Introduction
As increasing evidence supports the hypothesis that
brain inflammation is involved in the pathogenesis of epilepsy
(1), the pivotal role of immune
inflammatory reaction in epilepsy is increasingly being recognized
(2). Pro-inflammatory chemokines,
as chemotactic factors that control leukocyte migration under
physiological and pathological conditions, are important components
in neuro-inflammation. An increasing number of studies suggest that
they play a critical role in epileptogenesis (3,4). For
example, numerous chemokines and chemokine receptors, including
CCL2/CCR2 (5), CCL3/CCL4 (6), CCL5 (7), CCR5 (8,9),
CCR7/CCR8/CCR9/CCR10 (10), CXCL1
(11), CXCL12/CXCR4 (12) and CX3CL1/CX3CR1 (13,14)
are implicated in epilepsy. Among them, CCL2 (also known as MCP-1)
has been widely investigated in experimental rodents and clinical
epileptic patients (5,15–18).
It is considered that the inhibition of upregulated MCP-1
expression is likely to be beneficial in the treatment of
refractory epilepsy (3,17,18).
Microglia, the resident macrophages in the brain
parenchyma, play a key role in neuroinflammatory processes
(19). Seizure induced by
pilocarpine (PILO) results in rapid microglial activation in the
rat hippocampus (20) and it is
critical in the progression of neurodegeneration (21). However, it has been demonstrated
that the amelioration of tissue damage and PILO-induced seizure
severity may be achieved by attenuating microglial activation
(22).
Pyrrolidine dithiocarbamate (PDTC), a selective
nuclear factor-κB (NF-κB) inhibitor and antioxidant (23), exerts a significant anticonvulsant
effect and has demonstrated a neuroprotective effect on the CA1 and
CA3 regions of the hippocampus of a kainic acid (KA)-induced
seizure rat model (24,25) as well as on the piriform cortex in
the PILO status epilepticus (SE) model (26) by several mechanisms. However,
whether PDTC protects against hippocampal damage by inhibiting
MCP-1 upregulation and microglial activation in the PILO-induced SE
model has not yet been investigated.
In the present study, to the best of our knowledge,
we investigated the effects of PDTC on MCP-1 expression and
microglial activation in the hippocampus of SE model rats for the
first time. Moreover, seizure susceptibility, frequency and
severity as well as brain damage were also analyzed.
Materials and methods
Ethical approval
All the experimental procedures were conducted
according to the Guidance Suggestions for the Care and Use of
Laboratory Animals formulated by the Ministry of Science and
Technology of China (2006; Beijing, China). The study was approved
by The Medicine and Life Science Ethics Committee of Tongji
University (Shanghai, China).
PILO-induced seizures
Approximately 40 adult male Sprague-Dawley rats
(170–180 g) were purchased from Shang Hai Xipuer-BiKai experimental
animals Co., Ltd. Prior to the experiment, the rats were housed for
at least 1 week at a constant temperature of 22±1°C and relative
humidity (60%) and had free access to standard food and water under
a fixed 12-h light/dark cycle. Male Sprague-Dawley rats (230–250 g)
were randomly allocated into three groups: i) the saline group (NS
group), ii) the PILO-induced SE group (SE group), and iii) the SE
with PDTC-pretreatment group (PDTC group). The rats of the SE group
were treated with methylscopolamine (1 mg/kg, i.p; Sigma, St.
Louis, MO, USA) 30 min prior to the pilocarpine hydrochloride i.p.
injection (320 mg/kg; Sigma) (5).
The rats of the PDTC group were pretreated with PDTC (100 mg/kg,
i.p; Sigma) 24 h and 20 min prior to the administration of PILO.
The rats of the NS group were injected with an equivalent volume of
normal saline. The seizure activity was then scored according to
the system developed by Racine (27). Only the animals that reached a
seizure grade of ≥5 were selected for further analysis. All the
animals successfully established were sacrificed for analysis 24 h
after SE onset. Thirty-two rats were used in the analysis for
seizure onset time, 27 rats were used in the analysis for the
falling numbers, 15 rats were used in the analysis for the pattern
of MCP-1 and tissue damage 24h after SE, and 5 rats were used in
the analysis for the microglia activation in the three groups.
Seizure observation
Following the administration of PILO, the behavioral
changes of the rats were immediately observed using a video camera.
The seizure onset time to grades 3 (SOT3) and 5 (SOT5) as well as
the number of falls in the following 3 h after injecting PILO were
recorded by 2 independent observers blinded to the sample identity.
The seizure onset time to grade 3 and 5 were used to evaluate
seizure susceptibility (28),
while the number of falls was used to estimate seizure frequency
and severity.
Tissue processing
The rats were deeply anesthetized with 10% chloral
hydrate and then transcardially perfused with saline followed by 4%
paraformaldehyde. The brain was rapidly removed and post-fixed in
the same fixative for 24 h followed by rinsing with
phosphate-buffered saline (PBS) containing 30% sucrose at 4°C for
≥2 days (29). Tissue sections (50
μm thick) were cut using a cryostat.
Double immunofluorescence staining
Fifteen rats were used for studying the pattern of
MCP-1 24-h after SE. For double immunofluorescence analysis, after
pretreatment with 0.01 M citrate buffer (pH 6.0) for 5 min at 95°C,
the sections were incubated with a mixture of primary antibodies in
PBS containing 0.3% Triton X-100 and 1% normal bovine serum for 1 h
at room temperature (RT) followed by overnight incubation at 4°C.
