Introduction
Candida albicans (C. albicans), an
opportunistic pathogen, is able to attack hosts that are
immunocompromised or otherwise debilitated. With the increased use
of immunosuppressive and cytotoxic drugs, antibiotic abuse and
implanted devices, including urinary catheters and endotracheal
tubes, the prevalence of fungal infections has also increased
(1–3). C. albicans was the leading
cause of pulmonary fungal infection in 2011 and the drug resistance
rate demonstrates a tendency to increase year by year (4). A number of implanted devices,
including intravascular or urinary catheters and endotracheal
tubes, are associated with fungal infections, and a biofilm may be
detected on their surface (5–7). In
recent years, emerging cases of C. albicans drug resistance
have been primarily attributed to the formation of biofilms, with
resistance increasing in conjunction with the maturation of the
biofilm (8). Mature Candida
biofilms consist of a complex three-dimensional structure of layers
of yeast cells, hyphae and an abundant exopolysaccharide matrix
(9,10). As natural resistance barriers of
antifungal drugs, biofilms play a significant role in the invasion
and dissemination of C. albicans.
Chitosan is a natural linear polyatomic biopolymer
comprising N-acetyl-D-glucosamine and β-(1,4)-linked D-glucosamine. A previous study
revealed that surfaces coated with chitosan were able to resist
biofilm formation by bacteria and fungi in vitro (11). Chitosan is nontoxic to humans and
exhibits excellent biocompatibility. The biopolymer is easily
catalyzed to oligosaccharides by various biological enzymes, and is
considered biodegradable (12);
thus, can be absorbed by the body. Previous studies have
demonstrated that chitosan not only has immunoregulatory efficacy,
but also a naturally broad spectrum of antibacterial activity that
enables the inhibition of biofilm formation (11,13,14).
Chitosan is an extensively studied biomacromolecule
that is typically derived from chitin, a major component of
crustacean shells. Due to their biocompatible and biodegradable
properties, chitosan and its derivatives have been proposed for
application in novel drug studies. The effect of chitosan on
bacterial biofilms has been extensively reported; however, the aim
of the present study was to investigate the susceptibility of C.
albicans biofilms to chitosan, with an emphasis on determining
the effects of chitosan on various biofilm growth phases and
architectural organization.
Materials and methods
C. albicans
A C. albicans strain, previously isolated
from a patient, was obtained from the Clinical Laboratory of the
Children’s Hospital of Chongqing Medical University (Chongqing,
China). The fungus was inoculated on a chocolate agar plate and
grown at 37°C for 48 h. A single colony of the desired strain was
inoculated into 8 ml Sabouraud dextrose broth (SDB; Sangon Biotech
Co., Ltd., Shanghai, China) and incubated overnight at 37°C in a
rotary shaker at 220 rpm.
Chitosan susceptibility in C. albicans
planktonic cells
Chitosan (Sigma-Aldrich, St. Louis, MO, USA) was
solubilized in 0.2% acetic acid at a working concentration of 1% to
pH 5.0–5.5. Serial two-fold dilutions of chitosan stock solutions
in SDB were prepared in plastic centrifuges over a range of
0.0078–0.5% and stored at 4°C. A standard inoculum of
1v107 cells [optical density (OD)600] from
the overnight culture of the fungal strain was prepared prior to
each experiment. Briefly, 100 μl C. albicans planktonic
cells were mixed with 100 μl chitosan solution of various
concentrations and incubated for 48 h at 37°C. Data were obtained
from three independent experiments. For the positive control, C.
albicans planktonic cells were incubated in the presence of 200
μl SDB without chitosan, while in the negative control, 200 μl SDB
only was incubated in wells under otherwise identical conditions.
The minimum inhibitory concentrations (MIC)50 for the
biofilms and planktonic cells were defined as the minimum
antifungal concentration that caused ≥50% fungal damage compared
with the untreated controls.
