Introduction
Dendritic cells (DCs) are the most potent
antigen-presenting cells (APCs) and are able to induce primary
immune responses. DCs possess the capability to capture, process
and present antigens and induce immune diseases. DCs are special
APCs which are able to present antigens to naïve and quiescent T
cells and consequently play a significant role not only in the
initiation but also in the maintenance of inflammation and allergic
diseases (1). Nasal polyps are
frequently presented at the Department of Otolarynology-Head and
Neck Surgery, The Shantou Central Hospital/Affiliated Shantou
Hospital of Sun Yet-Sen University, Shantou, China. The
pathogenesis of nasal polyps is not clear and now the majority of
scholars believe that nasal polyps are the result of a number of
complications and that the pathological immune system may play a
key role in their onset and development. The penetration of T
lymphocytes and eosinophils (EOSs), an imbalance in the proportion
of T helper 1 to T helper 2 (Th1/Th2) cells and a predominance of
the Th2 cell cytokines are characteristics of nasal polyps. DCs are
able to present antigens to T cells and activate them through
co-stimulating molecules and cytokines. Therefore DCs are likely
play a significant role in the pathogenesis of nasal polyps.
However, there have not been any direct studies into the
correlation between DCs and nasal polyps. Therefore, the present
study aimed to investigate the expression, distribution and
function of DCs in nasal polyps and also to study their role in
nasal polyp pathogenesis. It is known that the S-100 protein is a
non-specific marker of DCs and that CD1a/CD40 double immunostaining
is a comparatively specific marker of DCs. CD40 is a significant
co-stimulating molecule that causes DCs to activate T cells. In the
present study, immunohistochemical and double immunostaining
methods were used to detect the expression and distribution of
S-100 and CD1a/CD40 in the nasal polyps, to gain information
concerning the role of DCs in their pathogenesis.
Subjects and methods
Subjects
The study group consisted of 45 patients with nasal
polyps (25 males and 20 females), ranging in age from 11–74 years
(mean, 35.53 years) and 10 patients (8 males and 2 females), 17–62
years old, (mean, 34 years) with a deviation of the nasal septum
requiring surgical treatment that were assigned as the control. The
cases were diagnosed by histological examination (hematoxylin and
eosin staining). None of the patients received any type of
corticosteroid therapy for 30 days prior to surgery. The study was
conducted in accordance with the Declaration of Helsinki and with
approval from the Ethics Committee of Shantou Central Hospital.
Written informed consent was obtained from all patients and control
subjects after the nature and purpose of the study had been
explained. The nasal polyp and normal inferior turbinate tissue
specimens were collected during surgery. All samples were fixed in
10% buffered formalin, embedded in paraffin and cut into
4-μm sections.
Staining methods
An immunohistochemical staining Elivision™ two-step
method was performed to detect the S-100 protein. The kits were
provided by Fuzhou Maixin Biotechnology Development Co., Ltd.,
Fuzhou, China. A rabbit anti-human polyclonal antibody to the S-100
protein (Pharmingen International, San Diego, CA, USA) was used as
the primary antibody and a mouse anti-rabbit IgG monoclonal
antibody was used as the secondary antibody. 3,3′-Diaminobenzidine
(DAB)-hydrogen peroxide was used as a chromogen.
The staining of CD1a and CD40 was performed with
certain modifications according to the previous S-100 staining
method. Two types of primary antibody [mouse anti-human monoclonal
antibody to CD1a (Lab Vision Corporation, Waltham, MA, USA) and
rabbit anti-human monoclonal antibody to CD40 (Santa Cruz
Biotechnology, Inc., Santa Cruz, CA, USA)] were added to the
specimens at the same time. The specimens were incubated at 40°C
overnight and then washed with PBS three times. The secondary
antibodies (anti-mouse antibody-FITC, KPL; anti-rabbit antibody-PE,
Santa Cruz Biotechnology, Inc.) were added and the specimens were
rein-cubated at 37°C with the avoidance of light for 1.5 h.
A known esophagal carcinoma sample was used as a
positive control; the negative control slides were processed with
PBS liquid instead of the primary antibody, but included all other
steps of the procedure.
