 in the media were analyzed by
enzyme immunoassay using a human neuregulin-1 ELISA kit according
to the manufacturer’s instructions (PlantSelect Biotechnology
Systems Ltd., Dartmouth, NS, Canada). Data were expressed as the
mean ± SEM.
Cardiomyocyte culture
Rat hearts were surgically removed from 1- to
3-day-old SD rats, washed instantly with phosphate-buffered saline
(PBS) solution, and then minced into 1- to 3-mm3 pieces.
The minced tissue was subjected to 6–8 cycles of proteolytic
dissociation by magnetic stirring (10 min, 37°C) in 0.06% trypsin
solution. The supernatants from each cycle were pooled and
centrifuged. The cell pellet was resuspended in DMEM supplemented
with 20% FBS. Selective adhesion was achieved by incubation at 37°C
for 1.5 h in a humidified atmosphere (5% CO2 and 95%
air) in order to obtain a high purity of cardiomyocytes.
Subsequently, 0.1 mM bromodeoxyuridine (Sigma) was added to the
medium for the first 48 h of culture to inhibit the growth of
fibroblasts.
Assessment of the bioactivity of
conditioned CMEC media
Cardiomyocytes were plated into 12-well dishes at a
cell density of 2×104/well and randomly allocated into
three groups. The cells in group A were cultured in fresh DMEM/20%
FBS (control); the cells in group B were cultured in 50% fresh
DMEM/20% FBS and 50% the medium previously harvested from
GFP-CMECs; and the cells in group C were cultured in 50% fresh
DMEM/20% FBS and 50% the medium previously harvested from
hNRG-1-CMECs. After 3 days of culture, some wells of cardiomyocytes
in each group were counted. The number of cardiomyocytes was
independently determined by investigators blinded to the type of
cell culture using a hemacytometer.
The remaining wells of cardiomyocytes of the three
groups were incubated with 20 ng/ml tumor necrosis factor-α (TNF-α)
(Peprotech, Rocky Hill, NJ, USA). The cardiomyocytes were harvested
24 h later, stained with Annexin V-APC/propidium iodide (PI; KeyGen
Biotech, Nanjing, China) and subjected to flow cytometric analysis
to assess apoptosis following the manufacturer’s instructions.
Animals and lentivirus injection
SD rats at a postnatal age of 6 weeks (body weight,
200–220 g) were allocated to the control (n=8) and diabetic groups
(n=30). Diabetes was induced by the intraperitoneal (i.p.)
injection of STZ (50 mg/kg; Sigma, L’Isle d’Abeaux, France)
(16). Tail vein blood glucose was
measured every 3 days during the first week; the rats with plasma
glucose levels ≥16.7 mmol/l were considered to be diabetic.
Concurrently, control rats were injected i.p. with 1 ml/kg body
weight 20 mmol/l citrate buffer (pH 4.5) vehicle. The control and
diabetic rats both raised on standard food and water for the whole
experimental period. Twelve weeks after the induction of diabetes,
24 diabetic rats were used for further analysis; the remaining six
rats died or were excluded due to unsuccessful induction of
diabetes. The 24 diabetic rats were randomly allocated into three
groups: the hNRG-1, GFP and DCM groups (n=8 rats per group).
Subsequently, all the diabetic rats were anesthetized with 4%
chloral hydrate solution (1 ml/100 g) by i.p. injection and put on
an animal ventilator. Thoracotomy was performed. Approximately 50
μl/heart (5×107 TU/ml) of hNRG-1-lentivirus
(hNRG-1 group) or GFP-lentivirus (GFP group) or an equivalent
volume of PBS alone (DCM group) was injected at five sites in the
left ventricles of the rats using a 30-gauge needle. Following the
injection of lentiviral vectors, the rats continued to be raised on
standard food and water for 4 weeks. All analyses were performed 16
weeks after the induction of diabetes.
Analysis of myocardial function
To evaluate the cardiac function, cardiac
catheterization was performed as previously described (17). Briefly, after the induction of
light general anesthesia [4% chloral hydrate solution (1 ml/100 g)
by i.p. injection], a catheter was inserted into the right carotid
artery and advanced into the left ventricle. Ventricular pressure
signals were measured with a transducer and conditioner (MLT0830;
ADInstruments, Bella Vista, Australia) and digitally recorded with
a data acquisition system (PowerLab; ADInstruments). The following
indices were obtained: heart rate (HR), left ventricular systolic
pressure (LVSP), left ventricular enddiastolic pressure (LVEDP), as
well as the maximum rates of left ventricular pressure rise and
fall (+dp/dt max and −dp/dt max, respectively). During this
process, the animals were placed on controlled heating pads. Core
temperature was measured via a rectal probe and was maintained at
37°C. The rats were sacrificed after analysis of myocardial
function and the hearts were harvested for subsequent
experiments.
Histopathological process and detection
of apoptotic cells by terminal deoxynucleotidyl
transferase-mediated dUTP nick-end labeling (TUNEL) staining
The samples were fixed in 10% formalin and were
paraffin-embedded in the Surgical Pathology Facility of Nanchang
University. TUNEL analysis was performed with a commercially
available kit (Dead End Colorimetric TUNEL System) according to the
manufacturer’s instructions (Promega, Madison, WI, USA). The slides
were counterstained with hematoxylin (blue). Three midventricular
sections (from the apex to the base) of each heart tissue were
analyzed. Cardiomyocyte nuclei were quantified by randomly counting
10 fields/section. The apoptotic index (percentage of apoptotic
nuclei) was calculated as apoptotic nuclei/total nuclei counted ×
100.
Analysis of myocardial collagen
content
Sections of left ventricles were stained with
Masson’s trichrome to measure interstitial fibrosis. Interstitial
collagen was quantified at a final magnification of ×200 using a
microscope (BX51; Olympus, Center Valley, PA, USA) connected to a
video camera (DS-Fi1; Nikon, Tokyo, Japan). The images captured
under the microscope were used to calculate the collagen volume
fraction of the myocardial interstitium using a Computer Imaging
Analysis System (MPIAS-500; Nikon, Tokyo, Japan). The content of
interstitial collagen, which was expressed as the fractional area
of the entire cross-section where the perivascular collagen was
excluded, was averaged on nine fields selected across the wall
thickness in the septum and free wall.
