Introduction
Genistein (4′,5,7-trihydroxyisoflavone) is a major
isoflavone present in plants, such as soybean, and therefore, in a
variety of human foods (1–3). In 1987, it was discovered that
genistein is a potent inhibitor of the tyrosine-specific protein
kinase activity of the epidermal growth factor receptor (4). Since then, numerous researchers have
studied the possible use of genistein as a cancer chemopreventive
agent based on the key role of protein tyrosine kinase inhibitors
in cancer cell growth and apoptosis (5,6). In
support of these studies, several epidemiological reports have
revealed significant correlations between genistein consumption and
a reduced risk of breast cancer (7–9).
Furthermore, a number of studies have demonstrated that genistein
exhibits significant anticancer activity against breast tumors
in vitro and in vivo (10,11).
Breast cancer belongs to a group of heterogeneous
diseases with multiple clinical, molecular and histopathological
forms, which makes achieving effective chemotherapy problematic
(12). To develop breast cancer
therapies, the targeting of estrogen receptor-α (ERα), which is
expressed in ~70% of breast cancers and which makes it difficult to
obtain a response to cancer drug treatment (13,14),
requires consideration.
Therefore, the aim of the present study was to
investigate the proliferative effects and induction of apoptosis by
genistein via ERα-related pathways in MCF-7 human breast cancer
cells and 3T3-L1 mouse preadipocytes.
Materials and methods
Reagents
All reagents and plasticware used for cell culture,
including fetal bovine serum (FBS), media and antibiotics, were
purchased from Invitrogen Life Technologies (Carlsbad, CA, USA) and
Corning Incorporated Life Sciences (Corning, NY, USA). Insulin,
dexamethasone, 3-isobutyl-1-methylxanthine (IBMX) and
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)
were purchased from Sigma-Aldrich Co. (St. Louis, MO, USA). The
protein analysis reagent and antibodies were purchased from Bio-Rad
Laboratories, Inc. (Hercules, CA, USA) and Santa Cruz
Biotechnology, Inc. (Santa Cruz, CA, USA), respectively. Genistein
was purchased from LC Laboratories (Woburn, MA, USA) and dissolved
in dimethyl sulfoxide (DMSO; final concentration of 0.1% in
medium).
Cell culture
The MCF-7 human breast cancer cells and 3T3-L1 mouse
preadipocytes were purchased from the Korean Cell Line Bank (Seoul,
South Korea) and American Type Culture Collection (Manassas, VA,
USA), respectively, for use in the present study. The cells were
maintained in Roswell Park Memorial Institute (RPMI)-1640 medium or
Dulbecco’s modified Eagle’s medium (DMEM) supplemented with 10% FBS
and antibiotics (50 U/ml penicillin and 50 μg/ml streptomycin) at
37°C in a humidified atmosphere containing 5% CO2. Two
days subsequent to reaching confluence (designated as day 0), DMEM
containing 10% FBS and differentiation inducers (10 μg/ml insulin,
0.5 μM dexamethasone and 0.5 mM IBMX) were added to the 3T3-L1
cells to induce differentiation.
MCF-7 cell proliferation assay
MCF-7 cell proliferation was examined using MTT
assays. Cells were plated at 2.5–5×105 cells/well in a
96-well tissue culture plate and incubated for 24 h following which
they were exposed to genistein solutions at concentrations of 50,
100, 150 and 200 μM. Following incubation for 24, 48 and 72 h, the
plated cells were incubated with MTT (final concentration 0.5
mg/ml; Sigma-Aldrich) for 4 h at 37°C. The medium was discarded
from the plates and 100 μl DMSO was added to each well. The plates
were incubated for 5 min at room temperature whilst being shaken,
so that the complete dissolution of formazan was achieved. The
absorbance of MTT formazan was determined at 540 nm using an
ultraviolet-visible (UV/VIS) spectrophotometric plate reader (EMax;
Molecular Devices, LLC, Sunnyvale, CA, USA).
Cytotoxicity assay using 3T3-L1
cells
Cellular toxicity was measured in 3T3-L1
preadipocytes using MTT and LDH assays with various concentrations
of genistein (5–100 μM) for 24, 48 and 72 h. To measure lactate
dehydrogenase (LDH) release, 100 μl/well supernatant medium was
transferred to the corresponding well of an optically clear 96-well
flat-bottom microtiter plate and analyzed using an LDH cytotoxicity
detection kit (Takara Bio, Inc., Otsu, Japan).