Measurement of chitosan susceptibility in
biofilms using a
2,3-bis-(2-methoxy-4-nitro-5-sulfophenyl)-2H-tetrazolium-5-
carboxanilide (XTT) reduction assay
C. albicans biofilm experiments were
performed in untreated 96-well plates. The wells of the 96-well
plates were incubated for various time periods (2, 8, 24 and 48 h)
in 100 μl C. albicans suspension (1×107 cells/ml)
at 37°C. The medium in each well was removed at the indicated time
points and the biofilms were washed twice with phosphate-buffered
saline (PBS). Following the removal of PBS, 100 μl chitosan
solution of various concentrations were added to one well of a
96-well plate and then incubated for additional 48 h at 37°C. A
formazan salt-based XTT reduction assay was performed to assess the
metabolic activity (15). All the
tests were performed in duplicate and the average was calculated.
Positive and negative controls were established as previously
described. A total of 100 μl XTT-menadione solution, consisting of
XTT (Sangon Biotech) salt solution (0.5 g/l in Ringer-Locke liquor)
mixed with menadione (Sangon Biotech Co., Ltd.) solution (1 mM in
acetone; Sigma-Aldrich), was added to each well. The 96-well plates
were incubated for 2 h at 37°C. Colorimetric changes were scanned
at 490 nm with a Varioskan™ Flash Multimode Reader
(Thermo Fisher Scientific, Waltham, MA, USA).
C. albicans biofilm formation with cover
slips
Cover slips (0.8×0.8 cm) that were used for biofilm
growth were soaked in concentrated sulfuric acid overnight. The
following day, the concentrated sulfuric acid was washed with
flowing water and the clean cover slips were immersed in 95%
alcohol overnight. All the cover slips were washed three times with
deionized water. For biofilm growth on the treated cover slips, as
aforementioned, the cover slips were placed in 24-well plates and
immersed in fetal bovine serum (FBS; Sigma-Aldrich, Beijing, China)
at 4°C overnight. Following this pretreatment, the cover slips were
washed with PBS (0.01 M) to remove the residual FBS. To ensure
uniform biofilm formation, the cover slips were immersed in 1 ml
standardized cell suspension (1×107 cells/ml) and
incubated for 90 min at 37°C. The cover slips were lifted carefully
using tweezers and gently placed in each well of the 24-well plate
containing 1.5 ml fresh SDB medium. Samples were incubated for
various durations at 37°C.
Susceptibility of the biofilms to
chitosan
To evaluate the chitosan susceptibility of C.
albicans cells grown in developing biofilms, the pretreated
cover slips were immersed in 1 ml standardized cell suspension
(1×107 cells/ml) and incubated for 90 min at 37°C. The
cover slips were lifted carefully using tweezers and gently placed
in each well of the 24-well plate containing 1 ml fresh SDB medium.
A 0.0625% chitosan solution was added to 24-well plates with cover
slips and incubated for an additional 8, 24 and 48 h at 37°C.
Fluorescence microscopy
Cover slips pretreated with biofilms were
transferred to microscope slides and stained for 1 min with 50 μl
Calcofluor White M2R [0.05% (vol/vol); Sigma-Aldrich] under minimal
ambient light or in a darkroom. The biofilms were observed
immediately under an ultraviolet (UV) range of 440 nm excitation
and 500–520 nm emission wavelengths. Any superfluous fluorochrome
on the cover slips was absorbed by the filter paper. The stained
biofilms were examined under a fluorescence microscope.
Scanning electron microscopy (SEM)
C. albicans biofilms were grown on pretreated
cover slips in 24-well plates, as described previously. In the
chitosan group, C. albicans biofilms were incubated in
0.0625% chitosan. For the positive control, C. albicans
biofilms were incubated in the presence of 200 μl SDB without
chitosan for 24 h. The cover slips with biofilms were subsequently
washed three times with PBS and transferred to an additional
24-well plate containing 2.5% glutaraldehyde at 4°C. The samples
were prepared using a regular method for electron microscopy
examination (16) and viewed under
an S-3000N scanning electron microscope (Hitachi High-Technologies,
Tokyo, Japan). Two separate sets of culture were prepared.
