Introduction
Hepatocellular carcinoma (HCC) is the fifth most
common malignancy worldwide (1,2).
Although surveillance of patients with risk factors for HCC and the
development of locoregional treatment options have improved
outcomes, to date, there are no effective curative methods due to
the high invasion, early metastasis and high tumor recurrence rates
of HCC following surgery or interventional treatment. Furthermore,
HCC progression, including metastasis, contributes to the high
fatality rates of liver cancer. Lymph node metastasis of tumors is
considered to be an important factor involved in HCC progression.
However, the underlying molecular mechanisms involved in lymph node
metastasis of HCC remain unclear.
Aurora kinases are key regulators of protein
phosphorylation during mitosis (3), consisting of three members that
differ with regard to subcellular localization, activation kinetics
and function. Aurora-B is one of the major protein kinases that
ensures the proper execution and fidelity of mitosis. As a member
of the chromosomal passenger complex (CPC), Aurora-B has been
implicated in various mitotic functions, including
chromosome-microtubule interactions, sister chromatid cohesion, the
spindle-assembly checkpoint and cytokinesis. Accumulating evidence
indicates that Aurora-B is an important antitumor target that is
strongly associated with lymph node metastasis in various tumor
types (4–7). Previous studies have demonstrated
that the expression of Aurora-B is increased in HCC cells, and the
inhibition of Aurora-B suppresses cell proliferation and invasion
in vitro and in vivo (8–11).
However, the potential molecular mechanisms underlying the
involvement of Aurora-B in HCC development and lymph node
metastasis remain unclear.
Matrix metalloproteinases (MMPs) are involved in the
degradation of the basement membrane and epimatrix, among which
MMP-2 and -9 markedly correlate with tumor invasion. A number of
studies have revealed that activation of the gene encoding nuclear
factor (NF)-κB, the upstream regulator of MMPs, promotes tumor cell
invasion and migration (12,13).
In addition, phosphorylation and activation of Akt has been
recognized as an important regulatory factor in the NF-κB signaling
pathway. A previous study demonstrated that an inhibitor of
Aurora-B decreased Akt phosphorylation at Ser 473 (14). Therefore, we hypothesized that the
inhibition of Aurora-B results in the suppression of HCC cell
invasion and migration via decreasing the activity of the
phosphoinositide 3-kinase (PI3K)/Akt/NF-κB signaling pathway.
Thus, the aim of the present study was to
investigate whether the inhibition of Aurora-B caused the
suppression of HepG2 cell invasion and migration via decreasing the
activity of the PI3K/Akt/NF-κB signaling pathway in
vitro.
Materials and methods
Construction of the recombinant plasmid
containing miRNA targeting the Aurora-B gene
A human cDNA sequence encoding the Aurora-B protein
was obtained from GenBank (NM-004217), and single-stranded DNA
oligos were designed and synthesized using the following primer
sequences: Forward,
5′-CCGGGAAGAGCTGCACATTTGACGACTCGAGTCGTCAAATGTGCAGCTCTTCTTTTTG-3′
and reverse,
5′-AATTCAAAAAGAAGAGCTGCACATTTGACGACTCGAGTCGTCAAATGTGCAGCTCTTC-3′.The
products were cloned into the express vector,
pcDNA6.2-GW/EmGFP-miR, using a BLOCK-iT™ Pol II miR RNAi Expression
Vector kit with EmGFP (K4936-00; Invitrogen Life Technologies,
Carlsbad, CA, USA). The DNA sequence of the plasmid was confirmed
using a PureLink HiPure Plasmid DNA kit (K2100-03; Invitrogen Life
Technologies).
Cell culture and transfection
A human HCC cell line, HepG2, (Shanghai Cell Bank of
the Chinese Academy of Sciences, Shanghai, China) was cultured in
Dulbecco’s modified Eagle’s medium (DMEM) supplemented with 10%
fetal bovine serum (FBS) and incubated at 37°C in 5%
CO2. The HepG2 cells were seeded in six-well plates at
30% confluence on the day prior to transfection. Transfection with
the recombinant plasmid targeting the Aurora-B gene (MiR-Aurora-B)
or the negative plasmid (MiR-Neg) was performed using Lipofectamine
2000 (Invitrogen Life Technologies). Transfection complexes were
prepared according to the manufacturer’s instructions.
