Introduction
Atrial fibrillation (AF) is one of the most
frequently occurring clinical diseases and is a common clinical
manifestation of rheumatic heart disease (RHD), hypertension,
coronary heart disease, congenital heart disease, cardiomyopathy,
pericardial diseases and other cardiovascular disease. AF presents
a serious threat to the health of the individual. In addition,
mural thrombus, induced by AF, may lead to serious cardiovascular
events with high rates of morbidity and mortality (1). Previous studies have shown that
radiofrequency catheter ablation may be used to narrow the left
atrial diameter (LAD) and significantly improve the left
ventricular ejection fraction (LVEF) and cardiac function in
patients with AF, indicating that the remodeling of atrial
structure is crucial in the development of AF (2–5).
Therefore, blocking the remodeling of the atrial structure maybe an
improved method of preventing the development of AF. Cardiac
fibrosis, which is a common pathology of numerous cardiovascular
diseases, has become of particular interest in recent years. This
is due to a desire to provide a theoretical basis for the
development of novel targets for anti-fibrotic therapies.
Clinical and non-clinical studies have demonstrated
that atrial fibrosis is the most prominent manifestation of atrial
structural remodeling in patients with AF (6–8). The
electrical conductivity heterogeneity in atrial fibrosis
facilitates the occurrence and maintenance of AF (9–12).
One of the primary factors leading to fibrosis is an imbalance
between fibrogenic and antifibrotic cell growth factors. Basic
fibroblast growth factor (bFGF) is a fibrogenic cell growth factor,
while hepatocyte growth factor (HGF) has been identified to be a
unique antifibrotic cell growth factor.
In addition to promoting cell differentiation,
mitosis, tumor occurrence and metastasis, HGF is also involved in
antifibrotic processes and exerts a variety of biological effects
(13–14). Iwata et al (15) demonstrated that a low degree of
myocardial fibrosis was present with lower levels of HGF in the
myocardial tissue of rats and that myocardial collagen expression
and distribution were significantly reduced following HGF
overexpression (15). HGF is the
ligand of c-Met, which has a domain with protein tyrosine kinase
activity (16). c-Met transduces
signals from the extracellular matrix into the cytoplasm by binding
with HGF/HGF ligand to regulate a number of physiological
processes, including cell proliferation, scattering, morphogenesis
and survival. Ligand binding at the extracellular domain induces
the autophosphorylation of c-Met in its intracellular domain, which
provides docking sites for downstream signaling molecules (17). Following activation by its ligand,
c-Met interacts with the phosphoinositide (PI) 3-kinase subunit
phosphoinositide-3-kinase regulatory subunit 1 (PIK3R1),
phospholipase C γ 1 (PLCG1), SRC, growth factor receptor-bound
protein 2 (GRB2), signal transducer and activator of transcription
3 (STAT3) or the adapter GRB2-associated-binding protein 1 (GAB1),
which is necessary for c-Met to activate a number of signaling
cascades, including RAS-extracellular signal-regulated kinase
(ERK), PI3-kinase-AKT and phospholipase C γ-protein kinase C
(PLCγ-PKC). The RAS-ERK activation is involved with morphogenetic
effects, while PI3K/AKT coordinates pro-survival effects (18). During embryonic development, c-Met
signaling is important in gastrulation, development and migration
of muscles and neuronal precursors, angiogenesis and kidney
formation. In adults, it participates in wound healing, as well as
organ regeneration and tissue remodeling (19). However, its role in atrial fibrosis
has not yet been clarified. A previous study demonstrated that
bFGF-fibroblast growth factor receptor (FGFR)-heparan sulfate
proteoglycan (HSPG) complexes were able to activate the
mitogen-activated protein kinase (MAPK; ERK1/2) signaling pathway,
thereby activating cardiac fibroblasts and leading to collagen
deposition, decreased degradation, disorder of metabolic balance
and, ultimately, fibrosis (20).
Therefore, this study aimed to investigate the interrelation of
bFGF, HGF and the MAPK signaling pathway with atrial fibrosis in
patients with AF and RHD. The results indicate that bFGF is able to
promote the development of atrial fibrosis, while HGF functions in
an opposite manner in patients with AF and RHD. The MAPK signaling
pathway may be the molecular basis for these roles in atrial
fibrosis.
Patients and methods
Study population
Twenty patients with RHD who underwent valve
replacement were included as the study subjects. The patients were
aged between 30 and 70 years and had heart function grades ranging
from I to III. The exclusion criteria were: infective endocarditis,
hyperthyroidism, serious liver, kidney or lung dysfunction,
malignant tumor, coronary athero-sclerotic heart disease and
chronic pulmonary heart disease. The patients were divided into two
groups, with 10 patients in the sinus rhythm (SR) group and 10 in
the AF group. The study was approved by the Ethics Committee of
Renmin Hospital of Wuhan University (Wuhan, China) and all patients
provided written informed consent.
