Introduction
Spinal cord injury (SCI), which is often caused by
trauma rather than disease, results in various symptoms such as
pain, paralysis or movement incontinence (1,2).
Treatments of SCI patients include restraining the spine and
controlling SCI-induced inflammation to prevent further damage
(3,4). Research into treatments for SCI
includes the use of controlled hypothermia and stem cells (5,6),
although the results of such research have seen little application
clinically.
For SCI patients, the secondary injuries include
neuronal losses driven by changes in levels of glucose, neuroactive
lipids and oxygen, and the release of free radicals, endogenous
opioids, amines and amino acids (7–13).
SCI-induced changes include activation of several molecular
signaling pathways during the first 48 h after SCI. For example,
cytoskeletal proteins have various effects on tissue survival, and
the expression levels of some genes might be altered, which may
have harmful effects on cell survival (14,15).
Thus, the identification of novel approaches that target the
signaling processes ensuing from traumatic injury to the spinal
cord is warranted.
The matrix metalloproteinases (MMPs), especially
MMP-2 and MMP-9, play key roles in tumor cell invasion and
metastasis by degrading type IV collagen, a major component of the
extracellular matrix (16–18). MMP-2 and MMP-9 are secreted as
inactive proenzymes and activated by other MMPs or other proteases.
MMP-9 is a potent regulator of acute neuroinflammation (19). It was recently reported that
reduced MMP-9 expression in the lumbar cord early after thoracic
SCI assists the recovery of learning ability in mice (20). Therefore, methods that decrease
MMP-9 expression may be useful for treating the impaired wound
healing in, for example, diabetic patients. Gene promoter
polymorphisms are often important for the roles of proteins. The
−1306C genotype ratio of the MMP-2 gene has been found to be
significantly higher in patients with lung cancers than in the
healthy population, and this genotype is associated with an
increased risk of lung cancer (21). The −1562 C to T (C/T) polymorphisms
in the MMP-9 gene promoter are considered to be important risk
factors associated with primary open-angle glaucoma (22).
It has been reported that a novel nutrient mixture
(NM), composed of lysine, ascorbic acid, proline, green tea
extracts and other micronutrients, has significant effects on MMP-2
and MMP-9 expression levels both in vitro and in vivo
(23). Therefore, in the present
study, a mouse SCI model was established to study the use of NM to
treat SCI. NM was administered to the mice and the changes in the
expression levels of MMP-2 and MMP-9 were determined.
Materials and methods
Animals and surgery
Male CD1 mice (22–28 g), aged 8–10 weeks, were used
in this experiment (Vital River Laboratory Animal Technology Co.,
Ltd, Beijing, China). The mice were kept in cages (5 mice/cage) and
maintained in one 12 h light-dark cycle. All animal experiments
were conducted according to the ethical guidelines of Xiangya
Hospital of Central South University (Changsha, China). Mice were
anesthetized with intraperitoneal ketamine and xylazine (20 and 10
mg/kg body weight, respectively). An incision on the midline of the
back was made to expose the paravertebral muscles. The spinal cord
was exposed by a T5–T8 laminectomy. The SCI was generated by
extradural compression of the T6–T7 spinal cord for 1 min with an
aneurysm clip. Following surgery, the mice were provided with food
and sterile water ad libitum.
Experimental grouping and the nutrient
mixture (NM) treatments
NM was prepared according to previously reported
methods (23). NM was composed of
the following ingredients: 700 mg vitamin C, 1,000 mg L-lysine, 750
mg L-proline, 500 mg L-arginine, 200 mg N-acetyl cysteine,
1,000 mg standardized green tea extract, 30 μg selenium, 2 mg
copper, and 1 mg manganese.
A total of 32 mice were grouped into four groups (8
mice/group) for this experiment, which comprised one sham and three
experimental groups. The mice in the sham group were subjected to
laminectomy only, without SCI being generated. The other 24 mice
were allocated into the three experimental groups treated with
different dosages of NM or vehicle (saline). The SCI model mice
received oral NM or saline in the 3 days following SCI. The sham
group received vehicle only. The SCI groups were treated orally
with saline, a low dose (500 μg 3 times/day) of NM (NM-LD) or a
high dose (2,000 μg 3 times/day) of NM (NM-HD).
