Introduction
Gastric cancer is one of the most common types of
cancer in the world, and in 2011 ~70% of all cases of gastric
cancer occurred in developing countries (1). Metastasis is the most common cause of
mortality of patients with gastric cancer, and one of the most
common causes of mortality in patients who have undergone radical
resection of gastric cancer (2).
Tissue factor (TF), a 47-kDa transmembrane glycoprotein, primarily
initiates the coagulation cascade through binding to activated
factor VII (FVIIa). Previous studies have reported that TF is
highly expressed in several types of tumor, including cancers of
the pancreas, prostate, breast, colon and lung (3,4), and
TF is detectable on the surface of tumor cells and TF-bearing
microparticles (MPs) in the blood circulation which are shed from
the cell surface (5,6). Furthermore, the expression levels of
TF are correlated with tumor growth, angiogenesis and metastasis.
Previous studies have suggested that TF plays an important role in
the pulmonary metastasis of melanoma (7,8).
However, to the best of our knowledge, no study thus far has
reported the antitumor effects and antitumor mechanism of the
inhibition of TF expression by small interfering RNA (siRNA) in
gastric cancer.
In the present study, RNA interference (RNAi)
technology was applied to silence the expression of TF in the
SGC7901 gastric cancer cell line.
Materials and methods
Design and synthesis of specific
siRNA
The mRNA sequence of the TF gene was obtained
through the GenBank sequence search, then siRNA design software was
used to design four TF-targeted siRNA sequences and a negative
control siRNA sequence (Table I).
To avoid the formation of a termination signal, TTDAAGAGA was
inserted into the loop-stem structure of the short hairpin RNA
(shRNA) template and the T6 structure was used as the transcription
termination sequence. At the 5′ end of the sense strand template,
CACC or CACCG, and a BasI restriction site were added, and
at the 5′ end of the antisense strand template, GATC and a
BamHI restriction site were added.
 | Table IInterference fragment sequences of the
TF gene. |
Table I
Interference fragment sequences of the
TF gene.
| Target gene | Sense sequence | Antisense
sequence |
|---|
| TF-1 |
5′-CACCGCGCTTCAGGCACTAC
AAATATTCAAGAGATATTTGTA
GTGCCTGAAGCGCTTTTTTG-3′ |
5′-GATCCAAAAAAGCGCTTCAG
GCACTACAAATATCTCTTGAAT
ATTTGTAGTGCCTGAAGCGC-3′ |
| TF-2 |
5′-CACCGGGAGCCTCTGTATGA
GAACTTTCAAGAGAAGTTCTC
ATACAGAGGCTCCCTTTTTTG-3′ |
5′-GATCCAAAAAAGGGAGCCT
CTGTATGAGAACTTCTCTTGAA
AGTTCTCATACAGAGGCTCCC-3′ |
| TF-3 |
5′-CACCGGAACCCAAACCCGT
CAATCATTCAAGAGATGATTGA
CGGGTTTGGGTTCCTTTTTTG-3′ |
5′-GATCCAAAAAAGGAACCCA
AACCCGTCAATCATCTCTTGAA
TGATTGACGGGTTTGGGTTCC-3′ |
| TF-4 |
5′-CACCGAATGTGACCGTAGA
AGATGATTCAAGAGATCATCTT
CTACGGTCACATTCTTTTTTG-3′ |
5′-GATCCAAAAAAGAATGTGA
CCGTAGAAGATGATCTCTTGAA
TCATCTTCTACGGTCACATTC-3′ |
| Negative |
5′-CACCGTTCTCCGAACGTGT
CACGTCAAGAGATTACGTGAC
ACGTTCGGAGAA TTTTTT |
5′-GATCCAAAAAATTCTCCGA
ACGTGTCACGTAATCTCTTGAC
G-3′ GTGACACGTTCGGAGAAC-3′ |
Construction and connection of
pGPU6/GFP/Neo-TF expression vector
The DNA oligonucleotide was dissolved with Tris-EDTA
buffer (pH 8.0), and the oligonucleotide solutions of the
corresponding sense and antisense strands were added. The
pGPU6/GFP/Neo vector (2 μg) was subjected to agarose gel
electrophoresis and recycled with a Takara DNA Purification kit,
version 2.0 (Takara Bio Inc., Otsu, Shiga, Japan). The
concentration was then diluted to 50 ng/μl for connection with the
pGPU6/GFP/Neo-shRNA vector (Shanghai GenePharma Co., Ltd.,
Shanghai, China), and the connected product was transformed into
Escherichia coli DH5α.
Extraction and identification of positive
clones
A monoclonal bacterial colony was selected,
inoculated in 5 ml LB resistant medium and incubated in a 37°C
incubation shaker overnight (4.44 × g). The plasmid was extracted
with a plasmid extraction kit [Sangon Biotech (Shanghai) Co., Ltd.,
Shanghai, China]. BglII was used for mono-enzyme digestion
to identify whether the targeted DNA fragment had been correctly
connected to the pGPU6/GFP/Neo vector. Two recombinant plasmids,
with the targeted fragment confirmed by mono-enzyme digestion, were
randomly selected and sent to Sangon Biotech (Shanghai) Co., Ltd.
for detection of the sequences, and the concentration and purity
detection was performed on 1 μl plasmid with an automatic
spectrophotometer.