Gene expression analysis by quantitative
reverse transcription polymerase chain reaction (qPCR)
Tissue samples obtained from the left ventricular
free wall were minced. Total RNA was extracted from the samples
with TRIzol reagent according to the manufacture’s instructions
(Invitrogen). For qPCR, cDNA was synthesized in a 20-μl
reaction volume containing 4 μg total RNA and SuperScript™
II Reverse Transcriptase (Fermentas, Burlington, ON, Canada)
according to the manufacturer’s instructions. qPCR was carried out
with a 7500 Real-Time PCR system (Applied Biosystems, Carlsbad, CA,
USA) using SYBR-Green I (Applied Biosystems) as a fluorescent dye
according to the manufacturer’s instructions. Relative quantitation
of the mRNA expression of the gene of interest was calculated using
the comparative threshold cycle number for each sample. To control
the variation in the amount of DNA, gene expression of the target
sequence was normalized in relation to the expression of an
internal control, β-actin. The PCR products of hNRG-1 were
size-fractioned by electrophoresis on 2% agarose gels. Primers for
hNRG-1, bcl-2, bax, collagen type I and III, as well as β-actin
were the following: hNRG-1, forward:
5′-TCACCATGGTGGCGACCGGTTCAGGCAGAGACAGAAAG-3′ and reverse:
5′-TCACCATGGTGGCGACCGGTTCAGGCAGAGACAGAAAG-3′; bcl-2, forward:
5′-CGGGAGATCGTGATGAAGT-3′ and reverse: 5′-CCACCGAACTCAAAGAAGG-3′;
bax, forward: 5′-GCAGGGAGGATGGCTGGGGAGA-3′ and reverse:
5′-TCCAGACAAGCAGCCGCTCACG-3′; collagen type I, forward:
5′-GTTCGTGGTTCTCAGGGTAG-3′ and reverse: 5′-TTGTCGTAGCAGGGTTCTTT-3′;
collagen type III, forward: 5′-TGCCCACAGCCTTCTACACCCT-3′ and
reverse: 5′-CAGCCATTCCTCCCACTCCAG-3′; β-actin, forward:
5′-TGTGCTATGTTGCCCTAGACTTC-3′ and reverse:
5′-CGGACTCATCGTACTCCTGCT-3′.
Western blot analysis
Samples of left ventricle myocardium were
homogenized in tissue protein extraction reagent (Beyotime,
Beijing, China) supplemented with protease inhibitors. After
centrifugation at 12,000 × g for 10 min at 4°C, the supernatants
were collected according to the manufacturer’s instructions.
Protein concentration was measured using the BCA Protein Assay kit
(Pierce Biotechnology, Rockford, IL, USA). Equal amounts of 20
μg protein were loaded onto 10% sodium dodecyl
sulfate-polyacrylamide gels. hNRG-1, total Akt, phospho-Akt (p-Akt;
phospho-Ser 473), total eNOS and phospho-eNOS (p-eNOS; phospho-Ser
1177) were detected with mouse anti-hNRG-1, rabbit
anti-Akt-1/-2/-3, rabbit anti-phospho-Akt-1/-2/-3 (Ser 473), mouse
anti-eNOS and rabbit anti-phospho-eNOS antibodies (Santa Cruz
Biotechnology, Inc., Santa Cruz, CA, USA). After probing with these
antibodies, the membranes were stripped of bound immunoglobulins.
The immunoblots were developed by the enhanced
chemiluminofluorescence method (Thermo Fisher Scientific, Hudson,
NH, USA). The signals were quantified by densitometric analysis
using a chemiluminescence imaging system (General Electric Company,
Fairfield, CT, USA) and normalized to those of β-actin, an
endogenous control protein.
Statistical analysis
All the data were expressed as the mean ± SEM.
Comparisons between groups were made using one-way analysis of
variance (ANOVA) with Fisher’s protected least significant
difference post hoc comparison test. P<0.05 was considered to
indicate a statistically significant difference.
Results
Effects of CMEC supernatants on
cardiomyocyte proliferation and survival
CMECs were characterized by positive staining for
factor VIII endothelial marker. The cells at passage 3 were
transfected with hNGR-1-lentivirus or GFP-lentivirus and the
expression of GFP was observed by fluorescence analysis.
Cytokines in the supernatant were measured by ELISA.
The level of hNRG-1 in the group transfected with hNGR-1-lentivirus
was detected to be 18±5.3 ng/ml, while no hNRG-1 was detected in
the group transfected with GFP-lentivirus.