The primary antibodies were as follows: polyclonal goat antibody
against MCP-1 (sc-1785, 1:100; Santa Cruz Biotechnology, Inc.,
Santa Cruz, CA, USA), mouse monoclonal antibody against NeuN
(1:1,000; Chemi-Con, Rosemont, IL, USA), MCP-1 and polyclonal
rabbit antibody against GFAP (1:500; DakoCytomation, Glostrup,
Denmark). After washing three times with PBS for 15 min, the
secondary antibodies donkey-anti-goat 488 (1:500; Invitrogen,
Carlsbad, CA, USA), donkey-anti-rabbit-Cy3 (711-165-152, 1:500;
Jackson ImmunoResearch, West Grove, PA, USA) and
donkey-anti-mouse-Cy3 (715-165-150, 1:500; Jackson ImmunoResearch)
were used to detect MCP-1, GFAP and NeuN. The tissues were then
washed with PBS, mounted onto glass slides and coverslips were
applied.
Immunohistochemistry (IHC)
IHC was performed to investigate microglial
activation. Briefly, all sections were initially incubated with
0.3% H2O2 for 15 min at RT. The sections were
then incubated with rabbit anti-Iba1 polyclonal antibody (1:4,000;
Wako Pure Chemical Industries, Ltd., Osaka, Japan) for 1 h at RT
followed by overnight incubation at 4°C. Then, the biotinylated
donkey anti-rabbit secondary antibody (711-065-152, 1:500; Jackson
ImmunoResearch) was used for a 3-h incubation at RT. After washing
three times for 5 min each, the sections were visualized with
3,3′-diaminobenzidine (DAB) in 0.1 M Tris buffer and mounted on
gelatin-coated slides.
Fluoro-Jade C (FJC) staining
Fifteen rats were used for studying tissue damage
24-h after SE. FJC staining was used to evaluate neuronal
degeneration. The procedures were conducted as described by Wang
et al(30).
Cell counting
Cell counts were performed by two different
investigators who were blind to the classification of tissues.
Regarding MCP-1-positive cells, 3 sections throughout the bilateral
hippocampus (at ~4.80–5.60 mm from the bregma) of each rat in the
SE and PDTC groups were selected. Regarding FJC-positive cells, 5
or 6 sections throughout the bilateral hippocampus (at ~2.88–4.16
mm from the bregma) were used and the cells were counted with a ×10
objective magnification. All the cells in every region of each rat
in the unilateral hippocampus were calculated using the following
formula: Total number of positive cells counted in bilateral
hippocampus/2 × corresponding cell numbers of sections. For the
quantification of microglial cells, the methods described by Yeo
et al(14) were used. All
the immunoreactive cells were counted regardless of the intensity
of labeling.
Statistical analysis
Data are provided as the mean ± SEM. Statistical
analysis was performed using SPSS software version 17.0 (SPSS,
Inc., Chicago, IL, USA). All the data with the exception of the
comparison of microglial activation were analyzed using an
independent sample test, while one-way ANOVA was used to determine
the statistical significance of microglial activation in the three
groups. P<0.05 was considered to indicate a statistically
significant difference.
Results
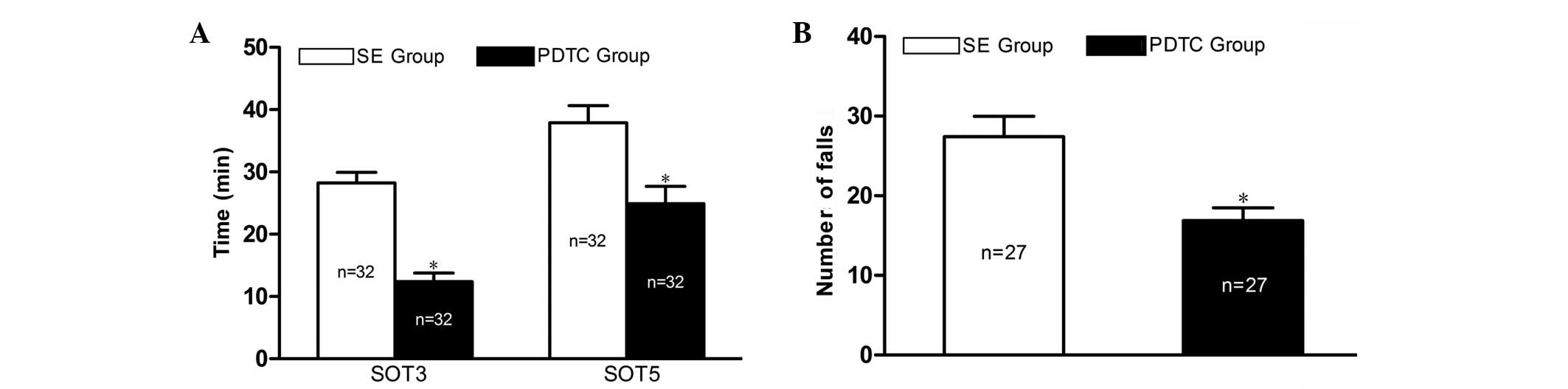
Pretreatment with PDTC enhances seizure
susceptibility, but reduces the frequency and severity of
PILO-induced seizures
No seizures were observed in the rats of the NS
group. The mean seizure onset times to grades 3 (SOT3) and 5 (SOT5)
of the rats in the SE group were 28.2±1.7 and 37.9±2.8 min,
respectively. The rats of the PDTC group exhibited significantly
shorter SOT3 and SOT5 (12.4±1.4 and 24.9±2.8 min, respectively,
P<0.01; Fig. 1A) compared with
the rats of the SE group. However, PDTC attenuated the frequency
and severity of the PILO-induced seizures, which was reflected by a
significantly reduced number of falls in the PDTC group (16.9±1.6
vs. 27.4±2.6, P<0.01) during the 3 h following the injection of
PILO (Fig. 1B).
PDTC exerts a neuroprotective effect on
the hippocampal CA1 and CA3 regions of PILO-induced SE model
rats
In the NS group, no FJC-positive cells were detected
in the CA1 and CA3 regions of the rat hippocampus (Fig. 2A and B). In addition, NeuN staining
indicated that the pyramidal cells of the hippocampal CA1 and CA3
regions were structurally intact with closely aligned cells and
clear nuclei (Fig. 2A′ and B′).