Cell culture and transfection
The SGC7901 gastric cancer cell line (stored in our
laboratory) was reanimated and placed in 10% fetal bovine serum
medium for cultivation and passage in a 37°C and 5% CO2
incubator. The cells in the logarithmic growth phase were obtained,
trypsinized, vaccinated and cultivated in G418 complete medium,
with the final concentrations of G418 as 800, 700, 600, 500, 400,
300 and 0 μg/ml to determine the minimum lethal concentration of
G418. The cells were seeded in a six-well plate and cultured to
90–95% confluence, then Opti-MEM (Gibco-BRL, Carlsbad, CA, USA) was
used to wash the cells twice. The lipofection method was used to
transfect the recombinant and empty plasmids into the cells. An
inverted fluorescence microscope was used to observe the results of
transfection. RPMI-1640 medium was then added to the complete
medium with the screened G418 concentration following the
cultivation passage, and after one week, the G418 complete medium
was changed to maintain the concentration at 600 μg/ml. When the
control cells were all dead and the colonies were observable with
the naked eye, the colonies were selected for detection.
Reverse transcription-polymerase chain
reaction (RT-PCR)
Following extraction, the total RNA was then
reverse-transcribed into cDNA for RT-PCR. The primer sequences of
GAPDH were as follows: Upstream, 5′-GCACCGTCAAGGCTGAGAAC-3′; and
downstream, 5′-TGGTGAAGACGCCAGTGGA-3′; with the amplified fragment
as 138 bp. The primer sequences of the TF gene were: Upstream,
5′-CTCCCGAACAGTTAACCGGAAG-3′; and downstream,
5′-GCCAGGATGATGACAAGGATGA-3′; with the amplified fragment as 136
bp. The PCR reaction conditions were as follows: 50 cycles, each
with denaturation at 95°C for 30 sec, 95°C for 5 sec, 60°C for 34
sec, and extension at 95°C for 15 sec. Agarose gel electrophoresis
(1.5%) was performed and the integrated optical density was
obtained with an image analysis system (Bio-Rad, Philadelphia, PA,
USA) for the semi-quantitative analysis.
CCK-8 (Cell Counting Kit-8) determination
of cell proliferation
Following conventional digestion, the cells in the
logarithmic growth phase were used to prepare a 1×105
cells/ml single cell suspension with 10% fetal bovine serum medium.
The experiment was divided into the blank control group, the
negative control group and the pGPU6/GFP/Neo/TF group, with four
repeated wells for each group, and the final volume of the culture
medium of each well was 200 μl. CCK-8 solution (10 μl; Beyotime
Institute of Biotechnology, Shanghai, China) was added to each well
after 24, 48 and 72 h of transfection and the cultivation was
terminated after 2 h at 37°C. The detection wavelength was selected
as 450 nm in the ELISA to measure the absorbance of each well, the
results were recorded and the growth curve was drawn according to
the time-absorbance value of each group to compare the difference
between the proliferative capacity of the three groups.
Chemotherapeutic drug resistance
testing
Following conventional digestion, the cells in the
logarithmic growth phase were used to prepare a 1×105
cells/ml single cell suspension with 10% fetal bovine serum medium,
and each well of a 96-well plate was inoculated with 100 μl. The
chemotherapy drug oxaliplatin (Jiangsu Hengrui Medicine Co., Ltd.,
Jiangsu, China) was diluted in autoclaved saline and configured
into the different working concentrations of 6.25, 12.5, 25, 50 and
100 μg/ml. Four repeated wells were set up for each group, with a
final volume of 200 μl for each well, then incubated for 48 h prior
to the addition of the different concentrations of oxaliplatin to
each well for a further 48-h incubation. CCK-8 (10 μl) was added to
each well and incubated 2 h prior to the termination of incubation.
A wavelength of 450 nm was selected for the ELISA assay, and the
absorbance values of each well were measured. The results recorded
were used to calculate the growth inhibition rates of oxaliplatin
towards the cells of each group. Inhibition (%) = (1- medication
group/control group) × 100%.
Transwell experiments
Matrigel gum (2 mg/ml; 15 μl) was added to each well
of the Transwell chamber, and the gum was spread on the surface of
the bottom membrane of the Transwell chamber. The base membrane was
hydrated for the preparation of the single cell suspension.