Human myocardium samples
Samples were collected from the right atrium of 20
patients with RHD who underwent valve replacement. Written informed
consent was obtained from the family of prospective donors and the
patient. The samples were obtained according to the regulations of
the Cardiovascular Research Institute of Wuhan University.
Materials
Primary antibodies against p38 (cat. no. 9212),
mitogen-activated protein kinase/extracellular signal-regulated
kinase 1/2 (MEK1/2; cat. no. 9122), c-Jun N-terminal kinase 1/2
(JNK1/2; cat. no. 9258), phospho-MEK1/2Ser217/221 (cat. no. 9154),
phospho-JNK1/2 (cat. no. 4668), ERK1/2 (cat. no. 4695),
phospho-ERK1/2Thr202/Thr204 (cat. no. 4370) and
phospho-p38Thr180/Thr182 (cat. no. 4511) were purchased from Cell
Signaling Technology, Inc. (Danvers, MA, USA). Antibodies against
HGF (ab10678) were purchased from Abcam Ltd. (Cambridge, UK).
Connective tissue growth factor (CTGF; sc-73869) and
glyceraldehyde-3-phosphate dehydrogenase (GAPDH; MB001) antibodies
were obtained from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA,
USA) and Bioworld Technology Inc. (Minneapolis, MN, USA)
respectively. The bicinchoninic acid (BCA) protein assay kit was
purchased from Pierce Biotechnology (Rockford, IL, USA).
Histological analysis
During surgery, ~200 mg of the right atrial
myocardium was collected prior to extracorporeal circulation being
established. This sample of myocardium was subsequently divided
into two parts; one part was rapidly placed in a liquid nitrogen
jar and immediately transferred to a −80°C refrigerator and the
remaining part was immediately washed with saline solution and
fixed with 10% neutral buffered formalin. Following this, several
sections (4–5 μm thick) were prepared and the hematoxylin
and eosin (H&E)-stained sections were used to determine the
cross-sectional area of the myocytes. Evidence of interstitial and
perivascular collagen deposition was visualized using Masson’s
trichrome staining and then high-magnification light micrographs
were captured using light microscopy. Collagen volume (%) was
measured using an image quantitative digital analysis system
(Image-Pro Plus 6.0, Media Cybernetics, Inc., Rockville, MD,
USA).
Immunohistochemistry
Using the EnVision™ two-step method, the
tissue specimens were fixed in formaldehyde solution, embedded in
paraffin and sliced. The paraffin was then removed. HGF and bFGF
antibodies were added at a concentration of 1:100, prior to the
specimens being incubated for ~60 min at room temperature and
rinsed three times in phosphate-buffered saline (PBS).
EnVision™ (50 μl) reagent was added to each
section and the sections were subsequently incubated for ~60 min at
room temperature, flushed with PBS, stained with
3,3’-diaminobenzidine (DAB), counterstained with hematoxylin and
placed on a neutral gum mount. The appearance of red or brownish
yellow granules in the cytoplasm indicated a positive result.
Computer image analysis was used to determine the density of the
positively stained area and for relative quantitative analysis.
Western blotting
A total of 50 μg protein was extracted from
the myocardial tissue, lysed in radio-immunoprecipitation assay
(RIPA) lysis buffer and used for sodium dodecyl
sulfate-polyacrylamide gel electrophoresis (SDS-PAGE). The proteins
were then transferred to nitrocellulose membranes and blocked with
5% non-fat dry milk in Tris-buffered saline (TBS) for 90 min at
room temperature. Following this, the membranes were probed with
various primary antibodies overnight. The next day, the membranes
were washed with 1X TBS and Tween 20 (TBST) and incubated for 1 h
with horseradish peroxidase-labeled mouse anti-rabbit antibody
(1:2,000) and anti-avidin antibodies (1:1,000) in double anti-TBST
fluid. Following the membrane being washed three times, the film
was placed in 10 ml LumiGLO® solution for 1 min. After
being developed, the images were placed into an automatic image
analyzer to determine the expression of the proteins and the
reference gray-scale values. A monoclonal anti-GAPDH antibody was
used separately as a loading control.
Statistical analysis
The data are presented as the mean ± standard error
of the mean. Comparisons between two groups were performed using an
unpaired Student’s t-test. P<0.05 was considered to indicate a
statistically significant difference.
Results
General clinical characteristics of the
two groups of patients
All patients underwent preoperative routine testing
of urine, stools, blood coagulation, blood biochemistry, chest
X-ray, electrocardiography and ultrasonic cardiogram. The general
clinical characteristics that were analyzed included age, gender,
LVEF, LAD and AF duration. With regard to gender, age, New York
classification of cardiac function (NYHA) and LVEF, the two groups
were not significantly different (Table I). However, in the AF group, the
LAD was significantly increased when compared with that of the SR
group (P<0.05).
 | Table I.General clinical characteristics of
the study population. |
Table I.