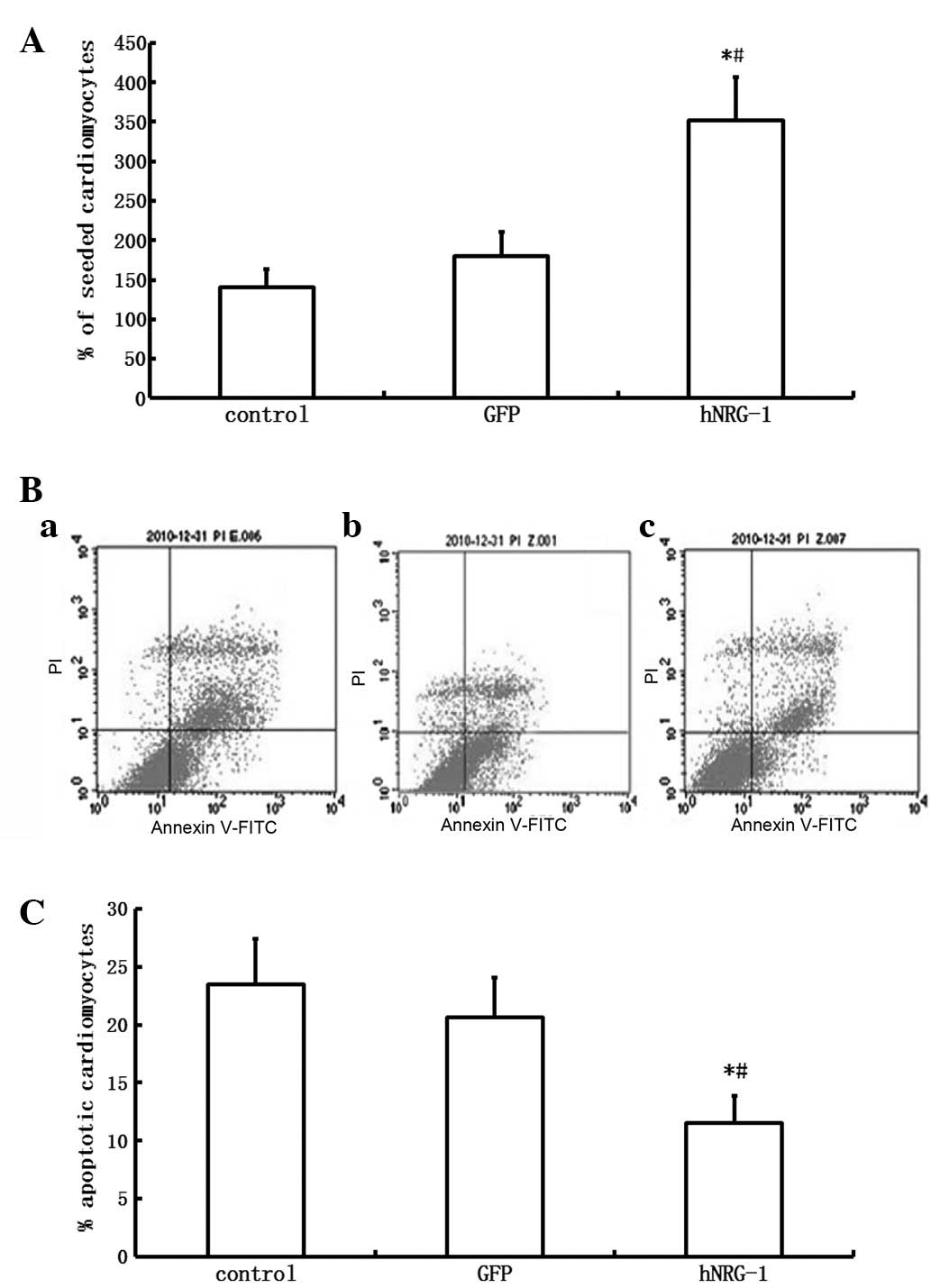
Since the proliferation and survival of
cardiomyocytes is an important aspect of DCM, the effects of
CMEC-conditioned media on cardiomyocyte growth and survival were
determined. The number of cardiomyocytes cultured for 3 days in
hNRG-1-CMEC conditioned basal medium was significantly higher
compared with the number of cardiomyocytes cultured in GFP-CMEC
conditioned basal medium (P<0.05). No significant difference was
detected in the number of cardiomyocytes between the control and
GFP groups (P>0.05, Fig.
1).
After co-culture with TNF-α for 24 h, the apoptotic
rates of cardiomyocytes cultured in hNRG-1-CMEC conditioned basal
medium was significantly lower compared with the number of
cardiomyocytes cultured in GFP-CMEC conditioned basal medium
(P<0.05). No significant difference was detected in the
apoptotic rates of cardiomyocytes between the control and GFP
groups (P>0.05, Fig. 1).
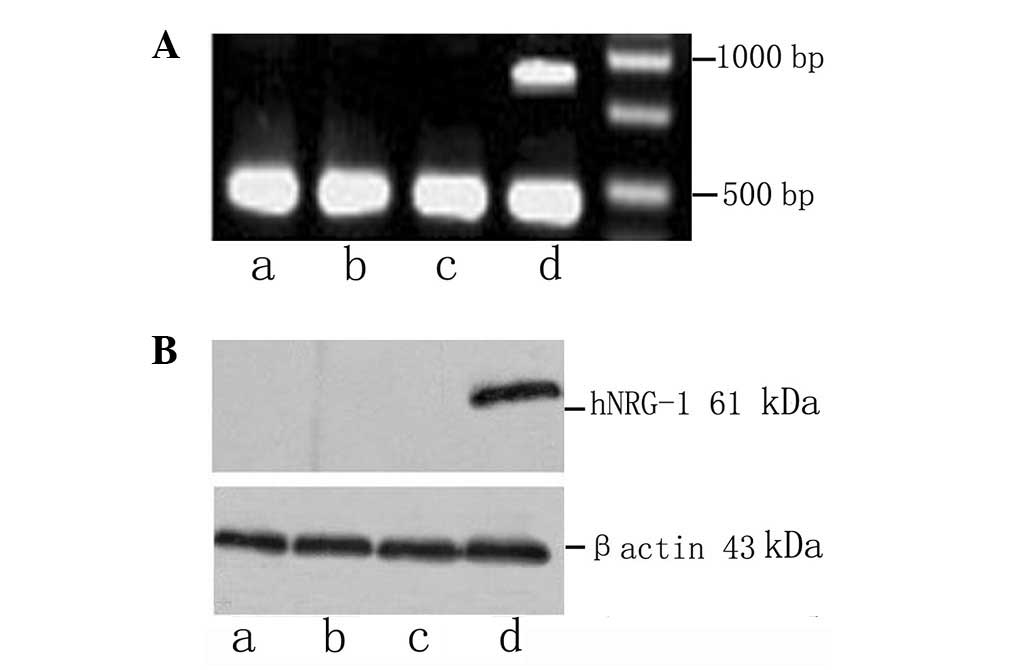
hNRG-1 expression in cardiac tissues
The expression of hNRG-1 in heart tissues was
confirmed by relative quantification of hNRG-1 mRNA and western
blot analysis. hNRG-1 mRNA and the fusion proteins of hNRG-1 and
GFP were detected in the cardiac samples extracted from the cardiac
tissues of the rats in the hNRG-1 group but not in those from the
rats in the control, DCM and GFP groups (Fig. 2).