Apoptosis detection
Apoptotic morphological changes were identified by
the 4′,6-diamidino-2-phenyl-indole (DAPI) staining of MCF-7 cells
and differentiating 3T3-L1 cells, which had been treated with
genistein at 50 μM for 48 h two days subsequent to reaching
confluence. Each cell line was seeded on poly-L-lysine-coated
slides and fixed with 4% methanol-free formaldehyde for 30 min.
Mounting medium containing DAPI was dispersed over the entire
slide. The mounted slides were stored at 4°C in the dark. Each
slide was observed under an LSM700 laser scanning microscope
equipped with Zen 2011 software (Carl Zeiss Microscopy GmbH, Jena,
Germany).
Immunoblotting
Following the exposure of MCF-7 and differentiating
3T3-L1 cells to genistein, each group of cells was subjected to
lysis in radio-immunoprecipitation assay (RIPA) buffer [1% nonyl
phenoxypolyethoxylethanol (NP)-40, 150 mM NaCl, 0.05% deoxycholic
acid (DOC), 1% sodium dodecyl sulfate (SDS) and 50 mM
tris(hydroxymethyl)aminomethane (Tris), pH 7.5] containing protease
inhibitors for 1 h at 4°C. The supernatant was separated by
centrifugation and the protein concentration was determined using a
Bradford protein assay kit 2 (Bio-Rad). The proteins were then
transferred to nitrocellulose membranes (0.45 μm). The membranes
were blocked with 1% bovine serum albumin (BSA) for 1.5 h, washed
twice with phosphate-buffered saline (PBS) containing 0.2%
Tween-20, and incubated with the respective primary antibodies
[cyclin D1, anti-ERα, -B-cell lymphoma 2 (-Bcl-2),
-Bcl-2-associated X protein (-Bax) and -β-actin; Santa Cruz
Biotechnology, Inc.] overnight at 4°C. The next day, the
immunoreaction was continued using secondary rabbit anti-rabbit