However, 24 h after SE, the hippocampal structures were markedly
damaged since numerous FJC-positive cells were detected and many
neurons were shown to be missing by NeuN staining (Fig. 2C, C′, D and D′). The numbers of
FJC-positive cells in the CA1 and CA3 regions were significantly
reduced in the PDTC group compared with those in the SE group
(15.0±2.3 vs. 29.4±4.7 and 34.6±3.9 vs. 66.5±6.6, respectively;
P<0.01; Fig. 2E, F and G).
Opposite results were obtained following NeuN staining (Fig. 2E′ and F′). In the PDTC group, the
NeuN staining indicated that the structure of the pyramidal cells
of the hippocampal CA1 and CA3 were damaged and small cells were
missing.
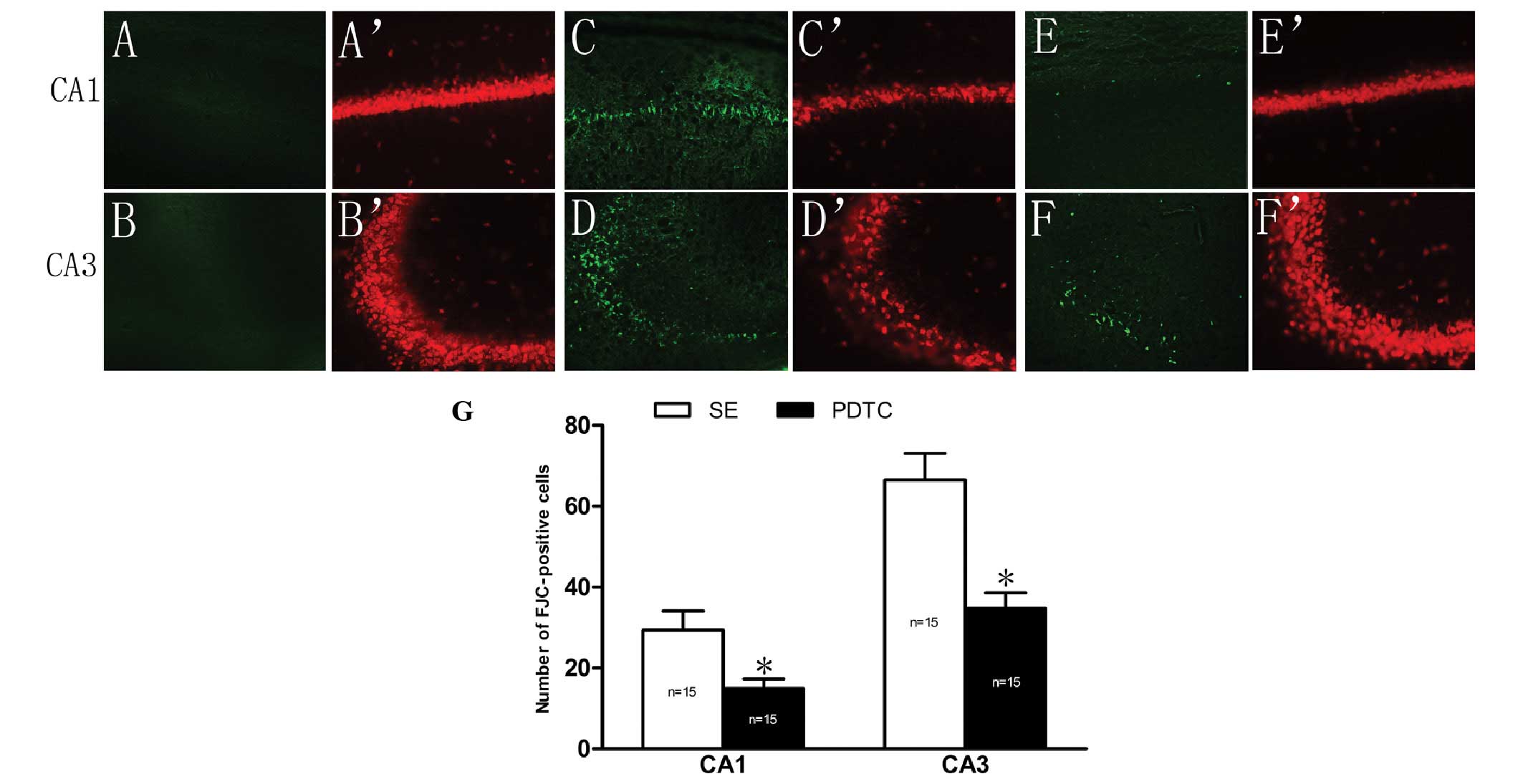 | Figure 2Effect of PDTC pretreatment on
PILO-induced neuronal damage in the hippocampal CA1 and CA3
regions. FJC staining in (A and B) the NS group, (C and D) the SE
group and (E and F) the PDTC group; NeuN staining in (A′ and B′)
the NS group, (C′ and D′) the SE group and (E′ and F′) the PDTC
group, SE resulted in significant neuronal damage in the
hippocampal CA1 and CA3 regions compared with that in the NS group
(A, B, A′ and B′), since a large number of FJC-positive cells were
detected by FJC staining and many neurons were shown to be missing
by NeuN staining (C, D, C′ and D′). The number of FJC-positive
cells was 29.4±4.7 and 66.5±6.6 in the CA1 and CA3 regions of the
rats in the SE group, respectively. By contrast, the numbers of
FJC-positive cells in the CA1 (15.0±2.3) and CA3 (34.6±3.9) regions
