Introduction
Mesenchymal stem cells (MSCs) are considered to be
one of the most promising therapeutic cell sources for regenerative
medicine, primarily due to their multipotency and immunosuppressive
functions. These cells can be isolated from multiple types of
tissue, including bone marrow (BM), skin, adipose and umbilical
cord tissue (1–3), and are able to differentiate into
osteoblasts, adipocytes, chondrocytes and myocardial cells in
vitro (4,5). Previous studies have confirmed that
MSCs are also able to secrete bioactive factors that alter the
milieu of dysfunctional tissues (6,7). These
observations provide strong evidence of the potential therapeutic
role of MSCs in the treatment of various types of diseases.
However, the therapeutic use of MSCs has been
limited due to a number of factors, including difficulties in
obtaining sufficient numbers of cells and the unsuccessful
engraftment of the cells following transplantation. Current methods
include the in vitro expansion of MSCs in plastic adherent
culture (8), and the subsequent use
of these cells for transplantation into patients. However, in
vitro expansion in a standard adherent culture can markedly
alter the cell phenotype, which may lead to lung entrapment of the
cells and little or no engraftment of the cells in the target
organs (8–10). At present, seldom studies have been
conducted with the aim to investigate the factors that cause a
variation in the MSC phenotype in vitro, and one of the
important reasons for this is the lack of suitable research
methods.
In 2011, Vunjak-Novakovic and Scadden classified the
cellular and acellular components of the stem cell niche (11). The authors demonstrated the important
role of the environment in determining the characteristics of MSCs.
In addition, the in vitro conditions of the adherent culture
may play a vital role in determining the stem cell niche; thus, may
affect MSCs. A previous study revealed that CD44− BM
cells contained almost all clonogenic cells with a multilineage
differentiation potential (12).
However, in vitro culture of CD44− BM cells
resulted in their conversion to a CD44+ phenotype. With
regard to these observations, it was hypothesized that plastic
adherence in culture may affect the cell phenotype of MSCs that
undergo amplification in vitro.
Stem cell antigen-1 (Sca-1) is enriched on freshly
isolated BM MSCs (13–15), and Sca-1+ MSCs are known
to have an important function in improving cardiac function in
myocardial infarction (16). To
determine whether adherence to plastic during culture affects the
expression of Sca-1 in MSCs, a novel method of seeding MSCs in
ultra-low-attachment culture plates was applied to analyze the
effects of plastic adherence on Sca-1 expression. The Sca-1 cell
surface marker on MSCs was analyzed using flow cytometry. In
addition, the mRNA expression of Sca-1 was assessed by quantitative
polymerase chain reaction (qPCR) to confirm the differences in
Sca-1 expression between the cells grown in adherent and
nonadherent culture conditions.
Materials and methods
Isolation of mononuclear cells from BM
in mice
FVB/N mice (age, 9–15 weeks) were obtained from the
Karolinksa Institute (Stockholm, Sweden) and 6 mice were sacrificed
for this study. Mice were sacrificed by cervical dislocation.
Animal experimental protocols were performed with approval from the
Local Ethics Committee at Karolinska Institute. Mononuclear cells
were isolated from the BM of the mice using a previously described
method (12). Briefly, femurs,
tibias and iliac crests were crushed in Dulbecco's
phosphate-buffered saline (DPBS; Gibco Life Technologies, Paisley,
Scotland) and 10% fetal bovine serum (FBS; #10500064; Gibco Life
Technologies, Divinopolis, Brazil). Cells from the bone samples
were obtained following treatment of the bone fragments with 0.1%
collagenase II (Worthington Biochemical Corporation, Lakewood, NJ,
USA) and 0.05% trypsin-EDTA (#25300062; Gibco Life Technologies,
Grand Island, NY, USA) for 30–45 min at 37°C. Bone and BM cells
were pooled and centrifuged at 300 × g (Sorvall ST 16R; Thermo
Fisher Scientific, Osterode, Germany) for 5–10 min at room
temperature, after which the cells were resuspended in DPBS plus
10% FBS for BM MSC isolation. This research was performed at the
Center for Hematology and Regenerative Medicine (HERM) of
Karolinska Institute.