Statistical analysis
Statistical analyses were performed using SPSS 19.0
software (SPSS, Inc., Chicago, IL, USA). All data were tested for
normality and transformed when necessary to meet the assumption of
normal distribution. P-values were calculated by one-way analysis
of variance and Fisher’s least significant difference test was
conducted to determine differences among the test groups. P<0.05
was considered to indicate a statistically significant
difference.
Results
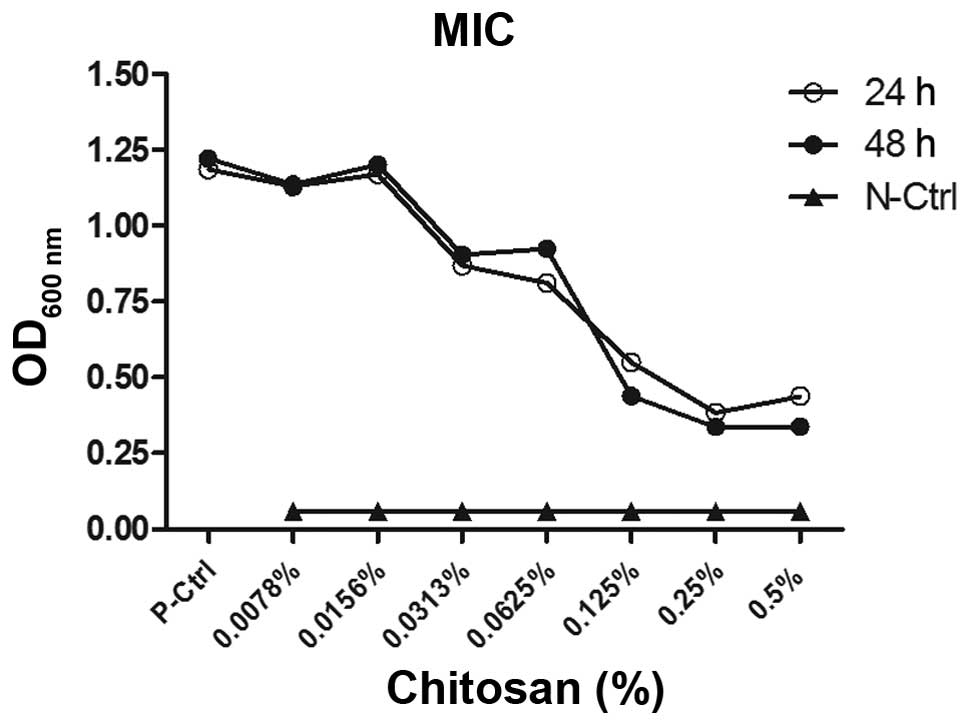
MIC values of chitosan on C. albicans
planktonic cells
Fig. 1 shows the
results of the experiment assessing the effect of chitosan at
various concentrations on C. albicans planktonic cell
growth. Chitosan, at a concentration of >0.0313%, was shown to
severely inhibit the activity of C. albicans planktonic
cells grown for 24 h. The OD decreased significantly when compared
with the positive control (P<0.05). When comparing the effect of
chitosan over 24 and 48 h, the MIC of chitosan exhibited no
statistically significant change on cell growth.
C. albicans biofilm formation
An XTT reduction assay was used to quantify the
effects of chitosan on biofilms produced by C. albicans. As
shown in Fig. 2, the different
phases of C. albicans biofilm formation were significantly
susceptible to chitosan. At 2 h, the OD values, as determined by an
enzyme-labeled instrument, were similar to when the concentration
of chitosan was >0.0313%. A statistically significant difference
was observed in the OD values between the 0.0313 and 0.0156%
chitosan groups (P<0.05). In addition, a marked difference was
observed between all the chitosan groups and the positive control
(P<0.05). The results did not demonstrate a statistically
significant difference between biofilm development at 2 and 8 h,
and the optimum concentration of chitosan was almost equal.