RPMI-1640 cell medium, containing 10% fetal bovine serum, was added
to a 24-well plate, with 600 μl in each well. The cell suspension
was added into the Transwell chamber, with 100 μl in each well, and
each well was repeated in three wells. The chamber was immersed in
the complete medium of the 24-well plate and incubated at 37°C and
5% CO2 for 24 h. The chamber was removed, rinsed with
phosphate-buffered saline (PBS), fixed with methanol for 10 min,
and then washed with PBS. Crystal violet staining was performed on
the chamber, and then it was rinsed and dried. The number of nuclei
on the bottom surface of the polyester film of the Transwell
chamber was detected under a microscope at high magnification to
determine the degrees of cellular invasion. Four view fields of the
high magnification were randomly selected to count the number of
nuclei and then the average number was calculated. The experiment
was repeated three times.
Wound-healing assay
The cells of each group were seeded in a
fibronectin-pre-custodite 24-well plate, with six wells for each
group and 4×105 cells/well. The cells were cultured
overnight to form cell monolayers, then a pipette was used to draw
a dash-shaped horizontal wound along the bottom of the plate.
Serum-free Dulbecco’s modified Eagle’s medium was used for
cultivation. The cell motilities were observed at 0 and 24 h, and
the distance from the starting point to the farthest migrated
nucleus was calculated, with the result expressed as the mobility
degree: The percentage of migration width to the original width.
The experiment was repeated five times.
Detection of apoptosis levels
The cells were transferred into a Falcon tube and
rinsed with the pre-cold 1X PBS buffer twice (111 × g for 5 min).
Subsequently, a 5×105 cells/ml cell suspension was
prepared with 1X binding buffer and then 5 μl Annexin V-fluorescein
isothiocyanate was added. After 15 min, 10 μl propidium iodide was
added and the suspension was incubated in the dark at room
temperature for 15 min. Binding buffer (1X; 400 μl) was added to
the suspension and it was gently vortexed. Flow cytometry was used
to detect the results 1 h later.
Statistical analysis
SPSS software package, version 17.0 (SPSS, Inc.,
Chicago, IL, USA) was used for the statistical analysis. The
measured data are expressed as the mean ± standard deviation.
One-way analysis of variance and intergroup Student-Newman-Keuls
test were performed for the statistical analysis. P<0.05 was
considered to indicate a statistically significant difference.
Results
Identification of the recombinant plasmid
pGPU6/GFP/Neo/TF
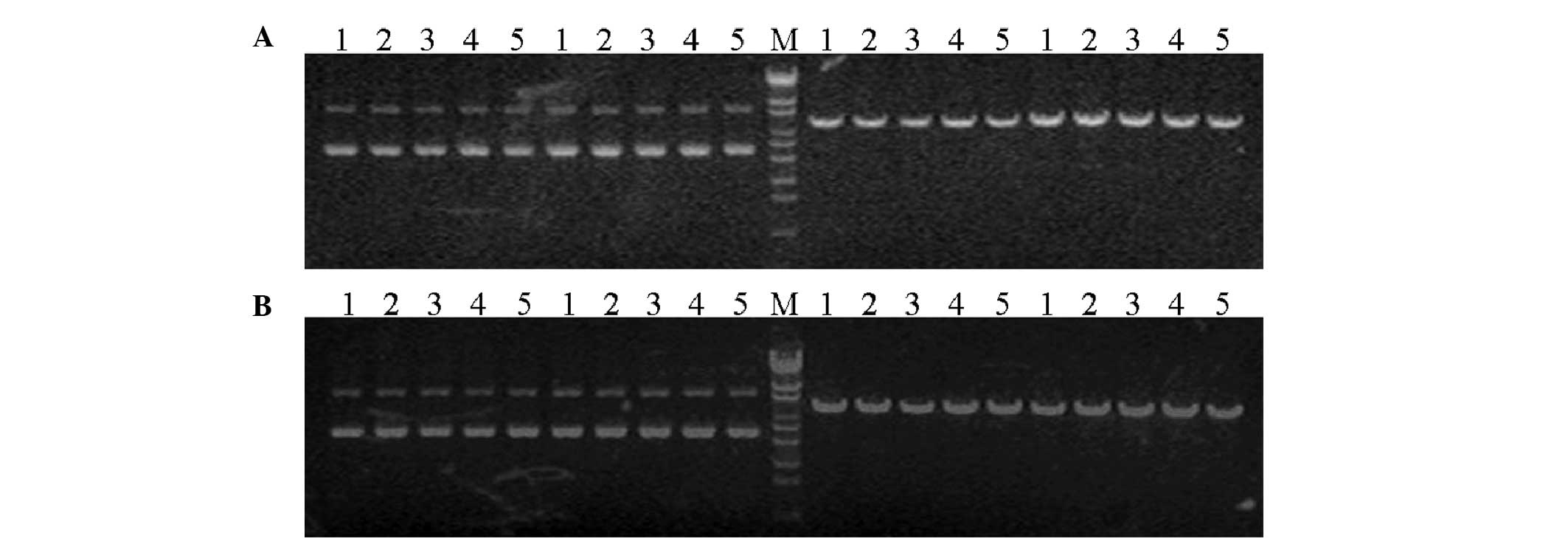
The enzyme digestion results showed that the
recombinant plasmid contained the BamHI digestion site
instead of the PstI digestion site, while the empty vector
contained the two sites. Therefore when using the restrictive
endonucleases BamHI and PstI for the digestion, the
restructured plasmid was digested with BamHI and appeared in
the linear form, while when digested by PstI, no change was
identified. The empty vector was digested by the two enzymes
(BamHI and PstI) and appeared in the linear form,
indicating that all digested plasmids were the positive recombinant
vectors. Liquid (20 μl) of each plasmid was obtained and delivered
to the Shanghai Invitrogen Biotechnology Co., Ltd. (Shanghai,
China) for sequence identification. The sequencing results showed
that the synthesized plasmid sequences were correct and
successfully cloned into the selected vector, and no presence of
abnormal bases was detected (Figs.