General clinical characteristics of
the study population.
| Characteristics | SR (n=10) | AF (n=10) |
|---|
| Gender
(male/female) | 4/6 | 5/5 |
| Age (years) | 46.01±10.38 | 49.51±11.04 |
| AF duration
(months) | - | 10.51±2.04 |
| NYHA (II/III) | 3/7 | 4/6 |
| LAD (mm) | 42.41±7.31 | 57.23±12.30a |
| LVEF (%) | 62.01±9.38 | 58.21±10.80 |
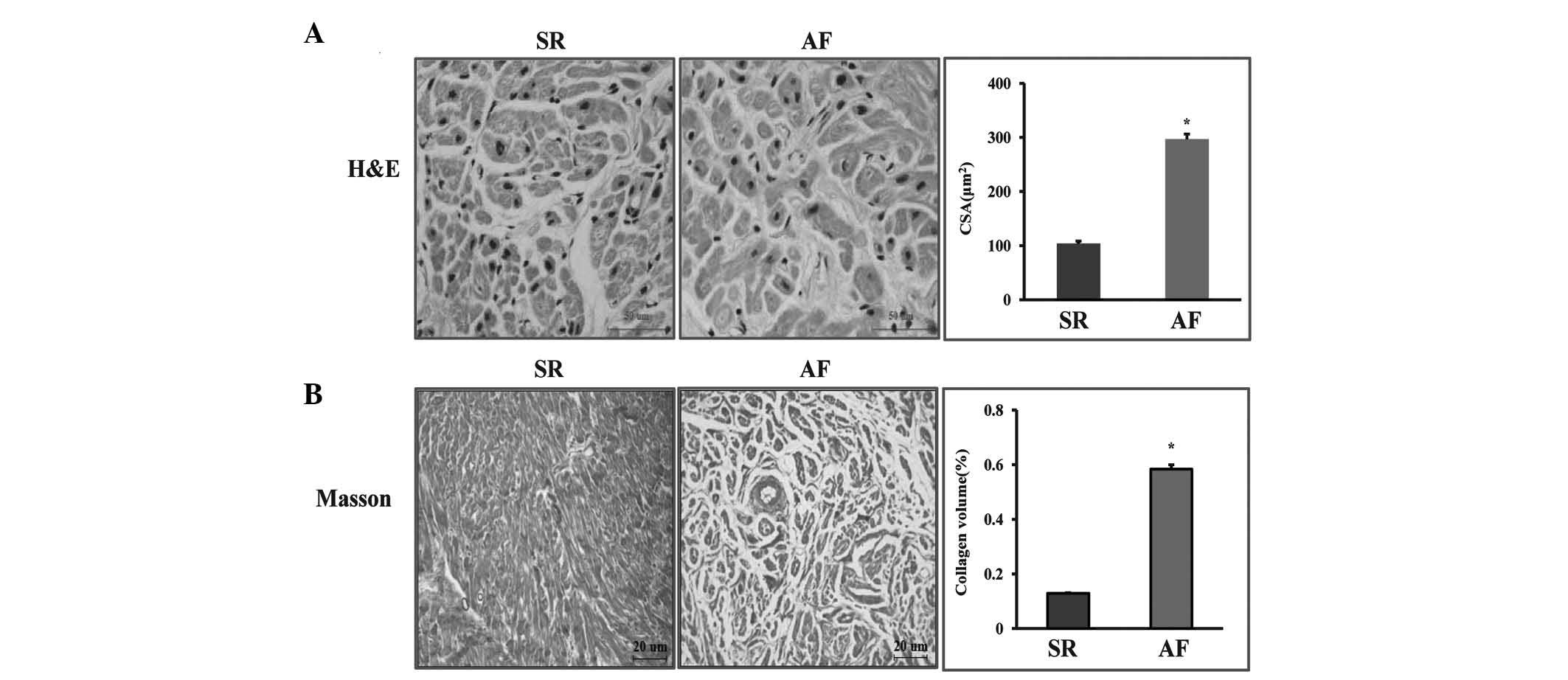
Myocardial cell and fibrosis
morphology
To investigate the role of AF in the morphology of
patients with RHD, samples were collected from the right atrium of
twenty patients with RHD who underwent valve replacement. H&E
and Masson’s trichrome staining indicated that AF had an adverse
effect on cardiac remodeling. From the H&E staining, it was
observed that the myocardial cell diameter of the patients in the
AF group was significantly expanded. Fibrosis was quantified by
visualizing the total amount of collagen present in the
interstitial spaces of the myocardial tissue and by determining the
collagen volume. Interstitial fibrosis was observed in the SR group
and the AF group; however, it was markedly increased in the AF
group. The AF group showed a significant increase in total collagen
volume compared with that in the SR group (P<0.05; Fig. 1).