Expression of the S-100 protein was located in the
nucleus and cytoplasm of the DCs. Expression was determined using
Frowits’s method with double-blind reading by two independent
observers. Five fields of view in 400 were chosen for
determination. Appearance of earthy yellow or brown-yellow granules
in the cytoplasm was considered as a positive result. The
percentage of positive cells and staining intensity in five fields
of view were calculated and analyzed, and the cell grading was
performed as follows: i) According to the percentage of positive
cells, grade 0: no cell coloration or positive cell percentage
<5%; grade 1: 5–35% of cells were colored; grade 2: 36–65% of
cells were colored; grade 3: more than 65% of cells were colored.
ii) According to color depth, grade 0: no cell coloration or
unclear coloration; grade 1: earthy yellow; grade 2: brown-yellow;
grade 3: seal brown. The final score was the average value of above
two grading methods. Scores of 0, 0.5–1 and 1.5–3 were defined as
negative (−), positive (+) and strong positive (++), respectively.
The positive staining intensity of S-100 was calculated using an
Image-pro plus 5.0 analysis system (Media Cybernetics, Inc., MD,
USA) and presented as the IOD (integrated optical density, IOD =
average positive grey value-average positive background value).
CD1a and CD40 were expressed in the cytoplasm of the
DCs. Positive staining of CD1a was demonstrated by olivine
fluorescence and that of CD40 by a red fluorescence. The double
immunostaining of CD1a and CD40 produced a yellow fluorescence.
Every slice was taken five eye-shot in 400 times and analyzed using
a KS400 analysis system (Carl Zeiss, Inc., New York, NY, USA) to
check the distribution areas, quantity and density of the DCs.
Statistical analysis
Statistical calculations were performed using SPSS
10.0 (SPSS, Inc., Chicago, IL, USA) statistical software. The data
are expressed as mean ± SD. The unpaired and paired t-test was used
to compare the staining intensities of S-100, the distribution
areas and the quantity and density of the double
immunostain-positive cells. The S-100 and CD1a/CD40-positive rates
were shown as percentages and the data were analyzed by the
χ2 test. P<0.05 was considered to indicate a
statistically significant difference.
Results
Expression of S-100 protein
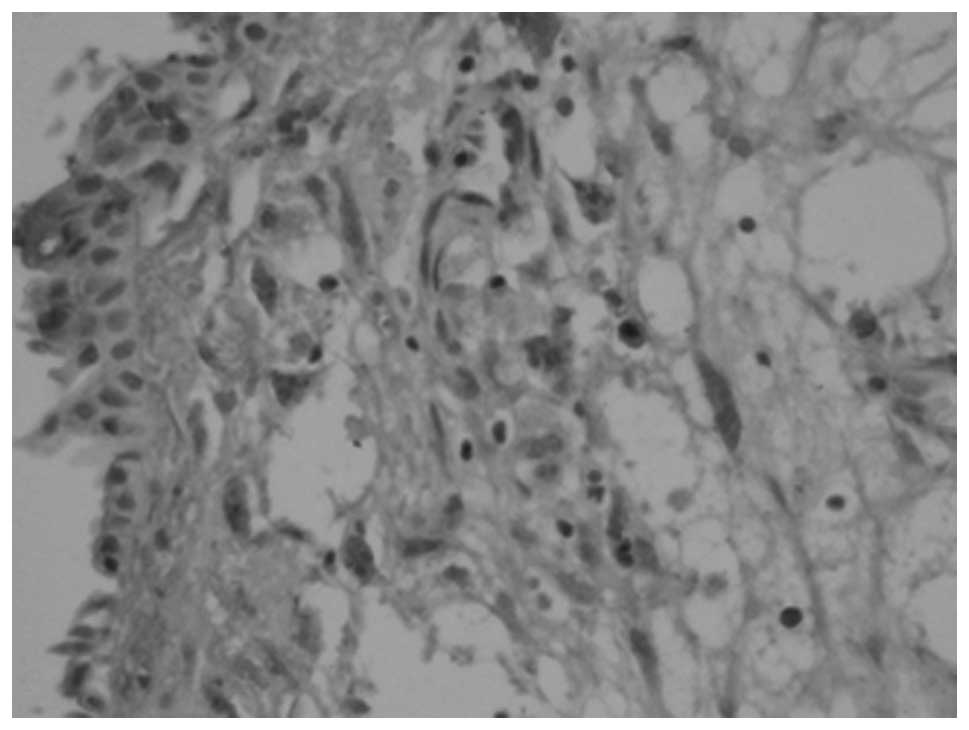
The S-100 protein-positive DCs were mainly
distributed in the submucosa of the nasal polyps (Fig. 1). In the inferior turbinate tissues
lined with ciliated epithelium, there were almost no S-100
protein-positive cells. The DCs were S-100 protein-positive in the
nasal polyp tissues in 40 cases; the positive rate was 88.9%.