1 and 2).
Plasmid transfection and screening
A UV spectrophotometer was used to detect the
concentration and purity of the extracted plasmid DNA, and the
results were satisfactory for the subsequent transfection step. The
medium, with the final concentration of G418 as 800, 700, 600, 500,
400, 300 and 0 μg/ml, was used for the cultivation, and the
screening concentration of SGC7901 was determined as 600 μg/ml and
the maintaining concentration as 300 μg/ml. During the stable
transfection stage, cell death was observed three days after the
addition of the screening concentration, appearing as cell
contraction, rounding, floating and gathering into groups. On the
8th day, the majority of cells were observed as dead, and a small
number of transfected cells remained. When switched to the medium
with the maintaining concentration, these transfected cells
restored growth and gradually formed positive clones. The expanded
resistant clones were transferred to a new culture flask on
approximately the 3rd week, and were cultured until the cells grew
to 70–80% confluence. Following sub-cultivation, the cells became
stably-transfected cells containing the hairpin-siRNA expression
plasmid.
Effect of time towards the cellular
transfection rate
An inverted fluorescence microscope was used to
observe the expression of the green fluorescent protein following
transfection. With 4 μg DNA, the transfection rates of the SGC7901
cells [transfection rate (%) = fluorescent cells under the same
view field/all cells under the same view field] at 24, 48 and 72 h
were 25, 40 and 30%, respectively, and the transfection rate at 48
h was significantly higher than that at 24 h, while the rate
decreased at 72 h.
RT-PCR
RT-PCR was used to detect the expression levels of
the TF gene, and the results showed that, compared with those of
the control and negative control groups, the transfected expression
levels of pGPU6/GFP/Neo/TF in each group decreased (P<0.05 or
P<0.01). The expression levels of the TF-2 gene were the lowest,
and the inhibitory effect was the most marked (P<0.01).
Therefore, in subsequent experiments this was considered the