NRG-1 improves myocardial function
Sixteen weeks after the induction of diabetes,
cardiac function was evaluated by invasive hemodynamic
measurements. No significant difference was detected in HR among
the four groups (P>0.05). A lower LVSP and higher LVEDP were
observed in the DCM group compared with those in the control group
(P<0.05). Resting maximum rates of rise (+dp/dt max) and fall
(−dp/dt max) in left ventricular pressure were also impaired after
the induction of diabetes (P<0.05), indicating that systolic and
diastolic functions were significantly impaired in the diabetic
rats. Following the injection of hNRG-1-lentivirus, these
hemodynamic abnormalities were markedly attenuated (P<0.05).
However, the hemodynamic abnormalities in the GFP group were
comparable with those in the DCM group (P>0.05, Table I).
 | Table I.Hemodynamic parameters evaluated by
invasive measurements. |
Table I.
Hemodynamic parameters evaluated by
invasive measurements.
| Group | HR (beats/min) | LVSP (mmHg) | LVEDP (mmHg) | +dp/dt
(mmHg/sec) | −dp/dt
(mmHg/sec) |
|---|
| Control (n=8) | 340±35 | 135±15 | 2.3±0.9 | 6451±408 | 5819±328 |
| DCM (n=8) | 320±31 | 103±7a | 11.8±3.2a | 4901±341a | 3856±275a |
| GFP (n=8) | 318±21 | 108±11a | 10.1±2.7a | 4887±322a | 3845±259a |
| hNRG-1 (n=8) | 329±29 | 120±12b,c |
6.4±2.3a–c |
5908±361a–c |
4890±311a–c |
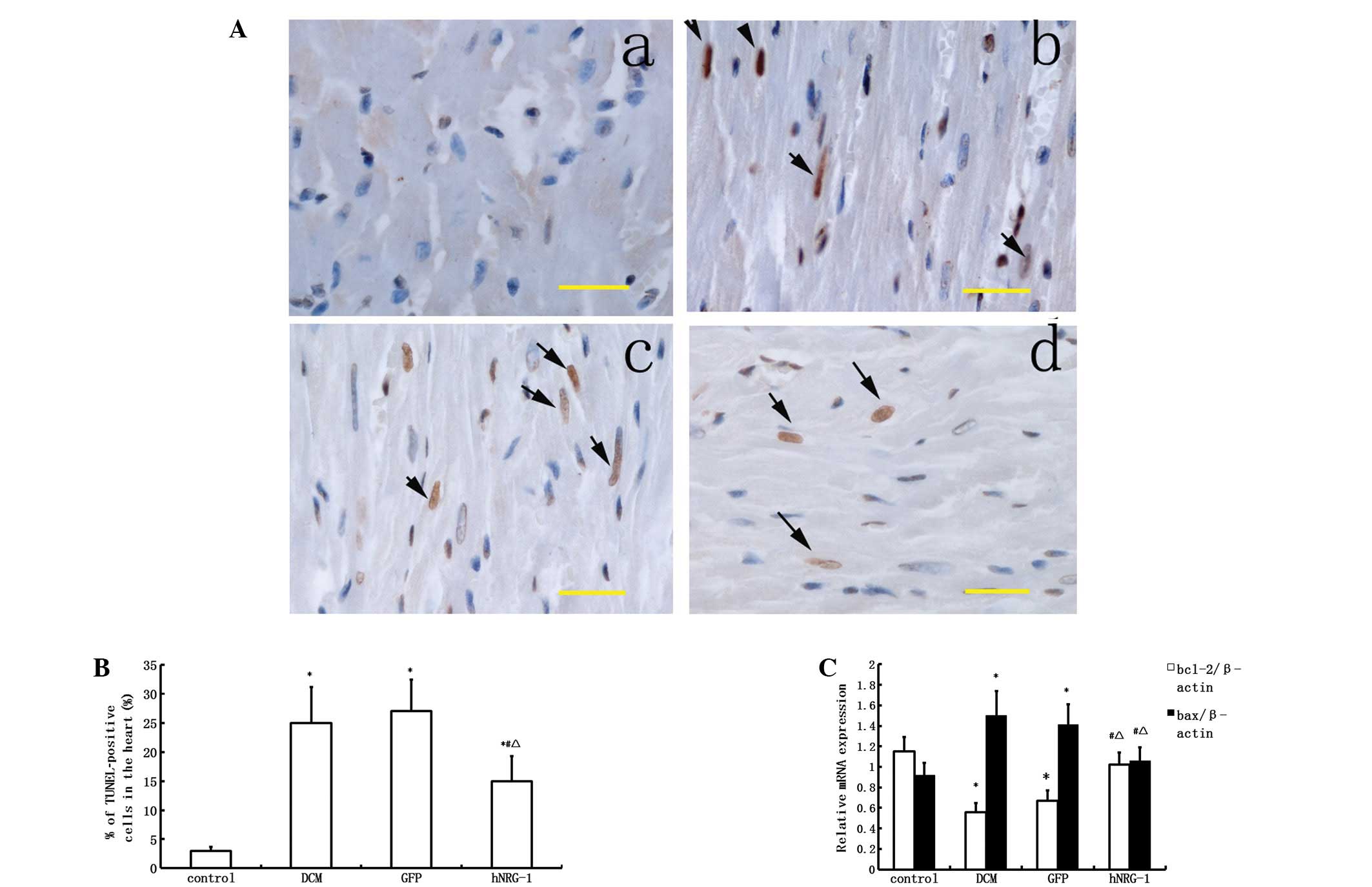
NRG-1 protects cardiomyocytes against
apoptosis
A TUNEL assay was performed to assess apoptosis
in vivo. The number of positively stained cells in the DCM
group was higher than that in the control group (P<0.05).
Transfection with hNRG-1-lentivirus significantly reduced the
number of apoptotic cells compared with those in the GFP and DCM
groups (P<0.05, Fig. 3). qPCR
was used to determine the mRNA expression levels of bcl-2 and bax,
which are known to be markers of apoptosis. Following qPCR, bcl-2
was shown to be downregulated and bax was upregulated in the DCM
group, while these alterations were attenuated following
transfection with hNRG-1-lentivirus (P<0.05). Since bcl-2 is
known to be an anti-apoptotic and bax a pro-apoptotic protein,
these results indicated that transfection with hNRG-1-lentivirus
protects cardiomyocytes against apoptosis (Fig. 3).