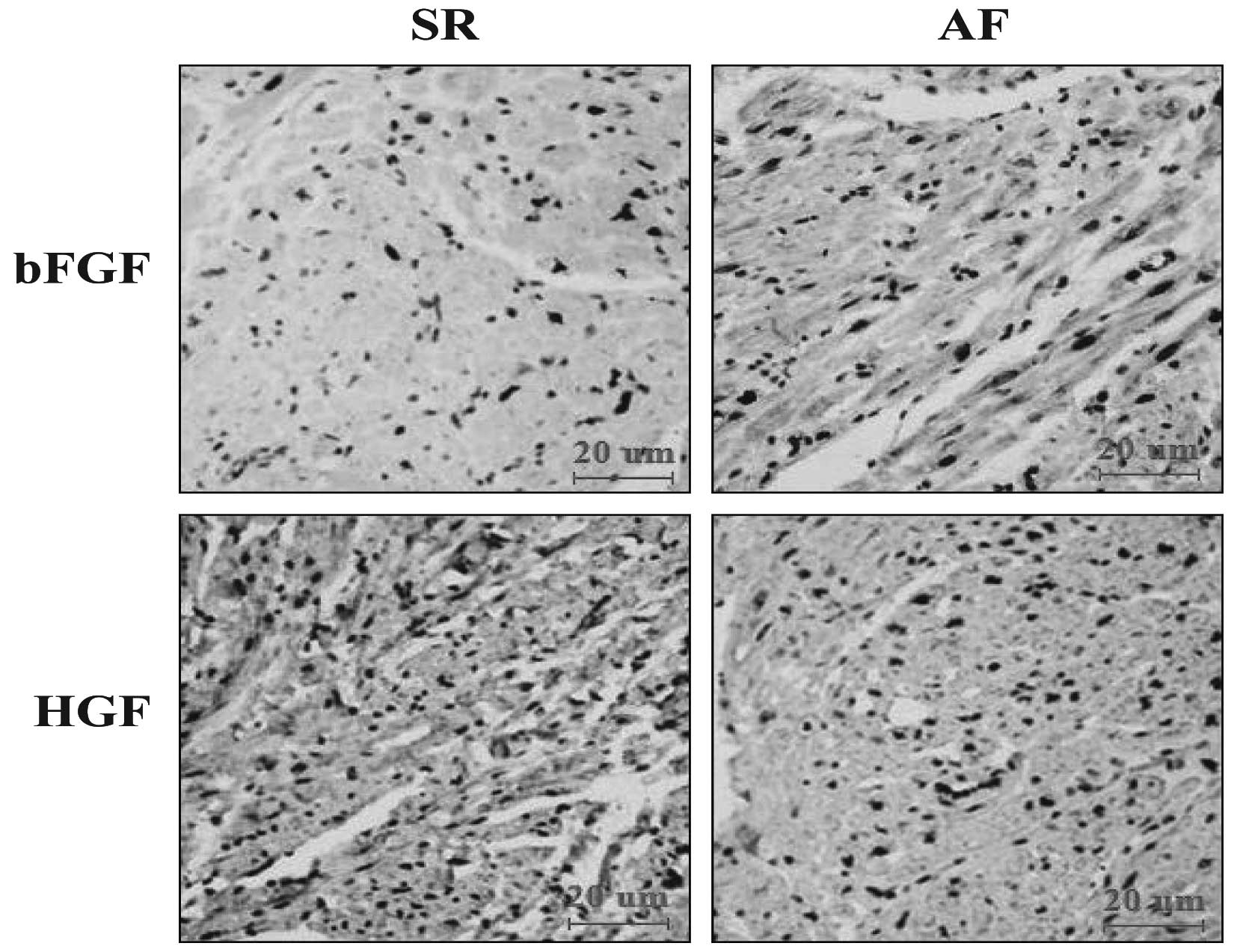
Effects of AF on bFGF and HGF
Immunohistochemical staining for bFGF and HGF was
performed in the tissue sections in order to assess the expression
levels of bFGF and HGF. The results showed that the intracellular
distribution of small bFGF granules in the atrial myocytes of the
SR group was lower than that in the AF group. By contrast, the
levels of HGF were significantly lower in the AF group compared
with those in the SR group (Fig.
2).