experimental group. No significant difference in the expression
levels of the TF gene was observed between the blank control and
negative control groups (P>0.05; Table II).
| Table IIExpression levels of the TF gene in
SGC7901 gastric cancer cells. |
Table II
Expression levels of the TF gene in
SGC7901 gastric cancer cells.
| Grouping | GAPDH Ct | TF Ct | ΔCt | ΔΔCt |
2−ΔΔCt |
|---|
| N | 32.95±0.55 | 23.87±0.04 | −9.08±0.39 | 0 | 1 |
| NC | 32.77±0.03 | 23.85±0.32 | −8.92±0.23 | 0.15717 | 0.864346 |
| TF-1 | 31.86±1.08 | 26.13±0.21 | −5.73±0.78 | 3.343803 | 0.483297a |
| TF-2 | 33.37±0.17 | 26.17±0.47 | −7.2±0.37 | 1.875092 | 0.095393b |
| TF-3 | 32.34±0.82 | 24.25±0.43 | −8.08±0.66 | 0.996823 | 0.310134a |
| TF-4 | 32.59±0.64 | 26.90±0.07 | −5.69±0.45 | 3.389969 | 0.27261a |
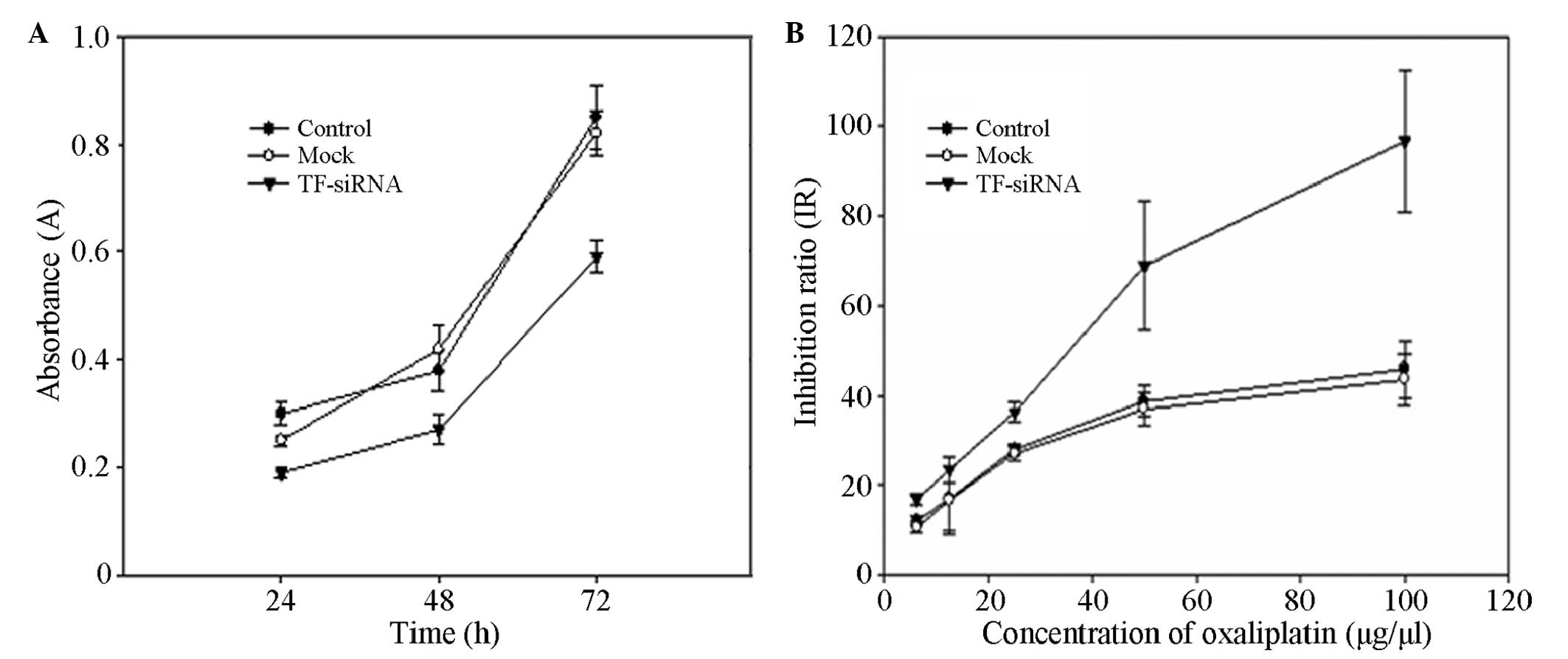
Inhibitory effect of TF-siRNA towards
cell proliferation
The CCK-8 method was performed to detect the
absorbance value A (the value is positively correlated with the
number of viable cells) of each well of the blank control group,
the negative control group and the experimental group at 24, 48 and
72 h. It was demonstrated that the levels of cell proliferation of
the experimental group were significantly lower than those of the
control group (P<0.05), while no significant difference between
the blank control and negative control groups was identified
(Fig. 3A).
TF-siRNA reduces the chemotherapeutic
drug resistance
A CCK-8 assay was used to detect the absorbance
value of each group following the addition of oxaliplatin, and the
inhibition rate was calculated. The results showed that the
inhibition rate of oxaliplatin towards the
pGPU6/GFP/Neo/TF-transfected cells was significantly higher than
those of the control groups (P<0.05; Fig. 3B).
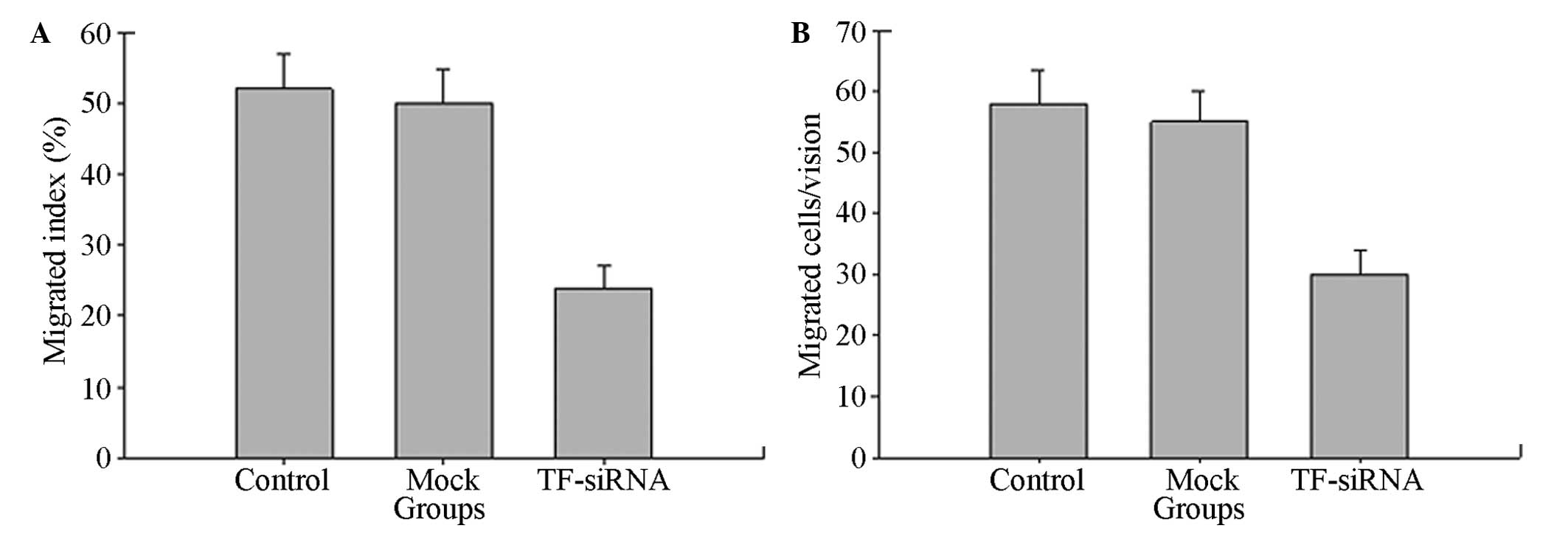
Inhibition of TF-siRNA towards the cell
invasion ability
To confirm the effect of TF-siRNA towards the
migration and invasion of the tumor cells, a wound-healing assay
and cell invasion experiment were performed on the SGC7901 gastric
cancer cell line. As shown in Fig.