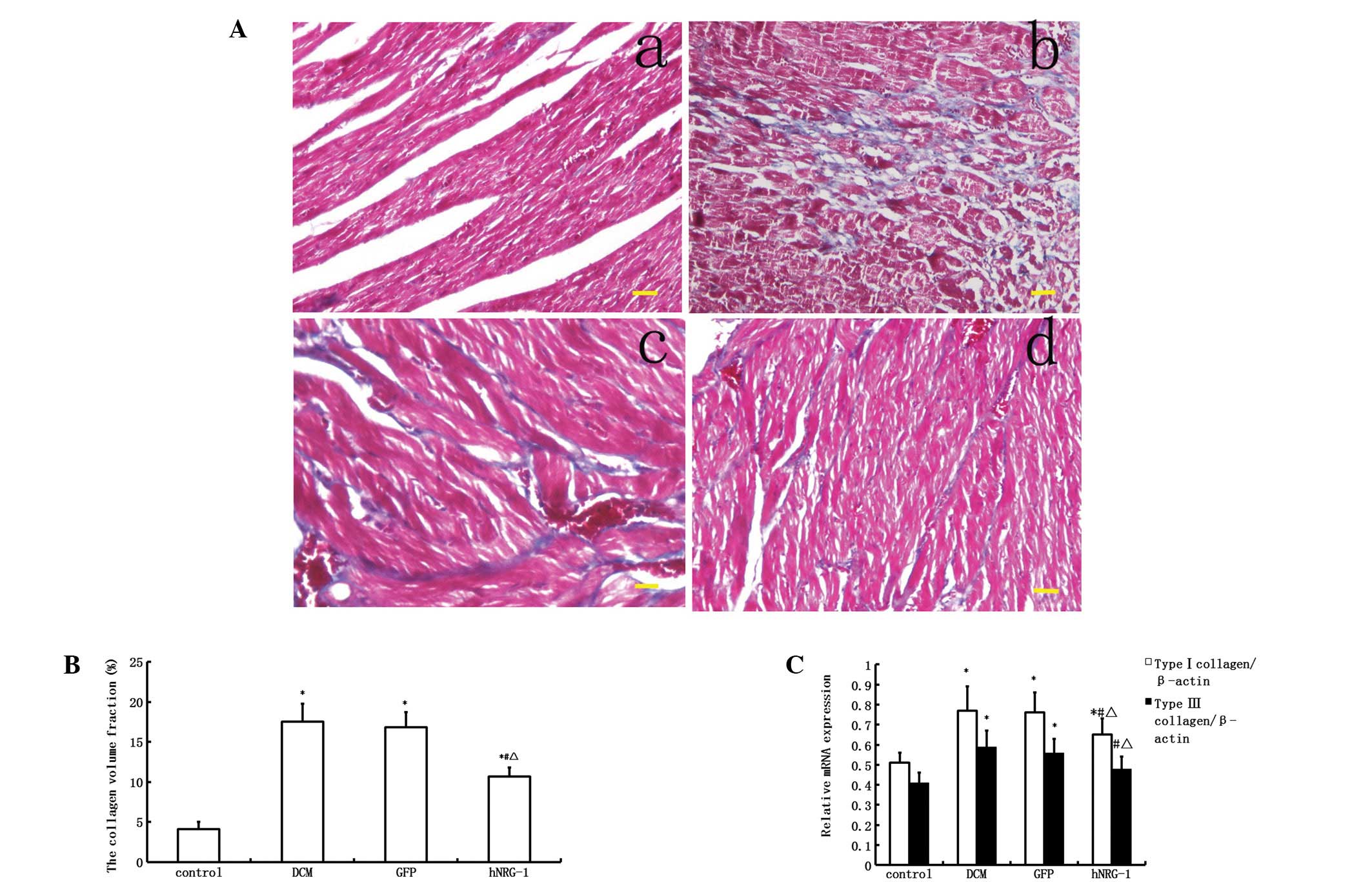
NRG-1 attenuates myocardial interstitial
fibrosis
The collagen volume fraction, which is an indicator
of interstitial fibrosis, was higher in the DCM group than in the
control group (P<0.05, Fig. 4).
Similarly, the collagen type I and III mRNA expression levels were
also significantly upregulated in the DCM group compared with those
in the control group (P<0.05, Fig.
4). The levels of myocardial fibrosis and of type I and III
pro-collagen mRNA in the myocardium were markedly inhibited
following cardiac transfection with rhNRG-1 rather than GFP,
suggesting that hNRG-1 treatment attenuates the myocardial
interstitial fibrosis caused by diabetes.