horseradish-peroxidase-conjugated antibodies following washing for
2 h at room temperature. Bands were detected with MicroChemi (DNR
Bio-Imaging System, Ltd., Jerusalem, Israel) using
WesternBright™ ECL solution (Advansta Inc., Menlo Park,
CA, USA).
Statistical analyses
All values are expressed as means ± standard
deviations. Data were analyzed by an unpaired Student’s t-test or
one-way analysis of variance followed by Dunnett’s multiple
comparison test (Sigma Stat software; Jandel Scientific Software,
San Rafael, CA, USA). For all comparisons, P<0.05 was considered
to indicate a statistically significant difference.
Results
Antiproliferative activity of genistein
toward MCF-7 cells
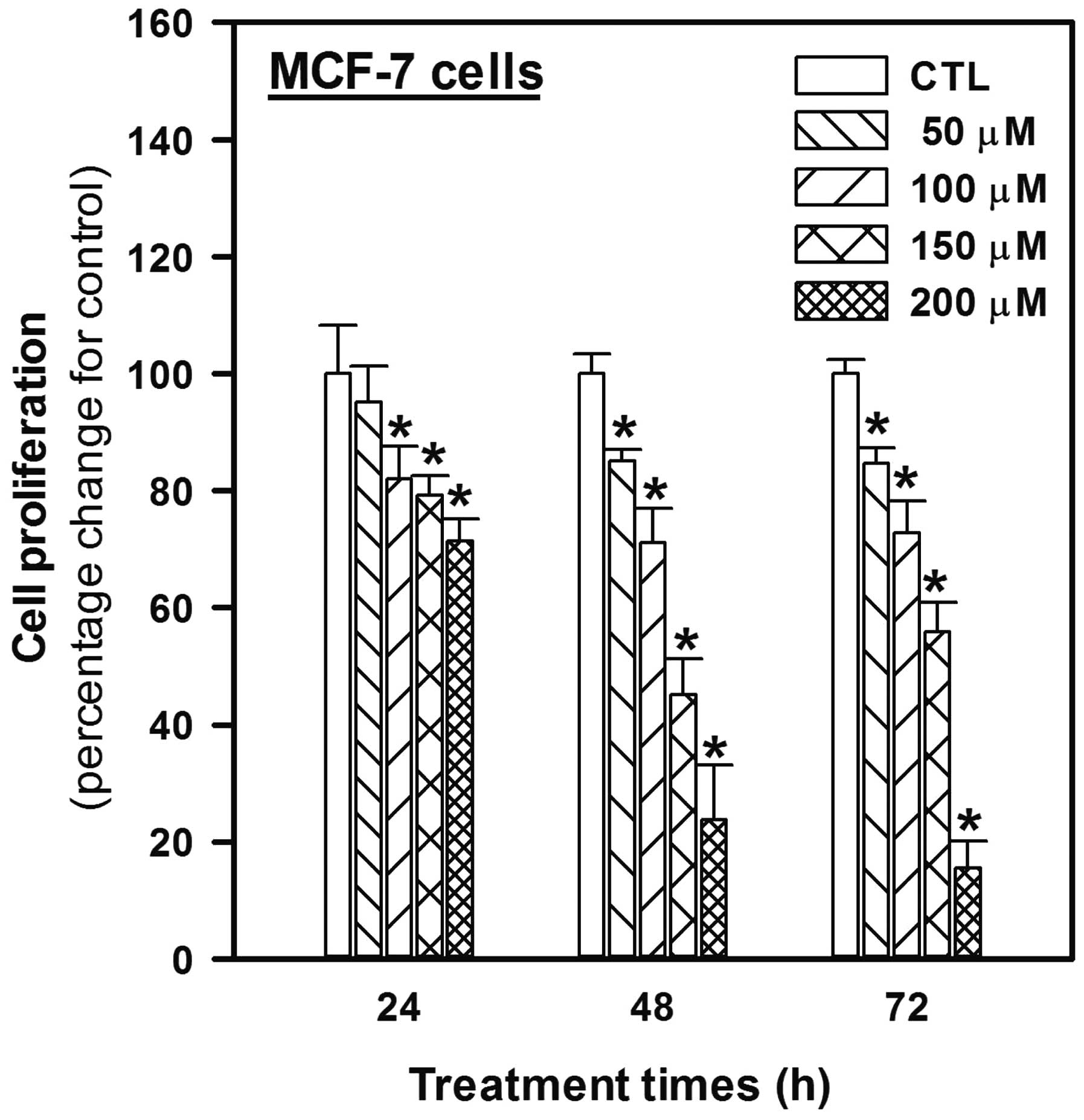
To investigate the possible anticancer effects of
the phytochemical genistein on MCF-7 human breast cancer cells, the
present study initially examined the antiproliferative effects of
genistein on MCF-7 cells using various concentrations (50, 100, 150
and 200 μM) of genistein for 24, 48 and 72 h (Fig. 1). Genistein inhibited the growth of
MCF-7 cells in a concentration-dependent manner and revealed
significant antiproliferative activity under all treatment
conditions, with the exception of 50 μM for 24 h. The
antiproliferative activity of genistein after 48 and 72 h was
stronger than that after 24 h, but there were no differences
between 48 and 72 h; reductions of 13, 29, 55 and 77% at 48 h and
15, 27, 45 and 85% at 72 h for 50, 100, 150 and 200 μM genistein,
were noted, as compared with the control levels after 48 and 72
h.
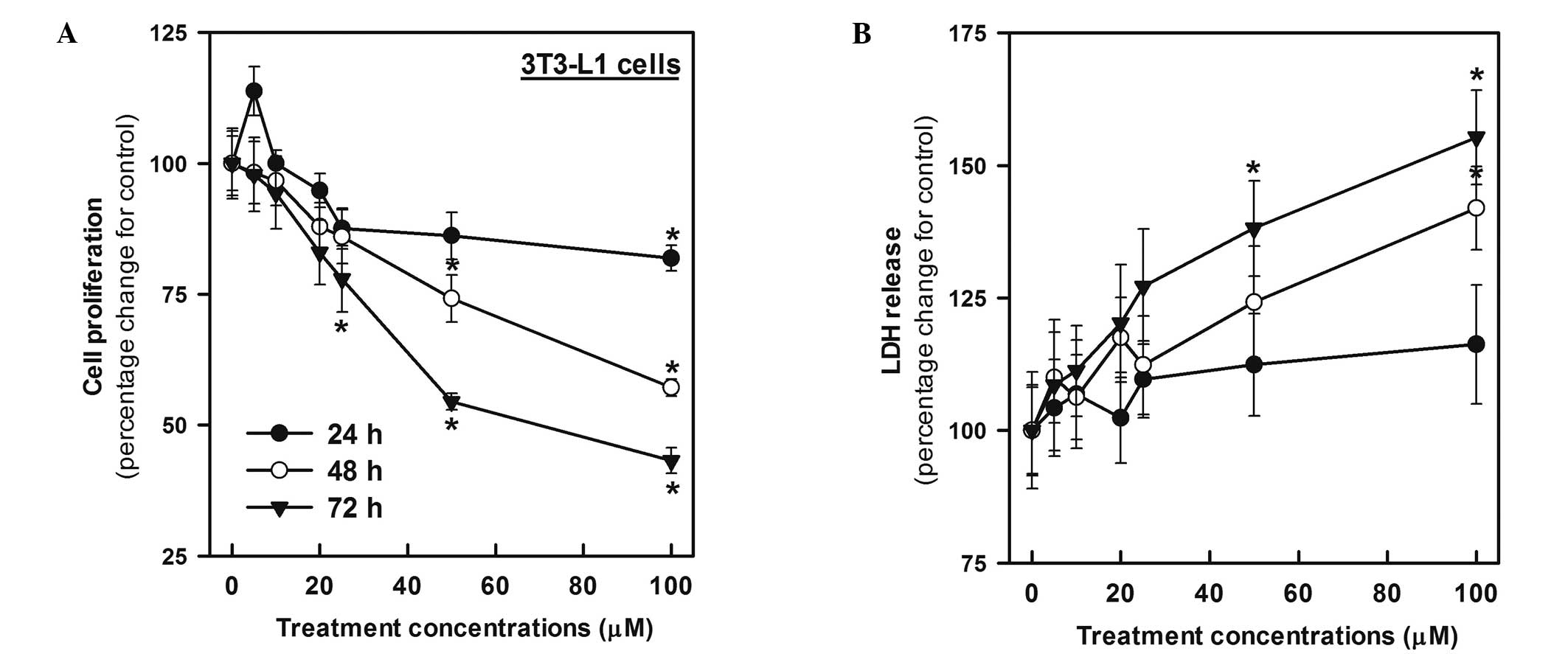
Cytotoxicity of genistein toward 3T3-L1
preadipocytes
To assess the cytotoxicity of genistein, cell growth