Multicolor fluorescence-activated cell
sorting (FACS) isolation of mouse MSCs
In a previous study, CD44− MSCs were
determined to have properties of MSCs (12). Therefore, BM
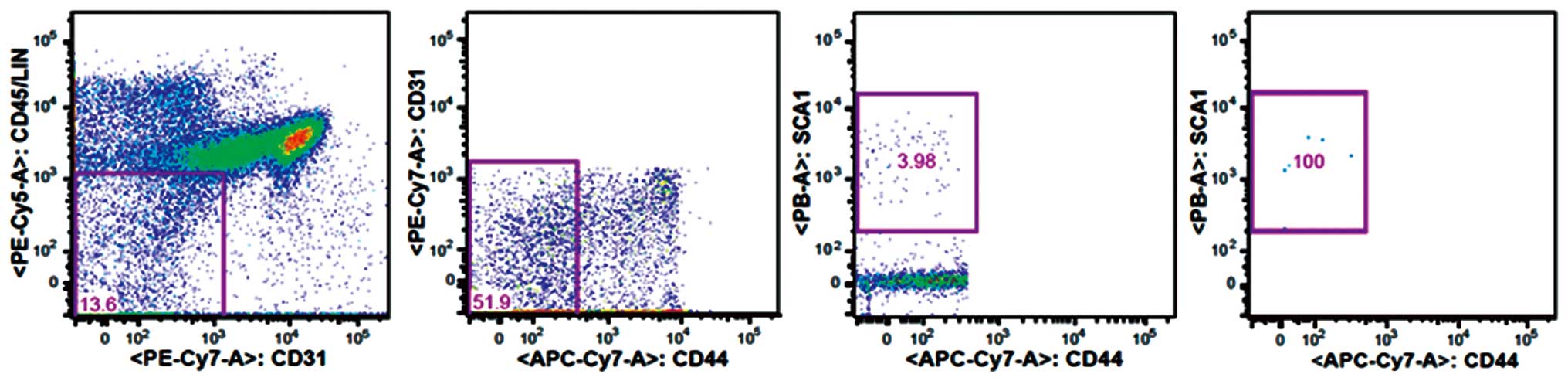
CD45−CD31−CD44−Sca-1+
MSCs were selected for the study. Briefly, the hematopoietic cells
in the BM mononuclear cell preparations were initially depleted by
incubating the cells with a purified rat anti-mouse CD45 primary
antibodies against CD45 (#140451; eBioscience Inc., San Diego, CA,
USA), TER119 (#116202), GR1 (#108402; 1:50), B220 (#103202; 1:100),
CD4 (#100506), CD8 (#100802) and MAC1 (#101202; 1:200; BioLegend
Inc., San Diego, CA, USA). Cells were subsequently incubated with
sheep anti-rat Dynabeads (Dynal Biotech, Inc., New York, NY, USA).
The remaining hematopoietic cells were visualized using a goat
anti-rat tricolor antibody and fluorescence-conjugated anti-CD45
(#553082; BD Biosciences, Franklin Lakes, NJ, USA), anti-TER119
(#116210) and anti-CD19 (#115510; BioLegend Inc.) antibodies to
remove any hematopoietic cells. Dead cells were excluded by
propidium iodide staining. CD44−Sca-1+
stromal cells were gated based on the Fluorescence Minus One
controls for CD44 Sca-1 expression using a FACSAria III flow
cytometer (BD Biosciences), as shown in Fig. 1. These experimental procedures were
performed at the Center for HERM of Karolinska Institute.
Expansion of MSCs and nonadherence in
vitro culture conditions
Sorted MSCs were plated in 20 ml complete Dulbecco's
modified Eagle's medium (#10569010; Gibco Life Technologies, Grand
Island, NY, USA), containing 10% FBS, 10 mM
N-2-hydroxyethylpiperazine-N-2-ethane sulfonic acid (#15630106;
Gibco Life Technologies, Paisley, Scotland), 100 U/ml penicillin
and 100 mg/ml streptomycin (15140122; Gibco Life Technologies,
Grand Island, NY, USA), in a T75 tissue culture flask (#430641;
Corning, Inc., Corning, NY, USA). A hypoxic environment is known to
greatly improve the genetic stability and expression of chemokine
receptors during in vitro expansion (17). Thus, to increase the accuracy of the
results, the cells were incubated in a incubator (FORMA3131; Thermo
Fisher Scientific) with 1% O2 and 5% CO2.
Complete medium was changed every 5–7 days. When the culture cells
exhibited 90% fusion, the cells were suspended by incubation in
0.05% trypsin-EDTA for 5 min at 37°C, and the MSCs were reseeded at
a density of 2,500 cells/cm2 in a T75 tissue culture
flask.
For this plastic-expansion approach, the cells were
passaged eight times. Cells in passage 9–11 were transferred into
the nonadherent cultures and grown for 24 or 72 h. The cells were
plated at a density of 2,500 cells/cm2 in
ultra-low-attachment tissue culture plates (#3471; Corning, Inc.).