Movement function evaluation
The Basso mouse scale (BMS) for locomotion was used
to evaluate the level of motor dysfunction following SCI (24). Prior to injury, the mice were
examined to ensure that they were all at normal level with a score
of 21. In 7 days after the completion of NM, the mice in every
group were scored. The scoring was initiated 3 days following SCI
and was conducted for 7 days. Scores for each hindlimb were
averaged for each day.
Western blotting
Mice were euthanized after completion of the
experiments and the spinal cords were quickly dissected, frozen and
stored at −80°C. Segments from L4–L5 were homogenized in lysis
buffer with the addition of protease inhibitor mixture (Roche
Diagnostics, Basel, Switzerland). Total proteins were separated on
10% SDS/PAGE gels, and then analyzed by immunoblotting. The primary
antibodies against MMP-2, MMP-9 and β-actin were purchased from
Santa Cruz, USA (anti-MMP-2, cat. no. sc-53630, 1:200; anti-MMP-9,
cat. no. sc-21733, 1:200; anti-β-actin, cat. no. sc-130301,
1:10,000). Secondary antibodies used in this study were goat
anti-mouse horseradish peroxidase-conjugated immunoglobulin G
(IgG-HRP; cat. no. sc-2005, 1:10,000; Santa Cruz Biotechnology,
Inc., Dallas, TX, USA). The bound antibodies were detected using an
electrochemiluminescence (ECL) system (Pierce Biotechnology Inc.,
Rockford, IL, USA). Image quantifications were performed using
ImageQuant software (GE Healthcare, Uppsala, Sweden). The
experiments were repeated at least three times.
Reverse transcription quantitative
polymerase chain reaction (RT-qPCR)
Mice were euthanized by CO2 inhalation
following completion of the experiments and the spinal cords were
quickly dissected for RNA isolation using the RNeasy kit (Qiagen,
Valencia, CA, USA) according to the manufacturer’s instructions.
One microliter of RNA was reverse transcribed into cDNA using
random primers with a Reverse Transcription II system (Promega
Corporation, Madison, WI, USA), according to the manufacturer’s
instructions. PCR was conducted using an ABI Prism Sequence
Detection System (Applied Biosystems, Foster City, CA, USA). A
VIC®-labeled probe (cat. no. 4310884E; Applied
Biosystems) was used to quantify the expression of endogenous GAPDH
mRNA, which was used as an internal control. Amplification of the
MMP-2 and MMP-9 cDNAs and the endogenous GAPDH cDNA were determined
using FAM™ and VIC fluorescence, respectively. The relative amounts
of MMP-2 and MMP-9 transcripts were expressed as ratios relative to
the levels of GAPDH mRNA. The experiments were repeated
independently at least three times. The primers used for MMP-2 were
5′-GGAGCA CGTCATGCAC and 5′-AGACACGCTAGTAGGC, and for MMP-9 were
5′-CACCACTGCAATTGCG and 5′-CACCAT CTCATACGT GAG.
Construction of firefly luciferase
constructs driven by human MMP-2 or MMP-9 promoters
A 1.6 kb segment at the 5′-flanking region of the
human MMP-2 gene or a 1.7 kb segment at the 5′-flanking region of
the human MMP-9 gene was generated by PCR using primers from the
human MMP-2 gene (Gene ID: 4313) and MMP-9 gene (GenBank accession
no. D10051). The primers used were: MMP-2 forward,
5′-AGCTAAGGCTTAGGGTACGGC; MMP-2 reverse, 5′-GCGTTAACGGACGCTAGCTAG;
MMP-9 forward, 5′-TGCACCGTGCATACCTTAG; and MMP-9 reverse,
5′-AGGGGCTGCCAGAAGCTTATGGT. The pGL2-Basic vector (Promega
Corporation) containing a polyadenylation signal upstream from the
luciferase gene was used to construct expression vectors by
subcloning PCR-amplified DNA of MMP-2 or MMP-9 promoters into the
SacI/HindIII site of the pGL2-Basic vector. Point
mutations at the loci of −1306 C/T and −1562 C/T were made using
the Site-Directed Mutagenesis kit (Agilent, Santa Clara, CA, USA).
Clones were confirmed by DNA sequencing.