Compared with the biofilms grown for 2 or 8 h, the OD values for
the biofilms grown for 24 or 48 h were significantly higher
following exposure to chitosan at a concentration of 0.0313%. The
results revealed that biofilms in the mature phase (24-8 h)
demonstrated less susceptibility to higher concentrations of
chitosan compared with those in the early phase (2–8 h). Thus,
biofilm formation exhibits a significant resistance to the
antifungal activity of chitosan.
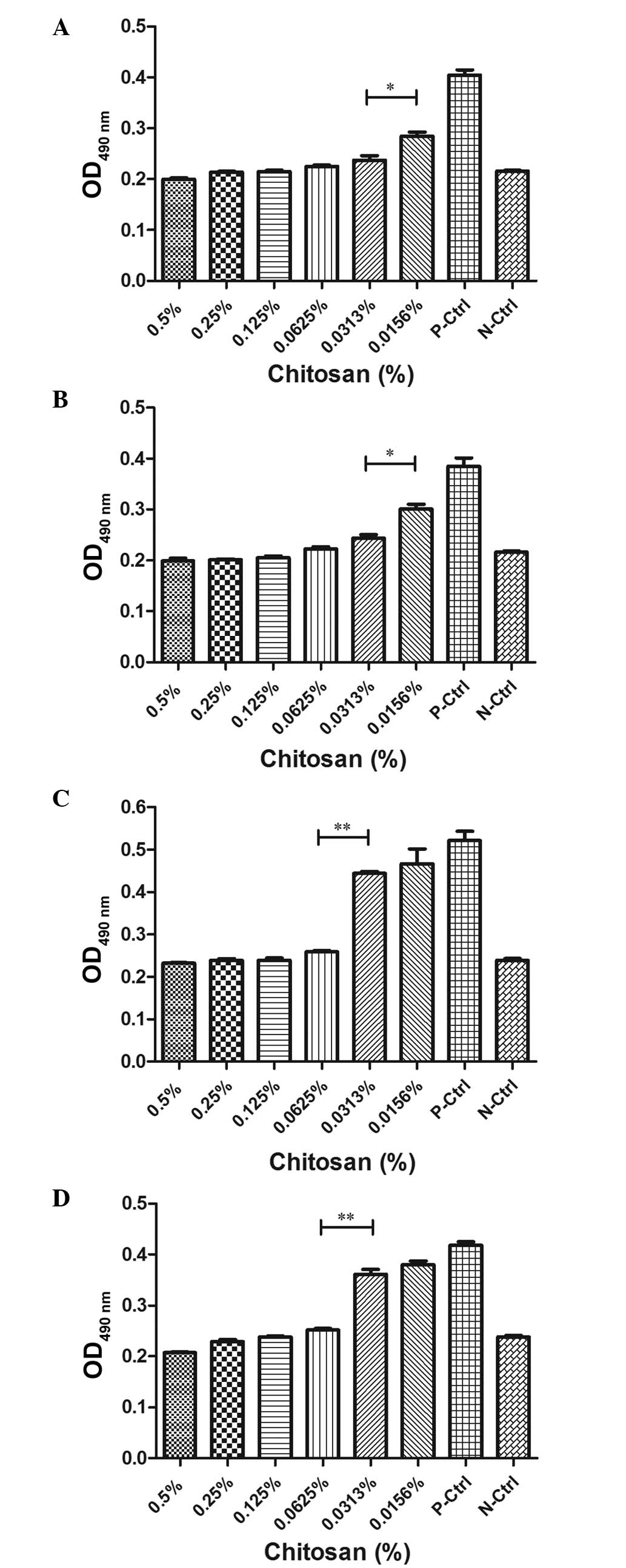 | Figure 2Candida albicans (C.
albicans) biofilm development with chitosan resistance. The
susceptibility of C. albicans biofilms, incubated for (A) 2
h, (B) 8 h, (C) 24 h and (D) 48 h, to various concentrations (0.5,
0.25, 0.125, 0.0625, 0.0313 and 0.0156%) of chitosan are
represented as histograms. For the positive control, C.