were significantly reduced in the PDTC group (P<0.01) (E and F),
which was also confirmed by NeuN staining (E′ and F′). (G)
Quantitative analysis of the number of FJC-positive cells in the
CA1 and CA3 regions of the rats in the SE and PDTC groups (mean ±
SEM). *P<0.01 vs. the SE group. PDTC, pyrrolidine
dithiocarbamate; PILO, pilocarpine; SE, status epilepticus; NS,
saline-treated; FJC, Fluoro-Jade C. (A–F′) Magnification, ×200. |
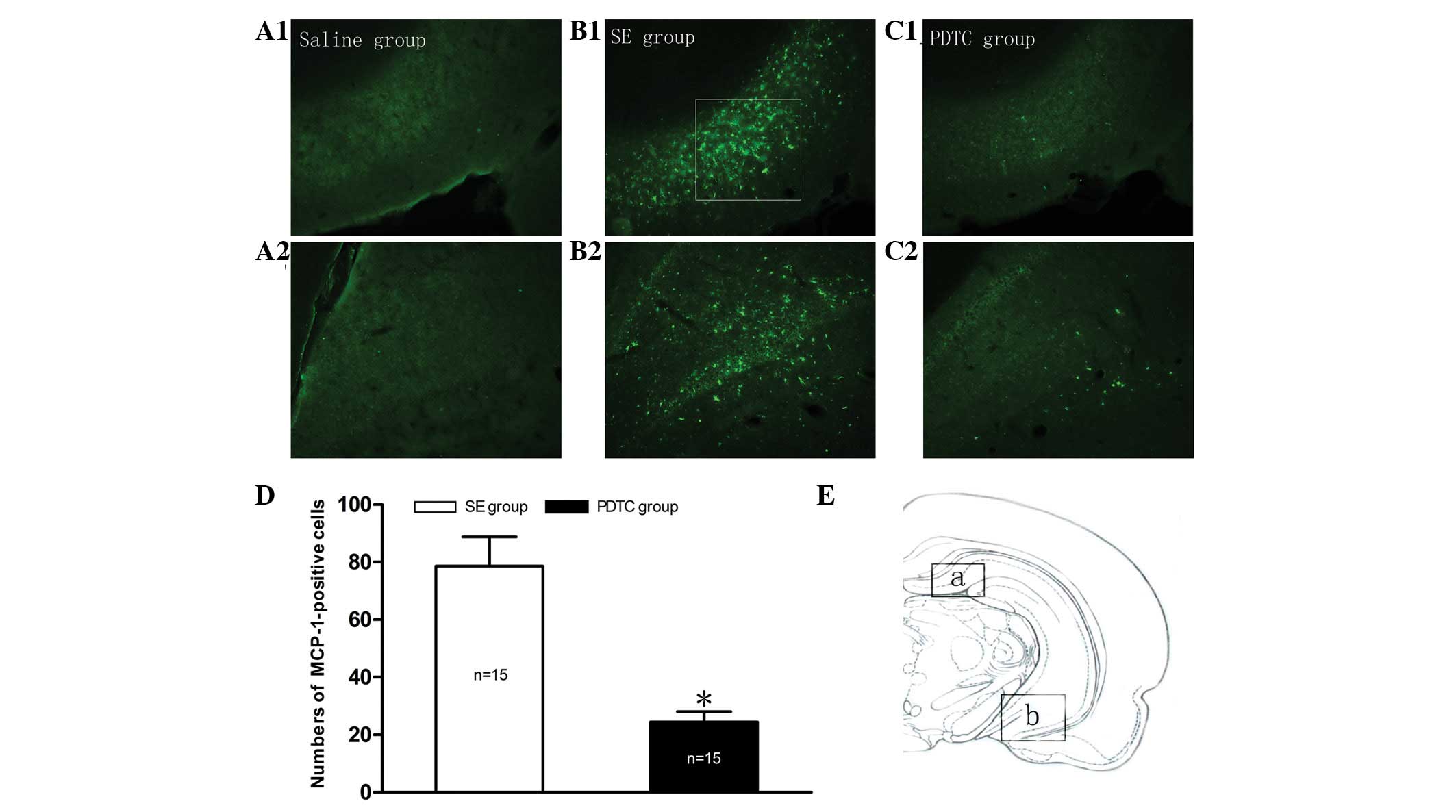
Increased chemokine MCP-1 expression in
the rat hippocampus following PILO-induced SE is inhibited by PDTC
pretreatment
No MCP-1 immunolabeled cells were observed in the
hippocampus of the rats in the NS group (Fig. 3A1 and A2). However, as shown in
Fig. 3B1 and B2, MCP-1 was
steadily expressed in the two regions of the hippocampus (boxed
areas a and b in Fig. 3E) 24 h
following PILO-induced SE. Therefore, these areas were selected for
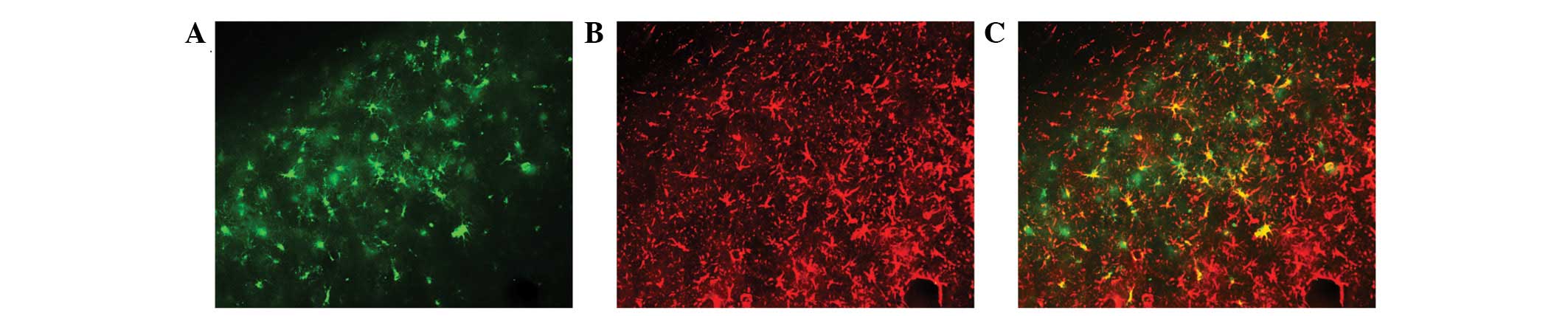
further study and quantitative analysis. As shown in Figs. 3 and 4, SE resulted in MCP-1 overexpression and
almost all the positive cells were co-localized with GFAP (Fig. 4C). However, the overexpression of
MCP-1 was markedly suppressed by PDTC pretreatment (Fig. 3C1 and C2). Quantitative analysis
indicated that the relative total number of MCP-1 immunopositive
cells in the PDTC group was significantly reduced compared with
that in the SE group (24.5±3.6 vs. 78.6±10.2, respectively;
P<0.01; Fig. 3D).