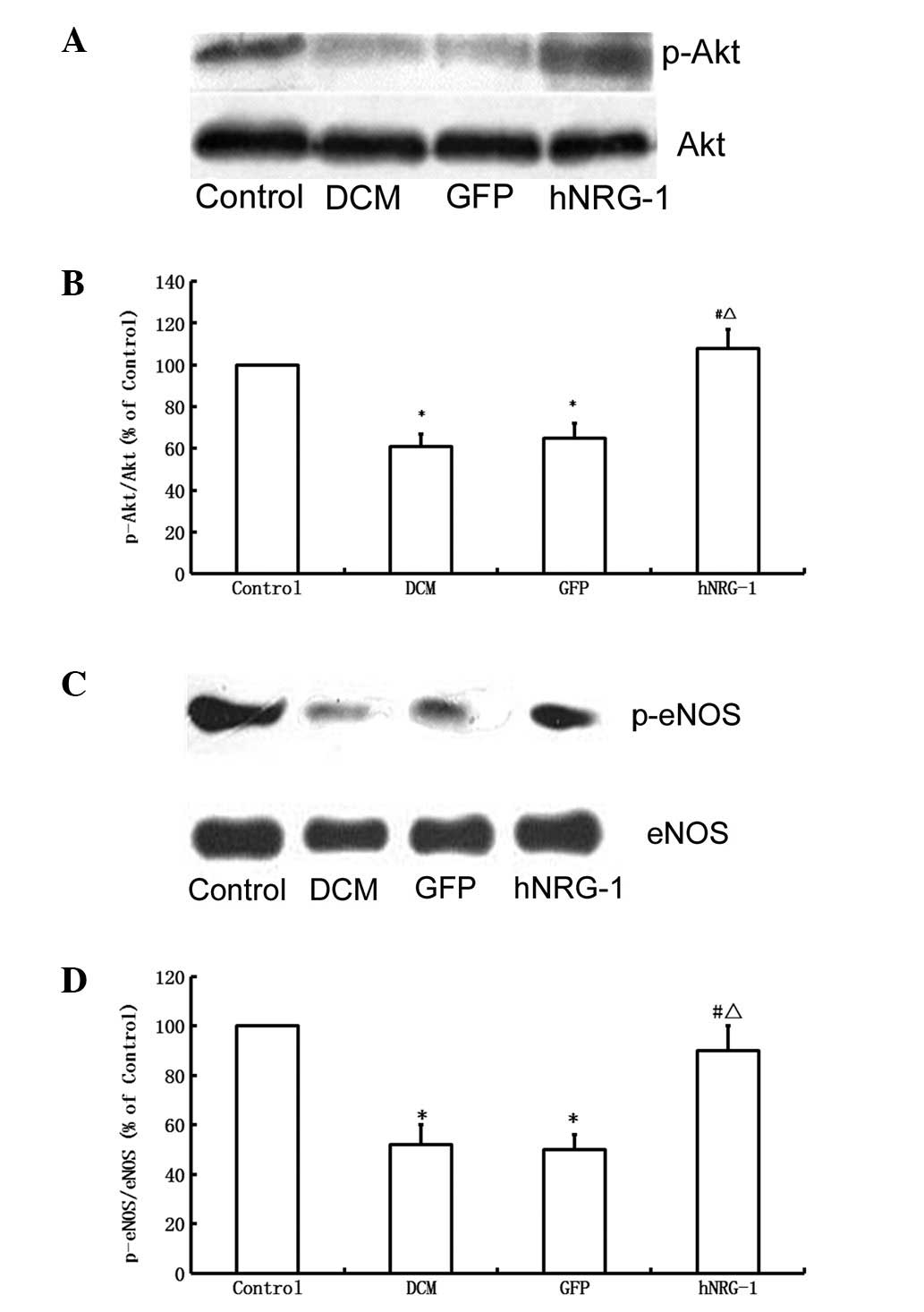
NRG-1 increases the expression of
phospho-Akt and phospho-eNOS
Western blot analysis showed that STZ treatment
reduced the level of Akt phosphorylation compared with that in the
control group, while hNRG-1 gene transfer increased the level of
phospho-Akt (Fig. 5). Total Akt
levels were not altered among the four groups. Similarly, hNRG-1
gene delivery significantly increased the level of the
phosphorylated form of eNOS compared with the levels in the GFP and
control groups. Total eNOS levels remained unaltered (Fig. 5). These results indicate that
hNRG-1 gene transfer resulted in the activation of Akt and eNOS
pathways by phosphorylation.
Discussion
The present study demonstrated that gene transfer of
hNRG-1 attenuated the remodeling of the hearts of DCM model rats by
regulating cardiomyocyte apoptosis and cardiac fibrosis, in
association with enhancement of systolic and diastolic cardiac
function. These findings support the hypothesis that NRG-1 plays an
important role in the regulation of heart function.
NRG-1 acts as a paracrine factor via the ErbB family
of tyrosine kinase receptors expressed in cardiomyocytes. ErbB
receptors are a family of four transmembrane receptors that bind
multiple growth factors including epidermal growth factor (EGF),
transforming growth factor-α (TGF-α) and NRG1-4. ErbB3 is expressed
in prenatal myocytes, while adult ventricular myocytes express only
ErbB1, ErbB2 and ErbB4. ErbB1, also known as the EGF receptor, does
not bind NRG-1; thus, only ErbB2 and ErbB4 serve as NRG-1 receptors
in adult cardiomyocytes (18,19).
NRG-1 acts through ErbB2 and ErbB4 in a paracrine fashion to
stimulate MEK/ERK, Akt/PI3-kinase, Src/FAK and NO synthase, which
act synergistically to promote myocyte function and survival in the
setting of cardiac stress (20–23).
Increased apoptosis has been shown to play a
critical role in the development of DCM (24,25).
In the present study, the overexpression of hNRG-1 inhibited
cardiomyocyte apoptosis in vivo and in vitro. The
mechanisms of apoptosis inhibition are suggested to be associated
with PI3-kinase/Akt pathway, one of the downstream signaling
pathways of NRG-1/ErbB. Previous studies have shown that the
PI3-kinase/Akt pathway is involved in the protection of
cardiomyocytes against cell death, as well as in the regulation of
metabolism and growth (26,27).
The exact mechanism by which NRG-1-dependent Akt signaling protects
myocytes has not been fully elucidated. An Akt-dependent change in
bcl-2 family expression has been implicated (28,29).
It has been reported that NRG-1 is involved in the regulation of
bcl-2 and p-Bad expression through the PI3K/Akt pathway following
transient focal cerebral ischemia (30). The results of the present study
suggest that the role of NRG-1 in cardiac protection is conferred
through activation of the Akt pathway, which is associated with an
increase in the level of bcl-2 expression and reduction in the
level of bax expression. The ratio of bax/bcl-2 is known to be an
important marker of cardiomyocyte apoptosis (31,32).
DCM is considered to involve interstitial and
perivascular fibrosis; fibrosis is one of the most important
characteristics of DCM (25).
Myocardial fibrosis is known to cause myocardial dysfunction in
diabetes. Our findings demonstrated that NGR-1 attenuates heart
fibrosis. However, the exact underlying mechanism remains unclear.
NO synthase, which may be stimulated by the NRG-1/ErbB signaling
pathway, is suggested to be involved in myocardial fibrosis. NO
synthase has been reported to attenuate myocardial fibrosis by
regulating renin release (33).