At the same time, an equal number of cells were transferred into
adherent culture plates (#353046; BD Biosciences) and grown for 24
or 72 h as a control. In addition, nonadherent culture cells and
adherent culture cells from the 72-h cultures were reseeded (2,500
cells/cm2) in the adherent culture plates and grown for
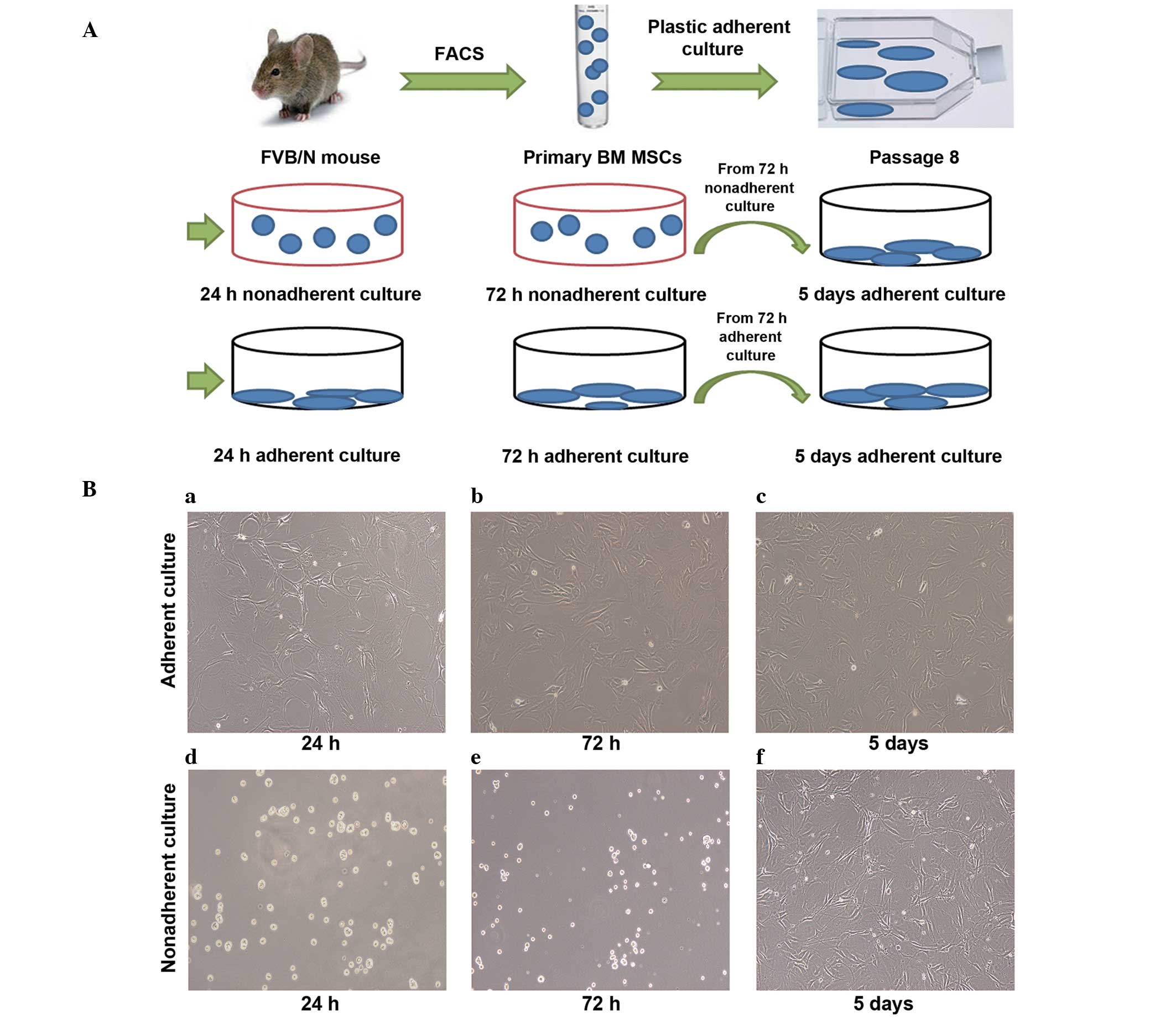
5 days under adherent culture conditions (Fig. 2A).