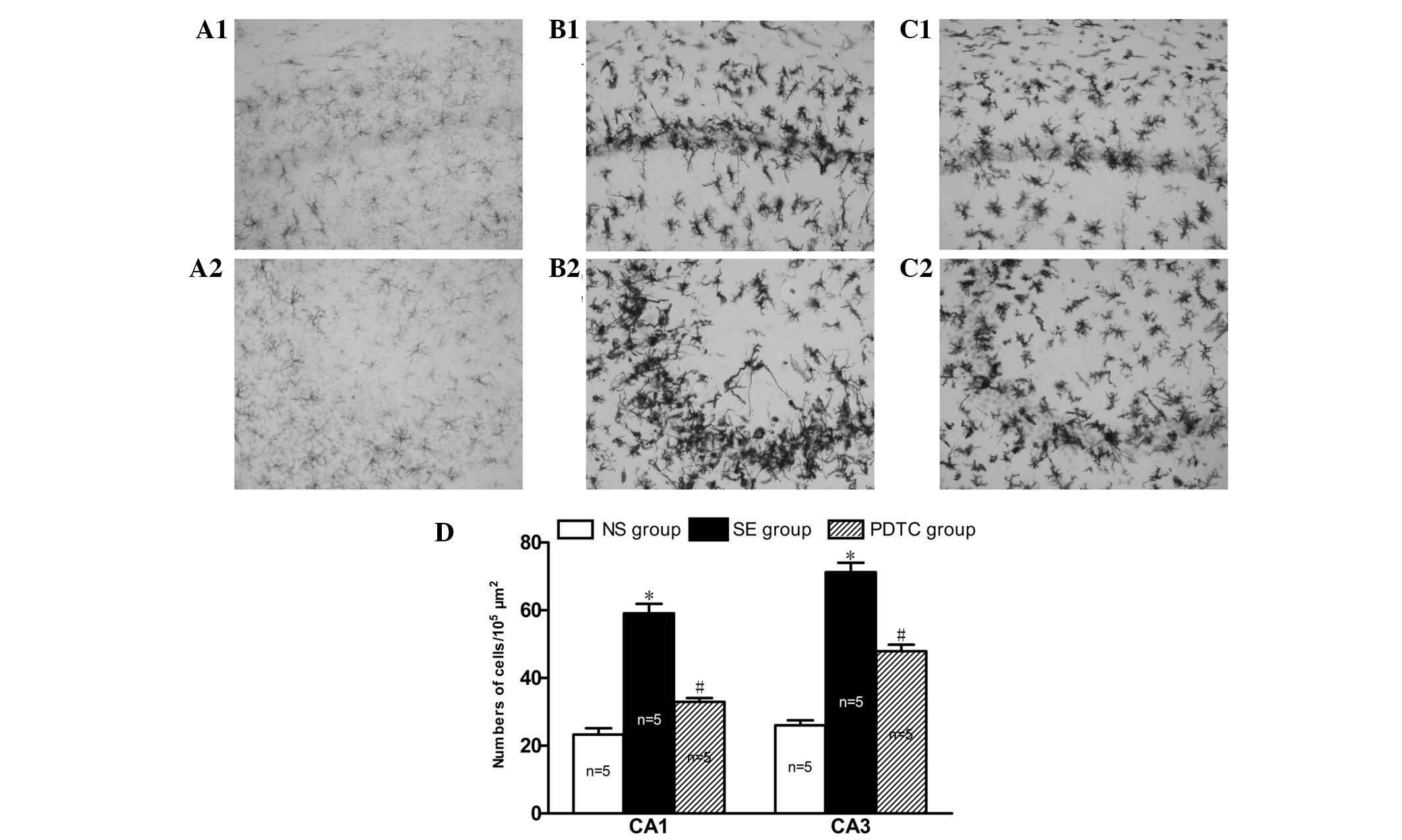
PDTC inhibits microglial activation in
the CA1 and CA3 regions of the hippocampus in PILO-induced SE
rats
Microglia cells were equally distributed and
exhibited resting states with thin cell bodies and slender
processes in the CA1 and CA3 regions of the hippocampus in the rats
in the NS group (Fig. 5A1 and A2).
However, 24 h after SE, the microglia were activated, demonstrating
a hyper-ramified and amoeboid or phagocytic appearance (Fig. 5B1 and B2). Notably, the numbers of
activated microglia migrating to the death-susceptible pyramidal
neuron cell layers CA1 (59.1±6.2 vs. 23.3±4.2, P<0.01) and CA3
(71.3±6.3 vs. 26.1±3.2, P<0.01) were significantly increased in
the SE group compared with the NS group (Fig. 5D). However, the accumulation of
activated microglia at these sites was significantly reduced in the
PDTC group compared with the SE group (33.0±2.6 vs. 59.1±6.2,
respectively, P<0.01 and 47.9±4.3 vs. 71.3±6.3, respectively,
P<0.01; Fig. 5C1, C2 and
D).
Discussion
The present study demonstrated that: i) pretreatment
with PDTC enhances seizure susceptibility and exerts an
anticonvulsant action; ii) PDTC has a neuroprotective effect on the
hippocampal CA1 and CA3 regions of rats with PILO-induced SE; and
iii) PDTC inhibits MCP-1 overexpression and attenuates microglial
activation in the hippocampus of the PILO-induced SE model
rats.