Quantitative polymerase chain reaction
(qPCR)
The qPCR procedure was performed as follows: 95°C
for 2 min; 35 cycles of 95°C for 20 sec, 55°C for 30 sec and 72°C
for 30 sec; and annealing between 65 and 95°C with 0.5°C
progressive increases. The primers used in the study were as
follows: Aurora-B (NM-004217) forward, 5′-ATAGCAGTGGGACACCCGACAT-3′
and reverse, 5′-GGGACTTGAAGAGGACCTTGAGC-3′; and glyceraldehyde
3-phosphate dehydrogenase (GAPDH; NM-002046) forward,
5′-CCTGTTCGACAGTCAGCCGCATC-3′ and reverse,
5′-CGACCAAATCCGTTGACTCCGACC-3′.
Western blot analysis
Total protein from the cells was extracted using
radioimmunoprecipitation assay lysis buffer containing 60 μg/ml
phenylmethylsulfonyl fluoride. Protein concentration was determined
using a Bradford assay. Equal amounts of protein were
electrophoresed using 8% SDS-PAGE and transferred onto a pure
nitrocellulose blotting membrane (0.22 μM). The membranes were
blocked with 5% skimmed milk for 1 h at room temperature, then
incubated with primary antibodies [rabbit anti-Aurora-B
immunoglobulin G (IgG), 1:200; rabbit anti-phosphorylated (p)-Akt
IgG, 1:800; goat anti-Akt IgG, 1:1,000; rabbit anti-NF-κB, 1:500;
and rabbit anti-MMP-2 and MMP-9, 1:1,000] overnight at 4°C. The
membranes were washed prior to incubation with the appropriate
horseradish peroxidase (HRP)-conjugated secondary antibodies
(anti-rabbit, -goat and -mouse, 1:2,000). The immune complexes were
detected with a pro-light HRP kit (Tiangen Biotech Co., Ltd.,
Beijing, China). GAPDH (1:1,000; Cell Signaling Technology, Inc.,
Danvers, MA, USA) protein expression was used as a normalization
control for protein loading. All the experiments were repeated six
times over multiple days.
Transwell invasion and migration
assays
Cell invasion was assayed using a Transwell chamber
(Millipore Corporation, Billerica, MA, USA) with Matrigel (BD
Biosciences, Franklin Lakes, NJ, USA). For the invasion assay, a
Transwell chamber was placed in a six-well plate and coated with 30
μl Matrigel, which was incubated for 40 min at 37°C. In the two
Transwell assays, 24 h following transfection, the cells were
trypsinized and seeded in chambers at a density of 5×104
cells/well and cultured in DMEM with 2% serum, while 600 μl
FBS-DMEM (10%) was added to the lower chamber. After 24 h, the
migrated cells were fixed with 100% methanol for 30 min. The
non-migrated cells were removed with cotton swabs. Next, the cells
on the bottom surface of the membrane were stained with crystal
violet for 20 min. Cell images were obtained under a phase-contrast
microscope (Olympus, Tokyo, Japan), and cell counts were performed
using Image J software (National Institutes of Health, Bethesda,
MD, USA). Cell migration was assessed by determining the ability of
the cells to move into a cellular space in a two-dimensional in
vitro wound healing assay’ In brief, cells were grown to
confluence in 6-well tissue culture plastic dishes to a density of
~5 × 106 cells/well. Cells were detached by dragging a
rubber policeman (Fisher Scientific, Hampton, NH, USA) through the
center of the plate. Cultures were rinsed with PBS and replaced
with fresh DMEM or DMEM containing 10% FBS, after which the cells
were incubated at 37°C for 24 h. Images were captured at 0 and 24 h
and the migrated distance was measured using ImageJ (NIH, Bethesda,
MD, USA). Transwell invasion and migration assays were repeated six
times over multiple days.
Statistical analysis
All measurement data are presented as the mean ±
standard deviation. Statistical analysis was performed using a
t-test for two independent-samples, where P<0.05 was considered
to indicate a statistically significant difference. All analyses
were performed using SPSS version 13.0 (SPSS, Inc., Chicago, IL,
USA).