Upregulation of the renin-angiotensin system (RAS) has been
described in diabetes and is associated with the development of
cardiac hypertrophy and fibrosis. In addition, cardiomyocytes and
endothelial cells in the hearts of individuals with diabetes and
end-stage heart failure provide evidence of oxidative stress,
apoptosis and necrosis that correlate with RAS activation (2). In the present study, certain actions
of NRG-1 relating to the inhibition of cardiac remodeling are
suggested to depend on the regulation of renin release through NO
synthase.
Other mechanisms may be involved in the cardiac
protective effect of NRG-1. Evidence indicates that increased
oxidative stress contributes to the development and progression of
DCM. The activation of RAS has been shown to be associated with
increased oxidative damage and cardiomyocyte apoptosis in the
diabetic heart, leading to cardiac fibrosis (34–36).
Previous studies have shown that NRG-1β treatment of myocytes in
vitro alters the expression of a number of genes related to
regulation of cellular oxidative stress, such as catalase and
superoxide dismutase (SOD), which are significantly upregulated
following treatment with NRG-1β, supporting the idea that NRG-1β is
important in the regulation of myocardial oxidative stress
(12). Based on these findings,
NRG-1 is postulated to inhibit cardiac remodeling at least
partially through the regulation of myocardial oxidative stress.
Cardiac autonomic neuropathy is an additional important mechanism
of DCM. Previous studies have shown that neuregulin proteins have
the ability to control excessive β-adrenergic activation. NRG-1 may
induce counterbalancing parasympathetic activity in animal models,
which is potentially an additional factor in the protective role of
neuregulin in heart failure (37,38).
It has been reported that NRG-1/ErbB expression
declined at a later stage of pump failure (39). The decline in NRG-1 expression
coincided with the development of eccentric ventricular hypertrophy
and pump failure, and was accompanied by a downregulation of the
ErbB2 and ErbB4 mRNA levels. This was suggested to be due to the
increased levels of angiotensin II and epinephrine, both of which
reduce NRG-1 mRNA synthesis in the cardiac endothelium (40). The replenishment of NRG-1 is
suggested to inhibit the physiopathological aggravation of DCM.
In conclusion, the gene transfer of hNRG-1 improves
cardiac dysfunction in diabetes. Although further studies are
needed, NRG-1 appears to be able to protect cardiomyocytes against
apoptosis and to reduce the extent of myocardial interstitial
fibrosis. Replenishing NRG-1 is suggesting to be an alternative
option for DCM treatment, although the exact mechanism of action
required investigation in future studies.
Acknowledgements
This study was supported by the
National Natural Science Foundation of China (30860101, 81041097).
The authors thank Dr Hong Xu and Dr Junyi Zeng for the expert
technical assistance and critical comments on the manuscript.
References
|
1.
|
Acar E, Ural D, Bildirici U, et al:
Diabetic cardiomyopathy. Anadolu Kardiyol Derg. 11:732–737.
2011.
|
|
2.
|
Battiprolu PK, Gillette TG, Wang ZV, et
al: Diabetic cardiomyopathy: mechanisms and therapeutic targets.
Drug Discov Today Dis Mech. 7:e135–e143. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
3.
|
An D and Rodrigues B: Role of changes in
cardiac metabolism in development of diabetic cardiomyopathy. Am J
Physiol Heart Circ Physiol. 291:H1489–H1506. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
4.
|
Boudina S and Abel ED: Diabetic
cardiomyopathy revisited. Circulation. 115:3213–3223. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
5.
|
Hsueh W, Abel ED, Breslow JL, et al:
Recipes for creating animal models of diabetic cardiovascular
disease. Circ Res. 100:1415–1427. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
6.
|
Sun D, Shen M, Li J, et al:
Cardioprotective effects of tanshinone IIA pretreatment via kinin
B2 receptor-Akt-GSK-3β dependent pathway in experimental diabetic
cardiomyopathy. Cardiovasc Diabetol. 10:42011.PubMed/NCBI
|
|
7.
|
Xu Y, Li X, Liu X and Zhou M:
Neuregulin-1/ErbB signaling and chronic heart failure. Adv
Pharmacol. 59:31–51. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8.
|
Odiete O, Hill MF and Sawyer DB:
Neuregulin in cardiovascular development and disease. Circ Res.
111:1376–1385. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9.
|
Lemmens K, Segers VF, Demolder M and De
Keulenaer GW: Role of neuregulin-1/ErbB2 signaling in
endothelium-cardiomyocyte cross-talk. J Biol Chem. 281:19469–19477.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
10.
|
Cote GM, Miller TA, Lebrasseur NK, et al:
Neuregulin-1alpha and beta isoform expression in cardiac
microvascular endothelial cells and function in cardiac myocytes in
vitro. Exp Cell Res. 311:135–146. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
11.
|
Pentassuglia L, Timolati F, Seifriz F, et
al: Inhibition of ErbB2/neuregulin signaling augments
paclitaxel-induced cardiotoxicity in adult ventricular myocytes.
Exp Cell Res. 313:1588–1601. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12.
|
Pentassuglia L and Sawyer DB: The role of
neuregulin 1β/ErbB signaling in the heart. Exp Cell Res.
315:627–637. 2009.
|
|
13.
|
Jabbour A, Hayward CS, Keogh AM, et al:
Parenteral administration of recombinant human neuregulin-1 to
patients with stable chronic heart failure produces favourable
acute and chronic haemodynamic responses. Eur J Heart Fail.
13:83–92. 2011. View Article : Google Scholar
|
|
14.
|
Li B, Zheng Z, Wei Y, et al: Therapeutic
effects of neuregulin-1 in diabetic cardiomyopathy rats. Cardiovasc
Diabetol. 10:692011.PubMed/NCBI
|
|
15.
|
Kilkenny C, Browne WJ, Cuthill IC, et al:
Improving bioscience research reporting: the ARRIVE guidelines for
reporting animal research. PLoS Biol. 8:e10004122010. View Article : Google Scholar : PubMed/NCBI
|
|
16.
|
Wold LE and Ren J: Streptozotocin directly
impairs cardiac contractile function in isolated ventricular
myocytes via a p38 map kinase-dependent oxidative stress mechanism.