albicans planktonic cells were incubated without chitosan; for
the negative control, sabouraud dextrose broth only was incubated
in the wells. The OD values at the various stages of biofilm
development were compared with those of the fungal cells and the
negative control. Experiments were performed twice, with similar
results obtained each time. OD, optical density. |
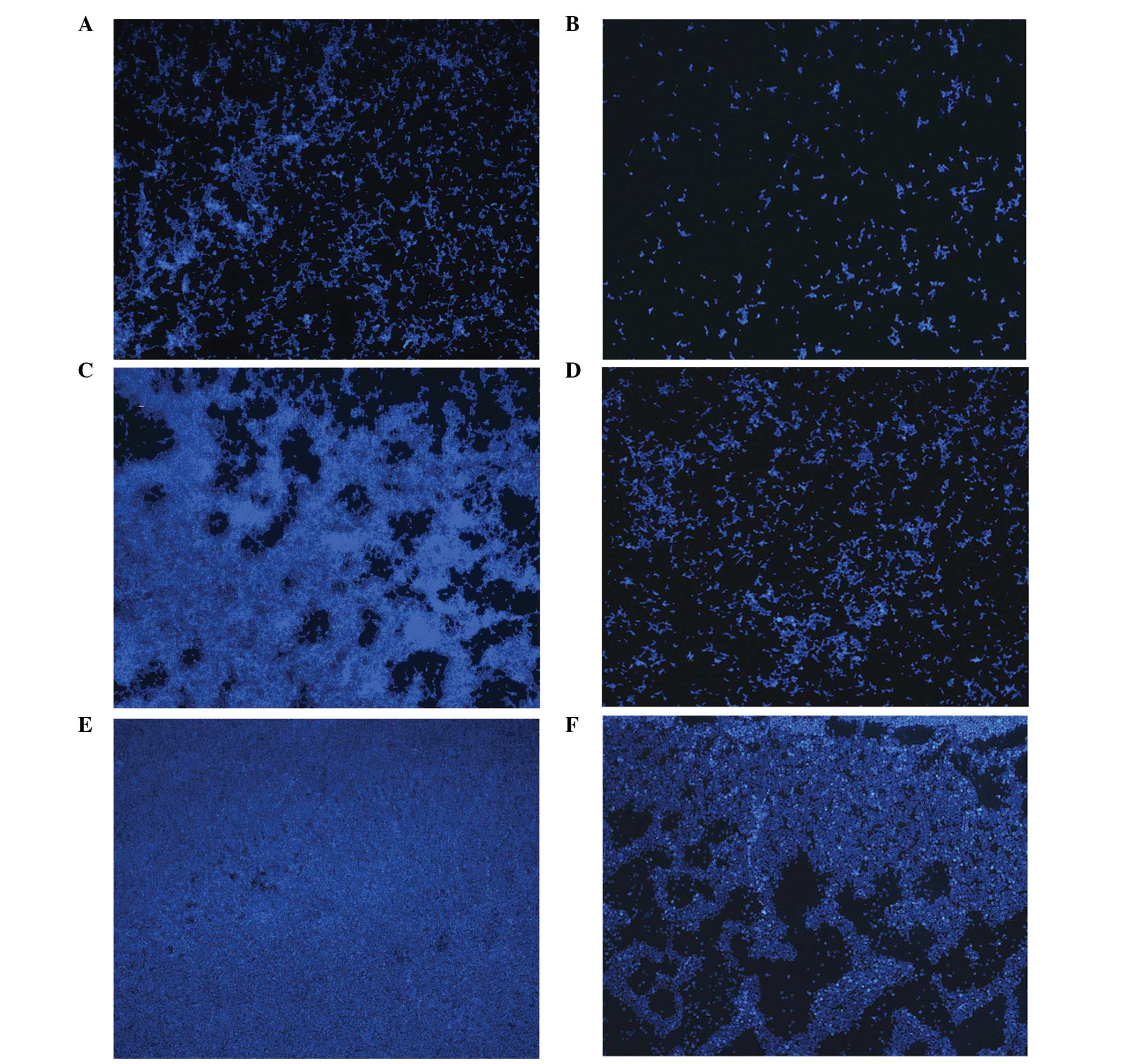
A previous study observed that C. albicans
biofilm formation on polymethylmethacrylate strips progresses in
three distinct developmental phases: Early, intermediate and
maturation (10). The effect of
chitosan on the temporal development of C. albicans biofilms
on cover slips was investigated with fluorescence microscopy using
Calcofluor White M2R, a UV-excitable dye that binds chitin and
β-glucan and has long been used to highlight the fungal cell wall
(17). Fluorescence microscopy was
performed to visualize and compare the three distinct developmental
phases between the untreated and chitosan-treated C.