Correspondingly, only one case from the control group was weakly
positive for the S-100 protein. When analyzed by IOD, the inferior
turbinate tissues were shown to seldomly express S-100 protein
(with a staining intensity of 890.02±723.24). By contrast, the
nasal polyp tissues had elevated expression levels of the S-100
protein (with a staining intensity of 8540.12±1249.79; Table I).
 | Table IS-100 expression in the nasal polyps
and normal inferior turbinate groups. |
Table I
S-100 expression in the nasal polyps
and normal inferior turbinate groups.
| Group | Positive rate
(%) | Frowits’ method
| IOD |
|---|
| − | + | ++ |
|---|
| Normal inferior
turbinate (n=10) | 10.0 | 9 | 1 | 0 | 890.02±723.24 |
| Nasal polyps
(n=45) | 88.9 | 5 | 19 | 21 | 8540.12±1249.79 |
| P-value | | | <0.01 | | <0.01 |
Expression of CD1a/CD40
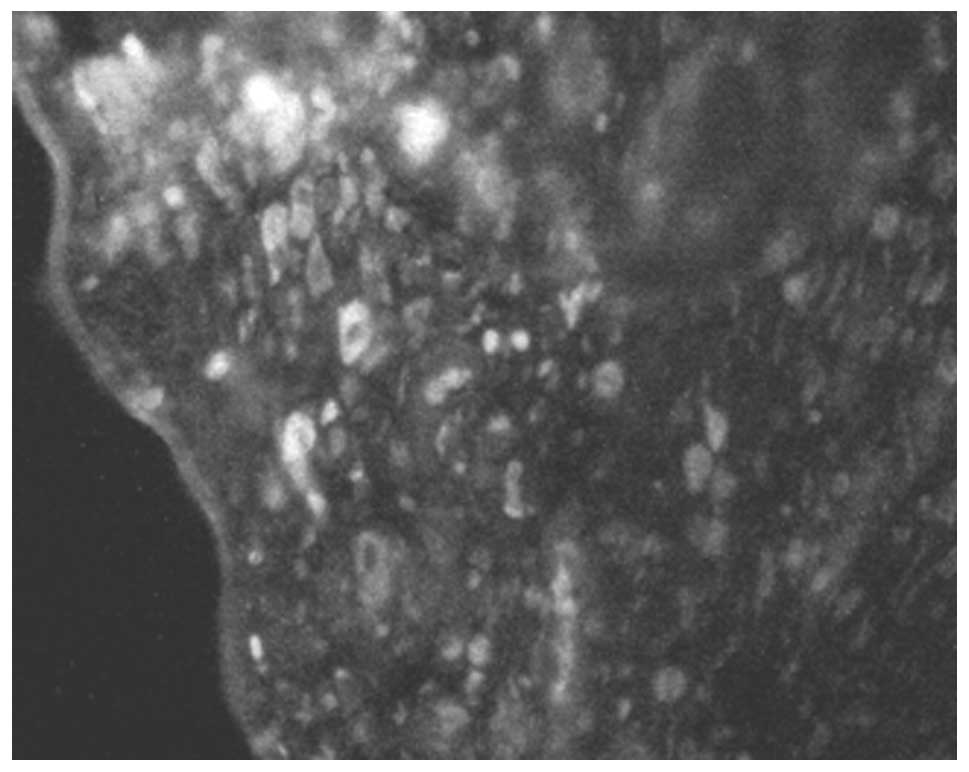
As was observed for S-100, the CD1a and CD40 double
stained-positive cells were mostly in the submucosa of the nasal
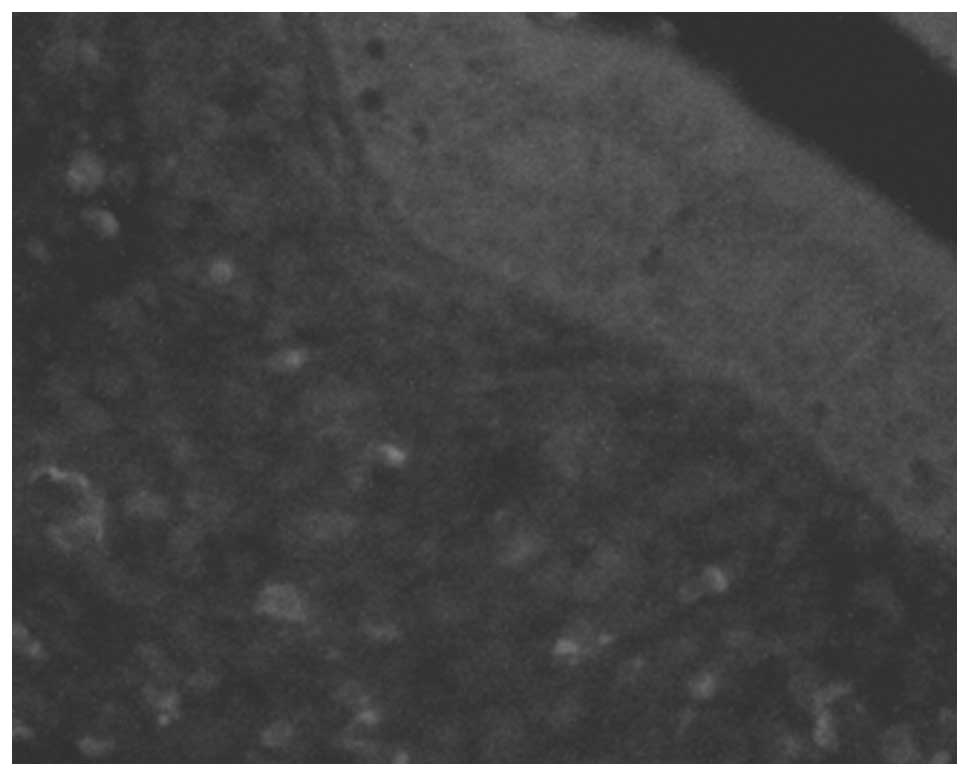
polyps (Fig. 2). In the inferior
turbinate tissues, there were rarely any double stained-positive
cells (Fig. 3). The positive rates
were 88.9 and 10.0% in the nasal polyps and inferior turbinate
tissues, respectively. The difference in the area, number and
density of the double stained cells between the control and study
groups was statistically significant (P<0.01; Table II).
| Table IIArea, number and density of the double
stained dentritic cells (DCs). |
Table II
Area, number and density of the double
stained dentritic cells (DCs).
| Group | Positive rate
(%) | Total area of double
stained cells (cells/mm2) | Total number of
double stained cells | Density of double