Biochem Biophys Res Commun. 318:1066–1071. 2004. View Article : Google Scholar
|
|
17.
|
Wichi R, Malfitano C, Rosa K, et al:
Noninvasive and invasive evaluation of cardiac dysfunction in
experimental diabetes in rodents. Cardiovasc Diabetol. 6:142007.
View Article : Google Scholar : PubMed/NCBI
|
|
18.
|
Jiang Z and Zhou M: Neuregulin signaling
and heart failure. Curr Heart Fail Rep. 7:42–47. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19.
|
Dammann O, Bueter W, Leviton A, et al:
Neuregulin-1: a potential endogenous protector in perinatal brain
white matter damage. Neonatology. 93:182–187. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20.
|
Kuramochi Y, Guo X and Sawyer DB:
Neuregulin activates erbB2-dependent src/FAK signaling and
cytoskeletal remodeling in isolated adult rat cardiac myocytes. J
Mol Cell Cardiol. 41:228–235. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
21.
|
Hertig CM, Kubalak SW, Wang Y and Chien
KR: Synergistic roles of neuregulin-1 and insulin-like growth
factor-I in activation of the phosphatidylinositol 3-kinase pathway
and cardiac chamber morphogenesis. J Biol Chem. 274:37362–37369.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
22.
|
Calvo M, Zhu N, Grist J, et al: Following
nerve injury neuregulin-1 drives microglial proliferation and
neuropathic pain via the MEK/ERK pathway. Glia. 59:554–568. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
23.
|
Ky B, Kimmel SE, Safa RN, et al:
Neuregulin-1 beta is associated with disease severity and adverse
outcomes in chronic heart failure. Circulation. 120:310–317. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
24.
|
Cai L, Li W, Wang G, et al:
Hyperglycemia-induced apoptosis in mouse myocardium: mitochondrial
cytochrome c-mediated caspase-3 activation pathway.
Diabetes. 51:1938–1948. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
25.
|
Mano Y, Anzai T, Kaneko H, et al:
Overexpression of human C-reactive protein exacerbates left
ventricular remodeling in diabetic cardiomyopathy. Circ J.
75:1717–1727. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26.
|
Wang B, Shravah J, Luo H, et al: Propofol
protects against hydrogen peroxide-induced injury in cardiac H9c2
cells via Akt activation and Bcl-2 up-regulation. Biochem Biophys
Res Commun. 389:105–111. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27.
|
Markou T, Barlaka E, Bartucci M and Lazou
A: Signal transduction pathways through cytoprotective, apoptotic
and hypertrophic stimuli: a comparative study in adult cardiac
myocytes. Cell Biochem Funct. 29:442–451. 2011. View Article : Google Scholar
|
|
28.
|
Jamnicki-Abegg M, Weihrauch D, Pagel PS,
et al: Isoflurane inhibits cardiac myocyte apoptosis during
oxidative and inflammatory stress by activating Akt and enhancing
Bcl-2 expression. Anesthesiology. 103:1006–1014. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
29.
|
Dhanasekaran A, Gruenloh SK, Buonaccorsi
JN, et al: Multiple antiapoptotic targets of the PI3K/Akt survival
pathway are activated by epoxyeicosatrienoic acids to protect
cardiomyocytes from hypoxia/anoxia. Am J Physiol Heart Circ
Physiol. 294:H724–H735. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
30.
|
Guo WP, Fu XG, Jiang SM and Wu JZ:
Neuregulin-1 regulates the expression of Akt, Bcl-2, and Bad
signaling after focal cerebral ischemia in rats. Biochem Cell Biol.
88:649–654. 2010.PubMed/NCBI
|
|
31.
|
Sapia L, Palomeque J, Mattiazzi A and
Petroff MV: Na+/K+-ATPase inhibition by
ouabain induces CaMKII-dependent apoptosis in adult rat cardiac
myocytes. J Mol Cell Cardiol. 49:459–468. 2010.
|
|
32.
|
Rosca AM, Matei C, Dragan E and Burlacu A:
Cardiomyocyte apoptosis in ischaemia-reperfusion due to the
exogenous oxidants at the time of reperfusion. Cell Biol Int.
36:1207–1215. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
33.
|
Chatziantoniou C, Pauti MD, Pinet F, et
al: Regulation of renin release is impaired after nitric oxide
inhibition. Kidney Int. 49:626–633. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
34.
|
Giacco F and Brownlee M: Oxidative stress
and diabetic complications. Circ Res. 107:1058–1070. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
35.
|
Zhou G, Li X, Hein DW, et al:
Metallothionein suppresses angiotensin II-induced nicotinamide
adenine dinucleotide phosphate oxidase activation, nitrosative
stress, apoptosis, and pathological remodeling in the diabetic
heart. J Am Coll Cardiol. 52:655–666. 2008. View Article : Google Scholar
|
|
36.
|
Varagic J, Ahmad S, Voncannon JL, et al:
Nebivolol reduces cardiac angiotensin II, associated oxidative
stress and fibrosis but not arterial pressure in salt-loaded
spontaneously hypertensive rats. J Hypertens. 30:1766–1774.
2012.
|
|
37.
|
Lemmens K, Fransen P, Sys SU, et al:
Neuregulin-1 induces a negative inotropic effect in cardiac muscle:
role of nitric oxide synthase. Circulation. 109:324–326. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
38.
|
Sawyer DB and Caggiano A: Neuregulin-1β
for the treatment of systolic heart failure. J Mol Cell Cardiol.
51:501–505. 2011.
|
|
39.
|
Rohrbach S, Niemann B, Silber RE and Holtz
J: Neuregulin receptors erbB2 and erbB4 in failing human
myocardium-depressed expression and attenuated activation. Basic
Res Cardiol. 100:240–249. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
40.
|
Lemmens K, Doggen K and De Keulenaer GW:
Role of neuregulin-1/ErbB signaling in cardiovascular physiology
and disease: implications for therapy of heart failure.
Circulation. 116:954–960. 2007. View Article : Google Scholar : PubMed/NCBI
|