albicans biofilms (Fig. 3). In
the early stage, characteristic microcolonies and short hyphae
appeared on the surface of the cover slips of the untreated cells
(Fig. 3A). As shown in Fig. 3B, a few yeast cells were dispersed
in the 0.0625% chitosan-treated group at the same stage. During the
intermediate developmental phase, a dense network of yeast cells
surrounded by a large amount of noncellular material was observed
in the untreated group, with the metabolically active cells
existing in a network (Fig. 3C).
The quantity of yeast cells in the chitosan-treated group at the
same stage was lower compared with the untreated group, and the
cells were unable to congregate and develop into a bioflim
(Fig. 3D). In the maturation
development phase, the surfaces of the cover slips were coated with
a thick layer of yeast cells and noncellular material; however, it
was difficult to distinguish between the cell types (Fig. 3E). By contrast, the
chitosan-treated group exhibited yeast cells lacking a network
structure and a significantly lower metabolic activity compared
with the untreated group (Fig.
3F).
Fluorescence microscopy images were used to
correlate the XTT reduction assay results with the visual effects
of the biofilm formation at the various exposure times. The results
indicated that chitosan was not only able to inhibit the metabolic
activity of cells in maturing biofilms, but also delay biofilm
formation by terminating the fusion process.
Direct visualization of the effect of
chitosan on C. albicans biofilms
SEM examination was used to visualize the structural
differences between normal and chitosan-treated C. albicans
biofilms. Normal C. albicans biofilms exhibited a dense
network of hyphae surrounded by vast amounts of exopolymeric matrix
(Fig. 4A). By comparison,
chitosan-treated C. albicans expressed a biofilm that lacked
a normal network structure and released polysaccharide (Fig. 4B). These results demonstrated that
chitosan significantly inhibited C. albicans biofilms in
vitro.
Discussion
Currently, fungal infections present an increasing
threat to the growing number of immunocompromised patients. These
contaminations, of which Candidiasis is the most common, represent
one of the most prevalent infections in US hospitals (18). A leading causative agent of
Candidiasis continues to be C. albicans (18). However, numerous problems remain in
the current therapies for fungal infections. Antifungal treatment
strategies for C. albicans are limited to a small
armamentarium of compounds, which mainly include azoles (such as
fluconazole), polyenes (such as amphotericin B) and echinocandins
(19). However, there are clinical
drawbacks to these drugs due to their toxicity (20). Therefore, identifying alternative
antifungal agents with fewer side effects is required. Furthermore,
since the majority of fungal infections are caused by biofilms,
this provides an additional reason for the low success rate of
treatment. Chitosan offers a flexible, biocompatible platform for
designing coatings to protect surfaces from infection and
decreasing the metabolic activity and survival rate of
Candida species biofilms (21). The characteristics of chitosan,
including its antiadherent and antifungal properties, mean that it
is a strong candidate for treatment against fungal biofilms. In a
normal conjunction, the C. albicans biofilm proceeds into
early, intermediate and maturation phases of development. The
extracellular material, as observed by microscopy, is predominantly
composed of cell wall-like polysaccharides containing mannose and
glucose residues. Biofilms grown on cover slips have a distinct
biphasic structure composed of an adherent blastospore layer
covered by sparser hyphal elements embedded in a deep layer of
extracellular material. The structure that meditates cell
interactions with the environment is the cell wall, which may be a
viable factor in the adhesion of fungi to solid surfaces.