4, the wound-healing assay indicated that, compared with those
of the negative control and blank control groups, the migration
levels of the TF-siRNA group were significantly reduced
(P<0.05). The results of the Transwell experiment showed that
the cells of the three groups all passed through the Matrigel gel
membrane. Compared with that of the negative control and blank
control groups, the number of cells of the TF-siRNA group which
penetrated the Matrigel gel membrane was significantly fewer
(P<0.05), resulting in the significantly reduced invasiveness.
These results suggested that TF-siRNA reduces the metastatic
potential of gastric cancer cells.
TF-siRNA increases the apoptosis of
gastric cancer cells
To assess the induction towards apoptosis following
the TF gene knockdown of SGC7901 gastric cancer cells, the
double-staining method was performed to detect apoptosis 48 h after
the transfection. The results showed that the percentage of
apoptotic cells of the TF-siRNA group was 18.35%, significantly
higher than that of the negative control (3.58%) and blank control
(2.35%) groups (P<0.05), indicating that specific TF-siRNA may
induce the apoptosis of tumor cells.
Discussion
In normal tissue cells, the expression levels of TF
are low or almost negative, but high expression levels of TF are
detectable in a variety of types of solid malignant tumor. Previous
studies have found that TF shows abnormal expression levels in
various types of tumor cell, including gastric, colon and
pancreatic cancer cells (9,10),
and that its expression levels are closely associated with
biological behaviors of tumor cells, including growth, invasion and
metastasis, which were confirmed by the present study. In
vitro and in vivo functional studies (11–13)
have shown that TF significantly promotes tumor angiogenesis and
enhances the invasion and metastasis abilities of tumor cells. In a
previous study, a recombinant vector with the pcDNA3.1/TFcDNA
plasmid was transfected into SGC7901 human gastric cancer cells,
and the endogenous increased expression levels of TF significantly
increased the levels of invasion and metastasis of the cells, and
thus confirmed that TF was closely associated with the processes of
invasion and metastasis of cancer (14). To further study the effects of the
TF gene in the biological behaviors of gastric cancer, based on the
high expression levels of TF in tumor cells and that siRNA
efficiently inhibits the expression of the purpose gene, siRNA,
which specifically encoded the TF gene, was designed, constructed,
and transfected into SGC7901 gastric cancer cells to observe the
siRNA-inhibited TF mRNA expression levels and function in the
present study. It was demonstrated that the specifically targeted
TF-siRNA effectively silenced the expression of the TF gene in
SGC7901 human gastric cancer cells. The aforementioned method was
used to study the changes in the biological behaviors of gastric
cancer cells following TF knockdown. The expression levels of TF
were correlated with the proliferative growth of the tumor cells
(15). Through the CKK-8 method,
it was observed that the growth of the gastric cancer cells was
inhibited following the transfection of TF-siRNA in vitro,
the inhibition was strongest after 48-h transfection, with an
inhibition rate of 37%, and the cell proliferation ability
significantly decreased. Following the chemotherapy of oxaliplatin,
the inhibition rate of the gastric cancer cells significantly
increased, indicating that following silencing of the TF gene, the
growth of the gastric cancer cells was more likely to be suppressed
by chemotherapy. Migration and invasion of tumor cells are the
important steps in the process of tumor metastasis (16). The wound-healing assay and cell
invasiveness test in the present study confirmed that TF-siRNA
significantly reduced the potential migration and invasion effects
of SGC7901 gastric cancer cells. These results suggested that TF is
involved in coagulation and angiogenesis and plays an important
role in the growth, invasion and metastasis of gastric cancer
cells.