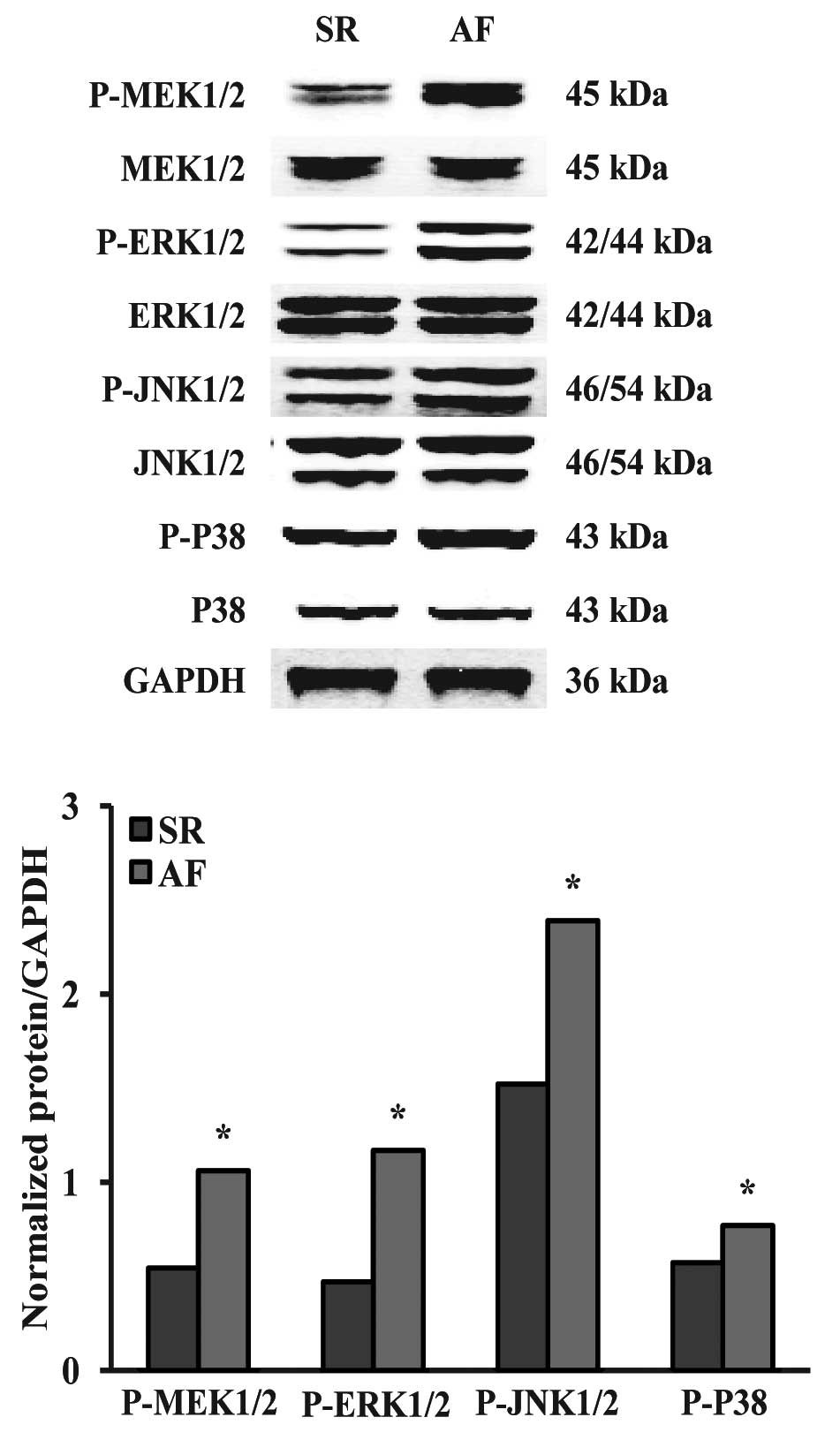
Effects of AF on the MAPK signaling
pathway
To explore the molecular mechanisms underlying the
increased bFGF levels and decreased HGF levels in the AF group, we
investigated the MAPK signaling pathway. It was observed that the
phosphorylation levels of MEK1/2, ERK1/2, p38 and JNK1/2 were
significantly increased in the AF group compared with those in the
SR group (Fig. 3).
Discussion
The most important observation in this study was
that myocardial cell diameter and levels of fibrosis were
significantly increased in patients with AF. Immunohistochemical
staining showed that levels of bFGF were increased, while levels of
HGF were reduced in the patients with AF compared with those in the
SR group. Further experiments showed that the phosphorylation level
of components of the MAPK pathway was increased markedly in the AF
group. To the best of our knowledge, these results are the first
direct indication that the expression levels of bFGF and HGF, which
are closely interrelated with fibrosis, are regulated by the MAPK
pathway in patients with AF.
The pathogenesis of AF is complex and has not been
completely elucidated. One recognized theory is that the occurrence
and maintenance of AF are closely associated with atrial
remodeling, including electrical and structural remodeling, and
that atrial fibrosis is the most important part of the structural
remodeling (21). An animal model
study of AF demonstrated the occurrence of atrial collagen
hyperplasia and AF from which the recovery of natural SR was rare.
In addition, while collagen hyperplasia and accumulation in the
interstitial cells affected the entire mechanics of the atrial
systolic and diastolic function, it also caused local
electro-cardiac heterogeneity in conduction, resulting in
arrhythmias and, in particular, AF (22). In the present study, we observed
that the atrial myocyte diameter and levels of fibrosis were
increased in patients with RHD and AF, using H&E and Masson’s
trichrome staining, respectively. In the SR group, only a few
collagen fibers were observed and there was little interstitial
fibrosis. The results of this study were consistent with a previous
study, which revealed that rheumatic mitral valve disease and
atrial structural remodeling were closely associated with the
occurrence of AF (23).
bFGF is a member of the FGF family. FGF family
members bind heparin and possess broad mitogenic and angiogenic
activities (24). bFGF has been
implicated in diverse biological processes, such as limb and
nervous system development (25),
wound healing (26) and tumor
growth (27). A previous study in
an animal model demonstrated that bFGF was important in continuous
hemodynamic-overload stimulation-induced myocardial cell
hypertrophy, myocardial fibrosis and myocardial collagen
hyperplasia (28). Our result
showed that bFGF was more diffusely distributed in the myocardial
cells of the patients with AF than in those in the SR group, .