The present study analyzed the role of chitosan on
C. albicans using the MIC. The results indicated that
chitosan had high fungistatic activity against these planktonic
cells. Repeated XTT reduction assay analysis revealed that chitosan
at a concentration of 0.0313% had the potential to kill >50% of
cells in the early and intermediate phases of biofilm development.
However, a higher concentration of chitosan was required to kill
cells in mature biofilms. Mature biofilms of C. albicans
were significantly less susceptible to chitosan than planktonic
cells and initial biofilms. The results of the current study
indicated that an addition of exogenous chitosan to C.
albicans biofilms reduced the metabolic activity of cells
significantly and apprehended the adhesion of yeast cells to the
polystyrene surface. Previous studies have demonstrated that the
biofilm phenotype confers resistance to antifungal therapy, which
is consistent with the results of the present study (22,23).
Physical stress of the biofilm structure due to permeabilization of
the cellular membrane, which permits higher levels of penetration
by chitosan and a more effective delivery of its antifungal
activity, may be the cause of this phenomenon (24,25).
Fluorescence and SEM techniques allowed the
morphology and structure of biofilms to be analyzed in the present
study. Fluorescence microscopy visualized the gross biofilm
morphology and the appearance of the extracellular matrix during
biofilm formation. SEM visualized the structure of the biofilms
surface without distortion of the native biofilm structure.
Progressing in three distinct developmental phases, C.
albicans biofilm formation is divided into early, intermediate
and maturation stages. The results of the present study revealed
that in the overall biofilm formation period, the chitosan-treated
biofilms exhibited a slower growth compared with those that were
untreated. Although chitosan treatment did not completely destroy
the yeast cells, the staining observed with fluorescence and SEM
demonstrated that chitosan-treated biofilms were significantly
inhibited in the formation of a complex network.
In conclusion, the results of the present study
demonstrated that chitosan may be developed as an antimicrobial
agent against the treatment of clinically-associated fungal biofilm
diseases. Although an inhibitory effect of chitosan on biofilm
production by C. albicans was reported in the laboratory and
in clinical isolates in vitro, further studies using animal
models of pulmonary C. albicans infection are required.
Acknowledgements
The study was supported in part by grants from the
National Natural Science Foundation of China for Young Scholars
(no. 81000745) and the Project of Medicine Science of Health Bureau
of Chongqing [nos. (2009)66 and (2010)51]. The authors thank the
Clinical Laboratory at the Children’s Hospital of Chongqing Medical
University for providing the C. albicans strain.
References
|
1
|
Hallahan AR, Shaw PJ, Rowell G, O’Connell
A, Schell D and Gillis J: Improved outcomes of children with
malignancy admitted to a pediatric intensive care unit. Crit Care
Med. 28:3718–3721. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Booy R, Habibi P, Nadel S, et al:
Reduction in case fatality rate from meningococcal disease
associated with improved healthcare delivery. Arch Dis Child.
85:386–390. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Tenner PA, Dibrell H and Taylor RP:
Improved survival with hospitalists in a pediatric intensive care
unit. Crit Care Med. 31:847–852. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Chen Yan-jun WF: Study on pathogens
causing lower respiratory tract infections and their drug
resistance. Zhong Guo Bing Yuan Sheng Wu Xue Za Zhi. 8:561–563.
2013.(In Chinese).
|
|
5
|
Adair CG, Gorman SP, Feron BM, et al:
Implications of endotracheal tube biofilm for ventilator-associated
pneumonia. Intensive Care Med. 25:1072–1076. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Crump JA and Collignon PJ: Intravascular
catheter-associated infections. Eur J Clin Microbiol Infect Dis.
19:1–8. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Maki DG and Tambyah PA: Engineering out
the risk for infection with urinary catheters. Emerg Infect Dis.