stained cells (cells/mm2) |
|---|
| Normal inferior
turbinate (n=10) | 10.0 | 299.3±177.6 | 46.3±38.3 | 589±456 |
| Nasal polyps
(n=45) | 88.9 | 3605.2±796.2 | 664.3±215.3 | 7125±2575 |
| P-value | | <0.01 | <0.01 | <0.01 |
Discussion
DCs are the most important APCs and are extremely
rare in normal tissues. Aimed at a variety of antigens, DCs express
various types of TOLL-like receptors and decide the type of
immunoreation (2). DCs are able to
capture antigens at the lymphocyte, process them into antigenic
epitopes and link with MHC II. DCs are activated following the
presentation of the MHC II-antigenic epitopes to the T cells and
produce numerous types of cytokines and chemokines. The chemokines
may then collect more DCs that are assembled in the pathological
tissues. DCs and T cells interact with each other through
co-stimulating molecules and cytokines. The DCs process the
antigenic epitope to the T cells and upregulate the CD40L on their
surface. The CD40L connects with the CD40 of the DCs to activate
the B7 molecules (CD80/CD86). The B7 molecules of the DCs activate
the CD28/CTLA-4 of the T cells, inducing the activation of the
cells and resulting in an immunoreaction (3).
The pathogenesis and mechanisms behind nasal polyps
are unclear. Bernstein’s hypothesis (4) has been accepted by the majority of
scholars. The effects of the changes in nasal aerodynamics,
inflammation and allergic factors induce serious inflammation in
the nasal mucosa. The infiltration of large numbers of inflammatory
cells and the effects of inflammation mediators lead to the nasal
mucosa becoming edemic. This causes a breaks in the epithelium,
submucosal extrusion and epithelialization, thus creating nasal
polyps.