Transfections and luciferase gene
assays
In brief, HeLa cells were plated onto six-well
plates at a density of 2×105 cells/well and grown
overnight. Cells were cotransfected with 1 μg construct template
(either wild-type or mutant pMMP-2-LUC and pMMP-9-LUC constructs)
and 1 μg pCMV-β-galactosidase construct using Lipofectamine reagent
(Life Technologies, Grand Island, NY, USA). After 4 h, cells were
treated with vehicle only (sham group), saline (saline group), 100
μg/ml NM (NM-LD group) or 500 μg/ml NM (NM-HD group). Luciferase
and β-galactosidase activity was determined according to the
manufacturer’s instructions and the luciferase activity of each
sample was normalized to β-galactosidase activity to calculate the
relative luciferase activities. Data are the mean ± SD from at
least five experiments. Luciferase assay and β-galactosidase assay
systems were purchased from Promega Corporation.
Statistical analysis
The experimental data are expressed as mean ±
standard deviation. Statistical software (SPSS version 10.0; SPSS,
Inc., Chicago, IL, USA) was used for independent sample t-tests,
followed by one-way variance analysis. In all analyses, P<0.05
was considered to indicate a statistically significant
difference.
Results
SCI-related activity was attenuated by
treatment with NM
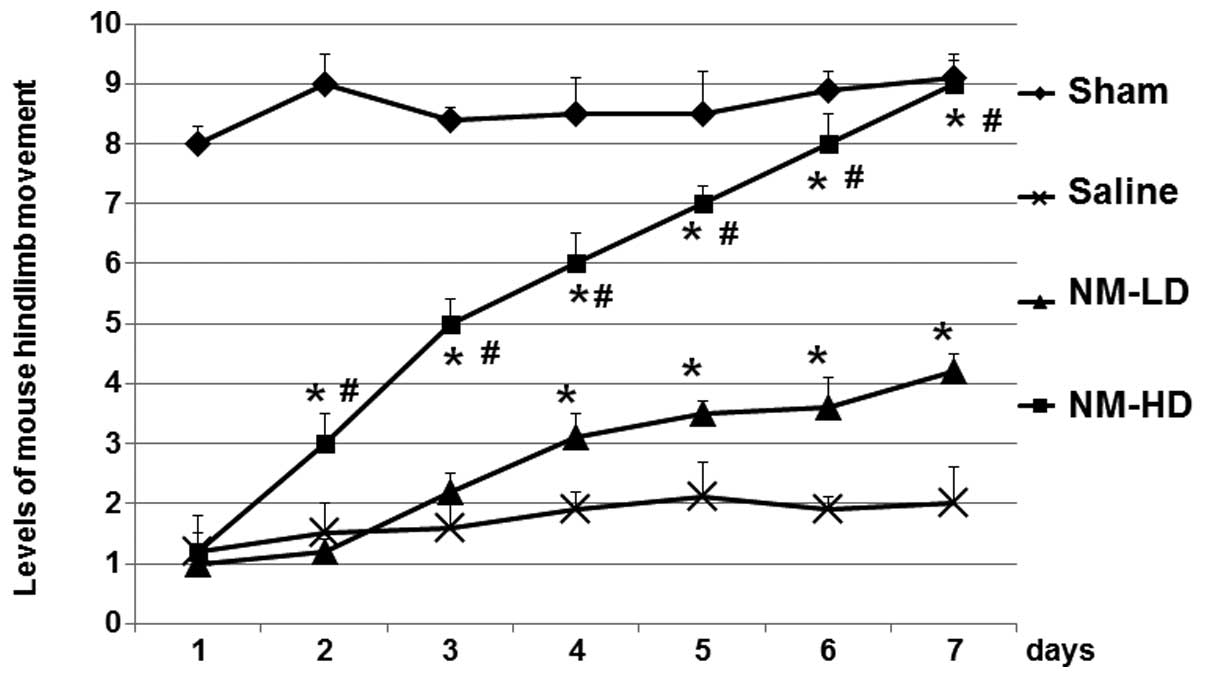
Changes in hindlimb movement of the mice, as an
indicator of the SCI-related consequences, were determined. As
shown in Fig. 1, the hindlimb
movements of mice in the sham, NM-LD and NM-HD groups were
significantly decreased at day 1 after SCI, suggesting that the
model was successfully established. The function levels of mice in
the saline group were not recovered within 7 days following the
surgery. However, the hindlimb movement levels of mice in the NM-LD
and NM-HD groups were significantly recovered when compared with
those in the saline group (Fig.
1). Furthermore, the recovery was better in the NM-HD group
than in the NM-LD group. These results suggest that NM
significantly increased the recovery of hindlimb movement of the
mice in comparison with that in the saline group.