Results
Effect of the recombinant plasmid
targeting the Aurora-B gene on Aurora-B expression in HepG2
cells
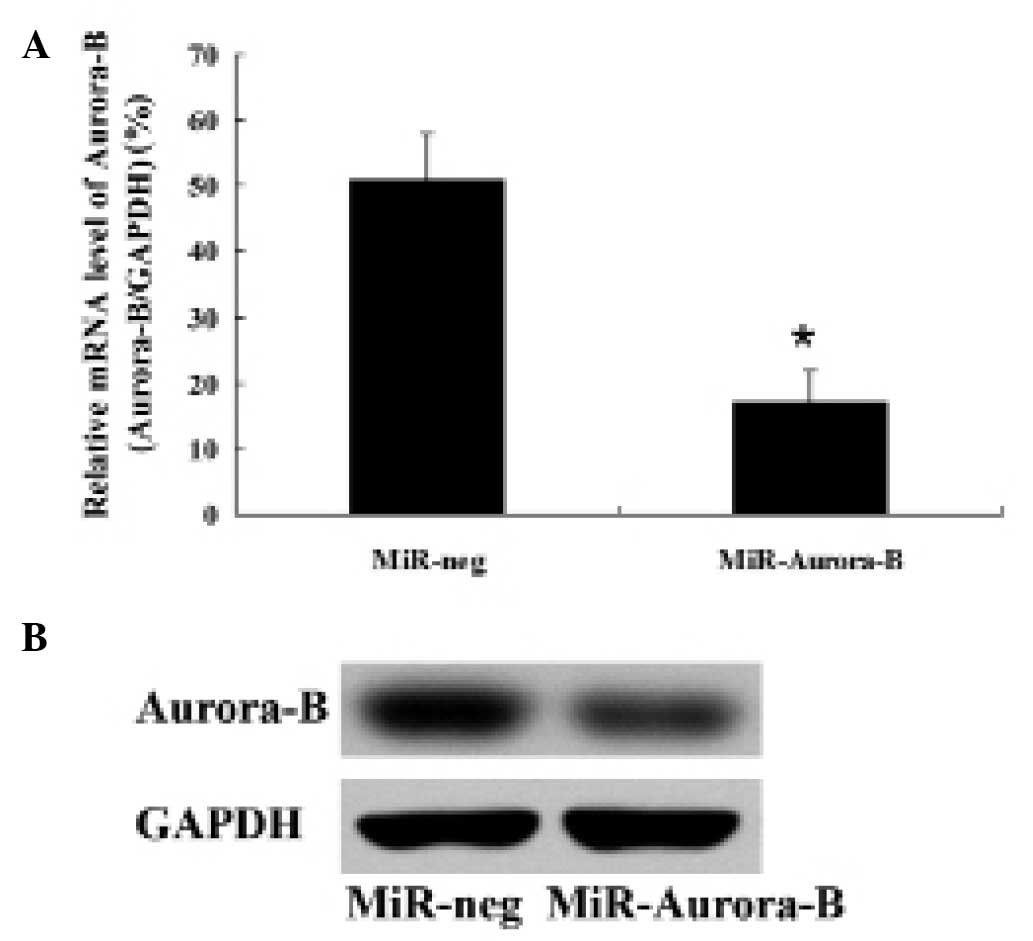
Cultured HepG2 cells were transfected with the
recombinant plasmid for 6 h and then cultured for 48 h. Aurora-B
mRNA and protein expression levels in the HepG2 cells were detected
by qPCR and western blot analysis, respectively. Aurora-B mRNA and
protein expression levels in the cells transfected with the
recombinant plasmid were significantly lower compared with those
transfected with the negative control plasmid (Fig. 1). These observations indicated that
the recombinant plasmid miRNA targeting the Aurora-B gene decreased
Aurora-B expression in HepG2 cells.