and LDH release were measured in 3T3-L1 cells exposed to genistein
at 5–100 μM for 24, 48 and 72 h. Genistein significantly decreased
cell growth after 48 and 72 h in a concentration-dependent manner
[the half maximal inhibitory concentration (IC50) for 48
and 72 h was 111.67 and 77.1 μM, respectively; Fig. 2]. Moreover, under the same
conditions, exposure to genistein for 24 h caused LDH release to
increase by 4–16%; however, the increase was not statistically
significant. After 48 and 72 h, although the release of LDH
increased in a concentration- and time-dependent manner,
significant inhibition of cell growth was first observed in cells
treated with 100 and 50 μM genistein for 48 and 72 h, respectively
(Fig. 2).
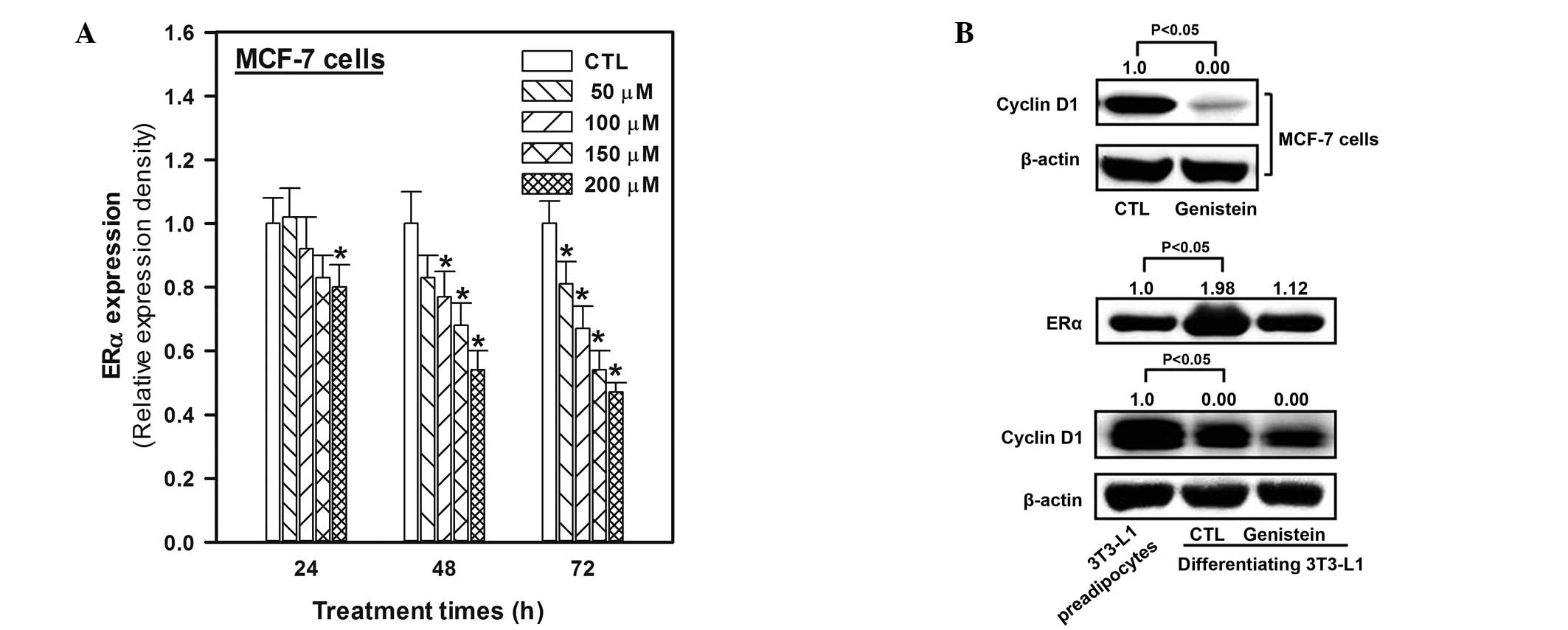
ERα, cyclin D1, and Bcl-2 expression in
genistein-treated MCF-7 or differentiating 3T3-L1 cells
To elucidate the mechanism of the ERα-dependent
antiproliferative activity of genistein, cells were exposed to 50,
100, 150 and 200 μM genistein for 24, 48 and 72 h. The results
revealed that the patterns of ERα expression and proliferation were
similar (Fig. 3). ERα expression
was downregulated by genistein at all concentrations; furthermore,
the effect of genistein was greater after 48 and 72 h. ERα
expression was upregulated 1.98-fold in 3T3-L1 cells following
inducer treatment for 48 h as compared with that in the negative
control (3T3-L1 preadipocytes, Fig.
3), in order to initiate differentiation. Genistein treatment
at a concentration of 50 μM for 48 h, which was selected as the
effective (no significant cellular toxicity) concentration,
restored ERα expression to almost the initial differentiating
levels.
Cyclin D1 expression in MCF-7 and 3T3-L1 cells was