PDTC, a selective NF-κB inhibitor and antioxidant
(23), has been previously
demonstrated to exert anticonvulsant activity and to have a
neuroprotective effect through stimulating the adenosine A1
receptor (25) and antagonizing
the activated NOS II-peroxynitrite signaling cascade (24) in a KA-induced seizure model. Our
data showed that PDTC enhanced seizure susceptibility, as indicated
by the shorter SOT3 and SOT5, but reduced the frequency and
severity of the PILO-induced seizures and protected the CA1 and CA3
regions of the hippocampus against neurodegeneration. In the
present study, the observation that PDTC enhanced seizure
susceptibility was similar to the findings of Lubin et
al(31) but not in agreement
with those by Yu et al(28). Lubin et al(31) suggested that PDTC is related to the
reduced expression of brain-derived neurotrophic factor (BDNF) gene
following inhibition of the NF-κB pathway, while Yu et
al(28) observed that PDTC
significantly extended the seizure onset time. The contrasting data
from Lubin et al(31) and
Yu et al(28) may be
related to the different drugs tested (DDTC and SN50 vs. PDTC) as
the same SE model (the KA model) was used in both. The
contradictory results between the present study and the study by Yu
et al(28) may be related
to the different SE models used (the PILO-induced seizure model vs.
the KA model), since the same drug (PDTC) was used. It is not
possible to determine the idiographic mechanisms of these
contradictory findings in this study and further investigation is
required. Furthermore, the current study demonstrated that PDTC
reduced the frequency and severity of PILO-induced seizures. The
following three aspects should be considered: i) PDTC has a
protective effect on the hippocampus. Shin et al(25) demonstrated that PDTC reduced the
number of animals developing generalized SE through stimulating the
adenosine A1 receptor and then blocking seizure-induced oxidative
stress and subsequent neuronal loss (25). ii) PDTC alleviates brain
inflammation. Liu et al(32) reported that PDTC exerted an
antiepileptic effect by inhibiting the NF-κB pathway and thereby
reducing the expression levels of NF-κB/P65, tumor necrosis
factor-α (TNF-α), interleukin (IL)-β and IL-10 in the hippocampus.
iii) PDTC attenuates microglial activation. Abraham et
al(22) showed that PDTC
alleviates seizures by attenuating microglial activation.
MCP-1 may act as a modulator of neuronal activity
and neuroendocrine functions in the brains of normal rats (33). However, it has been reported that
MCP-1 protein and mRNA levels as well as those of its receptor,
CCR2, are significantly increased in the brain tissues of epileptic
patients or experimental rodents (5,15–18).
The cell types expressing MCP-1 were mainly neurons and astrocytes
24 h after SE (17), and mainly
microglia 2 days post SE (15). In
the present study, the numbers of the MCP-1 immunopositive cells
were significantly increased in the rat hippocampus and were mainly
expressed by astrocytes 24 h after PILO-induced SE. The difference
in the cell types expressing MCP-1 may be associated with the
different time points after SE. However, the mechanism underlying
the differential expression of MCP-1 in various types of cells at
different time points after SE remains be elucidated, as the
available literature was not investigated. However, the
upregulation of MCP-1 was attenuated in rats pretreated with PDTC.
This may be explained by the selective NF-κB inhibition mechanism
of PDTC. It has been shown that inflammation induces the expression
of MCP-1 through the NF-κB signaling pathway under several
pathological conditions, such as progressive proteinuric
nephropathy (34), cardiac
ischemia-reperfusion injury (35)
and high glucose stimulation (36). Meanwhile, the NF-κB signaling
pathway has been shown to be involved in MCP-1 gene regulation
(37). Therefore, the results
obtained in the present study indicate that the inhibition of MCP-1
overexpression by PDTC in the context of epilepsy is likely to be
associated with the inhibition of NF-κB activation. Even though
PDTC exerts a dual mechanism as an NF-κB inhibitor and antioxidant,
the main mechanism by which PDTC acts in SE is likely to be the
inhibition of NF-κB since the pharmacological effect of PDTC has
been shown to be similar to that of double-stranded κB decoy DNA
which is a selective NF-κB inhibitor, while its antioxidant
properties are negligible (24).
Yu et al(28) confirmed
that PDTC pretreatment significantly decreased NF-κB activation 24
h after KA-induced SE.
Furthermore, to the best of our knowledge, our data
showed for the first time that PDTC attenuated microglial
activation and ameliorated neurodegenerative changes in the
hippocampus in SE model rats. Microglia cells became hyper-ramified
from their resting states after SE, indicating the expression of
inflammatory factors and the induction of cytotoxic activity
(38). In addition, activated
microglia synthesized and released IL-1β (39), osteopontin (OPN) (40), telomerase (41), IL-6 and TNF-α (42), which contribute to the occurrence
of brain injury. This may explain the increased number of activated
microglia that migrated to the death-susceptible pyramidal neuron
cell layers CA1 and CA3 24 h after SE. Regarding the attenuation of
microglial activation and ameliorative neurodegeneration, three
aspects should be considered. Firstly, these effects may be related
to the inhibition of MCP-1 since activated microglia migrate to
areas of injury guided by chemokines in the inflammatory process
(11) and MCP-1 was critical for
the microglia migration and subsequent neurodegeneration (17). Secondly, the dual mechanism of
action of PDTC should also be considered. PDTC was previously
demonstrated to affect microglia-mediated neuroinflammation through
inhibiting ROS and NF-κB pathways (43). Thirdly, the neuroprotective effect
of PDTC may be related to the inhibition of microglial function and
the reduction of cytokine releasing or cytotoxic activity.
However, the present study has several limitations.
Firstly, the activation of NF-κB in each group was not
investigated, since this has been previously examined in other
studies (24,28,31).
Secondly, the effects of PDTC on MCP-1 mRNA expression in the
context of epilepsy was not investigated, which will be examined in
future studies by our group.