NM decreases the expression of MMP-2 and
MMP-9 proteins
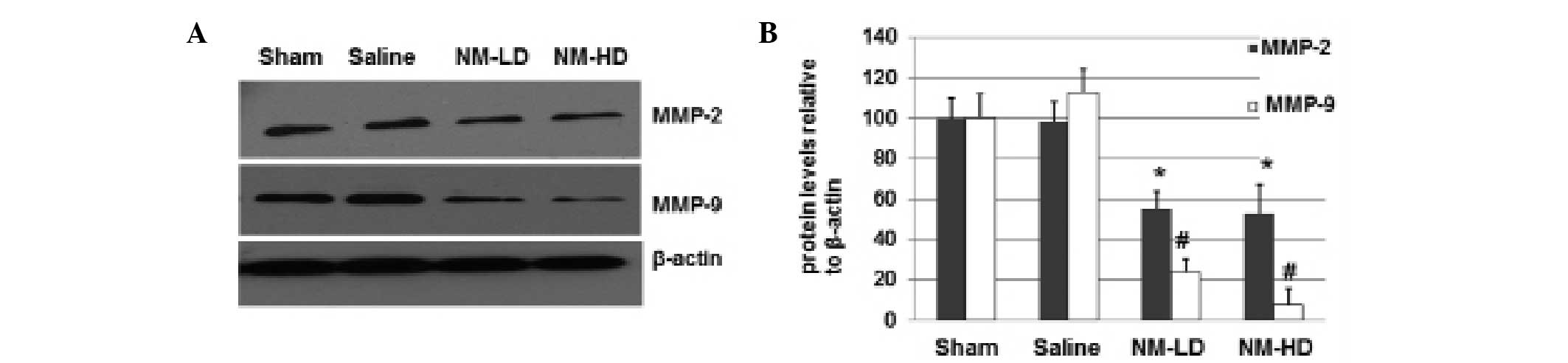
To determine whether NM is able to decrease the
expression of MMP-2 and MMP-9, mice in the sham, saline, NM-LD and
NM-HD groups were euthanized and the spinal cords were quickly
dissected for immunoblotting analyses. As shown in Fig. 2, the expression levels of MMP-2 in
the NM-LD and NM-HD groups were decreased by ~50% compared with the
saline group. The expression levels of MMP-9 in the NM-LD and NM-HD
groups were decreased to ~25 and ~10%, respectively. These results
suggest that NM significantly inhibits the expression of MMP-2 and
MMP-9, with greater inhibitory effects on MMP-9 expression than on
MMP-2 expression.
NM decreases the levels of MMP-2 and
MMP-9 mRNA
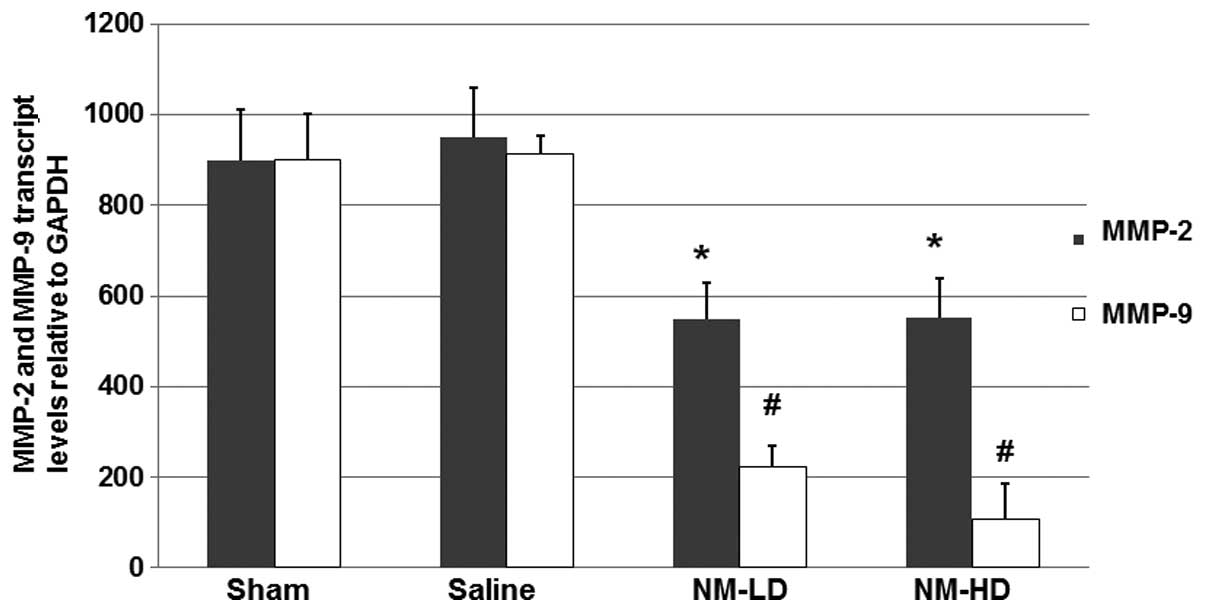
To further study the mechanism underlying the
inhibitory effects of NM on the increased MMP-9 expression, total
RNAs were harvested from the dissected spinal cords of the mice and
RT-qPCR was performed to analyze the mRNA levels of MMP-2 and
MMP-9. As shown in Fig. 3, the
RT-qPCR results indicated that NM significantly inhibited the
expression of MMP-2 and MMP-9 mRNA, respectively. These results
suggest that NM inhibits the expression of MMP-2 and MMP-9 via a
transcriptional mechanism.