decreased by treatment with genistein at 50 μM for 48 h (Fig. 3).
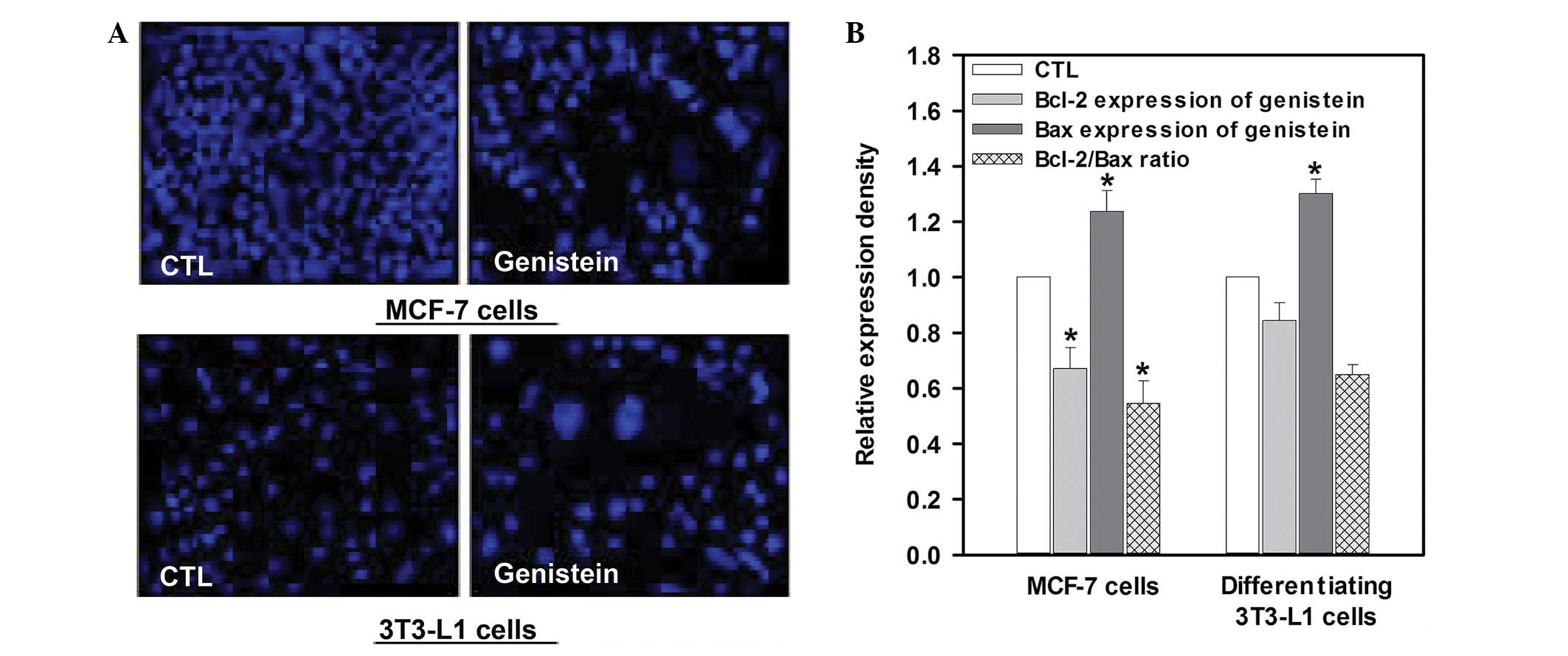
Apoptosis inducing effect of genistein on
MCF-7 and differentiating 3T3-L1 cells
As shown in Fig. 4,
exposure of MCF-7 cells to genistein at a concentration of 50 μM
for 48 h produced apoptotic morphological features, including cell
shrinkage and dot-shaped nuclear fragments. Compared with the
control cells, exposure to genistein resulted in an increase in
apoptotic morphological features on both MCF-7 and differentiating
3T3-L1 cells. This result is supported by the fact that genistein
significantly decreased the Bcl-2/Bax expression ratio by 45.5% and
35.2% in MCF-7 and differentiating 3T3-L1 cells, respectively,
compared with their respective control levels (P<0.05, Fig. 4).
Discussion
In the present study, the antiproliferative effects
of genistein on MCF-7 cells at concentrations of 50, 100, 150 and
200 μM were investigated. Genistein is a phytoestrogen, a
plant-derived phenolic compound that structurally mimics the
hormone 17β-estradiol (15). Thus,
the effects of genistein on estrogen receptors (ERs) are both
agonistic and antagonistic (16,17),
and may explain the biphasic effect of genistein on MCF-7 cell
proliferation. A previous study (18) revealed that the antiproliferative
effect of genistein on MCF-7 cells was biphasic (inhibitory at high
concentrations and stimulatory at low concentrations). This result
is consistent with that of other previous studies demonstrating
that estrogen-like bioactive molecules may stimulate breast cancer
cell growth (19–21). In the present study, following
treatment for 24, 48 and 72 h, genistein significantly inhibited