Inhibition of Aurora-B suppresses HepG2
cell invasion and migration in vitro
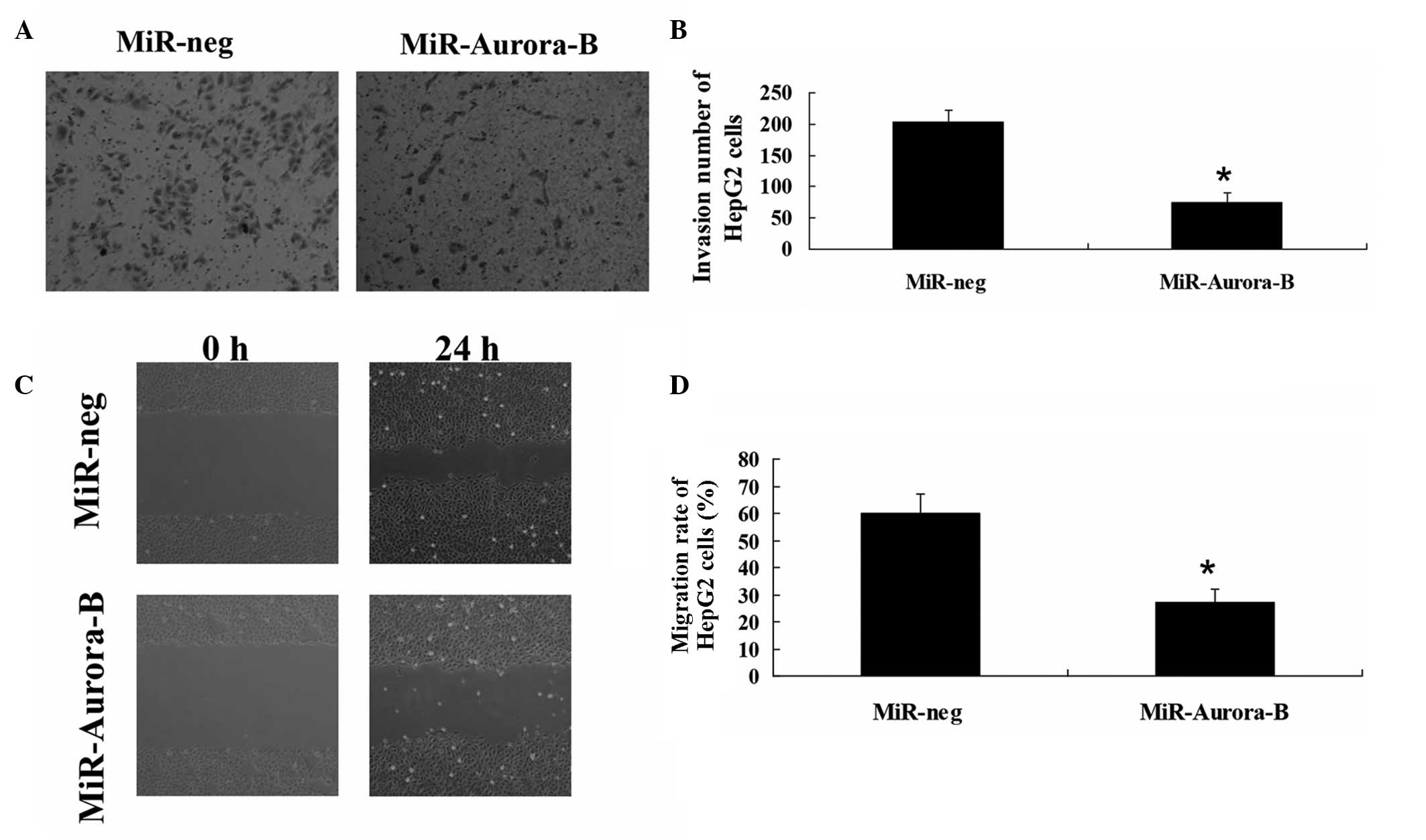
HepG2 cells were transfected with MiR-Aurora-B or
MiR-Neg. Transwell invasion and wound healing assays were performed
to investigate the migration and invasion of HepG2 cells. The
results revealed that the rates of invasion and migration in the
cells transfected with MiR-Aurora-B were significantly lower
compared with those transfected with MiR-Neg (Fig. 2). These results indicated that
Aurora-B inhibition was able to suppress HepG2 cell invasion and
migration in vitro.