NM decreases the transcriptional promoter
activities of MMP-2 and MMP-9 mRNA
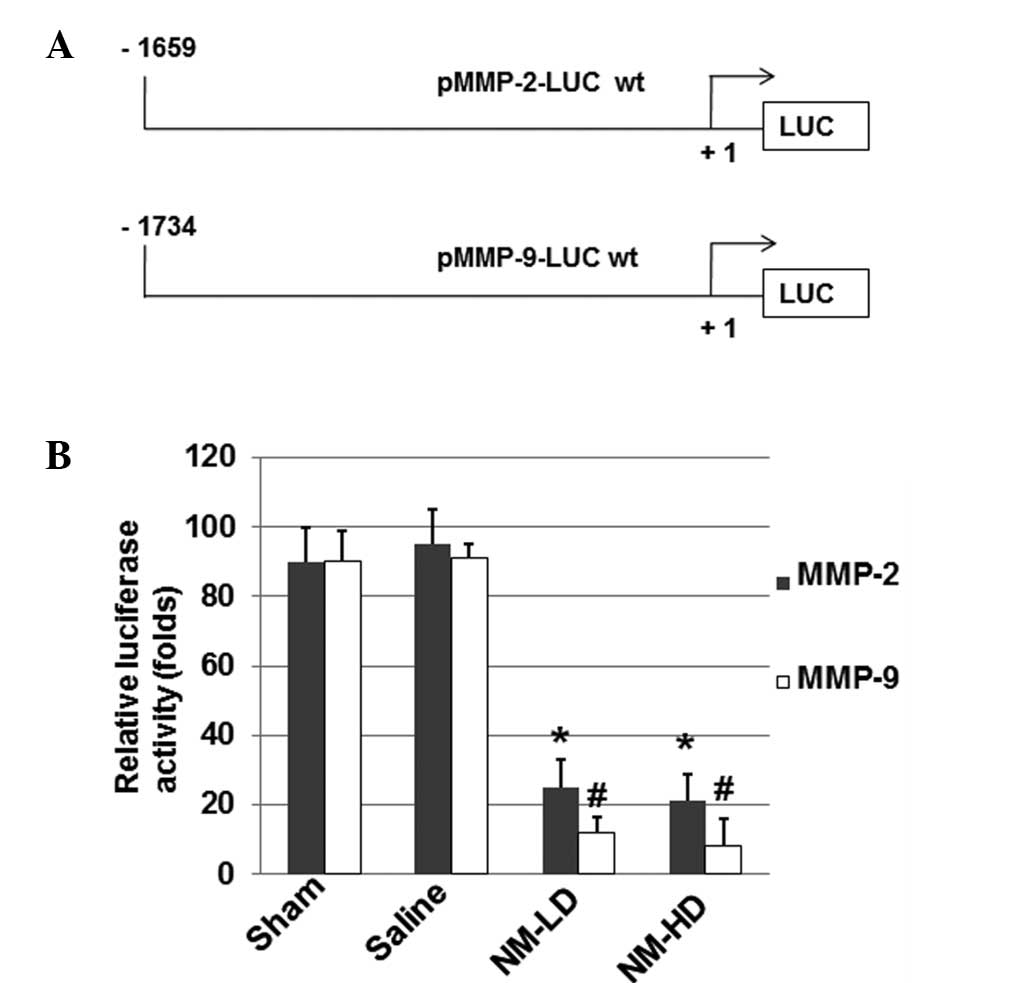
To further determine whether NM affects the
transcriptional activity of MMP-2 and MMP-9 mRNAs, luciferase
constructs driven by MMP-2 or MMP-9 promoter sequences were
prepared (Fig. 4A) and
investigated using a luciferase assay. As shown in Fig. 4B, NM significantly inhibited the
MMP-2 and MMP-9 promoter-directed luciferase activities,
respectively (P<0.05) when compared with those in the sham and
saline groups. These results suggest that NM inhibits the
expression of MMP-2 and MMP-9 via a mechanism related to the
regulation of their gene promoters.