FACS analysis of the Sca-1 MSC surface
marker
The control adherent cultured cells at 24 h, 72 h
and 5 days were detached with 0.05% trypsin-EDTA and collected in a
50-ml centrifuge tube. The nonadherent cultured cells were also
collected in a 50-ml centrifuge tube simultaneously. The cells were
centrifuged at 300 × g for 5 min and resuspended in
phosphate-buffered saline (PBS; Gibco Life Technologies, Taicang,
China) for analysis on a BD FACSCalibur (BD Biosciences). Adherent
and nonadherent cultured cells were stained with a rat anti-mouse
Sca-1 antibody (#557405; BD Biosciences) to analyze the expression
of this cell surface marker. An isotype control (PE-R3–34
immunoglobulin; #554685; BD Biosciences) was added at the indicated
concentration (0.25 µg), and BD FACSComp software (BD Biosciences)
was used for data analysis.
qPCR
Adherent and nonadherent cultured cells grown for 24
h, 72 h and 5 days were collected in 1.5-ml Eppendorf tubes. The
total RNA was extracted from the adherent and nonadherent culture
cells using a High Pure RNA isolation kit (#11828665001; Roche
Diagnostics, Laval, QC, Canada), according to the manufacturer's
instructions. The cDNA was synthesized using SuperScript III and
Oligo primers (#E6300S; New England Biolabs, Inc., Ipswich, MA,
USA), according to the manufacturer's instructions. The primers
used for Sca-1 were as follows: Forward,
5′-AGGAGGCAGCAGTTATTGTGG-3′, and reverse,
5′-CGTTGACCTTAGTACCCAGGA-3′. β-actin (ACTB) was used as the
reference gene, with the following primers: Forward,
5′-GGCTGTATTCCCCTCCATCG-3′, and reverse,
5′-CCAGTTGGTAACAATGCCATGT-3′ (designed by Gibco Life Technologies,
Shanghai, China). qPCR analysis of the Sca-1 gene was performed
using the Light-Cycler 480 platform (Roche Diagnostics, Basel,
Switzerland) with a SYBR Green I PCR kit (#4887352001; Roche
Diagnostics, Laval, QC, Canada). The mixture contained 10 ml SYBR
Green I Master, 6 ml RNase-free H2O, 1 ml PCR forward
primer (10 mM), 1 ml PCR reverse primer (10 mM) and 2 ml cDNA in a
final reaction volume of 20 ml. Sca-1 mRNA expression levels were
normalized against the Ct of the ACTB RNA (ΔΔCt). The Ct
method was applied to determine the fold changes using the
Light-Cycler 480 software. The experiment was repeated three times,
and each experiment was performed in triplicate.
Statistical analysis
GraphPad Prism software, version 5 (GraphPad
Software, Inc., La Jolla, CA, USA) was used for statistical
analysis. The Student's t-test was applied to identify the
statistical significance of the differences between the culture
condition groups. P<0.05 was considered to indicate a
statistically significant difference.
Results
Morphology of the adherent and
nonadherent cultured cells at different time points
MSCs seeded in tissue culture plates (2,500
cells/well) at 24 h, 72 h and 5 days were attached to the bottom of
the plastic tissue culture plates and were shown to exhibit flat
morphology (Fig. 2Ba–c). However,
MSCs seeded in the ultra-low-attachment culture plates at 24 and 72
h were suspended and scattered throughout the medium, exhibiting a
rounded morphology (Fig. 2Bd–e).
When nonadherent culture cells obtained after 72 h were reseeded in
the tissue culture plates and incubated for 5 days in the adherent
culture conditions, the cells exhibited a similar morphology to the
adherent culture cells (Fig.
2Bf).
FACS analysis of the Sca-1 MSC surface
marker
Sca-1 expression on the MSCs between passages 9 and
11 was analyzed by FACS. At 24 h, the expression of Sca-1 on the
nonadherent cultured cells was similar to that of the control group
of adherent cultured cells (P>0.05). However, Sca-1 expression
at 72 h differed significantly between the adherent and nonadherent
cultured cells. At 72 h, the expression level of Sca-1 in the
nonadherent cells was approximately one half of that observed in
the adherent cultured cells (P<0.05). However, when the
nonadherent cultured cells grown for 72 h were reseeded in the
adherent conditions and cultured for 5 days, Sca-1 expression
recovered to the level exhibited by the adherent cultured cells
(P>0.05; Fig. 3).