The causes of nasal polyps include allergies,
genetic predisposition, autonomic dysfunction of the blood vessels
of the nasal mucosa and inflammation (4). The infiltration of inflammatory
cells, particularly eosinophils (EOSs) and T lymphocytes, and
abnormalities in the cytokines, including the upregulation of
interleukin (IL)-4, IL-5 and IL-10, are characteristics of the
immune dysfunction of nasal polyps (5,6). The
increase in the number of T lymphocytes, the imbalance in the
proportion of Th1/Th2 cells and the predominance of cytokines
produced by the Th2 cells have been proposed to be the most
significant immunological abnormalities (7).
The present study demonstrated that the presence of
DCs (S-100 protein-positive cells) in the normal inferior turbinate
tissue was rare. Compared with in the turbinates, the number of DCs
in the nasal polyps was markedly increased. This suggested that DCs
may play a significant role in the pathogenesis of nasal
polyps.
In addition, the CD40-positive nature of the surface
of the DCs suggested that DCs may react in the nasal polyps through
reciprocal interaction with T lymphocytes. The S-100 protein and
double stained cells were mainly located in the lamina propria
below the mucous membrane. Yoshimi et al(8) reported that DCs were only detectable
in the squamous epithelium and the authors believed that the
migration of DCs into the squamous epithelium may have been
regulated by cytokines, including IL-1 and granulocyte-macrophage
colony-stimulating factor (GM-CSF), released from the keratinocytes
which constitute the squamous epithelium.
On the basis of the information presented in the
present study and based on a review of the literature, we propose a
hypothesis for the mechanism of pathogenesis in nasal polyps: In
the normal nasal mucosa, DCs are infrequently identified scouting
for pathogens. When invasion by a pathogen occurs, different
subsets of the DCs expressed various types of Toll-like receptors
(TRLs) to combine with the pathogen-associated molecular patterns
(PAMPs) and became mature themselves (9). In a previous study, Claeys et
al(10) observed that TRL-2
and TRL-4 were expressed in the lamina propria below the mucous
membrane of the nasal polyp tissue and that DCs presented the MHC
II-antigenic epitope to T cells. The DCs were then connected with
the T cells by the CD40/CD40L cell surface molecules. Next, the B7
molecules (CD80/CD86) on the surface of the DCs combined with the
CD28/CTLA-4 of the T cells to induce the activation of the T cells,
causing an immunoreaction. The result was that either the DCs or T
cells became mature so that they were able to release chemokines to
attract more DCs and T cells to assemble in the pathogenic tissues.
This enhanced the cycle leading to increases in the quantities of
DCs (as shown in the present study) and T cells (11) in the nasal polyps. The effects of
the hyperplastic quantities of DCs and T cells then followed.
Firstly, hyperplastic quantities of DCs and T cells
are the main cause of an imbalance in the proportion of Th1/Th2
cells (12) and the predominance
of Th2 cell cytokines (13). This
may involve the following mechanism: DCs are the main infection
factors for the differentiation of Th0 (14). DCs are of two types;
myeloid-derived DCs (DC1) and lymphoid-related DCs (DC2). In
general, DC1 lead to the preferential development of a Th1 response
and DC2 to a Th2 response (15). A
study by Jahnsen et al identified that the amount of DC2 was
increased in inflammatory and antigen-stimulated tissues. DC2
induced a Th2 response; the number of Th2 cells increased and the
expression of Th2 cytokines was enhanced (16). Alternatively, different subsets of
the DCs express varying types of TRLs to lead to different
immunoreaction types. It has been reported that low
lipopolysaccharide (LPS) levels led to a Th2 response by TRL-4 and
that mice expressed a strong Th2 response following immunization
with TRL-2 ligands and ovalbumin (OVA) (17). Nasal polyps express high levels of
TRL-2 and TRL-4 (18) which are
relevant to the Th2 response. The DCs induce the maturation and
differentiation of T cells using cytokines and co-stimulating
molecules on the cell surface. The CD80 of the DCs act as the
co-stimulating molecule of the Th1 cells while CD86 belongs to the
Th2 cells. In the nasal polyp tissues, DCs induce an imbalance in
the proportion of the Th1/Th2 cells by causing the expression of
CD80/CD86 to become unbalanced (19). The reciprocal interaction of the
DCs and T cells through CD40/CD40L may induce DCs to secrete IL-12
(20). IL-12 is the most
significant cytokine in the transformation of Th0 cells to Th1
cells. However, scholars have demonstrated that IL-10 is highly
expressed in nasal polyp tissues (21). DCs are not only able to secrete
IL-10 themselves but are also able to induce T cells to secrete
IL-10 (15). IL-10 may restrain
the Th1 cell cytokines. Reider et al(22) demonstrated that IL-10 could
decrease the capacity of DCs in secreting IL-12 so that the
differentiation from Th0 to Th1 was restrained but differentiation
to Th2 was induced. Nasal polyps highly express TRL-2 (10,18)
which in turn induces a Th2 immune response (17). DCs which express TRL-2 are able to
induce T cells to secrete IL-4 (17). IL-4 is the most significant
cytokine in the transformation of Th0 cells to Th2 cells. In nasal
polyp tissues, IL-4 levels are increased (7) and cause the Th0 cells to transform
into Th2 cells. Th2 is able to secrete IL-4 itself. This process
became a cycle that encourages the predominance of the cytokines
produced by the Th2 cells in the nasal polyps. Otherwise, IL-4
encourages the B cells to transform into plasmocytes which secrete
IgE (23). The high level of IgE
has been reported to be one of the demonstrably correlative factors
in nasal polyp pathogenesis and has been identified to be
positively correlated with EOS infiltration (6).