The effects of TF on tumor growth, invasion and
metastasis are the joint action of its coagulation and
non-coagulation pathways. Tumor cells themselves produce TF and
stimulate the body to produce interleukin-1 (IL-1) and vascular
endothelial growth factor (VEGF) (15). These procoagulant substances
produce a hypercoagulable state within patients with tumors when
they are exposed to endothelial cells and platelets in vivo
(17). It has been reported that
(18) the hypercoagulable state of
patients with malignant tumors may depend on the TF-positive MPs in
the circulation, and the MPs of different organizations are
involved in the pathogenesis of tumor thrombus. In tumor-associated
vascular diseases, the expression levels of TF in the circulation
increase. For example, in human colon cancer, the TF in the
circulation and on the membrane surface is associated with the
occurrence of tumor cells and blood vessels (13). Tesselaar et al (19) also demonstrated that MP-associated
TF was involved in the development of tumor-associated thrombosis
in various types of adenocarcinoma.
Compared with those of the coagulation pathway, the
functions of the TFs in the non-coagulation pathway are more
important towards tumor invasion and metastasis. TF may promote
tumor invasion and metastasis through intracellular signal
transduction, regulating tumor angiogenesis and degrading the
extracellular matrix (ECM), among other functions. In a previous
study (20) it was shown that
TF-FVII activates the signaling pathways of p44/42
mitogen-activated protein kinase (MAPK) and phosphatidylinositol 3
by activating protease activated receptor 2 (PAR2). TF-FVII may
hydrolyze prothrombin, therefore hydrolyzing and activating the
PARs, and then increasing the levels of cytokines, including IL-1β
and IL-8 (21),. These factors may
act as the messengers that regulate inflammation, and also
stimulate the invasion and metastasis of tumor cells. The
combination of TF and FVII may activate tyrosine kinase in the TF
cell area, causing a Ca2+ influx, and then increase the
transcription levels of VEGF through the MAPK or PKC signaling
pathways. Numerous types of tumor have the activities of
plasminogen activators, which is mainly caused by the increased
activities of tissue plasminogen activator (PA) (22). The combination of TF and FVII was
found to upregulate the expression levels of the urokinase PA in
SW979 human pancreatic cancer cells, and it was thought that TF may
increase the invasion and metastasis of tumor cells through
upregulation the expression levels of plasminogen activator
receptors (23). The plasmin
activated by PA has high levels of proteolytic activity, which may
degrade the ECM and thus promote tumor invasion and metastasis
(24). Tumor cells not only
secrete metalloproteinases (MMPs) by themselves, but also promote
vascular endothelial cells to produce MMPs through secreting VEGF.
They work together to degrade the basement membrane and ECM, and to
stimulate the formation of new blood vessels (25). These pathways all promote the
growth, invasion and metastasis of tumor cells.
Apoptosis inhibition is an important mechanism of
tumorigenesis. The double staining method was used in the present
study to detect the levels of apoptotic cells in each group, and
demonstrated that the reduced expression levels of TF induced the
apoptosis of SGC7901 gastric cancer cells. A study has shown that
TF-FVII activates PI3K/Akt signal transduction to inhibit the
doxorubicin-induced apoptosis in glioblastoma cells (26). Jiang, Guo and Bromberg (27) demonstrated that the complex of
TF-FVII-factor X inhibits the apoptosis of breast cancer cells via
the p44/42 MAPK and PKB/Akt signaling pathways. Whether or not its
mechanism of apoptosis induction and its long-term silencing would
lead to changes in the levels of anti-apoptotic factors requires
further studies.
Acknowledgements
This study was supported by the National Natural
Science Foundation of China (grant no. 31270532) and Gansu province
natural science fund research project (grant no. 1107RJZA250).
References
|
1
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:68–90. 2011. View Article : Google Scholar
|
|
2
|
Roukos DH and Kappas AM: Limitations in
controlling risk for recurrence after curative surgery for advanced
gastric cancer are now well-explained by molecular-based
mechanisms. Ann Surg Oncol. 8:620–621. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Callander NS, Varki N and Rao LV:
Immunohistochemical identification of tissue factor in solid
tumors. Cancer. 70:1194–1201. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Regina S, Rollin J, Bléchet C, Iochmann S,
Reverdiau P and Gruel Y: Tissue factor expression in non-small cell
lung cancer: relationship with vascular endothelial growth factor
expression, microvascular density, and K-ras mutation. J Thorac
Oncol. 3:689–697. 2008. View Article : Google Scholar
|
|
5
|
Aharon A and Brenner B: Microparticles,
thrombosis and cancer. Best Pract Res Clin Haematol. 22:61–69.
2009. View Article : Google Scholar
|
|
6
|
Zwicker JI: Predictive value of tissue
factor bearing microparticles in cancer associated thrombosis.