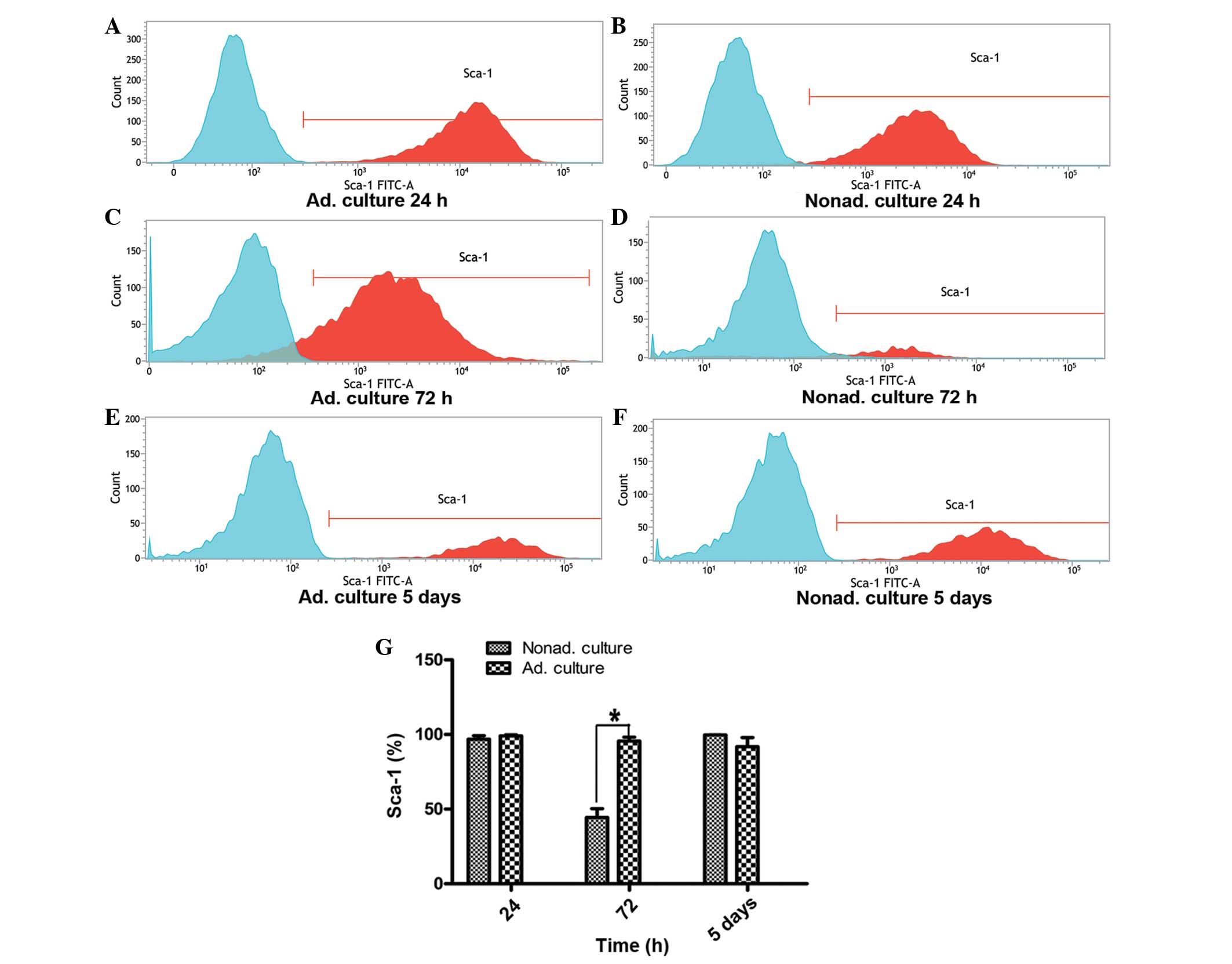 | Figure 3.Flow cytometry analysis of the
expression of the Sca-1 surface marker. (A, C and E) Graphs
represent Sca-1 expression on the adherent cultured cells at 24 h,
72 h and 5 days, respectively. (B, D and F) Graphs represent Sca-1
expression on the nonadherent cultured cells at 24 h, 72 h and 5
days, respectively. Expression of the Sca-1 surface marker was
analyzed by fluorescence-activated cell sorting, and the expression
levels were compared between the adherent and nonadherent cultured
cells. Three independent experiments were performed with the cells
between passages 9 and 11, which all showed similar Sca-1
expression. (G) Statistical histogram shows the differences in
Sca-1 expression levels at 24 h (P>0.05), 72 h (*P<0.05) and
5 days (P>0.05) between the nonadherent and adherent cultured
cells. FITC, fluorescein isothiocyanate; Ad, adherent; Nonad;
nonadherent; Sca, stem cell antigen. |
qPCR
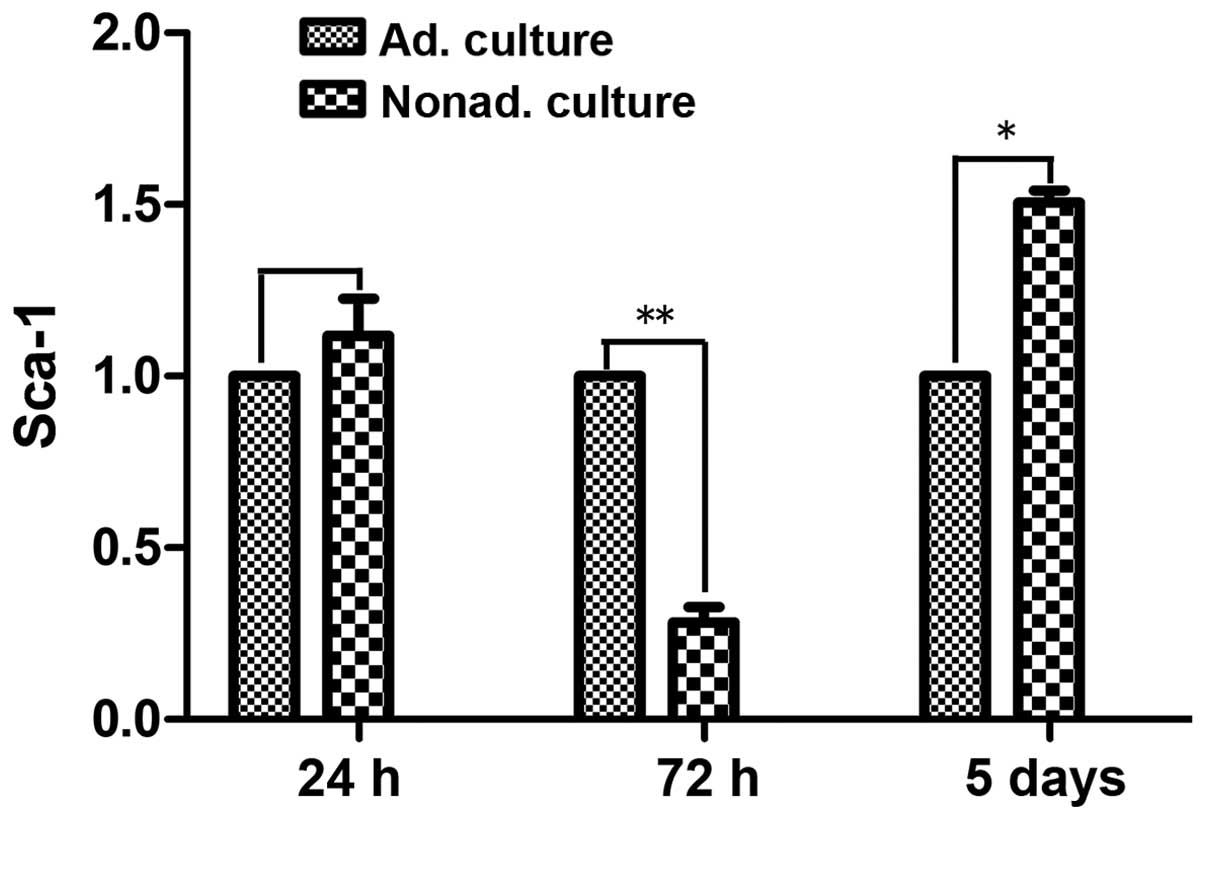
qPCR was used to compare the mRNA expression levels
of Sca-1 following culture for 24 h, 72 h and 5 days between the
adherent and nonadherent cultured cells. At 24 h, Sca-1 mRNA
expression levels did not differ significantly between the
nonadherent and adherent cultured cells (P>0.05). However, at 72
h, the mRNA expression levels of Sca-1 were ~3 times lower in the
nonadherent cultured cells, as compared with the control adherent
cultured cells (P<0.01). After culture for 5 days in the
adherent conditions, Sca-1 mRNA expression levels were upregulated,
and were 1.5 times higher in the nonadherent cultured cells
compared with the control adherent cultured cells (P<0.05;
Fig. 4).
Discussion
Sca-1 is widely recognized as a marker that can be
used to enrich stem cells in a number of tissues (18–20). In
addition, Sca-1 expression has been shown to be enriched on
isolated mouse BM MSCs with a regenerative and self-renewal
capacity (15,19,21,22). In
the present study, a novel method of seeding MSCs in
ultra-low-attachment culture plates was used, and Sca-1 expression
in MSCs was shown to change following culture in nonadherent
conditions.