the proliferation of MCF-7 cells in a concentration-dependent
manner. Notably, ERα expression was downregulated while
proliferation was inhibited in MCF-7 cells exposed to
genistein.
ERα is a member of the steroid receptor superfamily
that regulates processes such as growth and differentiation in
various target cells by affecting transcription. ERα also plays an
important role in the development and progression of breast cancer.
A novel strategy for breast cancer chemotherapy is the
identification of ERα regulators among phytoestrogens (22,23).
Obesity is associated with an increased risk of
developing cancer; in particular, obesity plays an important role
in the pathogenesis of breast cancer since it causes altered
adipokine levels, elevated circulating estrogen levels and insulin
resistance (24). The
differentiation of 3T3-L1 preadipocytes results in cells with the
biochemical characteristics of adipocytes (for example, increased
ERα expression). Upon the induction of differentiation in 3T3-L1
cells following inducer treatment for 48 h, ERα expression was
significantly downregulated by genistein to an extent similar to
that in 3T3-L1 preadipocytes. Based on these results, genistein may
be associated with the proliferation of MCF-7 cells and
differentiation of 3T3-L1 cells via ERα expression.
When genistein was applied to MCF-7 and 3T3-L1
cells, cyclin D1 expression was reduced. Cyclin D1 is a prominent
target of estrogens in breast cancer cells and its induction is
important for the progression of cells through the G1 phase of the
cell cycle (25). The results of
several studies suggest that cyclin D1 is overexpressed in breast
cancer (26,27) and that it is associated with ER
positivity in breast cancer (28–30).