HGF, another cell growth factor, is secreted by
mesenchymal cells and acts as a multi-functional cytokine on cells
of mainly epithelial origin. It regulates cell growth, motility and
morphogenesis by activating a tyrosine kinase signaling cascade
subsequent to binding to the proto-oncogenic c-Met receptor. The
ability of HGF to stimulate mitogenesis, cell motility and matrix
invasion makes it pivotal in angiogenesis, tumorigenesis and tissue
regeneration. Taniyama et al (29) studied hamster cardiomyopathy and
observed that, in lesions of the myocardium, the levels of HGF mRNA
and protein expression were reduced. Furthermore, myocardial
fibrosis and changes in cell shape were also observed. The results
of the immunohistochemical staining in our study showed that HGF
was highly expressed in the cytoplasm of the atrial myocytes of the
SR group, whereas the expression was significantly lower in the AF
group. Inoue et al (30)
revealed that HGF counteracted transforming growth factor β1
(TGFβ1) through the attenuation of CTGF induction and prevented
renal fibrogenesis in five out of six nephrectomized mice. Jun
et al (31) showed that
HGF/c-Met was able to enhance the proliferation and suppress the
expression of the fibrosis marker α-smooth muscle actin (α-SMA) in
ARPE-19 cells.
c-Met is a receptor tyrosine kinase that transduces
signals from the extracellular matrix into the cytoplasm by binding
to HGF/HGF ligand. It regulates a number of physiological
processes, including proliferation, scattering, morphogenesis and
survival. Ligand binding at the cell surface induces the
autophosphorylation of Met in its intracellular domain, which
provides docking sites for downstream signaling molecules.
Following activation by its ligand, c-Met interacts with the
PI3-kinase subunit PIK3R1, PLCG1, SRC, GRB2, STAT3 or the adapter
GAB1. The recruitment of these downstream effectors by Met leads to
the activation of numerous signaling cascades, including RAS-MAPK
kinase (MAPKK)-MAPKs (ERK/p38/JNK). The activation of
RAS-MAPKK-MAPK is associated with the morphogenetic effects.
Previous studies (32–34) have shown that MAPKs are important
in the process of fibrosis. The results in present study indicated
that the MAPK signaling pathway showed a significantly increased
level of activation in the AF group, and that the phosphorylation
levels of MEK1/2, ERK1/2, p38 and JNK1/2 were notably increased in
the AF group, compared with those in the SR group. Lu et al
(35) indicated that the
phosphorylation level of ERK1/2 was significantly lower in
claudin-7 transfected cells than control cells following HGF
treatment. In addition, Cohen et al (36) demonstrated that oncostatin M
(OSM)-induced HGF secretion was inhibited by PD-98059 (a specific
pharmacological inhibitor of ERK1/2), SB-203580 (a p38 MAPK
inhibitor) and SP-600125 (a JNK inhibitor) by 70, 82 and 100%,
respectively. Yang et al (37) showed that bFGF was able to induce
neuronal differentiation of mouse bone marrow stromal cells via
FGFR-1, MAPK/ERK and activator protein 1 (AP-1). bFGF has also been
demonstrated to activate the MAPK and nuclear factor κB (NFκB)
pathways to control the production of matrix metalloproteinase-13
in human adult articular chondrocytes (38).
In conclusion, bFGF may promote the development of
atrial fibrosis, while HGF may function in an opposite manner in
patients with RHD and AF. The MAPK signaling pathway may be the
molecular basis for these effects in atrial fibrosis.
Acknowledgements
The authors would like to thank
Professor Jun Xia, who helped in the collection of the samples of
the right atrium, and all of the members of the Department of
Cardiology and the Cardiovascular Research Institute of Renmin
Hospital of Wuhan University for their expert technical assistance
and advice. This study was supported by the National Natural
Science Foundation of China (grant no. 81170085).
References
|
1.
|
Cao X, Wang S, Jiang L, Liu L, Huang H and
Lu Z: Embolic events in 93 elderly Chinese patients with atrial
fibrillation. Chin Med J (Engl). 113:320–323. 2000.PubMed/NCBI
|
|
2.
|
Man J and Marchlinski FE: Atrial
fibrillation ablation and heart failure. Curr Cardiol Rep.
14:571–576. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3.
|
Ollivier R, Donal E, Veillard D, Pavin D,
Hamonic S, Daubert JC and Mabo P: Early and late cardiac
ventricular reverse remodeling after catheter ablation for lone
paroxysmal atrial fibrillation. Ann Cardiol Angeiol (Paris).
60:1–8. 2011. View Article : Google Scholar
|
|
4.
|
Efremidis M, Sideris A, Xydonas S, Letsas
KP, Alexanian IP, Manolatos D, Mihas CC, Filippatos GS and Kardaras
F: Ablation of atrial fibrillation in patients with heart failure:
reversal of atrial and ventricular remodelling. Hellenic J Cardiol.