Inhibition of Aurora-B decreases the
activity of the PI3K/Akt/NF-κB signaling pathway
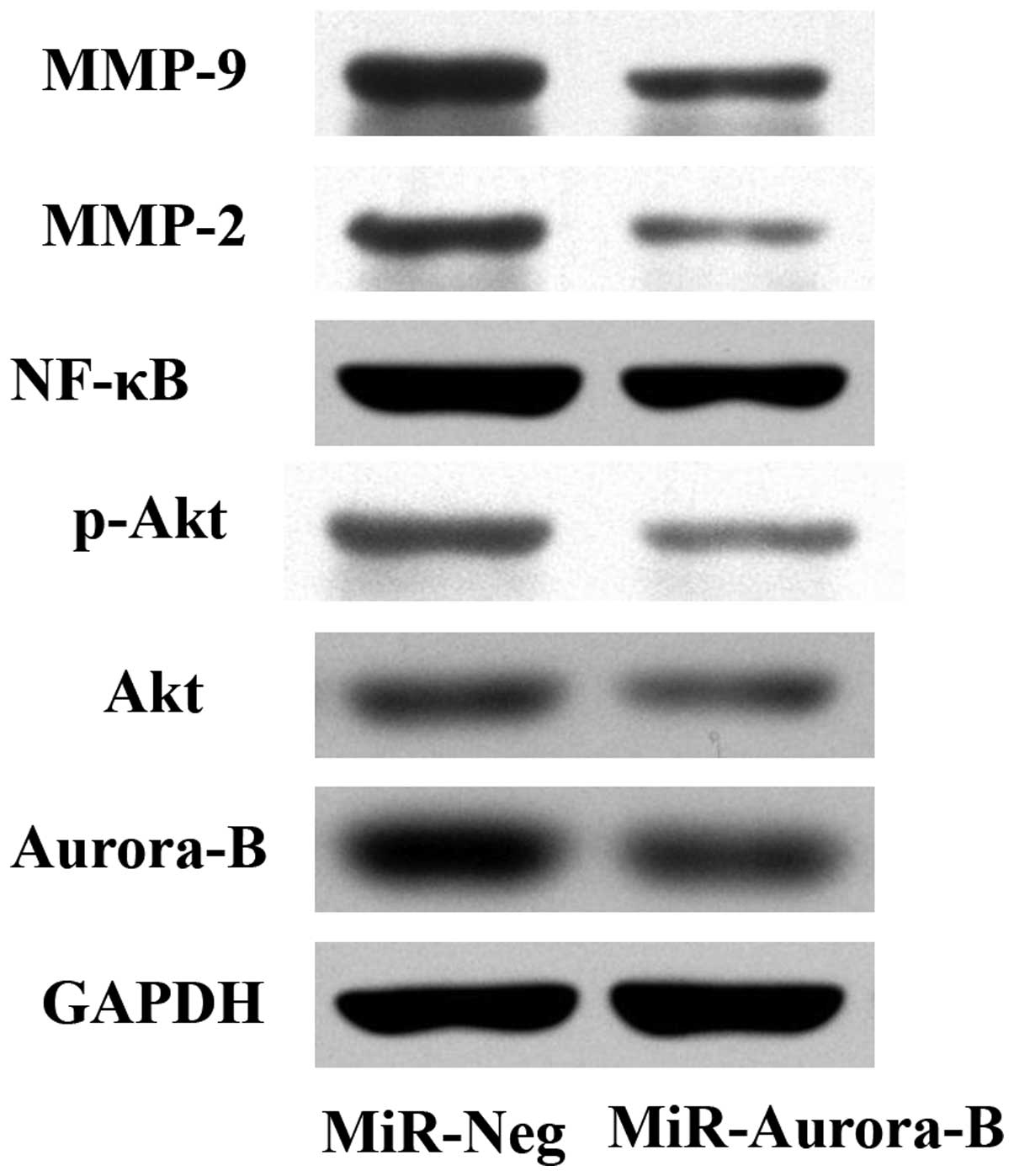
To investigate the effect of Aurora-B inhibition on
the phosphorylation of Akt, the protein expression level of p-Akt
was analyzed in HepG2 cells using western blot analysis. The
results demonstrated that the p-Akt protein expression level in the
cells transfected with MiR-Aurora-B was significantly lower
compared with the cells that had been transfected with MiR-Neg
(Fig. 3). This observation
indicated that the inhibition of Aurora-B may decrease the
phosphorylation of Akt. In addition, the protein expression levels
of Akt, NF-κB p65, MMP-2 and MMP-9 were detected, and were shown to
decrease significantly in the cells that had been transfected with
MiR-Aurora-B, as compared with the cells transfected with MiR-Neg.
These observations demonstrated that inhibition of Aurora-B gene
expression downregulated MMP-2, MMP-9 and NF-κB protein expression
levels in HepG2 cells (Fig. 3),
indicating that Aurora-B inhibition decreases the activity of the
PI3K/Akt/NF-κB signaling pathway.
Discussion
Aurora kinases are serine/threonine kinases that are
essential for cell cycle control and mitosis. Mammals have three
Aurora kinase family members (A, B and C), and these kinases are
expressed at maximum levels during mitosis. Aurora-B, part of the
CPC, is located on the chromosome arms during prophase and at the
centromeres during prometaphase and metaphase. Aurora-B
subsequently localizes to the midbody during cytokinesis. Increased
expression levels of Aurora-B have been identified in numerous
types of cancer (15–17). Various studies have demonstrated
that inhibition of Aurora-B inhibits cell proliferation and induces
cell apoptosis in a variety of tumor types (17–19).