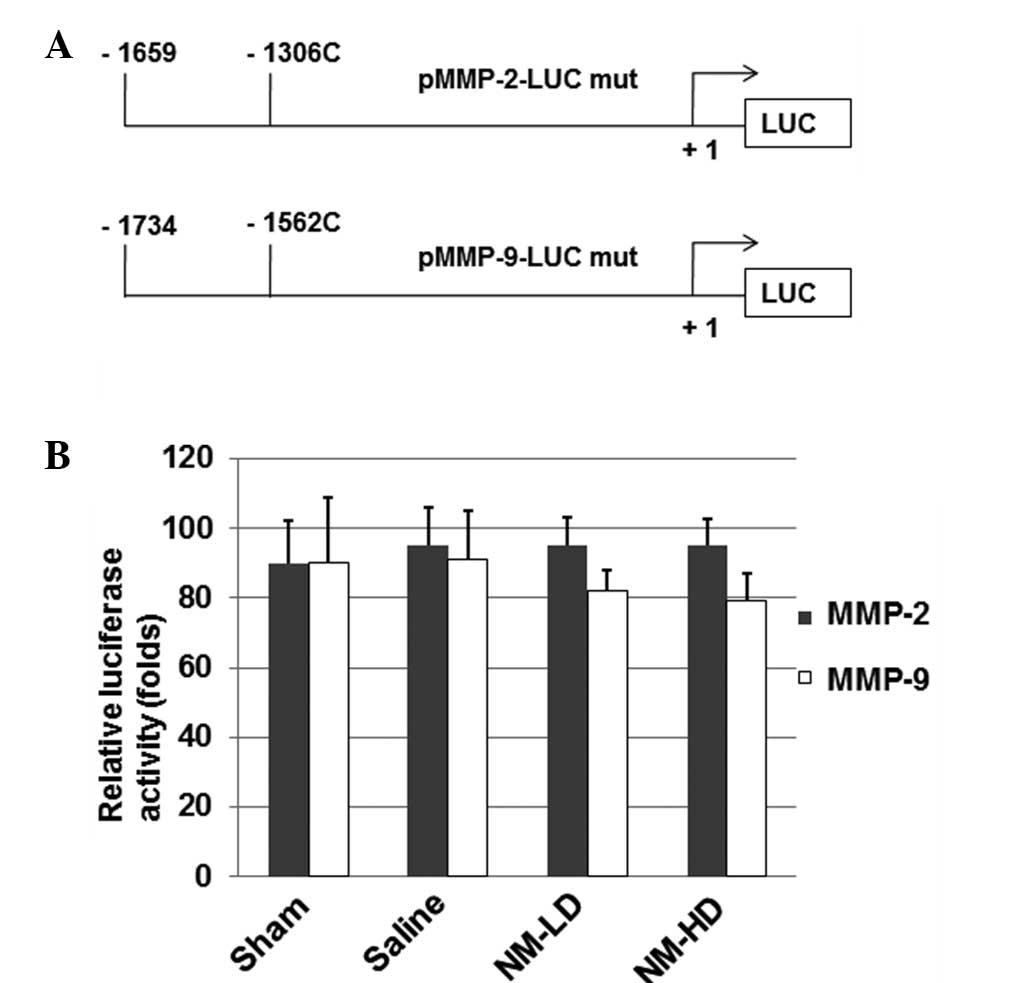
Site-directed mutagenesis abolishes the
inhibitory effects of NM on MMP-2 and MMP-9 promoters
Site-directed mutagenesis was performed to generate
the −1306 C/T base change on the MMP-2 promoter and the −1562 C/T
base change on the MMP-9 promoter in the luciferase constructs
(Fig. 5A). The luciferase assay
results (Fig. 5B) indicate that NM
did not significantly inhibit the MMP-2 and MMP-9 promoter-directed
luciferase activities when compared with those in the sham and
saline groups. These results suggest that these loci are important
for the inhibitory effect of NM on MMP-2 and MMP-9 gene
expression.