As previous research has shown, CD44−
MSCs exhibit similar properties to MSCs (12). Thus, to analyze Sca-1 expression, the
CD45−CD31−CD44−Sca-1+
MSCs were sorted. In addition, a previous study reported that MSCs
enriched by low-density culture undergo senescence and lose their
stem cell properties (23). However,
low-density hypoxic culture is a method for efficiently expanding
MSCs without losing their stem cell properties or increasing
tumorigenicity (17,24). Therefore, to ensure the presence of
MSCs with stable stem-cell properties, low-density hypoxic
conditions (1% O2) were utilized in the present
study.
The expression of Sca-1 on MSCs was analyzed using
FACS analysis, and the expression levels were compared between the
nonadherent and adherent cultured cells at three time points. At 24
h, there was no statistically significant difference in Sca-1
expression between the nonadherent and adherent cultured cells.
However, a notable change in the expression of the Sca-1 cell
surface marker was observed at 72 h and 5 days. At 72 h, the
nonadherent cultured cells had significantly lower expression
levels of Sca-1 compared with the adherent cultured cells. This
observation directly confirmed that the nonadherent culture
conditions downregulated Sca-1 expression on the MSCs. Notably, the
expression of the Sca-1 cell surface marker was shown to recover to
the level of the adherent cultured cells when the nonadherent
cultured cells, obtained after 72 h, were reseeded in the tissue
culture plates and incubated for 5 days in the adherent conditions.
These results indicate that the adherent culture conditions
increased the expression of Sca-1 following the downregulation by
the nonadherent culture. Therefore, these data suggest that
nonadherent culture treatment can downregulate the expression of
Sca-1 on MSCs. Accordingly, the results from the qPCR analysis
confirmed these observations. The pattern of Sca-1 mRNA expression
was similar to that of the cell surface expression of the protein
on the MSCs. Therefore, the results indicate that the cell surface
protein expression and the mRNA expression of Sca-1 can be changed
by a variation in culture conditions between nonadherent and
adherent culture. Consequently, the cell surface protein and mRNA
expression levels of Sca-1 in the MSCs were affected by the
nonadherent culture method. In addition, plastic adherence in
culture may affect the cell phenotype and gene expression in MSCs
that undergo amplification in vitro.
A previous study indicated that the upregulated
expression of Sca-1 is associated with a more destructive tumor
phenotype (25). Furthermore, in
mouse models, Sca-1 has been shown to be associated with greater
tumorigenic potential (26). Despite
the lack of clear evidence for the malignant transformation of MSCs
during in vitro culture, the susceptibility of these cells
to functional transformation should not be ignored. Consequently,
clinicians and scientists are concerned with regard to the
biological safety of MSC transplantation. Adherence to plastic
conditions is a critical factor for MSC proliferation in
vitro; however, the molecular processes that drive
proliferation are complex, and the underlying mechanisms remain
unclear. Therefore, the nonadherent culture technique used in the
present study may provide a novel method to study the mechanisms
underlying plastic adherent culture.
In conclusion, Sca-1 expression in the MSCs were
affected by the nonadherent culture method. Further study is
required to clarify the association between phenotype variation and
biological characteristics in MSCs that undergo amplification in
vitro.
Acknowledgements
The authors thank Hong Qian from the Center for HERM
at Karolinska Institute for the valuable advice and contribution to
the cell sorting (FACS) isolation undertaken in the present study.
This study was supported by grants from the International Program
of Project 985, Sun Yat-sen University and the ‘One Hundred
Talented Scholars’ of Sun Yat-sen University (no. F002009011).
References
|
1
|
Mundra V, Gerling IC and Mahato RI:
Mesenchymal stem cell-based therapy. Mol Pharm. 10:77–89. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kang EJ, Byun JH, Choi YJ, et al: In vitro
and in vivo osteogenesis of porcine skin-derived mesenchymal stem
cell-like cells with a demineralized bone and fibrin glue scaffold.
Tissue Eng Part A. 16:815–827. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Tong CK, Vellasamy S, Tan BC, et al:
Generation of mesenchymal stem cell from human umbilical cord
tissue using a combination enzymatic and mechanical disassociation
method. Cell Biol Int. 35:221–226. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Caplan AI: Adult mesenchymal stem cells
for tissue engineering versus regenerative medicine. J Cell
Physiol. 213:341–347. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kolf CM, Cho E and Tuan RS: Mesenchymal
stromal cells. Biology of adult mesenchymal stem cells: regulation
of niche, self-renewal and differentiation. Arthritis Res Ther.