Furthermore, the induction of apoptosis by genistein
is supported by evidence demonstrating that the apoptotic cell
population was increased in MCF-7 and 3T3-L1 cells. Apoptosis is
essential for tissue development and homeostasis. The mechanism of
apoptosis involves a balance between factors that induce and those
that inhibit the process. Pro-apoptotic agents have been proposed
as a novel strategy not only for cancer chemotherapy but also for
the treatment of obesity (31). It
has been reported (32,33) that the induction of apoptosis in
adipocytes, which are otherwise resistant to apoptosis due to high
levels of Akt/protein kinase B and Bcl-2, may be a method of
reducing adipocyte numbers. In MCF-7 and 3T3-L1 cells, genistein
treatment significantly reduced the Bcl-2/Bax ratio by decreasing
Bcl-2 expression and increasing Bax expression. Bax genes and
members of the Bcl-2 family, such as Bcl-2, are involved in the
control of apoptotic pathways (34). The decreased expression of Bcl-2
and increased expression of Bax is associated with the response of
cancer cell lines to anticancer compounds. In addition, a loss of
Bcl-2 expression may promote the induction of apoptosis.
Based on the results of the current study, genistein
inhibits the proliferation of MCF-7 and differentiation of 3T3-L1
cells via apoptosis induction and an ERα-related pathway. The
effects of genistein observed in the present study make it
potentially useful for further development as not only a
chemotherapeutical agent for breast cancer but also a
chemopreventive agent for obesity.
Acknowledgements
The present study was supported by the Basic
Research Program through the National Research Foundation of Korea
(NRF) funded by the Ministry of Education, Science and Technology
(NRF-2013R1A1A2008595).
References
|
1
|
Liggins J, Bluck LJ, Runswick S, et al:
Daidzein and genistein content of fruits and nuts. J Nutr Biochem.
11:326–331. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Liggins J, Bluck LJ, Runswick S, et al:
Daidzein and genistein contents of vegetables. Br J Nutr.
84:717–725. 2000.PubMed/NCBI
|
|
3
|
Liggins J, Mulligan A, Runswick S and
Bingham SA: Daidzein and genistein content of cereals. Eur J Clin
Nutr. 56:961–966. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Akiyama T, Ishida J, Nakagawa S, et al:
Genistein, a specific inhibitor of tyrosine-specific protein
kinases. J Biol Chem. 262:5592–5595. 1987.PubMed/NCBI
|
|
5
|
Ullrich A and Schlessinger J: Signal
transduction by receptors with tyrosine kinase activity. Cell.
61:203–212. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kyle E, Neckers L, Takimoto C, et al:
Genistein-induced apoptosis of prostate cancer cells is preceded by
a specific decrease in focal adhesion kinase activity. Mol
Pharmacol. 51:193–200. 1997.
|
|
7
|
Peeters PH, Keinan-Boker L, van der Schouw
YT and Grobbee DE: Phytoestrogens and breast cancer risk. Review of
the epidemiological evidence. Breast Cancer Res Treat. 77:171–183.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Qin LQ, Xu JY, Wang PY and Hoshi K:
Soyfood intake in the prevention of breast cancer risk in women: a
meta-analysis of observational epidemiological studies. J Nutr Sci
Vitaminol (Tokyo). 52:428–436. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Iwasaki M and Tsugane S: Risk factors for
breast cancer: epidemiological evidence from Japanese studies.
Cancer Sci. 102:1607–1614. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lamartiniere CA, Zhang JX and Cotroneo MS:
Genistein studies in rats: potential for breast cancer prevention
and reproductive and developmental toxicity. Am J Clin Nutr.
68:1400S–1405S. 1998.PubMed/NCBI
|
|
11
|
de Lemos ML: Effects of soy phytoestrogens
genistein and daidzein on breast cancer growth. Ann Pharmacother.
35:1118–1121. 2001.PubMed/NCBI
|
|
12
|
Kittaneh M, Montero AJ and Glück S:
Molecular profiling for breast cancer: a comprehensive review.
Biomark Cancer. 5:61–70. 2013.PubMed/NCBI
|
|
13
|
Esfahlan RJ, Zarghami N, Esfahlan AJ, et
al: The possible impact of obesity on androgen, progesterone and
estrogen receptors (ERα and ERβ) gene expression in breast cancer
patients. Breast Cancer (Auckl). 5:227–237. 2011.PubMed/NCBI
|
|
14
|
Williams C and Lin CY: Oestrogen receptors
in breast cancer: basic mechanisms and clinical implications.