Discussion
The reduction of further damage is very important in
the treatment of patients with SCI. In the present study, a mouse
model of SCI was established to study the use of NM in the
treatment of SCI. The mice were treated with NM and the changes in
the expression levels of MMP-2 and MMP-9 were detected. It was
found that NM significantly attenuated the SCI-induced impairment
in mice movement and also decreased MMP-2 and MMP-9 expression in a
dose-dependent manner.
MMP-2 and MMP-9 are secreted as inactive proenzymes
and activated by other MMPs or other proteases. As a potent
regulator of acute neuroinflammation (19), MMP-9 has recently been found to be
able to reduce MMP-9 expression in the lumbar cord early after
thoracic SCI, suggesting that MMP-9 might be helpful for the
recovery of learning ability in mice (20). The finding in the present study
that NM decreases MMP-9 expression in a dose-dependent manner
improves the understanding of the roles of MMP-9. It was noted that
the inhibitory effect of NM on MMP-9 protein expression was more
evident than that on MMP-2. The reason underlying this difference
remains to be studied in the future.
In SCI patients, secondary injuries are often
induced. The SCI-induced secondary injuries have various symptoms,
including the neuronal losses driven by changes in the levels of
glucose, neuroactive lipids and oxygen, and the release of free
radicals, endogenous opioids, amines and amino acids (7–13).
In the present study, the recovery of hindlimb movement of the mice
treated with NM in comparison with that in the saline group was
used as an indicator of the effect of NM on SCI. The BMS for
locomotion was used to evaluate the level of motor dysfunction
following SCI. Scores for each hindlimb were averaged for each day.
It was found that in 7 days, a high dose (2,000 μg 3 times/day) of
NM significantly facilitated the recovery of mouse hindlimb
movement generated by SCI, although a low dose (500 μg 3 times/day)
of NM also had detectable effects from the fourth day after SCI.
These findings imply that NM may have an important role in the
clinic upon further studies in the future.
Changes in protein expression are often associated
with the promoter activities of genes. In the present study,
luciferase experiments were performed to investigate the effects of
NM on MMP-2 and MMP-9 promoter activities. The results suggest that
the mutations on the −1306 C locus of the MMP-2 promoter and the
−1562 C locus of the MMP-9 promoter abolished the inhibitory
effects of NM on MMP-2 and MMP-9 promoters. Since numerous cellular
protein factors, such as AP-1 and CREB (25,26),
can bind to these sites on the promoters, further studies to
identify the cis-acting elements and trans-acting factors that may
be involved in the regulation of MMP-2 and MMP-9 expression are
planned.
Acknowledgements
This study was supported by the national natural
science foundation of China (grants No. 81171698 and No.
81371956).
References
|
1
|
Kenney K and Diaz-Arrastia R: Review of
traumatic brain and spinal cord injury: challenges and
developments. JAMA Neurol. 70:13332013. View Article : Google Scholar
|
|
2
|
Neirinckx V, Cantinieaux D, Coste C, et
al: Spinal cord injuries - how could adult mesenchymal and neural
crest stem cells take up the challenge? Stem Cells. 32:829–843.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Papa S, Rossi F, Ferrari R, et al:
Selective nanovector mediated treatment of activated
proinflammatory microglia/macrophages in spinal cord injury. ACS
Nano. 7:9881–9895. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Nelissen S, Vangansewinkel T, Geurts N, et
al: Mast cells protect from post-traumatic spinal cord damage in
mice by degrading inflammation-associated cytokines via mouse mast
cell protease 4. Neurobiol Dis. 62:260–272. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ahmad FU, Wang MY and Levi AD: Hypothermia
for acute spinal cord injury - a review. World Neurosurg.