7:342–347. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Douglas LJ: Candida biofilms and
their role in infection. Trends Microbiol. 11:30–36. 2003.
View Article : Google Scholar
|
|
9
|
Blankenship JR and Mitchell AP: How to
build a biofilm: a fungal perspective. Curr Opin Microbiol.
9:588–594. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Chandra J, Mukherjee PK and Ghannoum MA:
In vitro growth and analysis of Candida biofilms. Nat
Protoc. 3:1909–1924. 2008. View Article : Google Scholar
|
|
11
|
Carlson RP, Taffs R, Davison WM and
Stewart PS: Anti-biofilm properties of chitosan-coated surfaces. J
Biomater Sci Polym Ed. 19:1035–1046. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Alonso MJ and Sánchez A: The potential of
chitosan in ocular drug delivery. J Pharm Pharmacol. 55:1451–1463.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kulikov SN, Tiurin IuA, Fassakhov RS and
Varlamov VP: Antibacterial and antimycotic activity of chitosan:
mechanisms of action and role of the structure. Zh Mikrobiol
Epidemiol Immunobiol. 5:91–97. 2009.(In Russian).
|
|
14
|
Pasquantonio G, Greco C, Prenna M, et al:
Antibacterial activity and anti-biofilm effect of chitosan against
strains of Streptococcus mutans isolated in dental plaque.
Int J Immunopathol Pharmacol. 21:993–997. 2008.PubMed/NCBI
|
|
15
|
Meshulam T, Levitz SM, Christin L and
Diamond RD: A simplified new assay for assessment of fungal cell
damage with the tetrazolium dye,
(2,3)-bis-(2-methoxy-4-nitro-5-sul-
phenyl)-(2H)-tetrazolium-5-carboxanilide (XTT). J Infect Dis.
172:1153–1156. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Tsang PW, Bandara HM and Fong WP: Purpurin
suppresses Candida albicans biofilm formation and hyphal
development. PLoS One. 7:e508662012.PubMed/NCBI
|
|
17
|
Albani JR and Plancke YD: Interaction
between calcofluor white and carbohydrates of alpha 1-acid
glycoprotein. Carbohydr Res. 314:169–175. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wisplinghoff H, Ebbers J, Geurtz L,
Stefanik D, Major Y, Edmond MB, Wenzel RP and Seifert H: Nosocomial
bloodstream infections due to Candida spp. in the USA:
species distribution, clinical features and antifungal
susceptibilities. Int J Antimicrob Agents. 43:78–81. 2014.
|
|
19
|
Georgopapadakou NH and Walsh TJ: Human
mycoses: drugs and targets for emerging pathogens. Science.
264:371–373. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ramage G, VandeWalle K, Bachmann SP,
Wickes BL and López-Ribot JL: In vitro pharmacodynamic properties
of three antifungal agents against preformed Candida
albicans biofilms determined by time-kill studies. Antimicrob
Agents Chemother. 46:3634–3636. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Martinez LR, Mihu MR, Tar M, et al:
Demonstration of antibiofilm and antifungal efficacy of chitosan
against candidal biofilms, using an in vivo central venous catheter
model. J Infect Dis. 201:1436–1440. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hawser SP and Douglas LJ: Resistance of
Candida albicans biofilms to antifungal agents in vitro.
Antimicrob Agents Chemother. 39:2128–2131. 1995.
|
|
23
|
Mukherjee PK, Chandra J, Kuhn DM and
Ghannoum MA: Mechanism of fluconazole resistance in Candida
albicans biofilms: phase-specific role of efflux pumps and
membrane sterols. Infect Immun. 71:4333–4340. 2003.PubMed/NCBI
|
|
24
|
Rabea EI, Badawy ME, Stevens CV, Smagghe G
and Steurbaut W: Chitosan as antimicrobial agent: applications and
mode of action. Biomacromolecules. 4:1457–1465. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Sudarshan NR, Hoover DG and Knorr D:
Antibacterial action of chitosan. Food Biotechnol. 6:257–272. 1992.
View Article : Google Scholar
|