In conclusion, the present study demonstrated that
the protective effects of PDTC on hippocampal damage may be
associated with inhibited microglial activation in the PILO-induced
SE rat model. To the best of our knowledge, these results indicate
for the first time that the activation of the NF-κB pathway
contributes to MCP-1 upregulation and microglial activation under
the context of epilepsy.
Acknowledgements
We thank all the members of the Laboratory of Dr
Yu-Qiang Ding (Department of Anatomy and Neurobiology, Tongji
University School of Medicine) for their sincere help in this
study. This study was supported by the National Natural Science
Foundation of China (grant no. 81271441) to Professor Yongbo
Zhao.
References
|
1
|
Vezzani A, Friedman A and Dingledine RJ:
The role of inflammation in epileptogenesis. Neuropharmacology.
69:16–24. 2013. View Article : Google Scholar
|
|
2
|
Vezzani A and Rüegg S: The pivotal role of
immunity and inflammatory processes in epilepsy is increasingly
recognized: introduction. Epilepsia. 52(Suppl 3): 1–4.
2011.PubMed/NCBI
|
|
3
|
Fabene PF, Bramanti P and Constantin G:
The emerging role for chemokines in epilepsy. J Neuroimmunol.
224:22–27. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Sharma A: Genome-wide expression analysis
in epilepsy: a synthetic review. Curr Top Med Chem. 12:1008–1032.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Foresti ML, Arisi GM, Katki K, Montañez A,
Sanchez RM and Shapiro LA: Chemokine CCL2 and its receptor CCR2 are
increased in the hippocampus following pilocarpine-induced status
epilepticus. J Neuroinflammation. 6:402009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Guzik-Kornacka A, Sliwa A, Plucinska G and
Lukasiuk K: Status epilepticus evokes prolonged increase in the
expression of CCL# and CCL4 mRNA and protein in the rat
brain. Acta Neurobiol Exp (Wars). 71:193–207. 2011.PubMed/NCBI
|
|
7
|
Fiala M, Avagyan H, Merino JJ, Bernas M,
Valdivia J, Espinosa-Jeffrey A, Witte M and Weinand M: Chemotactic
and mitogenic stimuli of neuronal apoptosis in patients with
medically intractable temporal lobe epilepsy. Pathophysiology.
20:59–69. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Louboutin JP, Chekmasova A, Marusich E,
Agrawal L and Strayer DS: Role of CCR5 and its ligands in the
control of vascular inflammation and leukocyte recruitment required
for acute excitotoxic seizure induction and neural damage. FASEB J.
25:737–753. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Marusich E, Louboutin JP, Chekmasova AA
and Strayer DS: Lymphocyte adhesion to CCR5 ligands is reduced by
anti-CCR5 gene delivery. J Neurol Sci. 308:25–27. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Liu JX, Cao X, Tang YC, Liu Y and Tang FR:
CCR7, CCR8, CCR9 and CCR10 in the mouse hippocampal CA1 area and
the dentate gyrus during and after pilocarpine-induced status
epilepticus. J Neurochem. 100:1072–1088. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Johnson EA, Dao TL, Guignet MA, Geddes CE,
Koemeter-Cox AI and Kan RK: Increased expression of the chemokines
CXL1 and MIP-1α by resident brain cells precedes neutrophil
infiltration in the brain following prolonged soman-induced status
epilepticus in rats. J Neuroinflammation. 8:412011.
|
|
12
|
Lee TS, Mane S, Eid T, Zhao H, Lin A, Guan
Z, Kim JH, Schweitzer J, King-Stevens D, Weber P, Spencer SS,
Spencer DD and de Lanerolle NC: Gene expression in temporal lobe
epilepsy is consistent with increased release of glutamate by
astrocytes. Mol Med. 13:1–13. 2007.PubMed/NCBI
|
|
13
|
Xu Y, Zeng K, Han Y, Wang L, Chen D, Xi Z,
Wang H, Wang X and Chen G: Altered expression of CX3CL1 in patients
with epilepsy and in a rat model. Am J Pathol. 180:1950–1962. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yeo SI, Kim JE, Ryu HJ, Seo CH, Lee BC,
Choi IG, Kim DS and Kang TC: The roles of fractalkine/CX3CR1 system
in neuronal death following pilocarpine-induced status epilepticus.
J Neuroimmunol. 234:93–102. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kim JE, Ryu HJ, Yeo SI and Kang TC: P2X7
receptor regulates leukocyte infiltrations in rat frontoparietal
cortex following status epilepticus. J Neuroinflammation. 7:652010.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Manley NC, Bertrand AA, Kinney KS, Hing TC
and Sapolsky RM: Characterization of monocyte chemoattractant
protein-1 expression following a kainate model of status
epilepticus. Brain Res. 1182:138–143. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Sheehan JJ, Zhou C, Gravanis I, Rogove AD,
Wu YP, Bogenhagen DF and Tsirka SE: Proteolytic activation of
monocyte chemoattractant protein-1 by plasmin underlies excitotoxic
neurodegeneration in mice. J Neurosci. 27:1738–1745. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wu Y, Wang X, Mo X, Xi Z, Xiao F, Li J,
Zhu X, Luan G, Wang Y, Li Y and Zhang J: Expression of monocyte
chemoattractant protein-1 in brain tissue of patients with
intractable epilepsy. Clin Neuropathol. 27:55–63. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Miller G: Neuroscience. The dark side of
glia. Science. 308:778–781. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Shapiro LA, Wang L and Ribak CE: Rapid
astrocyte and microglial activation following pilocarpine-induced
seizures in rats. Epilepsia. 49(Suppl 2): 33–41. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Rogove AD and Tsirka SE: Neurotoxic
responses by microglia elicited by excitotoxic injury in the mouse
hippocampus. Curr Biol. 8:19–25. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Abraham J, Fox PD, Condello C, Bartolini A
and Koh S: Minocycline attenuates microglia activation and blocks
the long-term epileptogenic effects of early-life seizures.