82:207–214. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Yamamoto A, Matsubara K, Kano F and Sakai
K: Analysis of the neuroregenerative activities of mesenchymal stem
cells in functional recovery after rat spinal cord injury. Methods
Mol Biol. 1213:321–328. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bareyre FM and Schwab ME: Inflammation,
degeneration and regeneration in the injured spinal cord: insights
from DNA microarrays. Trends Neurosci. 26:555–563. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
DeWitt DS, Prough DS, Taylor CL and
Whitley JM: Reduced cerebral blood flow, oxygen delivery, and
electroencephalographic activity after traumatic brain injury and
mild hemorrhage in cats. J Neurosurg. 76:812–821. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kruman II and Mattson MP: Pivotal role of
mitochondrial calcium uptake in neural cell apoptosis and necrosis.
J Neurochem. 72:529–540. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Pedersen MO, Jensen R, Pedersen DS, et al:
Metallothionein-I+II in neuroprotection. Biofactors. 35:315–325.
2009. View
Article : Google Scholar : PubMed/NCBI
|
|
11
|
Takahashi H, Manaka S and Sano K: Changes
in extracellular potassium concentration in cortex and brain stem
during the acute phase of experimental closed head injury. J
Neurosurg. 55:708–717. 1981. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yamakami I and McIntosh TK: Effects of
traumatic brain injury on regional cerebral blood flow in rats as
measured with radiolabeled microspheres. J Cereb Blood Flow Metab.
9:117–124. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zemper ED: Analysis of cerebral concussion
frequency with the most commonly used models of football helmets. J
Athl Train. 29:44–50. 1994.PubMed/NCBI
|
|
14
|
Lau BY, Fogerson SM, Walsh RB and Morgan
JR: Cyclic AMP promotes axon regeneration, lesion repair and
neuronal survival in lampreys after spinal cord injury. Exp Neurol.
250:31–42. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Nakano N, Nakai Y, Seo TB, et al: Effects
of bone marrow stromal cell transplantation through CSF on the
subacute and chronic spinal cord injury in rats. PLoS One.
8:e734942013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Liotta LA, Tryggvason K, Garbisa A, et al:
Metastatic potential correlates with enzymatic degradation of
basement membrane collagen. Nature. 284:67–68. 1980. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Stetler-Stevenson WG: The role of matrix
metalloproteinases in tumor invasion, metastasis and angiogenesis.
Surg Oncol Clin N Am. 10:383–392. 2001.PubMed/NCBI
|
|
18
|
Stetler-Stevenson WG: Type IV collagenases
in tumor invasion and metastasis. Cancer Metastasis Rev. 9:289–303.
1990. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Vlodavsky E, Palzur E and Soustiel JF:
Hyperbaric oxygen therapy reduces neuroinflammation and expression
of matrix metalloproteinase-9 in the rat model of traumatic brain
injury. Neuropathol Appl Neurobiol. 32:40–50. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hansen CN, Fisher LC, Deibert RJ, et al:
Elevated MMP-9 in the lumbar cord early after thoracic spinal cord
injury impedes motor relearning in mice. J Neurosci.
33:13101–13111. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Miao X, Yu C, Tan W, et al: A functional
polymorphism in the matrix metalloproteinase-2 gene promoter
(−1306C/T) is associated with risk of development but not
metastasis of gastric cardia adenocarcinoma. Cancer Res.
63:3987–3990. 2003.
|
|
22
|
Markiewicz L, Majsterek I, Przybylowska K,
et al: Gene polymorphisms of the MMP1, MMP9, MMP12, IL-1β and TIMP1
and the risk of primary open-angle glaucoma. Acta Ophthalmol.
91:e516–e523. 2013.PubMed/NCBI
|
|
23
|
Roomi MW, Kalinovsky T, Niedzwiecki A and
Rath M: Modulation of u-PA, MMPs and their inhibitors by a novel
nutrient mixture in pediatric human sarcoma cell lines. Int J
Oncol. 43:1027–1035. 2013.
|
|
24
|
Basso DM, Fisher LC, Anderson AJ, et al:
Basso Mouse Scale for locomotion detects differences in recovery
after spinal cord injury in five common mouse strains. J
Neurotrauma. 23:635–659. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kankaanranta H, Ilmarinen P, Zhang X, et
al: Tumour necrosis factor-α regulates human eosinophil apoptosis
via ligation of TNF-receptor 1 and balance between NF-κB and AP-1.
PLoS One. 9:e902982014.
|
|
26
|
Tang M, Shi S, Guo Y, et al: GSK-3/CREB
pathway involved in the gx-50’s effect on Alzheimer’s disease.
Neuropharmacology. 81:256–266. 2014.PubMed/NCBI
|