These observations have led to an interest in Aurora-B as a
molecular target for cancer treatment. Notably, recent studies have
shown that upregulation of Aurora-B expression is associated with
tumor cell metastasis (5), while
downregulation of Aurora-B expression inhibits cell invasion and
migration in a number of tumor types (19,20).
A previous study revealed that inhibition of Aurora-B using a small
molecular inhibitor suppressed the growth of HCC cells (11). However, the effects of Aurora-B
inhibition on HCC cell invasion and migration are yet to be fully
elucidated. Therefore, the aim of the present study was to
investigate the effect of Aurora-B inhibition on HCC cell migration
and invasion. A plasmid targeting Aurora-B was used to inhibit
Aurora-B expression in HepG2 cells, and the rates of HepG2 cell
migration and invasion were investigated with Transwell assays. The
results demonstrated that the rates of migration and invasion were
significantly lower in the cells transfected with MiR-Aurora-B
compared with those treated with MiR-Neg, indicating that Aurora-B
inhibition suppresses HepG2 cell migration and invasion in
vitro.
In addition, potential molecular mechanisms
associated with Aurora-B inhibition of HCC cell migration and
invasion were analyzed. The PI3K/Akt/NF-κB signaling pathway is
known to be important in the metastasis of malignant tumors
(21). Long et al
demonstrated that inhibition of Aurora-B using a small molecular
inhibitor was able to decrease the phosphorylation of Akt at Ser
473 (14). Phosphorylation of Akt
is essential for NF-κB activation via the stimulation of the IκB
kinase complex, which phosphorylates and inactivates IκB, an
inhibitor of NF-κB. Previously, NF-κB was demonstrated to
upregulate MMP-9 expression (22),
while the inhibition of NF-κB was identified to downregulate MMP-2
expression (23). During the
development of metastases, cancer cells degrade the components of
the extracellular matrix. MMPs, particularly MMP-2 and -9, are
associated with this process due to their capacity to degrade the
extracellular matrix, promoting tumor invasion. In the present
study, the protein expression levels of Akt, p-Akt, NF-κB p65,
MMP-2 and MMP-9 were detected by western blot analysis to
investigate whether Aurora-B inhibition resulted in the decreased
activity of the PI3K/Akt/NF-κB signaling pathway. The results
demonstrated that the protein expression levels of p-Akt, Akt,
NF-κB p65, MMP-2 and MMP-9 were significantly decreased in the
cells transfected with MiR-Aurora-B when compared with those
transfected with MiR-Neg. These results indicate that inhibition of
Aurora-B decreases the activity of the PI3K/Akt/NF-κB signaling
pathway in HepG2 cells.
In conclusion, the observations of the present study
indicate that inhibition of Aurora-B suppresses HepG2 cell invasion
and migration via modulation of the PI3K/Akt/NF-κB signaling
pathway in vitro. Thus, inhibitors targeting Aurora-B and
the PI3K/Akt/NF-κB pathway may be potential therapeutic strategies
for treating HCC metastases.
References
|
1
|
Chen CJ, You SL, Lin LH, Hsu WL and Yang
YW: Cancer epidemiology and control in Taiwan: a brief review. Jpn
J Clin Oncol. 32(Suppl): S66–S81. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
El-Serag HB: Hepatocellular carcinoma: an
epidemiologic view. J Clin Gastroenterol. 35:S72–S78. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Keen N and Taylor S: Aurora-kinase
inhibitors as anticancer agents. Nat Rev Cancer. 4:927–936. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Tuncel H, Shimamoto F, Kaneko Guangying Qi
H, et al: Nuclear Aurora B and cytoplasmic Survivin expression is
involved in lymph node metastasis of colorectal cancer. Oncol Lett.