In the nasal polyp tissues, the reciprocal control
of the DCs and T cells, through CD40/CD40L, B7-CD28/CTLA-4
co-stimulating molecules, the high levels of IL-4 and IL-10 and the
low expresson levels of IL-12, induced an imbalance in the
proportion of the Th1/Th2 cells and caused the predominance of Th2
cell cytokines.
Secondly, the hyperplastic quantities of the DCs and
T cells cause high expression levels of IL-5 in the nasal polyp
tissues; active DCs induce CD4+ T cells to secrete IL-5 (15,24).
In previous studies, the high level of IL-5 has been identified to
be one of the most significant demonstrably correlative factors
with nasal polyps and one of the promotory factors in EOS
infiltration (5,13). In the initial stages, IL-5 is
generated mostly by Th2 cells. As the nasal polyps develop, EOSs
replace the Th2 cells to become the most significant source of
IL-5. Yi et al (25) demonstrated
that IL-5 was able to restrain the maturation of DCs directly. In
nasal polyp tissues, DCs induced T cells to secrete IL-5 by
increasing the amount of EOS infiltration. The infiltrating EOSs
secrete IL-5 to restrain the maturation of the DCs. This allows the
DCs to pause in their juvenile stage, encouraging an effective
immunoreaction and the development of nasal polyps.
In conclusion, the present study identified that
there was a large quantity of DC infiltration in the nasal polyp
tissues. The DCs recognized the PAMPs via TRLs and initiated the
immunoreaction of the nasal polyps. Next, the DCs induced the
infiltration and differentiation of T cells using co-stimulating
molecules and cytokines. Following this, the DCs induced an
imbalance in the proportion of the Th1/Th2 cells (12) and the predominance of Th2 cell
cytokines through cell and serum mechanisms. In addition, the DCs
were able to induce the infiltration of the EOSs by IL-5.
References
|
1.
|
Inaba K: Dendritic cells as
antigen-presenting cells in vivo. Immunol Cell Biol. 75:206–208.
1997. View Article : Google Scholar : PubMed/NCBI
|
|
2.
|
Watts C, Zaru R, Prescott AR, Wallin RP
and West MA: Proximal effects of Toll-like receptor activation in
dendritic cells. Curr Opin Immunol. 19:73–78. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
3.
|
Grewal IS, Foellmer HG, Grewal KD, et al:
Requirement for CD40 ligand in costimulation induction, T cell
activation, and experimental allergic encephalomyelitis. Science.
273:1864–1867. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
4.
|
Bernstein JM, Gorfien J, Noble B and
Yankaskas JR: Nasal polyposis: immunohistochemistry and
bioelectrical findings (a hypothesis for the development of nasal
polyps). J Allergy Clin Immunol. 99:165–175. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
5.
|
Fan GK, Wang H and Takenaka H: Eosinophil
infiltration and activation in nasal polyposis. Acta Otolaryngol.
127:521–526. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
6.
|
Hirschberg A, Jókúti A, Darvas Z, et al:
The pathogenesis of nasal polyposis by immunoglobulin E and
interleukin-5 is completed by transforming growth factor-beta1.