Ecancermedicalscience. 7:3702013.PubMed/NCBI
|
|
15
|
Sirtori CR, Arnoldi A and Johnson SK:
Phytoestrogens: end of a tale? Ann Med. 37:423–438. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Mueller SO, Simon S, Chae K, et al:
Phytoestrogens and their human metabolites show distinct agonistic
and antagonistic properties on estrogen receptor alpha (ERalpha)
and ERbeta in human cells. Toxicol Sci. 80:14–25. 2004. View Article : Google Scholar
|
|
17
|
Bovee TF, Schoonen WG, Hamers AR, et al:
Screening of synthetic and plant-derived compounds for
(anti)estrogenic and (anti)androgenic activities. Anal Bioanal
Chem. 390:1111–1119. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Choi EJ and Kim GH: Antiproliferative
activity of daidzein and genistein may be related to ERα/c-erbB-2
expression in human breast cancer cells. Mol Med Rep. 7:781–784.
2013.PubMed/NCBI
|
|
19
|
Hsu JT, Hung HC, Chen CJ, et al: Effects
of the dietary phytoestrogen biochanin A on cell growth in the
mammary carcinoma cell line MCF-7. J Nutr Biochem. 10:510–517.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Choi EJ and Kim T: Equol induced apoptosis
via cell cycle arrest in human breast cancer MDA-MB-453 but not
MCF-7 cells. Mol Med Rep. 1:239–244. 2008.PubMed/NCBI
|
|
21
|
Rajah TT, Du N, Drews N and Cohn R:
Genistein in the presence of 17beta-estradiol inhibits
proliferation of ERbeta breast cancer cells. Pharmacology.
84:68–73. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Björnström L and Sjöberg M: Mechanisms of
estrogen receptor signaling: convergence of genomic and nongenomic
actions on target genes. Mol Endocrinol. 19:833–842.
2005.PubMed/NCBI
|
|
23
|
Liu MM, Huang Y and Wang J: Developing
phytoestrogens for breast cancer prevention. Anticancer Agents Med
Chem. 12:1306–1313. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Iyengar NM, Hudis CA and Dannenberg AJ:
Obesity and inflammation: new insights into breast cancer
development and progression. Am Soc Clin Oncol Educ Book. 46–51.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Musgrove EA, Lee CS, Buckley MF and
Sutherland RL: Cyclin D1 induction in breast cancer cells shortens
G1 and is sufficient for cells arrested in G1 to complete the cell
cycle. Proc Natl Acad Sci USA. 91:8022–8026. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Buckley MF, Sweeney KJ, Hamilton JA, et
al: Expression and amplification of cyclin genes in human breast
cancer. Oncogene. 8:2127–2133. 1993.PubMed/NCBI
|
|
27
|
Bartkova J, Lukas J, Müller H, et al:
Cyclin D1 protein expression and function in human breast cancer.
Int J Cancer. 57:353–361. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Barbareschi M, Pelosio P, Caffo O, et al:
Cyclin-D1-gene amplification and expression in breast carcinoma:
relation with clinicopathologic characteristics and with
retinoblastoma gene product, p53 and p21WAF1 immunohistochemical
expression. Int J Cancer. 74:171–174. 1997. View Article : Google Scholar
|
|
29
|
Utsumi T, Yoshimura N, Maruta M, et al:
Correlation of cyclin D1 MRNA levels with clinico-pathological
parameters and clinical outcome in human breast carcinomas. Int J
Cancer. 89:39–43. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Butt AJ, McNeil CM, Musgrove EA and
Sutherland RL: Downstream targets of growth factor and oestrogen
signalling and endocrine resistance: the potential roles of c-Myc,
cyclin D1 and cyclin E. Endocr Relat Cancer. 12(Suppl 1): S47–S59.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Herold C, Rennekampff HO and Engeli S:
Apoptotic pathways in adipose tissue. Apoptosis. 18:911–816. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Nelson-Dooley C, Della-Fera MA, Hamrick M
and Baile CA: Novel treatments for obesity and osteoporosis:
targeting apoptotic pathways in adipocytes. Curr Med Chem.
12:2215–25. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhang Y and Huang C: Targeting adipocyte
apoptosis: a novel strategy for obesity therapy. Biochem Biophys
Res Commun. 417:1–4. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Cory S and Adams JM: Killing cancer cells
by flipping the Bcl-2/Bax switch. Cancer Cell. 8:5–6. 2005.
View Article : Google Scholar : PubMed/NCBI
|