3:1109–1114. 2012.PubMed/NCBI
|
|
5
|
Takeshita M, Koga T, Takayama K, et al:
Aurora-B overexpression is correlated with aneuploidy and poor
prognosis in non-small cell lung cancer. Lung Cancer. 80:85–90.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Chen YJ, Chen CM, Twu NF, et al:
Overexpression of Aurora B is associated with poor prognosis in
epithelial ovarian cancer patients. Virchows Arch. 455:431–440.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Qi G, Ogawa I, Kudo Y, et al: Aurora-B
expression and its correlation with cell proliferation and
metastasis in oral cancer. Virchows Arch. 450:297–302. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Tovuu LO, Utsunomiya T, Imura S, et al:
The role of Aurora B expression in non-tumor liver tissues of
patients with hepatocellular carcinoma. Int J Clin Oncol. Jul
27–2013.(Epub ahead of print).
|
|
9
|
Sun DQ, Wang Y and Liu DG: Overexpression
of hnRNPC2 induces multinucleation by repression of Aurora B in
hepatocellular carcinoma cells. Oncol Lett. 5:1243–1249.
2013.PubMed/NCBI
|
|
10
|
Lin ZZ, Jeng YM, Hu FC, et al:
Significance of Aurora B overexpression in hepatocellular
carcinoma. Aurora B overexpression in HCC. BMC Cancer. 10:4612010.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Benten D, Keller G, Quaas A, et al: Aurora
kinase inhibitor PHA-739358 suppresses growth of hepatocellular
carcinoma in vitro and in a xenograft mouse model. Neoplasia.
11:934–944. 2009.PubMed/NCBI
|
|
12
|
Tao T, Cheng C, Ji Y, et al: Numbl
inhibits glioma cell migration and invasion by suppressing
TRAF5-mediated NF-κB activation. Mol Biol Cell. 23:2635–2644.
2012.PubMed/NCBI
|
|
13
|
Jiang L, Wu J, Yang Y, et al: Bmi-1
promotes the aggressiveness of glioma via activating the
NF-κB/MMP-9 signaling pathway. BMC Cancer. 12:4062012.PubMed/NCBI
|
|
14
|
Long ZJ, Xu J, Yan M, et al: ZM 447439
inhibition of aurora kinase induces Hep2 cancer cell apoptosis in
three-dimensional culture. Cell Cycle. 7:1473–1479. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Tatsuka M, Katayama H, Ota T, et al:
Multinuclearity and increased ploidy caused by overexpression of
the aurora- and Ipl1-like midbody-associated protein mitotic kinase
in human cancer cells. Cancer Res. 58:4811–4816. 1998.PubMed/NCBI
|
|
16
|
Kurai M, Shiozawa T, Shih HC, et al:
Expression of Aurora kinases A and B in normal, hyperplastic, and
malignant human endometrium: Aurora B as a predictor for poor
prognosis in endometrial carcinoma. Hum Pathol. 36:1281–1288. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Sorrentino R, Libertini S, Pallante PL, et
al: Aurora B overexpression associates with the thyroid carcinoma
undifferentiated phenotype and is required for thyroid carcinoma
cell proliferation. J Clin Endocrinol Metab. 90:928–935. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Spartà AM, Bressanin D, Chiarini F, et al:
Therapeutic targeting of Polo-like kinase-1 and Aurora kinases in
T-cell acute lymphoblastic leukemia. Cell Cycle. 13:May
29–2014.(Epub ahead of print).
|
|
19
|
Lu Y, Liu Y, Jiang J, et al: Knocking down
the expression of Aurora-A gene inhibits cell proliferation and
induces G2/M phase arrest in human small cell lung cancer cells.
Oncol Rep. 32:243–249. 2014.PubMed/NCBI
|
|
20
|
Zhu XP, Liu ZL, Peng AF, et al: Inhibition
of Aurora-B suppresses osteosarcoma cell migration and invasion.
Exp Ther Med. 7:560–564. 2014.PubMed/NCBI
|
|
21
|
Graham TR, Odero-Marah VA, Chung LW,
Agrawal KC, Davis R and Abdel-Mageed AB: PI3K/Akt-dependent
transcriptional regulation and activation of BMP-2-Smad signaling
by NF-kappaB in metastatic prostate cancer cells. Prostate.
69:168–180. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Andela VB, Gordon AH, Zotalis G, et al:
NF-κB: a pivotal transcription factor in prostate cancer metastasis
to bone. Clin Orthop Relat Res. (415 Suppl): S75–S85. 2003.
|
|
23
|
Felx M, Guyot MC, Isler M, et al:
Endothelin-1 (ET-1) promotes MMP-2 and MMP-9 induction involving
the transcription factor NF-κB in human osteosarcoma. Clin Sci
(Lond). 110:645–654. 2006.PubMed/NCBI
|