Neurobiol Dis. 46:425–430. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Nurmi A, Goldsteins G, Närväinen J,
Pihlaja R, Ahtoniemi T, Gröhn O and Koistinaho J: Antioxidant
pyrrolidine dithiocarbamate activates AKT-GSK signaling and is
neuroprotective in neonatal hypoxia-ischemia. Free Radic Biol Med.
40:1776–1784. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chuang YC, Chen SD, Lin TK, Chang WN, Lu
CH, Liou CW, Chan SH and Chang AY: Transcriptional upregulation of
nitric oxide synthase II by nuclear factor-kappaB promotes
apoptotic neuronal cell death in the hippocampus following
experimental status epilepticus. J Neurosci Res. 88:1898–1907.
2010.
|
|
25
|
Shin EJ, Jhoo JH, Kim WK, Jhoo WK, Lee C,
Jung BD and Kim HC: Protection against kainate neurotoxicity by
pyrrolidine dithiocarbamate. Clin Exp Pharmacol Physiol.
31:320–326. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Soerensen J, Pekcec A, Fuest C, Nickel A
and Potschka H: Pyrrolidine dithiocarbamate protects the piriform
cortex in the pilocarpine status epilepticus model. Epilepsy Res.
87:177–183. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Racine RJ: Modification of seizure
activity by electrical stimulation. II. Motor seizure.
Electroencephalogr Clin Neurophysiol. 32:281–294. 1972. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yu N, Di Q, Liu H, Hu Y, Jiang Y, Yan YK,
Zhang YF and Zhang YD: Nuclear factor-kappa B activity regulates
brain expression of P-glycoprotein in the kainic acid-induced
seizure rats. Mediators Inflamm. 2011:6706132011.PubMed/NCBI
|
|
29
|
Ryu HJ, Kim JE, Yeo SI, Kim MJ, Jo SM and
Kang TC: ReLA/P65-serine 536 nuclear factor-kappa B phosphorylation
is related to vulnerability to status epilepticus in the rat
hippocampus. Neuroscience. 187:93–102. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wang L, Liu YH, Huang YG and Chen LW:
Time-course of neuronal death in the mouse pilocarpine model of
chronic epilepsy using Fluoro-Jade C staining. Brain Res.
1241:157–167. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Lubin FD, Ren Y, Xu X and Anderson AE:
Nuclear factor-kappa B regulates seizure threshold and gene
transcription following convulsant stimulation. J Neurochem.
103:1381–1395. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Liu GJ, Huang JM, Li XB, Meng LQ and Huang
RY: Effect of pyrrolidine dithiocarbamate on expression of nuclear
factor-κB and related inflammatory factors in rat hippocampus after
epilepsy. Chin J Behav Med & Brain Sci. 20:784–786. 2011.(In
Chinese).
|
|
33
|
Banisadr G, Gosselin RD, Mechighel P,
Rostène W, Kitabgi P and Mélik Parsadaniantz S: Constitutive
neuronal expression of CCR2 chemokine receptor and its
colocalization with neurotransmitters in normal rat brain:
functional effect of MCP-1/CCL2 on calcium mobilization in primary
cultured neurons. J Comp Neurol. 492:178–192. 2005. View Article : Google Scholar
|
|
34
|
Donadelli R, Abbate M, Zanchi C, Corna D,
Tomasoni S, Benigni A, Remuzzi G and Zoja C: Protein traffic
activates NF-κB gene signaling and promotes MCP-1-dependent
interstitial inflammation. Am J Kidney Dis. 36:1226–1241. 2000.
|
|
35
|
Kim YS, Kim JS, Kwon JS, Jeong MH, Cho JG,
Park JC, Kang JC and Ahn Y: BAY 11–7082, a nuclear factor-κB
inhibitor, reduces inflammation and apoptosis in a rat cardiac
ischemia-reperfusion injury model. Int Heart J. 51:348–353.
2010.
|
|
36
|
Quan Y, Jiang CT, Xue B, Zhu SG and Wang
X: High glucose stimulates TNFα and MCP-1 expression in rat
microglia via ROS and NF-κB pathways. Acta Pharmacol Sin.
32:188–193. 2011.
|
|
37
|
Yadav A, Saini V and Arora S: MCP-1:
chemoattractant with a role beyond immunity: a review. Clin Chim
Acta. 411:1570–1579. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Streit WJ, Walter SA and Pennell NA:
Reactive microgliosis. Prog Neurobiol. 57:563–581. 1999. View Article : Google Scholar
|
|
39
|
Vezzani A, Conti M, De Luigi A, Ravizza T,
Moneta D, Marchesi F and De Simoni MG: Interleukin-1beta
immunoreactivity and microglia are enhanced in the rat hippocampus
by focal kainate application: functional evidence for enhancement
of electrographic seizures. J Neurosci. 19:5054–5065.
1999.PubMed/NCBI
|
|
40
|
Kim SY, Choi YS, Choi JS, Cha JH, Kim ON,
Lee SB, Chung JW, Chun MH and Lee MY: Osteopontin in kainic
acid-induced microglial reactions in the rat brain. Mol Cells.
13:429–435. 2002.PubMed/NCBI
|
|
41
|
Fu W, Lee J, Guo Z and Mattson MP:
Seizures and tissue injury induce telomerase in hippocampal
microglial cells. Exp Neurol. 178:294–300. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Ravizza T, Rizzi M, Perego C, Richichi C,
Velísková J, Moshé SL, De Simoni MG and Vezzani A: Inflammatory
response and glia activation in developing rat hippocampus after
status epilepticus. Epilepsia. 46(Suppl 5): 113–117. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Quan Y, Du J and Wang X: High glucose
stimulates gro secretion from rat microglia via ROS, PKC, and
NF-kappaB pathways. J Neurosci Res. 85:3150–3159. 2007. View Article : Google Scholar : PubMed/NCBI
|