Thromb Res. 125(Suppl 2): S89–S91. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Amarzguioui M, Peng Q, Wiiger MT, et al:
Ex vivo and in vivo delivery of anti-tissue factor short
interfering RNA inhibits mouse pulmonary metastasis of B16 melanoma
cells. Clin Cancer Res. 12:4055–4061. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wang X, Wang M, Amarzguioui M, Liu F,
Fodstad Ø and Prydz H: Downregulation of tissue factor by RNA
interference in human melanoma LOX-L cells reduces pulmonary
metastasis in nude mice. Int J Cancer. 112:994–1002. 2004.
View Article : Google Scholar
|
|
9
|
Nitori N, Ino Y, Nakanishi Y, et al:
Prognostic significance of tissue factor in pancreatic ductal
adenocarcinoma. Clin Cancer Res. 11:2531–2539. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yamashita H, Kitayama J, Ishikawa M and
Nagawa H: Tissue factor expression is a clinical indicator of
lymphatic metastasis and poor prognosis in gastric cancer with
intestinal phenotype. J Surg Oncol. 95:324–331. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Xin H, Zheng YJ and Han ZG: Effect of
tissue factor on intravascular migration of tumor cells. Chinese
Journal of Pathophysiology. 20:960–963. 2004.(In Chinese).
|
|
12
|
Yu YJ, Ma HT, Cao N, Jiao ZY and Li YM:
Tissue factor influence on invasion and metastasis of human gastric
cancer cells line. Chinese Journal of Bases and Clinics in General
Surgery. 19:942–946. 2012.(In Chinese).
|
|
13
|
Yu YJ, Li YM, Hou XD, Guo C, Cao N and
Jiao ZY: Effect of tissue factor on invasion inhibition and
apoptosis inducing effect of oxaliplatin in human gastric cancer
cell. Asian Pac J Cancer Prev. 13:1845–1849. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Rickles FR, Shoji M and Abe K: The role of
the hemostatic system in tumor growth, metastasis, and
angiogenesis: tissue factor is a bifunctional molecule capable of
inducing both fibrin deposition and angiogenesis in cancer. Int J
Hematol. 73:145–150. 2001. View Article : Google Scholar
|
|
15
|
Chambers AF, Groom AC and MacDonald IC:
Dissemination and growth of cancer cells in metastatic sites. Nat
Rev Cancer. 2:563–572. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
16
|
Rickles FR and Falanga A: Molecular basis
for the relationship between thrombosis and cancer. Thromb Res.
102:V215–V224. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Davila M, Amirkhosravi A, Coll E, et al:
Tissue factor-bearing microparticles derived from tumor cells:
impact on coagulation activation. J Thromb Haemost. 6:1517–1524.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yu JL, May L, Lhotak V, et al: Oncogenic
events regulate tissue factor expression in colorectal cancer
cells: implications for tumor progression and angiogenesis. Blood.
105:1734–1741. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Tesselaar ME, Romijn FP, Van Der Linden
IK, Prins FA, Bertina RM and Osanto S: Microparticle-associated
tissue factor activity: a link between cancer and thrombosis? J
Thromb Haemost. 5:520–527. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Rao LV and Pendurthi UR: Tissue
factor-factor VIIa signaling. Arterioscler Thromb Vasc Bio1.
25:47–56. 2005.PubMed/NCBI
|
|
21
|
Hjortoe GM, Petersen LC, Albrektsen T, et
al: Tissue factor-factor VIIa-specific up-regulation of IL-8
expression in MDA-MB-231 cells is mediated by PAR-2 and results in
increased cell migration. Blood. 103:3029–3037. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Schmitt M, Janicke F, Moniwa N,
Chucholowski N, Pache L and Graeff H: Tumor-associated
urokinase-type plasminogen activator: biological and clinical
significance. Biol Chem Hoppe Seyler. 373:611–622. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Taniguchi T, Kakkar AK, Tuddenham EG,
Williamson RC and Lemoine NR: Enhanced expression of urokinase
receptor induced through the tissue factor-factor VIIa pathway in
human pancreatic cancer. Cancer Res. 58:4461–4467. 1998.PubMed/NCBI
|
|
24
|
Francis JL, Biggerstaff J and Amirkhosravi
A: Hemostasis and malignancy. Semin Thromb Hemost. 24:93–109. 1998.
View Article : Google Scholar
|
|
25
|
Zucker S, Mirza H, Conner CE, et al:
Vascular endothelial growth factor induces tissue factor and matrix
metalloproteinase production in endothelial cells: conversion of
prothrombin to thrombin results in progelatinase A activation and
cell proliferation. Int J Cancer. 75:780–786. 1998. View Article : Google Scholar
|
|
26
|
Tang H, Fang J, Shu K, et al: Tissue
factor/FVII regulates doxorubicin-induced apoptosis in glioblastoma
via activating PI3K/Akt signaling. The Chinese-German Journal of
Clinical Oncology. 6:487–491. 2007. View Article : Google Scholar
|
|
27
|
Jiang X, Guo YL and Bromberg ME: Formation
of tissue factor-factor VIIa-factor Xa complex prevents apoptosis
in human breast cancer cells. Thromb Haemost. 96:196–201.
2006.PubMed/NCBI
|