49:19–25. 2008.PubMed/NCBI
|
|
5.
|
Olasinska-Wisniewska A, Mularek-Kubzdela
T, Grajek S, Marszalek A, Sarnowski W, Jemielity M, Seniuk W,
Lesiak M, Prech M and Podzerek T: Impact of atrial remodeling on
heart rhythm after radiofrequency ablation and mitral valve
operations. Ann Thorac Surg. 93:1449–1455. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6.
|
de Oliveira IM, Oliveira BD, Scanavacca MI
and Gutierrez PS: Fibrosis, myocardial crossings, disconnections,
abrupt turns, and epicardial reflections: do they play an actual
role in human permanent atrial fibrillation? A controlled necropsy
study. Cardiovasc Pathol. 22:65–69. 2013.
|
|
7.
|
Platonov PG, Mitrofanova LB, Orshanskaya V
and Ho SY: Structural abnormalities in atrial walls are associated
with presence and persistency of atrial fibrillation but not with
age. J Am Coll Cardiol. 58:2225–2232. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8.
|
Kim SJ, Choisy SC, Barman P, Zhang H,
Hancox JC, Jones SA and James AF: Atrial remodeling and the
substrate for atrial fibrillation in rat hearts with elevated
afterload. Circ Arrhythm Electrophysiol. 4:761–769. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
9.
|
Cha TJ, Ehrlich JR, Zhang L, Shi YF,
Tardif JC, Leung TK and Nattel S: Dissociation between ionic
remodeling and ability to sustain atrial fibrillation during
recovery from experimental congestive heart failure. Circulation.
109:412–418. 2004. View Article : Google Scholar
|
|
10.
|
Hong CS, Cho MC, Kwak YG, Song CH, Lee YH,
Lim JS, Kwon YK, Chae SW and Kim DH: Cardiac remodeling and atrial
fibrillation in transgenic mice overexpressing junctin. FASEB J.
16:1310–1312. 2002.PubMed/NCBI
|
|
11.
|
John B, Stiles MK, Kuklik P, et al:
Electrical remodelling of the left and right atria due to rheumatic
mitral stenosis. Eur Heart J. 29:2234–2243. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12.
|
de Groot NM, Zeppenfeld K, Wijffels MC,
Chan WK, Blom NA, Van der Wall EE and Schalij MJ: Ablation of focal
atrial arrhythmia in patients with congenital heart defects after
surgery: role of circumscribed areas with heterogeneous conduction.
Heart Rhythm. 3:526–535. 2006.PubMed/NCBI
|
|
13.
|
Ishikawa H, Jo JI and Tabata Y: Liver
anti-fibrosis therapy with mesenchymal stem cells secreting
hepatocyte growth factor. J Biomater Sci Polym Ed. 23:2259–2272.
2012.PubMed/NCBI
|
|
14.
|
Okunishi K, Dohi M, Nakagome K, Tanaka R,
Mizuno S, Matsumoto K, Miyazaki J, Nakamura T and Yamamoto K: A
novel role of hepatocyte growth factor as an immune regulator
through suppressing dendritic cell function. J Immunol.
175:4745–4753. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
15.
|
Iwata K, Sawa Y, Kitagawa-Sakakida S,
Kawaguchi N, Matsuura N, Nakamura T and Matsuda H: Gene
transfection of hepatocyte growth factor attenuates the progression
of cardiac remodeling in the hypertrophied heart. J Thorac
Cardiovasc Surg. 130:719–725. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
16.
|
Cecchi F, Rabe DC and Bottaro DP:
Targeting the HGF/Met signaling pathway in cancer therapy. Expert
Opin Ther Targets. 16:553–572. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17.
|
Trusolino L, Bertotti A and Comoglio PM:
MET signalling: principles and functions in development, organ
regeneration and cancer. Nat Rev Mol Cell Biol. 11:834–848. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
18.
|
Faletto DL, Kaplan DR, Halverson DO, Rosen
EM and Vande Woude GF: Signal transduction in c-met mediated
motogenesis. EXS. 65:107–130. 1993.PubMed/NCBI
|
|
19.
|
Zhang YW and Vande Woude GF: HGF/SF-met
signaling in the control of branching morphogenesis and invasion. J
Cell Biochem. 88:408–417. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
20.
|
Ornitz DM: FGFs, heparan sulfate and
FGFRs: complex interactions essential for development. Bioessays.
22:108–112. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
21.
|
Tan AY and Zimetbaum P: Atrial
fibrillation and atrial fibrosis. J Cardiovasc Pharmacol.