9:2042007. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Schinköthe T, Bloch W and Schmidt A: In
vitro secreting profile of human mesenchymal stem cells. Stem Cells
Dev. 17:199–206. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Morrison SJ and Scadden DT: The bone
marrow niche for haematopoietic stem cells. Nature. 505:327–334.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Rabani V, Shahsavani M, Gharavi M, Piryaei
A, Azhdari Z and Baharvand H: Mesenchymal stem cell infusion
therapy in a carbon tetrachloride-induced liver fibrosis model
affects matrix metalloproteinase expression. Cell Biol Int.
34:601–605. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Qian H, Badaloni A, Chiara F, et al:
Molecular characterization of prospectively isolated multipotent
mesenchymal progenitors provides new insight into the cellular
identity of mesenchymal stem cells in mouse bone marrow. Mol Cell
Biol. 33:661–677. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Karp JM and Leng Teo GS: Mesenchymal stem
cell homing: the devil is in the details. Cell Stem Cell.
4:206–216. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Vunjak-Novakovic G and Scadden DT:
Biomimetic platforms for human stem cell research. Cell Stem Cell.
8:252–261. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Qian H, Le Blanc K and Sigvardsson M:
Primary mesenchymal stem and progenitor cells from bone marrow lack
expression of CD44 protein. J Biol Chem. 287:25795–25807. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Morikawa S, Mabuchi Y, Kubota Y, et al:
Prospective identification, isolation, and systemic transplantation
of multipotent mesenchymal stem cells in murine bone marrow. J Exp
Med. 206:2483–2496. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Nakamura Y, Arai F, Iwasaki H, et al:
Isolation and characterization of endosteal niche cell populations
that regulate hematopoietic stem cells. Blood. 116:1422–1432. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Forte G, Franzese O, Pagliari S, et al:
Interfacing Sca-1(pos) mesenchymal stem cells with biocompatible
scaffolds with different chemical composition and geometry. J
Biomed Biotechnol. 2009:9106102009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hughey CC, Ma L, James FD, et al:
Mesenchymal stem cell transplantation for the infarcted heart:
therapeutic potential for insulin resistance beyond the heart.
Cardiovasc Diabetol. 12:1282013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Haque N, Rahman MT, Abu Kasim NH and
Alabsi AM: Hypoxic culture conditions as a solution for mesenchymal
stem cell based regenerative therapy. Scientific World Journal.
2013:6329722013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Forte G, Carotenuto F, Pagliari F, et al:
Criticality of the biological and physical stimuli array inducing
resident cardiac stem cell determination. Stem Cells. 26:2093–2103.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Nadri S, Soleimani M, Hosseni RH, Massumi
M, Atashi A and Izadpanah R: An efficient method for isolation of
murine bone marrow mesenchymal stem cells. Int J Dev Biol.
51:723–729. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Gong X, Sun Z, Cui D, et al: Isolation and
characterization of lung resident mesenchymal stem cells capable of
differentiating into alveolar epithelial type II cells. Cell Biol
Int. 38:405–411. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Holmes C and Stanford WL: Concise review:
stem cell antigen-1: expression, function, and enigma. Stem Cells.
25:1339–1347. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lee CH, Park JH, Lee JH, et al:
Replacement of mouse embryonic fibroblasts with bone marrow stromal
cells for use in establishing and maintaining embryonic stem cells
in mice. Cell Biol Int. 36:537–543. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Shibata KR, Aoyama T, Shima Y, et al:
Expression of the p16INK4A gene is associated closely with
senescence of human mesenchymal stem cells and is potentially
silenced by DNA methylation during in vitro expansion. Stem Cells.
25:2371–2382. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Tsai CC, Chen YJ, Yew TL, et al: Hypoxia
inhibits senescence and maintains mesenchymal stem cell properties
through down-regulation of E2A-p21 by HIF-TWIST. Blood.
117:459–469. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Witz IP: Differential expression of genes
by tumor cells of a low or a high malignancy phenotype: the case of
murine and human Ly-6 proteins. J Cell Biochem Suppl. 34:61–66.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yuan H, Upadhyay G, Yin Y, Kopelovich L
and Glazer RI: Stem cell antigen-1 deficiency enhances the
chemopreventive effect of peroxisome proliferator-activated
receptorγ activation. Cancer Prev Res (Phila). 5:51–60. 2012.
View Article : Google Scholar : PubMed/NCBI
|