Laryngoscope. 113:120–124. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
7.
|
Pawankar R: Nasal polyposis: an update:
editorial review. Curr Opin Allergy Clin Immunol. 3:1–6. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
8.
|
Yoshimi R, Takamura H, Takasaki K and
Kumagami H: Immunohistologic study of the nasal mucosa with
reference to Langerhans cells. Nihon Jibiinkoka Gakkai Kaiho.
96:1252–1257. 1993.(In Japanese).
|
|
9.
|
Drohan L, Harding JJ, Holm B, et al:
Selective developmental defects of cord blood antigen-presenting
cell subsets. Hum Immunol. 65:1356–1369. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
10.
|
Claeys S, de Belder T, Holtappels G, et
al: Human beta-defensins and toll-like receptors in the upper
airway. Allergy. 58:748–753. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
11.
|
Hao J, Pang YT and Wang DY: Diffuse
mucosal inflammation in nasal polyps and adjacent middle turbinate.
Otolaryngol Head Neck Surg. 134:267–275. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
12.
|
Morinaka S and Nakamura H: Inflammatory
cells in nasal mucosa and nasal polyps. Auris Nasus Larynx.
27:59–64. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
13.
|
Woodworth BA, Joseph K, Kaplan AP and
Schlosser RJ: Alterations in eotaxin, monocyte chemoattractant
protein-4, interleukin-5, and interleukin-13 after systemic steroid
treatment for nasal polyps. Otolaryngol Head Neck Surg.
131:585–589. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
14.
|
Villadangos JA and Schnorrer P: Intrinsic
and cooperative antigen-presenting functions of dendritic-cell
subsets in vivo. Nat Rev Immunol. 7:543–555. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
15.
|
Rissoan MC, Soumelis V, Kadowaki N, et al:
Reciprocal control of T helper cell and dendritic cell
differentiation. Science. 283:1183–1186. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
16.
|
Jahnsen FL, Lund-Johansen F, Dunne JF,
Farkas L, Haye R and Brandtzaeg P: Experimentally induced
recruitment of plasmacytoid (CD123high) dendritic cells in human
nasal allergy. J Immunol. 165:4062–4068. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
17.
|
Redecke V, Häcker H, Datta SK, et al:
Cutting edge: activation of Toll-like receptor 2 induces a Th2
immune response and promotes experimental asthma. J Immunol.
172:2739–2743. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
18.
|
Claeys S, de-Belder T, Holtappels G, et
al: Human beta-defensins and toll-like receptors in the upper
airway. Allergy. 58:748–753. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
19.
|
Kuchroo VK, Das MP, Brown JA, et al: B7-1
and B7-2 costimulatory molecules activate differentially the
Th1/Th2 developmental pathways: application to autoimmune disease
therapy. Cell. 80:707–718. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
20.
|
Kelsall BL, Stüber E, Neurath M and
Strober W: Interleukin-12 production by dendritic cells. The role
of CD40-CD40L interactions in Th1 T-cell responses. Ann NY Acad
Sci. 795:116–126. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
21.
|
Chen YS, Arab SF, Westhofen M and Lorenzen
J: Expression of interleukin-5, interleukin-8, and interleukin-10
mRNA in the osteomeatal complex in nasal polyposis. Am J Rhinol.
19:117–123. 2005.PubMed/NCBI
|
|
22.
|
Reider N, Reider D, Ebner S, et al:
Dendritic cells contribute to the development of atopy by an
insufficiency in IL-12 production. J Allergy Clin Immunol.
109:89–95. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
23.
|
Sánchez-Segura A, Brieva JA and Rodriguez
C: T lymphocytes that infiltrate nasal polyps have a specialized
phenotype and produce a mixed TH1/TH2 pattern of cytokines. J
Allergy Clin Immunol. 6:953–960. 1998.PubMed/NCBI
|
|
24.
|
Sokolovska A, Hem SL and HogenEsch H:
Activation of dendritic cells and induction of CD4(+) T cell
differentiation by aluminum-containing adjuvants. Vaccine.
25:4575–4585. 2007.
|
|
25.
|
Yi H, Zhang L, Zhen Y, He X and Zhao Y:
Dendritic cells induced in the presence of GM-CSF and IL-5.
Cytokine. 37:35–43. 2007. View Article : Google Scholar : PubMed/NCBI
|