57:625–629. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22.
|
Xing XQ, Xu J, Su H and Lu YW: Association
between myocardial connexin 40 and 45 expression and myocardial
fibrosis in the rapid atrial pacing canine model. Zhonghua Xin Xue
Guan Bing Za Zhi. 39:176–180. 2011.(In Chinese).
|
|
23.
|
Chen YQ, Wang L, Su X, Tao L and Chen XF:
Calpain-I, calpastatin, caspase-3 and apoptosis in the human left
atrium in rheumatic atrial fibrillation. Zhonghua Xin Xue Guan Bing
Za Zhi. 34:303–307. 2006.(In Chinese).
|
|
24.
|
Flamme I, Schulze-Osthoff K and Jacob HJ:
Mitogenic activity of chicken chorioallantoic fluid is temporally
correlated to vascular growth in the chorioallantoic membrane and
related to fibroblast growth factors. Development. 111:683–690.
1991.
|
|
25.
|
Toledo RN, Borin A, Cruz OL, Ho PL, Testa
JR and Fukuda Y: The action of topical basic fibroblast growth
factor in facial nerve regeneration. Otol Neurotol. 31:498–505.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
26.
|
Abe M, Yokoyama Y and Ishikawa O: A
possible mechanism of basic fibroblast growth factor-promoted
scarless wound healing: the induction of myofibroblast apoptosis.
Eur J Dermatol. 22:46–53. 2012.PubMed/NCBI
|
|
27.
|
Felix AS, Edwards RP, Stone RA, Chivukula
M, Parwani AV, Bowser R, Linkov F and Weissfeld JL: Associations
between hepatocyte growth factor, c-Met, and basic fibroblast
growth factor and survival in endometrial cancer patients. Br J
Cancer. 106:2004–2009. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28.
|
Virag JA, Rolle ML, Reece J, Hardouin S,
Feigl EO and Murry CE: Fibroblast growth factor-2 regulates
myocardial infarct repair: effects on cell proliferation, scar
contraction, and ventricular function. Am J Pathol. 171:1431–1440.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
29.
|
Taniyama Y, Morishita R, Aoki M, Hiraoka
K, Yamasaki K, Hashiya N, Matsumoto K, Nakamura T, Kaneda Y and
Ogihara T: Angiogenesis and antifibrotic action by hepatocyte
growth factor in cardiomyopathy. Hypertension. 40:47–53. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
30.
|
Inoue T, Okada H, Kobayashi T, Watanabe Y,
Kanno Y, Kopp JB, Nishida T, Takigawa M, Ueno M, Nakamura T and
Suzuki H: Hepatocyte growth factor counteracts transforming growth
factor-beta1, through attenuation of connective tissue growth
factor induction, and prevents renal fibrogenesis in 5/6
nephrectomized mice. FASEB J. 17:268–270. 2003.
|
|
31.
|
Jun EJ, Kim HS and Kim YH: Role of
HGF/c-Met in serum-starved ARPE-19 cells. Korean J Ophthalmol.
21:244–250. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
32.
|
Gu J, Liu X, Wang QX, Tan HW, Guo M, Jiang
WF and Zhou L: Angiotensin II increases CTGF expression via
MAPKs/TGF-β1/TRAF6 pathway in atrial fibroblasts. Exp Cell Res.
318:2105–2115. 2012.PubMed/NCBI
|
|
33.
|
Ambrosino C, Iwata T, Scafoglio C,
Mallardo M, Klein R and Nebreda AR: TEF-1 and C/EBPbeta are major
p38alpha MAPK-regulated transcription factors in proliferating
cardiomyocytes. Biochem J. 396:163–172. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
34.
|
Nagai Y, Miyata K, Sun GP, Rahman M,
Kimura S, Miyatake A, Kiyomoto H, Kohno M, Abe Y, Yoshizumi M and
Nishiyama A: Aldosterone stimulates collagen gene expression and
synthesis via activation of ERK1/2 in rat renal fibroblasts.
Hypertension. 46:1039–1045. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
35.
|
Lu Z, Ding L, Hong H, Hoggard J, Lu Q and
Chen YH: Claudin-7 inhibits human lung cancer cell migration and
invasion through ERK/MAPK signaling pathway. Exp Cell Res.
317:1935–1946. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
36.
|
Cohen M, Marchand-Adam S, Lecon-Malas V,
Marchal-Somme J, Boutten A, Durand G, Crestani B and Dehoux M: HGF
synthesis in human lung fibroblasts is regulated by oncostatin M.
Am J Physiol Lung Cell Mol Physiol. 290:L1097–L1103. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
37.
|
Yang H, Xia Y, Lu SQ, Soong TW and Feng
ZW: Basic fibroblast growth factor-induced neuronal differentiation
of mouse bone marrow stromal cells requires FGFR-1, MAPK/ERK, and
transcription factor AP-1. J Biol Chem. 283:5287–5295. 2008.
View Article : Google Scholar
|
|
38.
|
Muddasani P, Norman JC, Ellman M, van
Wijnen AJ and Im HJ: Basic fibroblast growth factor activates the
MAPK and NFkappaB pathways that converge on Elk-1 to control
production of matrix metalloproteinase-13 by human adult articular
chondrocytes. J Biol Chem. 282:31409–31421. 2007.
|